

Abstract
Objective
Obsessive-compulsive disorder (OCD) is a common and debilitating psychiatric illness. Although a genetic component contributes to its etiology, no single gene or mechanism has been identified to the OCD susceptibility. The catechol-O-methyltransferase (COMT) and monoamine oxidase A (MAO-A) genes have been investigated in previous OCD studies, but the results are still unclear. More recently, Taylor (2013) in a comprehensive meta-analysis of genetic association studies has identified COMT and MAO-A polymorphisms involved with OCD. In an effort to clarify the role of these two genes in OCD vulnerability, a family-based association investigation was performed as an alternative strategy to the classical case-control design.
Methods
Transmission disequilibrium analyses were performed after genotyping 13 single-nucleotide polymorphisms (eight in COMT and five in MAO-A) in 783 OCD trios (probands and their parents). Four different OCD phenotypes (from narrow to broad OCD definitions) and a SNP x SNP epistasis were also analyzed.
Results
OCD, broad and narrow phenotypes,were not associated with any of the investigated COMT and MAO-A polymorphisms. In addition, the analyses of gene-gene interaction did not show significant epistatic influences on phenotype between COMT and MAO-A.
Conclusions
The findings do not support an association between DSM-IV OCD and the variants of COMT or MAO-A. However, results from this study cannot exclude the contribution of these genes in the manifestation of OCD. The evaluation of broader spectrum phenotypes could help to understand the role of these and other genes in the pathophysiology of OCD and its spectrum disorders.
Introduction
Obsessive-compulsive disorder (OCD) is characterized by repetitive thoughts (obsessions) and repetitive behaviors (compulsions) that are unwanted, time consuming, egodystonic, and result in significant functional impairment [1]. Various studies have demonstrated that the etiology of OCD has a genetic component [2] with reported heritability rates ranging between 27% and 65% [3]. Although more than 140 candidate gene studies have been conducted, the findings have been inconclusive due to small sample size and few study replications [4]. The etiology of OCD seems to involve an interaction between environmental factors and several genes of small effect [2]. Studies employing different methodological strategies (e.g., cerebral spinal fluid metabolites measurements, pharmacological challenges, and drug treatment responses) all suggest that abnormal serotonergic neurotransmission is one of the most consistent biological findings in OCD [5]. Studies have also reported dopaminergic abnormalities in the basal ganglia and nucleus accumbens, as well as altered glutamate transmission in other locations in OCD samples, thus suggesting that complex interactions between multiple neurotransmitter systems may contribute to the phenotypic presentation of OCD [6]. This could be understood by a potential dysfunction in enzymes that metabolize central nervous system neurotransmitters in OCD. Two important enzymes of this type are catechol-O-methyltransferase (COMT) and monoamine oxidase A (MAO-A).
COMT is an Mg2+-dependent enzyme involved in the inactivation of certain catecholamines (norepinephrine, epinephrine, and dopamine). The important role that COMT plays in neuropsychiatric disorders has previously been described [7]. The most widely studied COMT polymorphism is a single-nucleotide polymorphism (SNP) leading to a valine-to-methionine substitution at codon 158 (val158met or rs4680), which results in a low-activity thermolabile and a high-activity thermostable forms of the enzyme—the met(158) and val(158) alleles, respectively [8].
A deletion within the 22q11 region, which includes the COMT region, causes velocardiofacial syndrome, which increases the risk for many psychiatric disorders [9], including OCD [10]. Although various studies have evaluated the association between COMT polymorphisms and OCD, the findings have been inconclusive [4].
The monoamine oxidases (MAO) are enzymes that catalyze the oxidation of monoamines, which exist in two forms: MAO-A and MAO-B. Both forms are found bound to the outer membrane of mitochondria in most cell types in the body but show different specificities. MAO-A is present in catecholaminergic neurons in the brain and plays a preferencial role in the metabolic degradation of several neurotransmitters, including serotonin, norepinephrine, epinephrine, and dopamine. The MAO-A gene maps to the p11.3 region on the X chromosome. The first evidence that MAO-A could play an important role in human behavior was the description of a large Dutch family that presented with a new form of mental retardation with prominent behavioral abnormalities linked to the X chromosome. Several males in the family were affected by this disturbance and exhibited aggressive behavior, with significant impulsivity, pyromania, suicide attempts, and sexually aberrant behavior [11,12].
Some studies that have investigated the association between MAO-A polymorphisms and OCD, and they have produced controversial results [4]. The various methodologies, different phenotype definitions, small sample sizes and diverse ethnic backgrounds of the populations studied could be sources of bias contributing to the discrepant results obtained in previous studies.
Recently, Taylor [13] performed a comprehensive meta-analysis of OCD genetic association studies. A total of 230 polymorphisms from 113 genetic association studies were identified. In the main meta-analysis, COMT and MAO-A polymorphisms were found to be associated with OCD in males.
In this study, the transmission disequilibrium of COMT and MAO-A SNPs was analyzed in OCD patients, using a family-based association approach. Given that family studies have shown familial aggregation between OCD and different psychiatric disorders (e.g., tic disorders, body dysmorphic disorder, skin picking, trichotillomania and anxiety disorders) [14,15,16,17,18,19], these disorders were included as broader phenotypes in the analyses.
Methods
Participants
The sample consisted of 783 individuals meeting DSM-IV criteria for OCD, and their biological parents. The OCD probands were recruited from specialized OCD clinics at Harvard University, the University of California- San Francisco, the University of Michigan, the University of Toronto, as well as from two universities in Brazil—the University of São Paulo and the University of Pernambuco that participate from the Obsessive-Compulsive Foundation Genetics Collaborative Subgroup. Most of the sample was studied in a genome-wide association study [20]. The probands and their parents underwent structured clinical interviews. For subjects who were ≥ 16 years of age, we employed the Structured Clinical Interview for DSM-IV Axis I Disorders, Clinician Version [21]. For subjects who were < 16 years of age, we employed the Schedule for Affective Disorders and Schizophrenia for School Aged Children—Present and Lifetime Version (K-SADS-PL) [22]. In addition, all subjects were interviewed with the Yale-Brown Obsessive Compulsive Scale [23] and the Yale Global Tic Severity Scale [24]. All of the interviewers were trained mental health professionals (psychiatrists or psychologists). Reliability evaluations were performed and best-estimate diagnoses were made. The assessment procedures have previously been described in detail [25].
