

Abstract
Poststroke depression, the second most serious psychosomatic complication after brain stroke, leads to delay of the rehabilitation process and is associated with an increased disability and cognitive impairment along with increase in term mortality. Research into the biochemical changes in depression is still insufficiently described. The aim of our study was therefore to evaluate the possible association between plasma protein oxidative/nitrative damages and the development of poststroke depression. We evaluated oxidative/nitrative modifications of specific proteins by measurement of 3-nitrotyrosine and carbonyl groups levels using ELISA test. Additionally, we checked differences in proteins thiol groups by spectrophotometric assay based on reaction between DTNB and thiols. We also evaluated catalase activity in erythrocytes measured as ability to decompose H2O2. Correlation analysis was performed using Spearman's rank. We observed significant (P < 0.001) differences in all oxidative/nitrative stress parameters in brain stroke patients compared to healthy group. Our research shows that oxidative damage of proteins is correlated with the degree of poststroke depression, while nitrative changes do not show any relationship. We demonstrate a positive correlation between the concentration of carbonyl groups and the Geriatric Depression Scale and a negative correlation between the degree of depression and the concentration of -SH groups or catalase activity.
1. Introduction
In the past few years, notable attention is paid to mental disorders particularly prevalent among stroke patients, affecting about one-third of population. Physical disability and psychological stress occurring due to stroke need psychological treatment conducted together with physical treatment and rehabilitation [1]. Although affective disorders have not been formally established as independent risk factor for stroke and adverse prognostic factor responsible for impeding convalescing patients, attention to this physical/psychological symptom has continued to increase in the past decade [2]. A growing body of evidence acknowledges the existence of poststroke depression (PSD) [3].
Studies suggest that poststroke depression is the second most serious psychosomatic complication after brain stroke. Depression can occur at any stage after stroke (acute, subacute, and chronic), although some authors classify this condition as common mainly in the second phase of the disease [4]. Poststroke depression is associated with an increased disability and increased cognitive impairment and leads to delay of the rehabilitation process, due to the deterioration of motivation and low mood [5]. What is important in patients with PSD appears to be 3-4-fold increase in short- and long-term mortality compared with nondepressed patients [6].
However, research into the biochemical changes in depression is still not systematized and is lacking in both quality and quantity. Our aim in this study was therefore to evaluate the possible association between plasma protein oxidative and nitrative damages and the development of PSD in patients with acute ischemic stroke. In the past few years, oxidative stress has received much notice with regard to psychiatric illnesses including depression and it has been proposed as a contributing factor in the pathogenesis of depression [7]. Measurements of impaired redox balance after the ischemic stroke occurrence might be useful to monitor the outcome of patients who suffered an ischemic stroke in terms of stroke recurrence and other vascular events. This is particularly important for patients whose convalescence may be slowed down due to the presence of PSD.
Oxidative stress is thought to be a contributing factor in many chronic neurodegenerative pathologies, as well as acute cerebrovascular disorders such as stroke. The pathogenesis of acute ischemic stroke is highly complex and embroils multiple mechanisms. It is well proven that oxidative stress is involved in the pathogenesis of acute ischemic stroke. In ischemic stroke, blood flow is interrupted to a portion of the brain by a formation of clot blocking an artery. The later reperfusion of the engaged brain area when blood flow is restored is associated with a rapid increase in oxidative damage. Imbalance between the cellular production of free radicals and the ability of cells to defend against them is one of key mechanisms contributing to neuronal damage [8]. These conditions support oxidative and nitrative modifications in some amino acids. Proteins constitute one of the major targets of ROS/RNS (reactive oxygen/nitrogen species). Oxidative and nitrative changes in proteins include carbonyl groups formation, oxidation of the thiol groups and nitrotyrosine (3-NT) generation [9].
Some neurological disorders, including stroke, are associated with oxidative and nitrative modification of specific proteins and the accumulation of oxidative damage. There are several evidences that markedly indicate involvement of oxidative and nitrative stress in the pathophysiology of depression [10, 11]. Increased levels of ROS and RNS in depression, mainly peroxide [10] and NO [12, 13], and decreased levels of antioxidants, such as glutathione (GSH), in the postmortem depression brain have been demonstrated [14]. For this reason, oxidative and nitrative mechanisms have been proposed as targets for novel antidepressants [15].
Oxidative stress leads to inactivation and modification of antioxidant enzymes and weakening of antioxidant defense. The oxidation of proteins can be responsible for their conversion to forms more susceptible to degradation by proteinases [8, 16]. Protein carbonyl formation is an index of oxidative stress, as a result of amino acid modifications. Carbonylation of proteins is a nonenzymatic addition of aldehydes or ketones to specific amino acid residues, mainly to arginine, lysine, threonine, proline, cysteine, or histidine [17].
Shifts of redox-equilibrium in thiol-disulfide plasma system are implicated as a key mechanism of oxidative damage and restoration of redox homeostasis in postacute stroke patients. ROS-dependent oxidative modification of redox active cysteines within proteins is responsible for regulation of activity of a variety of protein functions, including enzyme activity, protein expression and abundance, protein localization, and interaction with other biomolecules in controlling of cell signaling and gene expression [18]. Oxidized cysteine residues can react with nitric oxide (NO) to form a nitrosothiol or with another protein thiol group to give a disulfide bond [19].