Ethics Statement
This work was performed in accordance with Declaration of Helsinki. The project was approved by the research ethics committees (IRB) at each of the participating institutions (In Pernambuco, Brazil: Complexo Hospitalar do Hospital Universitario Oswaldo Cruz e Pronto Socorro Cardiologico de Pernambuco, University of Pernambuco – HUOC/PROCAP #29/2010; and In Sao Paulo, Brazil: Comissao de Etica para Analise de Projetos de Pesquisa do Instituto de Psiquiatria da Faculdade de Medicina da Universidade de Sao Paulo—CAPPesq IPqHCFMUSP #968/05; United States: University of California, San Francisco Committee on Human Research—#10–00157; Institutional Review of the University of Michigan Medical School (IRBMED) protocol #1992-0191; Canada: Toronto: Centre for Addiction and Mental Health Research Ethics Board #368-2008). Written informed consent was obtained from all adult participants and from the parents of those who were minors.
Narrow and Broad Phenotypes categories
In addition to the association analysis between COMT and MAO-A and DSM-IV OCD, additional analyses were performed in the Brazilian subsample (83 trios) using the broader phenotypes described below.
Hypothesizing that OCD phenomenology (and not severity) could be influenced by COMT and MAO-A genes, we examined the association of COMT and MAO-A polymorphisms and a broadly-defined OCD, which includes cases with OCD symptoms even when they did not meet full DSM-IV criteria for OCD (subclinical OCD).Acknowledging that OCD is part of a spectrum [26,27] that includes other anxiety disorders [19,28], body dysmorphic disorder, and pathological grooming disorders (skin picking and trichotillomania) [17], these conditions were collectively referred to as OCD spectrum disorders. The term ‘OCD spectrum disorders’ includes DSM-IV OCD, subclinical OCD, together with the DSM-IV anxiety disorders, body dysmorphic disorder, and pathological grooming disorders (skin picking and trichotillomania).
In addition, since some studies have suggested that OCD could be grouped based on tic disorder comorbidity [29,30], analyses in the Brazilian subsample were also completed to examine an association between COMT and MAO-A polymorphisms and OCD with tic disorder (Tourette syndrome or chronic tic disorder) comorbidity.
Genotyping
DNA from peripheral blood samples were extracted using the “salting-out” protocol [31]. On the basis of Tagger software [32], COMT and MAOA genomic regions were specified and tag SNPs were picked (a local copy of HapMap data was used) within them. Tagger produced a list of tag SNPs and corresponding statistical tests to capture all variants of interest, and a summary coverage report of the selected tag SNPs. Eleven SNPs in the COMT gene region and six SNPs in the MAO-A gene region were selected. These SNPs were genotyped at the Psychiatric and Neurodevelopmental Genetics Unit of Massachusetts General Hospital, a teaching hospital associated with Harvard University. Genotyping was performed in 384-well plates with the Sequenom MassARRAY platform (Sequenom, San Diego, CA, USA). Primers for polymerase chain reaction (PCR) amplification and single-base extension assays were designed using Assay Design software, version 3.1 on the basis of FASTA sequences surrounding the SNPs, derived from SNPper [33]. Multiplex PCR was performed, followed by a pooled single-base extension reaction (iPLEX Gold). Samples were analyzed with a MassARRAY RT mass spectrometer, in automated mode. The resulting spectra were analyzed with SpectroTyper software after baseline correction and peak identification (Sequenom). Information about the ten COMT SNPs and the six MAO-A SNPs was obtained from the dbSNP, Celera, and HapMap databases. Three markers were excluded because of quality control issues: COMT rs2239393, because the genotyping success rate was lower than 75%; MAO-A rs2179098, because the minor allele frequency was ≤ 0.025; and COMT rs1544325, because the Hardy-Weinberg p value was < 0.05. The thirteen remaining SNPs collectively covered the COMT and MAO-A gene regions: the eight selected COMT SNPs spanned 25.6 kb, with a density of 2.84 kb/SNP; and the five selected MAO-A SNPs spanned 63 kb, with a density of 12.6 kb/SNP. Single-marker and haplotype analyses were performed in order to identify associations.
Statistical analysis
Statistical analyses were performed with the programs PLINK [34,35] and Haploview 3.32 [36]. Using the Haploview software, SNPs were selected on the basis of quality control criteria. 95% confidence bounds on D prime are generated and each comparison is called "strong LD", "inconclusive" or "strong recombination". A haplotype block is created if 95% of informative (i.e., non-inconclusive) comparisons are "strong LD" [37]. Following this, the haplotype TDT analysis was performed.
For single markers association, the results of the standard transmission/disequilibrium test (TDT) and the `parental discordance test` [34] were combined in order to calculate the p-value using PLINK [34,35]. No covariate was added in the TDT analysis model. The `parental discordance test`is based on a comparison between affected and unaffected parents in terms of the number of alleles they carry, treating each parental pair as a matched case-control pair. The `parental discordance test` assumes homogeneity, in terms of population stratification, within rather than between nuclear families. This test can add power to family-based association analyses, as well as provide considerable protection against population stratification [38].
In addition to the association analysis between COMT and MAO-A and DSM-IV OCD, a secondary investigation of the COMT and MAO-A associations in our sample was completed. First, probands were grouped into gender-matched trios in order to evaluate the influence of gender on the results. This approach was based on the knowledge that COMT has influence on estrogens [39] and that MAO-A is located on the X chromosome.
Second, exploratory analyses were performed on the Brazilian subsample to consider as affected status, some OCD-related phenotypes such as broadly-defined OCD (clinical plus subclinical OCD), OCD spectrum disorders and OCD with comorbid tic disorders.
Third, since both COMT and MAOA are involved in dopamine catabolism, the gene- gene interaction was evaluated in contributing to OCD etiology. It was used as the case-only epistatic analysis under the SNP x SNP model, using PLINK software [34,35] in the whole sample (783 trios). Only SNPs that are more than 1 Mb apart, or on different chromosomes, are included in case-only analyses.
The statistical power of this study was calculated using Genetic Power Calculator [40].
The results were subject to permutation analysis (100,000 permutations) using Plink, in order to control for false-positive association, and to Bonferroni`s correction for multiple analysis (corrected p-value threshold = 0.0004).
Results
Strictly-defined DSM-IV OCD phenotype
The results of the OCD association analysis of the eight COMT SNPs and five MAO-A SNPs are described in Table 1. The TDT, the “parental discordance test”, and the combined test were all not significant for the female (Table 2) and the male proband trios (Table 3).
Table 1. Single-marker analysis of OCD association with catechol-O-methyltransferase and monoamine oxidase A single-nucleotide polymorphisms.
| Gene | SNP | OR | CHISQ | P | CHISQ_PAR | P_PAR | CHISQ_COM | P_COM |
|---|---|---|---|---|---|---|---|---|
| COMT | rs737866 | 0.756 | 6.127 | 0.013 | 2.286 | 0.131 | 7.86 | 0.005 |
| rs933271 | 1.238 | 3.191 | 0.074 | 0.222 | 0.637 | 3.413 | 0.065 | |
| rs5993883 | 1.164 | 2.042 | 0.153 | 1.882 | 0.17 | 3.133 | 0.077 | |
| rs740603 | 0.97 | 0.091 | 0.762 | 2.574 | 0.109 | 0.057 | 0.812 | |
| rs4680 | 1.034 | 0.099 | 0.752 | 1 | 0.317 | 0.002 | 0.96 | |
| rs4646316 | 0.851 | 1.766 | 0.184 | 0.143 | 0.706 | 1.325 | 0.25 | |
| rs165774 | 0.988 | 0.012 | 0.911 | 0.037 | 0.847 | 0.026 | 0.872 | |
| rs9332377 | 0.88 | 0.832 | 0.362 | 0.6 | 0.439 | 1.174 | 0.278 | |
| MAO-A | rs1465107 | 0.957 | 0.066 | 0.798 | 0.4 | 0.527 | 0.277 | 0.599 |
| rs1465108 | 1.016 | 0.008 | 0.929 | 0.026 | 0.873 | 0 | 1 | |
| rs6323 | 1.014 | 0.007 | 0.934 | 1.4 | 0.237 | 0.2 | 0.655 | |
| rs979606 | 1.094 | 0.269 | 0.604 | 0.471 | 0.493 | 0.024 | 0.877 | |
| rs979605 | 1.103 | 0.343 | 0.558 | 0.641 | 0.423 | 0.022 | 0.882 |
SNP: single-nucleotide polymorphism; TDT: transmission/disequilibrium test; OR: TDT odds ratio; CHISQ: TDT chi-square value; P: TDT p value; CHISQ_PAR: parental discordance test chi-square value; P_PAR: parental discordance test p value; CHISQ_COM: combined test chi-square value; P_COM: combined test p value; COMT: catechol-O-methyltransferase; MAO-A: monoamine oxidase-A
Table 2. Analysis of OCD association with catechol-O-methyltransferase and monoamine oxidase A single-nucleotide polymorphisms in female proband trios.
| Gene | SNP | OR | CHISQ | P | CHISQ_PAR | P_PAR | CHISQ_COM | P_COM |
|---|---|---|---|---|---|---|---|---|
| COMT | rs737866 | 0.649 | 5.541 | 0.019 | 0.4 | 0.527 | 5.939 | 0.015 |
| rs933271 | 1.137 | 0.449 | 0.503 | 0.667 | 0.414 | 0.217 | 0.641 | |
| rs5993883 | 1.463 | 4.699 | 0.03 | 2.579 | 0.108 | 6.737 | 0.009 | |
| rs740603 | 1.295 | 2.314 | 0.128 | 4.167 | 0.041 | 4.78 | 0.029 | |
| rs4680 | 1.293 | 2.173 | 0.14 | 0.4 | 0.527 | 1.573 | 0.21 | |
| rs4646316 | 0.893 | 0.34 | 0.56 | 0 | 1 | 0.3 | 0.585 | |
| rs165774 | 0.875 | 0.533 | 0.465 | 1 | 0.317 | 0.194 | 0.66 | |
| rs9332377 | 0.783 | 1.22 | 0.269 | 0.111 | 0.739 | 0.89 | 0.345 | |
| MAO-A | rs1465107 | 1.083 | 0.08 | 0.777 | 0.067 | 0.796 | 0.015 | 0.901 |
| rs1465108 | 1.087 | 0.083 | 0.773 | 0.067 | 0.796 | 0.016 | 0.9 | |
| rs6323 | 1.24 | 0.643 | 0.423 | 0.692 | 0.405 | 0.13 | 0.718 | |
| rs979606 | 1.2 | 0.455 | 0.5 | 0.692 | 0.405 | 0.059 | 0.808 | |
| rs979605 | 1.269 | 0.83 | 0.362 | 0.692 | 0.405 | 0.222 | 0.637 |
SNP: single-nucleotide polymorphism; TDT: transmission/disequilibrium test; OR: TDT odds ratio; CHISQ: TDT chi-square value; P: TDT p value; CHISQ_PAR: parental discordance test chi-square value; P_PAR: parental discordance test p value; CHISQ_COM: combined test chi-square value; P_COM: combined test p value; COMT: catechol-O-methyltransferase; MAO-A: monoamine oxidase-A
Table 3. Analysis of OCD association with catechol-O-methyltransferase and monoamine oxidase A single-nucleotide polymorphisms in male proband trios.
| Gene | SNP | OR | CHISQ | P | CHISQ_PAR | P_PAR | CHISQ_COM | P_COM |
|---|---|---|---|---|---|---|---|---|
| COMT | rs737866 | 0.827 | 1.705 | 0.192 | 0.889 | 0.346 | 2.327 | 0.127 |
| rs933271 | 1.28 | 2.579 | 0.108 | 0.111 | 0.739 | 2.689 | 0.101 | |
| rs5993883 | 1.037 | 0.073 | 0.786 | 0.067 | 0.796 | 0.107 | 0.743 | |
| rs740603 | 0.851 | 1.613 | 0.204 | 0.043 | 0.835 | 1.332 | 0.248 | |
| rs4680 | 0.955 | 0.114 | 0.736 | 1 | 0.317 | 0.345 | 0.557 | |
| rs4646316 | 1.099 | 0.424 | 0.515 | 0.889 | 0.346 | 0.12 | 0.73 | |
| rs165774 | 0.918 | 0.214 | 0.644 | 2.667 | 0.102 | 0.658 | 0.417 | |
| rs9332377 | 0.954 | 0.048 | 0.827 | 0 | 1 | 0.037 | 0.847 | |
| MAO-A | rs1465107 | 1.054 | 0.053 | 0.818 | 0.037 | 0.847 | 0.087 | 0.768 |
| rs1465108 | 0.954 | 0.046 | 0.829 | 0.222 | 0.637 | 0.154 | 0.695 | |
| rs6323 | 1.111 | 0.21 | 0.646 | 0.048 | 0.827 | 0.094 | 0.761 | |
| rs979606 | 1.077 | 0.111 | 0.739 | 0 | 1 | 0.087 | 0.768 | |
| rs979605 | 0.827 | 1.705 | 0.192 | 0.889 | 0.346 | 2.33 | 0.127 |
SNP: single-nucleotide polymorphism; TDT: transmission/disequilibrium test; OR: TDT odds ratio; CHISQ: TDT chi-square value; P: TDT p value; CHISQ_PAR: parental discordance test chi-square value; P_PAR: parental discordance test p value; CHISQ_COM: combined test chi-square value; P_COM: combined test p value; COMT: catechol-O-methyltransferase; MAO-A: monoamine oxidase-A
Haplotypes
Two haplotype blocks were identified in each gene (Figs. 1 and 2). In the COMT gene (Fig. 1), SNPs rs737866 and rs933271 compose block 1; and SNPs rs4646316 and rs165774 compose block 2. In the MAO-A gene (Fig. 2), SNPs rs1465107 and rs1465108 compose block 1; and the SNPs rs6323, rs979606, and rs979605 compose block 2. None of these haplotype blocks were found to be associated with OCD (Figs. 1 and 2).