The nitrative stress can promote nitrosative posttranslational modifications of proteins that are implicated in the pathogenesis of the associated cardiovascular dysfunction [20, 21]. However, simultaneous formation of nitric oxide and superoxide radicals may reduce S-nitrosylation of protein due to concurrently generating the potent tyrosine-nitrating agent peroxynitrite (ONOO−). Protein nitration and accumulation of 3-nitrotyrosine is a biomarker of nitrative stress induced by peroxynitrite and other reactive nitrogen species. Tyrosine nitration as an usual modification is also a relatively specific marker of oxidative damage mediated by ONOO−, which is formed by the reaction of superoxide with nitric oxide and is important for the regulation of protein structure and function [22].
The antioxidant defense system in the organism comprises enzymatic and nonenzymatic components. In the pathogenesis of acute ischemic stroke low molecular antioxidants are consumed in the reaction with free radicals generated during the oxidative stress. The endogenous catalytic system includes a variety of antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase. Among them, the first line of defense is a catalase which decomposes hydrogen peroxide, since its accumulation can lead to the formation of free radicals [23, 24]. The body of evidence suggests that depression belongs to the spectrum of (neuro)degenerative disorders and is accompanied by a decreased antioxidant status and by induction of oxidative and nitrative pathways. A significant association between depression and polymorphisms in genes involved in oxidative and nitrative pathways, like manganese superoxide dismutase, catalase, and myeloperoxidase, is also known [10].
In our study we compare the activities of CAT in the erythrocytes of PSD patients and nondepression patients.
Our study was aimed to investigate the dynamic of some oxidative and nitrative markers of plasma proteins during the convalescent of postacute stroke patients with depression.
2. Materials and Methods
2.1. Chemicals
Anti-dinitrophenyl (DNP) antibodies developed in rabbit, bicinchoninic acid protein assay kit, o-phenylenediamine dihydrochloride (OPD peroxidase substrate), and DNTB (5,5′-dithiobis(2-nitrobenzoic acid)) were purchased from Sigma-Aldrich (St. Louis, MO, USA). 2,4-Dinitrophenylhydrazine (DNPH) was obtained from Polish Chemical Reagents (POCH, Gliwice). All other reagents were of analytical grade and were provided by commercial suppliers.
2.2. Patients
35 patients following ischemic stroke were enrolled for the study and 29 poststroke patients were carefully selected who met inclusion criteria and did not meet any criterion for exclusion. A total of 35 patients (aged 74.3 ± 16.3) with moderate stroke severity (NIHSS scores 5.8 ± 3.5) who agreed to participate were included in this analysis. 29 patients were screened for positive results for depressive symptoms (GDS ≥ 11). Subjects with and without depressive symptoms were similar in demographics and clinical characteristics (Table 1). Patients with a medical history of prestroke dementia, hemorrhagic stroke, decreased consciousness, significant acute or chronic inflammatory factor or neurological illness other than stroke, and psychiatric disorder other than depression were excluded. All blood samples (control groups and patients) were taken in the morning (between 7 am to 9 am) in fasting status and stored using the same protocol. The protocol was approved by the Ethics Committee of the Faculty of Biology and Environmental Protection of University of Lodz, Poland number 16/2013. All participants provided written informed consent prior to participation.
Table 1.
Clinical demographic characteristics. ADL: activity daily living; BMI: body mass index; GDS: Geriatric Depression Scale; NIHSS: National Institutes of Health Stroke Scale; NSAID: nonsteroidal anti-inflammatory; ASA: acetylsalicylic acid.
| GDS < 11 (non-D) n = 6 |
GDS ≥ 11 (D) n = 29 |
Healthy n = 20 |
|
|---|---|---|---|
| Demographics | |||
| Age (mean ± SD) | 70.0 ± 14.6 | 74.3 ± 16.3 | 53.9 ± 10.6 |
| Sex (% male) | 66.7 | 47.2 | 25.6 |
| Living alone (%) | 48.2 | 56.7 | 33.8 |
|
| |||
| Vascular risk | |||
| Hypertension (%) | 92.6 | 97.9 | |
| Diabetes (%) | 20.3 | 27.8 | |
| Dyslipidemia (%) | 65.8 | 76.8 | |
| BMI ≥ 30 (%) | 16.8 | 27.9 | |
|
| |||
| Concomitant medications | |||
| Antidepressants (%) | 3.4 | 23.7 | |
| ASA | 62.8 | 78.8 | |
| NSAID | 8.7 | 20.8 | |
|
| |||
| Stroke characteristics | |||
| Weeks since stroke (mean ± SD) | 5.3 ± 3.7 | 5.6 ± 4.2 | |
| NIHSS scores mean ± SD | 5.2 ± 4.7 | 5.8 ± 3.5 | |
| ADL mean ± SD | 11 ± 5.6 | 10.7 ± 6.7 | |
|
| |||
| Lesion location | |||
| Anterior (%) | 12.0 | 15.8 | |
| Posterior (%) | 52.6 | 36.7 | |
| Intermediate (%) | 12.0 | 27.0 | |
|
| |||
| Lesion side | |||
| Left (%) | 56.7 | 58.4 | |
| Right (%) | 43.3 | 41.6 | |
Depression was screened using the Geriatric Depression Scale (GDS) on which a score ≥11 is a reliable and sensitive indicator of poststroke depression [13]. A trained psychologist researcher administered the GDS scale. The GDS, ADL, and NIHSS were administered either on the same day as the blood draw or on the afternoon before.