Fig 1. Catechol-O-methyltransferase haplotype blocks composed by single nucleotide polymorphisms evaluated.
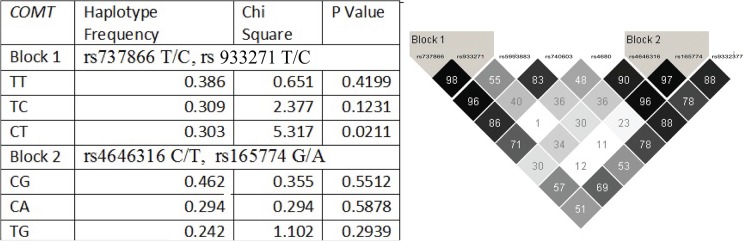
Two haplotype blocks, with two SNPs each, were defined in Cathecol-O-methyltransferase. Block 1 comprises SNP rs737866 and rs933271 and Block 2 comprises SNPs rs4646316 and rs165774. Darker squares mean the high D prime and high LOD scores. Haplotype blocks were defined with D prime scores higher than 95. When D prime is 100, the number is not shown inside the square. Two haplotype blocks, with two SNPs each, were defined in Cathecol-O-methyltransferase. Data shown in the table refers to each haplotypes allele combination association results.
Fig 2. Monoamine oxidase A haplotype Block comprises SNPs rs1465107 and rs1465108 and Block 2 comprises the SNPs rs6323, rs979606, and rs979605.
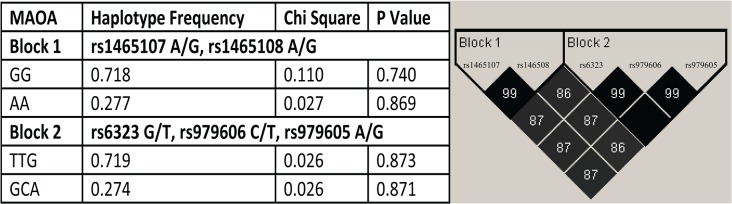
1 composed by single nucleotide polymorphisms evaluated. Darker squares mean the high D prime and high LOD scores. Haplotype blocks were defined with D prime scores higher than 95. When D prime is 100, the number is not shown inside the square. Data shown in the table refers to each haplotypes allele combination association results.
Epistasis analysis
None of the COMT or MAOA SNPs showed an epistatic interaction in association with OCD (data not shown).
Power calculation for whole sample
The statistical power for this association analysis (783 trios, high risk allele frequency = 0.478; OCD prevalence = 0.025; alpha = 0.05) was 65% in average for COMT gene (1120 trios were needed to achieve 80% power) and 58% in average for MAOA (783 trios, high risk allele frequency = 0.257; OCD prevalence = 0.025; alpha = 0.05; and 1305 trios needed to achieve 80% power).
A secondary analysis was performed, with the Brazilian subsample, as described below:
Strictly-defined OCD with comorbid tic disorders
When the Brazilian patients with OCD and comorbid tic disorders (Tourette syndrome or chronic tic disorders; N = 19 trios) were analyzed separately, OCD with comorbid tic disorder was not found to be significantly associated with any COMT or MAO-A SNP (S1 Table).
Broadly-defined OCD phenotype
When the Brazilian subjects meeting criteria for subclinical and clinical OCD are included together, there was preferential transmission of the A allele of the MAO-A SNP rs1465108 (p = 0.0469) (S2 Table). This finding did not remain significant after 100,000 permutations nor after Bonferroni`s correction.
Broadly-defined OCD spectrum disorders
Subsequent analyses revealed that the A allele of the MAO-A SNP rs979605 was associated with OCD spectrum disorders (as defined above: including patients with OCD, subclinical OCD, skin picking, trichotillomania, body dysmorphic disorder, or DSM-IV-defined anxiety disorders) (p = 0.027) (S3 Table). This finding also did not remain significant after 100,000 permutations nor after Bonferroni`s correction.
Power calculation for Brazilian subsample
The statistical power for the analyses performed with the Brazilian subsample (83 trios; high risk allele frequency = 0.39; OCD prevalence = 0.025; alpha = 0.05) was 11% (1282 trios needed for 80% power) for COMT; and for MAOA would be 10.5% power (83 trios; high risk allele frequency = 0.32; OCD prevalence = 0.025; alpha = 0.05; and 1402 trios needed for 80% power)
Discussion
The results presented here do not support the hypothesis that DSM-IV OCD diagnosis is associated with MAO-A or COMT. Results were also negative in the additional analyses performed: 1) one that examined a strictly defined phenotype that consisted of individuals with OCD and tic disorders; 2) one that examined a broadly defined phenotype that included OCD, subclinical OCD; 3) one that included other OCD spectrum disorders; and 4) one COMT-MAOA epistasis analysis.
This result is consistent with the findings of a previous Brazilian study where no association between DSM-IV OCD and the COMT val-158-met (rs4680) variant in a case-control OCD study was observed (Meira-Lima et al. 2004).
As with any genetic association study, the present work is not without its limitations. The main limitation is the small sample size. The statistical power to refute the null hypothesis in this study was 23% (COMT) and 21% (MAOA) for the whole sample and 11% (COMT) and 10.5% (MAOA) for the analyses performed solely on the Brazilian subsample.
The most recent meta-analysis evaluated about 810 OCD cases and found a positive association between COMT rs4680 and OCD in males [41]. These analyses had a power of 49.9% for the total sample and 30.75% for the male sample. As mentioned previously, about 1800 OCD trios were needed to achieve 80% statistical power in a COMT rs4680 and OCD association study [40].