2.3. Determination of Protein Carbonyl Groups in Human Blood Plasma by ELISA Method
The detection of protein carbonyl groups was performed in human blood plasma using ELISA method described by Buss et al. [25] and modified by Alamdari et al. [26]. The microplates were incubated overnight at 4°C where plasma proteins were nonspecifically adsorbed onto ELISA plates. The wells were washed 3 times with 300 μL PBS, reacted with dinitrophenylhydrazine (DNPH; 0.05 mM, 200 μL, pH 6.2), incubated for 45 min at room temperature in the dark, and washed 5 times with 300 μL PBS: ethanol (1 : 1, v/v) and a last time with 300 μL PBS, all in accordance with Alamdari et al. [26]. Carbonyl groups were detected by anti-DNP antibodies (I step) and then a second antibody conjugated with horseradish peroxidase was added for quantification (II step) according to Alamdari et al. [26] procedure. This sensitive method was calibrated using oxidized albumin and required only 5 μg protein per well. The standard curve was linear in the range of 0–3.36 nmol carbonyls/mg albumin [27].
2.4. Determination of Thiol Groups
Thiol groups in blood plasma proteins were determined using 5,5′-dithio-bis(2-nitro-benzoic acid) (Ellman's reagent, DTNB) [28]. The thiol-disulfide interchange reaction between DTNB and thiol is the basis of this spectrophotometric assay. The samples were mixed with DTNB and at the end of the incubation (1 h, 37°C) period the absorbances were recorded at 412 nm. The concentrations were calculated by using ε = 13600 M−1 cm−1 and the results were expressed as μmol/L.
2.5. Determination of 3-Nitrotyrosine in Plasma Proteins by the Competitive ELISA Test
The detection of 3-nitrotyrosine in blood plasma was performed according to the method described by Khan et al. [29] and was modified as was described previously [27]. The concentrations of nitrated plasma proteins were estimated from the standard curve, constructed with the use of 3-nitrotyrosine-containing fibrinogen (3-NT-Fg). 3-NT-Fg was prepared by the exposure of human fibrinogen to 1000 μM peroxynitrite action. The presence of 3-nitrotyrosine in fibrinogen was confirmed spectrophotometrically at 302 nm, at pH 11.5 (molar absorption coefficient for nitrotyrosine, ε = 4400 M−1 × cm−1) [30], and then the obtained nitro-fibrinogen was used for the preparation of the standard curve, ranging from 10 to 1000 nmol/L of 3-nitrotyrosine-fibrinogen equivalent.
2.6. Catalase Activity Estimation
Catalase activity in erythrocytes was estimated spectrophotometrically using Beers and Sizer method [31]. Changes in absorbance were recorded at 240 nm. CAT activity was calculated in terms of nM H2O2 consumed/min using molar extinction coefficient ε = 36000 M−1 cm−1 and shows as Bergmeyer unit [32].
2.7. Statistical Analysis
The statistical analysis was performed using StatsDirect statistical software V. 2.7.2. All values in this study were expressed as mean ± SD. The obtained results were analyzed under the account of normality with Shapiro-Wilk test. The significance of differences between the values was analyzed depending on the normality by unpaired t-Student or U-Mann-Whitney tests. Correlation analysis between obtained numerical data was performed using Spearman's rank correlation [33]. A level of P < 0.05 was accepted as statistically significant.
3. Results
In our study we determined organism oxidative and nitrative stress parameters in stroke patients, especially in patients with poststroke depression. A comparative analysis shows that oxidative and nitrative stress parameters are significantly higher in stroke patients (both patients with PSD and nondepression patients) than in control groups (healthy people). All measured parameters were changed in stroke group (Figures 1–4). We observed a statistically significant increased level of carbonyl groups in plasma proteins (Figure 1), decreased level of plasma protein thiol groups (Figure 2), decreased catalase activity in erythrocytes (Figure 3), and increased plasma protein 3-nitrotyrosine level (Figure 4) in stroke patients in relation to control group. In the case of carbonyl groups and 3-nitrotyrosine, the observed increase in oxidative and nitrative protein damage after stroke was more than two times higher in patients than in controls (Figures 1 and 4).
Figure 1.
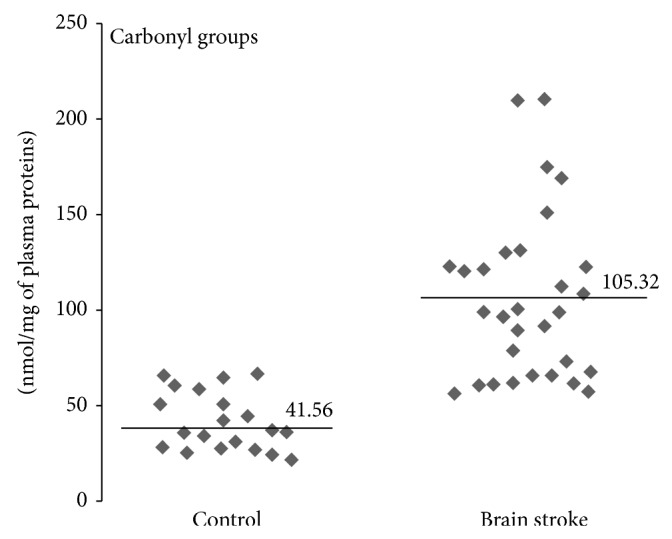
Differences of carbonyl group levels between stroke patients and control group. Carbonyl group level was measured in plasma proteins by ELISA method, n = 31, P < 0.0001.