The same meta-analysis evaluated about 452 OCD probands regarding the MAOA EcoRV polymorphism and found a significant association between T allele and OCD in males [41]. This meta-analysis reached 17.2% statistical power for the MAOA polymorphism. The sample number needed to reach 80% power would be more than 2,000 trios [40].
Therefore, the accumulated empirical data to date on the association of COMT and MAOA polymorphisms and OCD are not enough to refute the null hypothesis and thus, additional association studies are needed to add power in future meta-analyses.
Another limitation was the number of tests performed in this study (more than 120 tests). The multitesting could increase the odds of false positive results. None of the results remained significant after Bonferroni`s correction or permutation analysis. Other investigators, who also had small sample studies, have reported associations between COMT and other psychiatric conditions, such as schizophrenia [42,43,44,45,46,47,48,49,50], bipolar disorder [47,50,51]; alcoholism [52,53], substance use disorders [54,55], depression [56,57], and anorexia nervosa [58]. Likewise, MAO-A has been associated with attention deficit hyperactivity disorder [59], anxiety disorders [60], major depressive disorder [61], and other psychiatric conditions [9,62,63]. Besides the odds of some false-positive associations, the non-specificity of the phenotypes associated with the COMT and MAO-A genes might be related to their metabolic functions. Because the COMT and MAO-A enzymes metabolize a number of important neurotransmitters in the limbic pathways, it is possible that, from a phenomenological perspective, impairment of their function could lead to various psychological presentations. Besides, underlying psychopathology of correlated mental disorders are hypothesized to be dimensional and continuous and to share genetic etiologic factors [64].
Additionally, the genetic heterogeneity of the sample could be another limitation, since the Brazilian population is an admixture of individuals of European, African and indigenous descent. However, the use of a family-based approach in conjunction with the `parental discordance test` minimized the odds of population stratification bias.
In conclusion, the present work investigated eight SNPs from COMT gene, five from MAO-A gene and the interaction between these two genes in 783 OCD trios using transmission disequilibrium linkage analyses. Analyses using three phenotype variations (from narrow to broad definitions of OCD phenotypes), showed only no significant association. Despite the lack of association, it appears that the broadly-defined OCD, as well as the OCD spectrum disorders, may be interesting phenotypes to be studied in future association analysis. Further genetic association studies with larger samples involving a more comprehensive coverage of polymorphisms in the COMT and MAO-A genes, as well as other meta-analyses, are needed in order to clarify the biological effect of those genes over the OCD susceptibility phenotypes.
Supporting Information
Legend: SNP: single-nucleotide polymorphism; TDT: transmission/disequilibrium test; OR: TDT odds ratio; CHISQ: TDT chi-square value; P: TDT p value; CHISQ_PAR: parental discordance test chi-square value; P_PAR: parental discordance test p value; CHISQ_COM: combined TDT and parental discordance test chi-square value; P_COM: combined TDT and parental discordance test p value; COMT: catechol-O-methyltransferase; MAO-A: monoamine oxidase-A; NA: not applicable, not enough sample to perform the analysis.
(DOCX)
Legend: SNP: single-nucleotide polymorphism; TDT: transmission/disequilibrium test; OR: TDT odds ratio; CHISQ: TDT chi-square value; P: TDT p value; CHISQ_PAR: parental discordance test chi-square value; P_PAR: parental discordance test p value; CHISQ_COM: combined test chi-square value; P_COM: combined test p value; COMT: catechol-O-methyltransferase; MAO-A: monoamine oxidase-A.
(DOCX)
Legend: SNP: single-nucleotide polymorphism; TDT: transmission/disequilibrium test; OR: TDT odds ratio; CHISQ: TDT chi-square value; P: TDT p value; CHISQ_PAR: parental discordance test chi-square value; P_PAR: parental discordance test p value; CHISQ_COM: combined test chi-square value; P_COM: combined test p value; COMT: catechol-O-methyltransferase; MAO-A: monoamine oxidase-A
(DOCX)
Legend: FAMILY ID: Given code for each family; SUBJECT ID: given code for each subject; FATHER: subject’s father code, 0 if it is not included in the sample; MOTHER: subject’s mother code, 0 if it is not included in the sample; GENDER: subject’s gender = 1 if male, and = 2 if female; DSM OCD: 1 = did not fulfill obsessive-compulsive disorder criteria according the Diagnostic and Statistical Manual for Mental Disorders – forth version, and 2 = did fulfill those criteria; BroadOCD 1 = did not have at least 75% of obsessive-compulsive disorder criteria according the Diagnostic and Statistical Manual for Mental Disorders – forth version, and 2 = did have at least 75% of obsessive-compulsive disorder criteria according the Diagnostic and Statistical Manual for Mental Disorders – forth version; OCD+TIC: 1 = did not fulfill obsessive-compulsive disorder criteria plus Tourette’s syndrome or chronic tic disorder criteria according the Diagnostic and Statistical Manual for Mental Disorders – forth version, and 2 = did fulfill both obsessive-compulsive disorder criteria and Tourette’s syndrome or chronic tic disorder criteria according the Diagnostic and Statistical Manual for Mental Disorders – forth version; SpecOCD: 1 = did not fulfill any of the called Obsessive-compulsive spectrum disorders, which includes broad obsessive-compulsive diagnosis as well as the anxiety disorders, body dysmorphic disorder, and pathological grooming disorders (skin picking and trichotillomania) according the Diagnostic and Statistical Manual for Mental Disorders – forth version.
(XLSX)
Acknowledgments
We acknowledge Jeff Boyles, ELS, from Precise Editing, and Marissa Williams for the English review and editing.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This study received financial support in the form of grants provided by the following Brazilian governmental agencies to Dr. Miguel: the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, National Council for Scientific and Technological Development, Grant number: 573974/2008-0) and the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP, Foundation for the Support of Research in the State of São Paulo, Grant number: 2005/55628-08 and 2008/57896-8); to Dr. Sampaio: by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES – Coordination for Education of Graduated People), Brazil, Fundo de Aprimoramento Acadêmico (FUAA- Grant for Academic Improvement), Department of Psychiatry University of São Paulo School of Medicine. Genotyping expenses were paid by Obsessive-Compulsive Foundation Genetics Collaborative Study. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. American-Psychiatric-Association, editor (2000) Diagnostic and Statistical Manual of Mental Disorders. 4th edition, Text Revision ed. Washington D.C.: American Psychiattic Press. [Google Scholar]
- 2. Pauls DL (2010) The genetics of obsessive-compulsive disorder: a review. Dialogues Clin Neurosci 12: 149–163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. van Grootheest DS, Cath DC, Beekman AT, Boomsma DI (2005) Twin studies on obsessive-compulsive disorder: a review. Twin Res Hum Genet 8: 450–458. [DOI] [PubMed] [Google Scholar]
- 4. Sampaio AS, Lins RMP, Daltro-Oliveira R, Quarantini LdC, Hounie AG, et al. (2013) Systematic Review of Genetic Association Studies in Obsessive-Compulsive Disorder. Revista de Psiquiatria Clínica 40 15279152 [Google Scholar]
- 5. Micallef J, Blin O (2001) Neurobiology and clinical pharmacology of obsessive-compulsive disorder. Clin Neuropharmacol 24: 191–207. [DOI] [PubMed] [Google Scholar]
- 6. Rolls ET, Loh M, Deco G (2008) An attractor hypothesis of obsessive-compulsive disorder. Eur J Neurosci 28: 782–793. 10.1111/j.1460-9568.2008.06379.x [DOI] [PubMed] [Google Scholar]
- 7. Hosak L (2007) Role of the COMT gene Val158Met polymorphism in mental disorders: a review. Eur Psychiatry 22: 276–281. [DOI] [PubMed] [Google Scholar]
- 8. Lachman HM, Papolos DF, Saito T, Yu YM, Szumlanski CL, et al. (1996) Human catechol-O-methyltransferase pharmacogenetics: Description of a functional polymorphism and its potential application to neuropsychiatric disorders. Pharmacogenetics 6: 243–250. [DOI] [PubMed] [Google Scholar]
- 9. Bearden CE, Jawad AF, Lynch DR, Monterossso JR, Sokol S, et al. (2005) Effects of COMT genotype on behavioral symptomatology in the 22q11.2 Deletion Syndrome. Child Neuropsychol 11: 109–117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Gothelf D, Michaelovsky E, Frisch A, Zohar AH, Presburger G, et al. (2007) Association of the low-activity COMT 158Met allele with ADHD and OCD in subjects with velocardiofacial syndrome. Int J Neuropsychopharmacol 10: 301–308. [DOI] [PubMed] [Google Scholar]
- 11. Brunner HG, Nelen M, Breakefield XO, Ropers HH, van Oost BA (1993) Abnormal behavior associated with a point mutation in the structural gene for monoamine oxidase A. Science 262: 578–580. [DOI] [PubMed] [Google Scholar]
- 12. Brunner HG, Nelen MR, van Zandvoort P, Abeling NG, van Gennip AH, et al. (1993) X-linked borderline mental retardation with prominent behavioral disturbance: phenotype, genetic localization, and evidence for disturbed monoamine metabolism. Am J Hum Genet 52: 1032–1039. [PMC free article] [PubMed] [Google Scholar]
- 13.Taylor S (2013) Molecular genetics of obsessive-compulsive disorder: a comprehensive meta-analysis of genetic association studies. Mol Psychiatry. [DOI] [PubMed]
- 14. Miguel EC, Leckman JF, Rauch S, do Rosario-Campos MC, Hounie AG, et al. (2005) Obsessive-compulsive disorder phenotypes: implications for genetic studies. Mol Psychiatry 10: 258–275. [DOI] [PubMed] [Google Scholar]
- 15. Pauls DL, Alsobrook JP 2nd, Goodman W, Rasmussen S, Leckman JF (1995) A family study of obsessive-compulsive disorder. Am J Psychiatry 152: 76–84. [DOI] [PubMed] [Google Scholar]
- 16. Pauls DL, Raymond CL, Stevenson JM, Leckman JF (1991) A family study of Gilles de la Tourette syndrome. Am J Hum Genet 48: 154–163. [PMC free article] [PubMed] [Google Scholar]
- 17.Bienvenu OJ, Samuels JF, Wuyek LA, Liang KY, Wang Y, et al. Is obsessive-compulsive disorder an anxiety disorder, and what, if any, are spectrum conditions? A family study perspective. Psychol Med: 1–13. [DOI] [PMC free article] [PubMed]
- 18. Nestadt G, Di CZ, Riddle MA, Grados MA, Greenberg BD, et al. (2009) Obsessive-compulsive disorder: subclassification based on co-morbidity. Psychol Med 39: 1491–1501. 10.1017/S0033291708004753 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Seixas AA, Hounie AG, Fossaluza V, Curi M, Alvarenga PG, et al. (2008) Anxiety disorders and rheumatic Fever: is there an association? CNS Spectr 13: 1039–1046. [DOI] [PubMed] [Google Scholar]
- 20. Stewart SE, Yu D, Scharf JM, Neale BM, Fagerness JA, et al. Genome-wide association study of obsessive-compulsive disorder. Mol Psychiatry 18: 788–798. 10.1038/mp.2012.85 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. First M SR, Gibbon M, Williams J, editor (1996) Structured Clinical Interview for DSM-IV Axis I Disorders, Clinician Version. Washington, DC: American Psychiatric Press, Inc. [Google Scholar]
- 22. Kaufman J BB, Brent D, Rao U, Flynn C, Moreci P, Williamson D, Ryan N. (1997) Schedule for affective disorders and schizophrenia for school age children – Present and lifetime version (KSADS-PL): initial reliability and validity data. J Am Acad Child Adolesc Psychiatry 36: 980–988. [DOI] [PubMed] [Google Scholar]
- 23. Goodman W PL, Rasmussen S, Mazure C, Fleischman R, Hill C, Heninger G, Charney D. (1989) The Yale-Brown Obsessive Compulsive Scale: I. Development use and reliability. Arch Gen Psychiatry 46: 1006–1011. [DOI] [PubMed] [Google Scholar]
- 24. Leckman J RM, Hardin M, Ort S, Swartz K, Steveson J, Cohen D (1989) The Yale Global Tic Severity Scale: Initial testing of a clinician-rated scale of tic severity. J Am Acad Child Adolesc Psychiatry 28: 566–573. [DOI] [PubMed] [Google Scholar]
- 25. Miguel EC, Ferrao YA, do Rosario MC, de Mathis MA, Torres AR, et al. (2008) The Brazilian Research Consortium on Obsessive-Compulsive Spectrum Disorders: recruitment, assessment instruments, methods for the development of multicenter collaborative studies and preliminary results. Rev Bras Psiquiatr 30: 185–196. [DOI] [PubMed] [Google Scholar]
- 26. Pallanti S, Hollander E (2008) Obsessive-compulsive disorder spectrum as a scientific "metaphor". CNS Spectr 13: 6–15. [DOI] [PubMed] [Google Scholar]
- 27. Nestadt G, Addington A, Samuels J, Liang KY, Bienvenu OJ, et al. (2003) The identification of OCD-related subgroups based on comorbidity. Biol Psychiatry 53: 914–920. [DOI] [PubMed] [Google Scholar]
- 28.Bienvenu OJ, Samuels JF, Wuyek LA, Liang KY, Wang Y, et al. (2012) Is obsessive-compulsive disorder an anxiety disorder, and what, if any, are spectrum conditions? A family study perspective. Psychol Med: 1–13. [DOI] [PMC free article] [PubMed]
- 29. de Mathis MA, Diniz JB, do Rosario MC, Torres AR, Hoexter M, et al. (2006) What is the optimal way to subdivide obsessive-compulsive disorder? CNS Spectr 11: 762–768, 771–764, 776–769. [DOI] [PubMed] [Google Scholar]
- 30. de Mathis MA, do Rosario MC, Diniz JB, Torres AR, Shavitt RG, et al. (2008) Obsessive-compulsive disorder: influence of age at onset on comorbidity patterns. Eur Psychiatry 23: 187–194. 10.1016/j.eurpsy.2008.01.002 [DOI] [PubMed] [Google Scholar]
- 31. Miller SA, Dykes DD, Polesky HF (1988) A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res 16: 1215 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.DeBakker TAGGER
- 33. Riva A, Kohane IS (2002) SNPper: retrieval and analysis of human SNPs. Bioinformatics 18: 1681–1685. [DOI] [PubMed] [Google Scholar]
- 34. Purcell S, Neale B, Todd-Brown K, Thomas L, Ferreira MAR, et al. (2007) PLINK: a toolset for whole-genome association and population-based linkage analysis. American Journal of Human Genetics 81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Purcell S PLINK v. 1.06. 1.06 ed. pp. Software. Available: http://pngu.mgh.harvard.edu/purcell/plink/.