Figure 4.
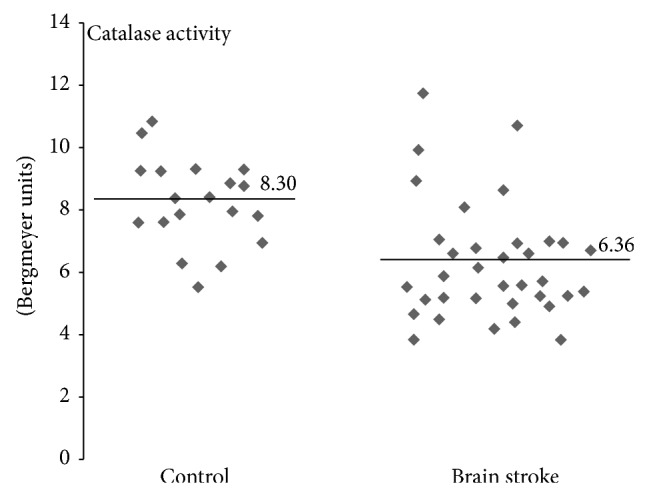
Differences of catalase activity between stroke patients and control group. Catalase activity was measured in erythrocytes using H2O2 disproportionation method, n = 35, P < 0.0001.
Figure 2.
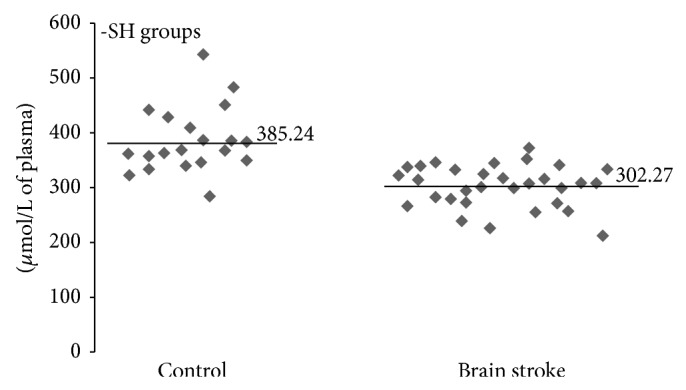
Differences of thiol group levels between stroke patients and control group. The concentration of thiol groups was measured in plasma proteins using Ellman's reagent, n = 31, P < 0.0001.
Figure 3.
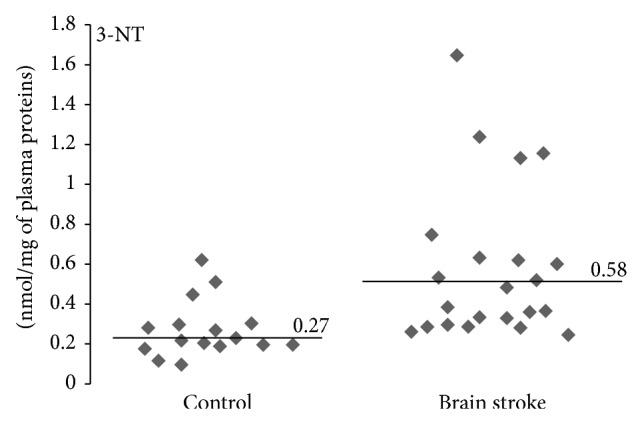
Differences of 3-nitrotyrosine levels between stroke patients and control group. 3-Nitrotyrosine level was measured in plasma proteins by c-ELISA method, n = 22, P < 0.0001.
Additionally, we observed dependence between changes in oxidative and nitrative stress parameters and poststroke depression intensity measured by Geriatric Depression Scale (GDS). As the distribution of the results of the GDS deviates from the normal, the method of Spearman correlation was more suitable for use in these analyzes. Obtained correlation parameters (Table 2) indicate the significant positive correlation between carbonyl group level and GDS, whereas between thiol group concentration/catalase activity and GDS we observed a significant negative correlation. For nitrative stress the marker which is a 3-nitrotyrosine showed no correlation with GDS. For dependence of carbonyl group level (Figure 5(a)), thiol groups level (Figure 5(b)), catalase activity (Figure 5(c)), and GDS regression plots were determined. We have shown the detailed course of these correlations, which confirms correlation and received numeric data presented in Table 2.
Table 2.
Correlation coefficients values obtained for oxidative stress parameters and Geriatric Depression Scale. Correlation was analyzed using Spearman's rank correlation method and probability for correlation was presented in parentheses.
| Carbonyl groups | Thiol groups | Catalase activity | 3-Nitrotyrosine | |
|
| ||||
| Spearman's rank correlation coefficient (Rho) | 0.578941 (P = 0.0014) |
−0.519103 (P = 0.0073) |
−0.57903 (P = 0.0028) |
0.071829 (P = 0.754) |
Figure 5.