- 36. Barrett JC FB, Maller J, Daly MJ. (2005) Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics 21: 263–265. [DOI] [PubMed] [Google Scholar]
- 37. Sabeti PC, Reich DE, Higgins JM, Levine HZ, Richter DJ, et al. (2002) Detecting recent positive selection in the human genome from haplotype structure. Nature 419: 832–837. [DOI] [PubMed] [Google Scholar]
- 38. Purcell S, Sham P, Daly MJ (2005) Parental phenotypes in family-based association analysis. Am J Hum Genet 76: 249–259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Dawling S, Roodi N, Mernaugh RL, Wang X, Parl FF (2001) Catechol-O-methyltransferase (COMT)-mediated metabolism of catechol estrogens: comparison of wild-type and variant COMT isoforms. Cancer Res 61: 6716–6722. [PubMed] [Google Scholar]
- 40. Purcell S CS, Sham PC. (2003) Genetic Power Calculator: design of linkage and association genetic mapping studies of complex traits. Bioinformatics 19: 149–150. [DOI] [PubMed] [Google Scholar]
- 41. Taylor S (2013) Molecular genetics of obsessive-compulsive disorder: a comprehensive meta-analysis of genetic association studies. Mol Psychiatry 18: 799–805. 10.1038/mp.2012.76 [DOI] [PubMed] [Google Scholar]
- 42. Lee SG, Joo Y, Kim B, Chung S, Kim HL, et al. (2005) Association of Ala72Ser polymorphism with COMT enzyme activity and the risk of schizophrenia in Koreans. Hum Genet 116: 319–328. [DOI] [PubMed] [Google Scholar]
- 43. Li T, Sham PC, Vallada H, Xie T, Tang X, et al. (1996) Preferential transmission of the high activity allele of COMT in schizophrenia. Psychiatr Genet 6: 131–133. [DOI] [PubMed] [Google Scholar]
- 44. Nicodemus KK, Kolachana BS, Vakkalanka R, Straub RE, Giegling I, et al. (2007) Evidence for statistical epistasis between catechol-O-methyltransferase (COMT) and polymorphisms in RGS4, G72 (DAOA), GRM3, and DISC1: influence on risk of schizophrenia. Hum Genet 120: 889–906. [DOI] [PubMed] [Google Scholar]
- 45. Palmatier MA, Pakstis AJ, Speed W, Paschou P, Goldman D, et al. (2004) COMT haplotypes suggest P2 promoter region relevance for schizophrenia. Mol Psychiatry 9: 859–870. [DOI] [PubMed] [Google Scholar]
- 46. Sanders AR, Rusu I, Duan J, Vander Molen JE, Hou C, et al. (2005) Haplotypic association spanning the 22q11.21 genes COMT and ARVCF with schizophrenia. Mol Psychiatry 10: 353–365. [DOI] [PubMed] [Google Scholar]
- 47. Shifman S, Bronstein M, Sternfeld M, Pisante A, Weizman A, et al. (2004) COMT: a common susceptibility gene in bipolar disorder and schizophrenia. Am J Med Genet B Neuropsychiatr Genet 128B: 61–64. [DOI] [PubMed] [Google Scholar]
- 48. Shifman S, Bronstein M, Sternfeld M, Pisante-Shalom A, Lev-Lehman E, et al. (2002) A highly significant association between a COMT haplotype and schizophrenia. Am J Hum Genet 71: 1296–1302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Wonodi I, Stine OC, Mitchell BD, Buchanan RW, Thaker GK (2003) Association between Val108/158 Met polymorphism of the COMT gene and schizophrenia. Am J Med Genet B Neuropsychiatr Genet 120B: 47–50. [DOI] [PubMed] [Google Scholar]
- 50. Abdolmaleky HM, Cheng KH, Faraone SV, Wilcox M, Glatt SJ, et al. (2006) Hypomethylation of MB-COMT promoter is a major risk factor for schizophrenia and bipolar disorder. Hum Mol Genet 15: 3132–3145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.(1997) No association between bipolar disorder and alleles at a functional polymorphism in the COMT gene. Biomed European Bipolar Collaborative Group. Br J Psychiatry 170: 526–528. [DOI] [PubMed] [Google Scholar]
- 52. Sery O, Didden W, Mikes V, Pitelova R, Znojil V, et al. (2006) The association between high-activity COMT allele and alcoholism. Neuro Endocrinol Lett 27: 231–235. [PubMed] [Google Scholar]
- 53. Tiihonen J, Hallikainen T, Lachman H, Saito T, Volavka J, et al. (1999) Association between the functional variant of the catechol-O-methyltransferase (COMT) gene and type 1 alcoholism. Mol Psychiatry 4: 286–289. [DOI] [PubMed] [Google Scholar]
- 54. Li T, Chen CK, Hu X, Ball D, Lin SK, et al. (2004) Association analysis of the DRD4 and COMT genes in methamphetamine abuse. Am J Med Genet B Neuropsychiatr Genet 129B: 120–124. [DOI] [PubMed] [Google Scholar]
- 55. Shiels MS, Huang HY, Hoffman SC, Shugart YY, Bolton JH, et al. (2008) A community-based study of cigarette smoking behavior in relation to variation in three genes involved in dopamine metabolism: Catechol-O-methyltransferase (COMT), dopamine beta-hydroxylase (DBH) and monoamine oxidase-A (MAO-A). Prev Med 47: 116–122. 10.1016/j.ypmed.2008.03.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Massat I, Souery D, Del-Favero J, Nothen M, Blackwood D, et al. (2005) Association between COMT (Val158Met) functional polymorphism and early onset in patients with major depressive disorder in a European multicenter genetic association study. Mol Psychiatry 10: 598–605. [DOI] [PubMed] [Google Scholar]