Regression plots of selected parameters of oxidative stress in patients with poststroke depression and GDS. (a) Plot for correlation between carbonyl group level and GDS; (b) plot for correlation between thiol group level and GDS; (c) plot for correlation between catalase activity and GDS.
4. Discussion
The development of poststroke depression is a frequent mood disorder that affects around one-third of stroke patients; however, the etiology of PSD is not well understood. Probably, both the psychosocial and biological factors are significant in the development of poststroke depression [34].
Previous studies have identified relationships between PSD and increased functional disability [35, 36]. In the last studies poststroke depression was also associated with inflammation [37]. Previous studies have reported the elevated levels of some inflammatory mediators in PSD, including IL-18, [38, 39], C-reactive protein [40, 41], and IL-17 which is able to disrupt the blood brain barrier and contribute to neurodegeneration through increased production of reactive oxygen species [42, 43].
The brain with its high oxygen consumption and a lipid-rich environment is considered highly susceptible to oxidative stress or redox imbalances. Therefore, it is very probable that oxidative stress is implicated in several mental disorders, including depression. Although some studies have established a link between oxidative stress and psychiatric disorders, the causal relationship between them is not fully determined. Several mechanisms including genetic predisposition, abnormalities in traditional signal transduction pathways, and oxidative stress theory have been proposed to be involved in depression pathogenesis [7, 44, 45]. It has been demonstrated that depression in medically healthy patients has been associated with increased concentrations of blood cytokines [46], elevated levels of ROS and RNS [10–13], and increased F2 isoprostane excretion [47]. Moreover, considerably lower plasma concentrations of several key antioxidants, such as vitamin E, zinc, and coenzyme Q10, and reduced antioxidant enzyme activity, as well as reduced level of brain GSH, have been reported in major depression [10, 14]. The existence of a possible link between depression and central nervous system (CNS) inflammation is suggested in a few recent papers on the neuroimmunology of stroke [48, 49]. The mechanisms related with PSD may involve an imbalance between proinflammatory and anti-inflammatory activity responsible for augmenting of oxidative stress, which may weaken cognitive susceptibility. Recent studies demonstrate that oxidative and nitrative stress may contribute to the pathogenesis of depression through participation in neurogenesis/neuroplasticity, neuroinflammation, and monoamine reuptake process [50, 51]. Recent trend is that antidepressants exert a therapeutic effect by suppressing the production of inflammatory cytokines and ROS/RNS or increasing the antioxidant defense [15]. Therefore, inhibition of inflammation and concomitant oxidative stress may possibly contribute to the improvement of mental and physical efficiency of stroke patients. However, research into the changes and oxidative damage to major biomolecules in depression still lacks in both quality and quantity. Although the depression is one of the many consequences of stroke, it is still poorly diagnosed and inadequately treated [52]. Accordingly, these studies place particular emphasis on the examination of biochemical parameters that may directly affect the improvement of the functionality of patients depending on the severity of their depression.
Brain ischemia initiates a complex cascade of metabolic events, and most of them involve the generation of nitrogen and oxygen free radicals. Oxidative stress, associated with inflammation or postischemic reperfusion, may lead to oxidative and nitrative modifications of protein structure, leading to the alterations in their functional properties, and may cause accumulation of modified protein products, that has been observed under various conditions, such as aging, cell differentiation, and apoptosis [53]. The increase of oxidative stress parameters during a stroke is well documented in the literature. Oxidative stress plays a significant role in the damage associated with ischemic cascade arising from reperfusion, which is restoration of blood flow in the blood vessel after the hypoxia. Oxidative stress which occurs during a stroke significantly affects the irreversible neuronal damage [54–57]. After ischemia-reperfusion ROS may be generated from many sources, including an increased oxidation of catecholamines and hemoglobin, associated with a massive release of iron ions. Furthermore, ROS are produced due to changes in the mitochondrial electron transport, the activation of xanthine oxidoreductase and NAD(P)H oxidase, and during the metabolism of arachidonic acid [58]. As a result of hypoxia, a brain tissue is depleted of oxygen and glucose which are essential products for the ATP synthesis. Thus, it inhibits the activity of sodium-potassium pump and causes an inflow of calcium ions into the cell. Increased concentration of Ca2+ in the cell leads to the activation of Ca2+-dependent enzymes, including proteases and nucleases that are responsible for degradation processes of neurons [59].
Based on the above facts, the present study has been designed to explore the possible relationship between the level of typical markers of oxidative and nitrative protein modifications (carbonyl groups, 3-NT, SH-groups, CAT-activity) and the degree of poststroke depression. Oxidation of amino acid residues leads to the formation of relatively stable carbonyl groups that can be qualitative and quantitative markers enabling the assessment of the oxidative damage of proteins [60, 61]. In our studies, oxidative damage of proteins has been documented as an increased concentration of carbonyl groups in plasma proteins in patients after cerebral ischemia-reperfusion injury (Figure 1). Furthermore, a significant positive correlation between the level of carbonylation and Geriatric Depression Scale (GDS) has also been seen (Figure 5(a)). We have demonstrated more than twofold increase in protein carbonyl groups in poststroke patients as compared to healthy subjects. Our findings are consistent with previous reports. Wang et al. showed a significantly higher increasing the amount of carbonyl groups in the brain tissue of rats with hypoxia-induced brain damage [62]. The link between free radical generation and formation of carbonyl groups in protein molecules in different pathological conditions, including stroke [7], is well documented. However, our findings demonstrate for the first time a significant correlation between the degree of oxidation of the amino acid residues in plasma proteins and the GDS in post stroke patients.
The most important barrier that protects the brain tissue from damage is the blood-brain barrier (BBB). Particular role played the integrity of monolayer of brain capillary endothelial cells. Glutathione is low molecular weight antioxidant that protects endothelial cells from oxidative damage and affects the regeneration process after oxidative damage of cells occurring during a stroke [18]. Moreover, thiol compounds are the first line of defense against ROS and increasing of their oxidation is marker of oxidative stress [63]. Regulation of protein function through thiol-based redox switches plays an important role in maintaining redox homeostasis. Our study showed that concentration of thiol groups in plasma proteins in poststroke patients is approximately 20% lower than in the control group of healthy subjects (Figure 2). Analogous results are obtained by Tsai et al. and Wang et al. who studied the relationship between oxidative stress and the occurrence of other side effects in patients with acute stroke [64, 65]. Moreover, Tsai et al. postulated that the level of free thiol groups was significantly lower in patients with large-vessel disease than in those with small-vessel disease in the acute phase of stroke [64]. This proves that the level of free thiol groups depends on the extent of brain damage caused by oxidative stress, while our studies indicate that the degree of the thiol group oxidation in the plasma proteins is dependent on depression scale in poststroke patients. A significant negative correlation is indicated in Figure 5(b).
Protein nitration process resulting in the formation of 3-nitrotyrosine is mainly caused by strong biological oxidant peroxynitrite (PN) [66]. PN is a potent oxidant and nitrating compound, generated in vivo from the rapid reaction of two relatively less reactive but commonly found of free radicals: nitrogen monoxide (•NO) and superoxide (O2 •−) [8]. Nitric oxide, a water- and lipid-soluble free radical, is generated by the action of nitric oxide synthases. Besides synthesis of nitric oxide, cerebral ischemia leads to the formation of the anion, mainly by the action of nitric oxide synthase, xanthine oxidase, and leakage of mitochondrial electron transport chain. There is increasing evidence that at high concentrations PN is the major compound responsible for ischemia-reperfusion injury or tissue damage by inflammation [67–71]. Ischemic stroke is an acute vascular event that obstructs blood supply to the brain and causes irreversible damage to both neurons and brain vessel cells. Immediately after the stroke, the ischemic tissue produces nitric oxide to restore blood perfusion but also produces superoxide anion. The interaction of these reactive species leads to the formation of peroxynitrite, which irreversibly nitrates protein tyrosines. Recent studies have emphasized the role of peroxynitrite in vascular damages. Increased 3-nitrotyrosine level was observed in various cardiovascular diseases [72]. Vadseth et al. reported the augmented level of nitrated tyrosine residues in plasma proteins derived from patients with coronary artery disease [73]. Bas et al. have demonstrated a significant increase in the concentration of 3-NT positively correlated with elevated activity of metalloproteinases activity (MMPs) in patients after ischemic stroke [74]. Similarly, increased concentration of NT-3 was observed in the studies Coucha et al. who investigated effect of neuronal damage during a stroke on the reactivity of cerebral vessels in rat. They confirmed the hypothesis that the loss of tone of blood vessel was affected by excessive production of peroxynitrite. On this basis, the authors postulated that ischemia-reperfusion injury impairs vessel reactivity via nitration [75]. Moreover, Tajes et al. who studied the effects of PN on the viability of the different brain cells also reported an important role of nitrooxidative stress in the pathogenesis of ischemic stroke [76]. Our studies have shown that the concentration of 3-nitrotyrosine in plasma proteins was approximately twice higher in poststroke patients in comparison with healthy controls (Figure 3). However, unlike in the case of oxidative stress, no relationship was found between protein nitrative damage and Geriatric Depression Scale.
Superoxide anion in addition to the formation of peroxynitrite is involved in a range of toxic mechanisms leading to tissue damage. The production of radicals in the brain is due to catecholamine metabolism such as dopamine and norepinephrine and is increased by the presence of transition metals and by a deficiency of antioxidant agents. Due to the high toxicity of free radicals the organism has developed a number of specialized and efficient methods to deactivate them. The superoxide dismutase enzyme (SOD) catalyzes a dismutation of the superoxide radical into hydrogen peroxide and oxygen. Afterwards hydrogen peroxide is reduced to water and molecular oxygen by peroxidase glutathione and catalase enzymes [77]. Catalase is not directly involved in the elimination of ROS but does not allow for Fenton's reaction to take place in which hydrogen peroxide is converted to highly toxic hydroxyl radicals. Following ischemia there is an increased enzymatic production of ROS, especially superoxide anion and hydrogen peroxide in a reaction catalyzed by xanthine oxidase. During reperfusion molecular oxygen is delivered and xanthine oxidase converts the hypoxanthine to xanthine. In such conditions superoxide anion is released in large quantities. It provides the opportunity for the production of hydrogen peroxide and consequently highly toxic hydroxyl radicals [59]. According to the literature, in stroke, the SOD value does not vary, while the CAT value decreases [77]. Reduced activity of catalase in stroke patients diminishes the effectiveness of deactivation of H2O2 [78], which can freely diffuse across the mitochondrial membrane, and therefore increases oxidative stress. In our study we noticed that that catalase activity in erythrocytes of stroke patients was about 25% lower in comparison with a control group of healthy (Figure 4). The similar effect of reduction in CAT activity in stroke-induced rats was also observed in studies conducted by Safwen et al. [54]. However, our studies have showed that decrease in the activity of catalase is dependent on Geriatric Depression Scale in poststroke patients. A significant negative correlation is indicated in Figure 5(c).