- 57. Wray NR, James MR, Dumenil T, Handoko HY, Lind PA, et al. (2008) Association study of candidate variants of COMT with neuroticism, anxiety and depression. Am J Med Genet B Neuropsychiatr Genet 147B: 1314–1318. 10.1002/ajmg.b.30744 [DOI] [PubMed] [Google Scholar]
- 58. Michaelovsky E, Frisch A, Leor S, Stein D, Danziger Y, et al. (2005) Haplotype analysis of the COMT-ARVCF gene region in Israeli anorexia nervosa family trios. Am J Med Genet B Neuropsychiatr Genet 139B: 45–50. [DOI] [PubMed] [Google Scholar]
- 59. Manor I, Tyano S, Mel E, Eisenberg J, Bachner-Melman R, et al. (2002) Family-based and association studies of monoamine oxidase A and attention deficit hyperactivity disorder (ADHD): preferential transmission of the long promoter-region repeat and its association with impaired performance on a continuous performance test (TOVA). Mol Psychiatry 7: 626–632. [DOI] [PubMed] [Google Scholar]
- 60. Samochowiec J, Hajduk A, Samochowiec A, Horodnicki J, Stepien G, et al. (2004) Association studies of MAO-A, COMT, and 5-HTT genes polymorphisms in patients with anxiety disorders of the phobic spectrum. Psychiatry Res 128: 21–26. [DOI] [PubMed] [Google Scholar]
- 61. Yu YW, Tsai SJ, Hong CJ, Chen TJ, Chen MC, et al. (2005) Association study of a monoamine oxidase a gene promoter polymorphism with major depressive disorder and antidepressant response. Neuropsychopharmacology 30: 1719–1723. [DOI] [PubMed] [Google Scholar]
- 62. Avramopoulos D, Stefanis NC, Hantoumi I, Smyrnis N, Evdokimidis I, et al. (2002) Higher scores of self reported schizotypy in healthy young males carrying the COMT high activity allele. Mol Psychiatry 7: 706–711. [DOI] [PubMed] [Google Scholar]
- 63. Barr CL, Wigg KG, Sandor P (1999) Catechol-O-methyltransferase and Gilles de la Tourette syndrome. Molecular Psychiatry 4: 492–495. [DOI] [PubMed] [Google Scholar]
- 64. Lahey BB, Van Hulle CA, Singh AL, Waldman ID, Rathouz PJ (2011) Higher-order genetic and environmental structure of prevalent forms of child and adolescent psychopathology. Arch Gen Psychiatry 68: 181–189. 10.1001/archgenpsychiatry.2010.192 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Legend: SNP: single-nucleotide polymorphism; TDT: transmission/disequilibrium test; OR: TDT odds ratio; CHISQ: TDT chi-square value; P: TDT p value; CHISQ_PAR: parental discordance test chi-square value; P_PAR: parental discordance test p value; CHISQ_COM: combined TDT and parental discordance test chi-square value; P_COM: combined TDT and parental discordance test p value; COMT: catechol-O-methyltransferase; MAO-A: monoamine oxidase-A; NA: not applicable, not enough sample to perform the analysis.
(DOCX)
Legend: SNP: single-nucleotide polymorphism; TDT: transmission/disequilibrium test; OR: TDT odds ratio; CHISQ: TDT chi-square value; P: TDT p value; CHISQ_PAR: parental discordance test chi-square value; P_PAR: parental discordance test p value; CHISQ_COM: combined test chi-square value; P_COM: combined test p value; COMT: catechol-O-methyltransferase; MAO-A: monoamine oxidase-A.
(DOCX)
Legend: SNP: single-nucleotide polymorphism; TDT: transmission/disequilibrium test; OR: TDT odds ratio; CHISQ: TDT chi-square value; P: TDT p value; CHISQ_PAR: parental discordance test chi-square value; P_PAR: parental discordance test p value; CHISQ_COM: combined test chi-square value; P_COM: combined test p value; COMT: catechol-O-methyltransferase; MAO-A: monoamine oxidase-A
(DOCX)
Legend: FAMILY ID: Given code for each family; SUBJECT ID: given code for each subject; FATHER: subject’s father code, 0 if it is not included in the sample; MOTHER: subject’s mother code, 0 if it is not included in the sample; GENDER: subject’s gender = 1 if male, and = 2 if female; DSM OCD: 1 = did not fulfill obsessive-compulsive disorder criteria according the Diagnostic and Statistical Manual for Mental Disorders – forth version, and 2 = did fulfill those criteria; BroadOCD 1 = did not have at least 75% of obsessive-compulsive disorder criteria according the Diagnostic and Statistical Manual for Mental Disorders – forth version, and 2 = did have at least 75% of obsessive-compulsive disorder criteria according the Diagnostic and Statistical Manual for Mental Disorders – forth version; OCD+TIC: 1 = did not fulfill obsessive-compulsive disorder criteria plus Tourette’s syndrome or chronic tic disorder criteria according the Diagnostic and Statistical Manual for Mental Disorders – forth version, and 2 = did fulfill both obsessive-compulsive disorder criteria and Tourette’s syndrome or chronic tic disorder criteria according the Diagnostic and Statistical Manual for Mental Disorders – forth version; SpecOCD: 1 = did not fulfill any of the called Obsessive-compulsive spectrum disorders, which includes broad obsessive-compulsive diagnosis as well as the anxiety disorders, body dysmorphic disorder, and pathological grooming disorders (skin picking and trichotillomania) according the Diagnostic and Statistical Manual for Mental Disorders – forth version.
(XLSX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.