In conclusion, oxidative and nitrative stress parameters have been seen in the present study. However, only oxidative damage of proteins seems to be correlated with the degree of poststroke depression, while nitration of proteins did not show such a relationship. This indicates that the strategies attenuating oxidative stress in poststroke patients can be evaluated for efficacy as novel antidepressants in this population.
Acknowledgment
This study was supported by the Department of General Biochemistry, Faculty of Biology and Environmental Protection, University of Lodz (no. 506/1136).
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.Li I. G., Han H. Y., Lee H. S. Emotional disorder of stroke patients. Journal of Korean Academy of Rehabilitation Medicine. 1995;12:33–38. [Google Scholar]
- 2.Pan A., Sun Q., Okereke O. I., Rexrode K. M., Hu F. B. Depression and risk of stroke morbidity and mortality: a meta-analysis and systematic review. The Journal of the American Medical Association. 2011;306(11):1241–1249. doi: 10.1001/jama.2011.1282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ellis C., Zhao Y., Egede L. E. Depression and increased risk of death in adults with stroke. Journal of Psychosomatic Research. 2010;68(6):545–551. doi: 10.1016/j.jpsychores.2009.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Masskulpan P., Riewthong K., Dajpratham P., Kuptniratsaikul V. Anxiety and depressive symptoms after stroke in 9 rehabilitation centers. Journal of the Medical Association of Thailand. 2008;91(10):1595–1602. [PubMed] [Google Scholar]
- 5.Paolucci S. Epidemiology and treatment of post-stroke depression. Neuropsychiatric Disease and Treatment. 2008;4(1):145–154. doi: 10.2147/ndt.s2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hadidi N., Treat-Jacobson D. J., Lindquist R. Poststroke depression and functional outcome: a critical review of literature. Heart and Lung. 2009;38(2):151–162. doi: 10.1016/j.hrtlng.2008.05.002. [DOI] [PubMed] [Google Scholar]
- 7.Ng F., Berk M., Dean O., Bush A. I. Oxidative stress in psychiatric disorders: evidence base and therapeutic implications. International Journal of Neuropsychopharmacology. 2008;11(6):851–876. doi: 10.1017/s1461145707008401. [DOI] [PubMed] [Google Scholar]
- 8.Radak D., Resanovic I., Isenovic E. R. Link between oxidative stress and acute brain ischemia. Angiology. 2014;65(8):667–676. doi: 10.1177/0003319713506516. [DOI] [PubMed] [Google Scholar]
- 9.Miller E., Walczak A., Saluk J., Ponczek M. B., Majsterek I. Oxidative modification of patient's plasma proteins and its role in pathogenesis of multiple sclerosis. Clinical Biochemistry. 2012;45(1-2):26–30. doi: 10.1016/j.clinbiochem.2011.09.021. [DOI] [PubMed] [Google Scholar]
- 10.Maes M., Galecki P., Chang Y. S., Berk M. A review on the oxidative and nitrosative stress (O&NS) pathways in major depression and their possible contribution to the (neuro)degenerative processes in that illness. Progress in Neuro-Psychopharmacology and Biological Psychiatry. 2011;35(3):676–692. doi: 10.1016/j.pnpbp.2010.05.004. [DOI] [PubMed] [Google Scholar]
- 11.Maes M. An intriguing and hitherto unexplained co-occurrence: depression and chronic fatigue syndrome are manifestations of shared inflammatory, oxidative and nitrosative (IO&NS) pathways. Progress in Neuro-Psychopharmacology and Biological Psychiatry. 2011;35(3):784–794. doi: 10.1016/j.pnpbp.2010.06.023. [DOI] [PubMed] [Google Scholar]
- 12.Suzuki H., Colasanti M. NO: a molecule with two masks of ‘NO’ theatre. BioFactors. 2001;15(2–4):123–125. doi: 10.1002/biof.5520150216. [DOI] [PubMed] [Google Scholar]
- 13.Dhir A., Kulkarni S. K. Nitric oxide and major depression. Nitric Oxide. 2011;24(3):125–131. doi: 10.1016/j.niox.2011.02.002. [DOI] [PubMed] [Google Scholar]
- 14.Gawryluk J. W., Wang J. F., Andreazza A. C., Shao L., Young L. T. Decreased levels of glutathione, the major brain antioxidant, in post-mortem prefrontal cortex from patients with psychiatric disorders. International Journal of Neuropsychopharmacology. 2011;14(1):123–130. doi: 10.1017/s1461145710000805. [DOI] [PubMed] [Google Scholar]
- 15.Lee S.-Y., Lee S.-J., Han C., Patkar A. A., Masand P. S., Pae C.-U. Oxidative/nitrosative stress and antidepressants: targets for novel antidepressants. Progress in Neuro-Psychopharmacology and Biological Psychiatry. 2013;46:224–235. doi: 10.1016/j.pnpbp.2012.09.008. [DOI] [PubMed] [Google Scholar]
- 16.Dröge W. Free radicals in the physiological control of cell function. Physiological Reviews. 2002;82(1):47–95. doi: 10.1152/physrev.00018.2001. [DOI] [PubMed] [Google Scholar]
- 17.Uchida K., Stadtman E. R. Covalent attachment of 4-hydroxynonenal to glyceraldehyde-3-phosphate dehydrogenase. A possible involvement of intra- and intermolecular cross- linking reaction. The Journal of Biological Chemistry. 1993;268(9):6388–6393. [PubMed] [Google Scholar]
- 18.Li W., Busu C., Circu M. L., Aw T. Y. Glutathione in cerebral microvascular endothelial biology and pathobiology: implications for brain homeostasis. International Journal of Cell Biology. 2012;2012:14. doi: 10.1155/2012/434971.434971 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wang Y., Yang J., Yi J. Redox sensing by proteins: oxidative modifications on cysteines and the consequent events. Antioxidants and Redox Signaling. 2012;16(7):649–657. doi: 10.1089/ars.2011.4313. [DOI] [PubMed] [Google Scholar]
- 20.Sun J., Murphy E. Protein S-nitrosylation and cardioprotection. Circulation Research. 2010;106(2):285–296. doi: 10.1161/CIRCRESAHA.109.209452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Manolescu B. N., Berteanu M., Oprea E., et al. Dynamic of oxidative and nitrosative stress markers during the convalescent period of stroke patients undergoing rehabilitation. Annals of Clinical Biochemistry. 2011;48(4):338–343. doi: 10.1258/acb.2011.010243. [DOI] [PubMed] [Google Scholar]
- 22.Reiter C. D., Teng R.-J., Beckman J. S. Superoxide reacts with nitric oxide to nitrate tyrosine at physiological pH via peroxynitrite. The Journal of Biological Chemistry. 2000;275(42):32460–32466. doi: 10.1074/jbc.m910433199. [DOI] [PubMed] [Google Scholar]
- 23.Cojocaru I. M., Cojocaru M., Sapira V., Ionescu A. Evaluation of oxidative stress in patients with acute ischemic stroke. Romanian Journal of Internal Medicine. 2013;51(2):97–106. [PubMed] [Google Scholar]
- 24.Golden T. R., Patel M. Catalytic antioxidants and neurodegeneration. Antioxidants & Redox Signaling. 2009;11(3):555–569. doi: 10.1089/ars.2008.2256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Buss H., Chan T. P., Sluis K. B., Domigan N. M., Winterbourn C. C. Protein carbonyl measurement by a sensitive ELISA method. Free Radical Biology and Medicine. 1997;23(3):361–366. doi: 10.1016/S0891-5849(97)00104-4. [DOI] [PubMed] [Google Scholar]
- 26.Alamdari D. H., Kostidou E., Paletas K., et al. High sensitivity enzyme-linked immunosorbent assay (ELISA) method for measuring protein carbonyl in samples with low amounts of protein. Free Radical Biology and Medicine. 2005;39(10):1362–1367. doi: 10.1016/j.freeradbiomed.2005.06.023. [DOI] [PubMed] [Google Scholar]
- 27.Bijak M., Kolodziejczyk-Czepas J., Ponczek M. B., Saluk J., Nowak P. Protective effects of grape seed extract against oxidative and nitrative damage of plasma proteins. International Journal of Biological Macromolecules. 2012;51(3):183–187. doi: 10.1016/j.ijbiomac.2012.05.009. [DOI] [PubMed] [Google Scholar]
- 28.Koster J. F., Biemond P., Swaak A. J. G. Intracellular and extracellular sulphydryl levels in rheumatoid arthritis. Annals of the Rheumatic Diseases. 1986;45(1):44–46. doi: 10.1136/ard.45.1.44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Khan J., Brennand D. M., Bradley N., Gao B., Bruckdorfer R., Jacobs M. 3-Nitrotyrosine in the proteins of human plasma determined by an ELISA method. Biochemical Journal. 1998;330, part 2:795–801. doi: 10.1042/bj3300795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ischiropoulos H., Al-Mehdi A. B. Peroxynitrite-mediated oxidative protein modifications. FEBS Letters. 1995;364(3):279–282. doi: 10.1016/0014-5793(95)00307-U. [DOI] [PubMed] [Google Scholar]
- 31.Beers R. F., Jr., Sizer I. W. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. The Journal of Biological Chemistry. 1952;195(1):133–140. [PubMed] [Google Scholar]
- 32.Wallace W. R., Dimopoullos G. T. Catalase activity in anaplasma marginale. Journal of Bacteriology. 1965;90:309–311. doi: 10.1128/jb.90.2.309-311.1965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Hauke J., Kossowski T. Comparison of values of pearson's and spearman's correlation coefficients on the same sets of data. Quaestiones Geographicae. 2011;30(2):87–93. doi: 10.2478/v10117-011-0021-1. [DOI] [Google Scholar]
- 34.Whyte E. M., Mulsant B. H. Post stroke depression: epidemiology, pathophysiology, and biological treatment. Biological Psychiatry. 2002;52(3):253–264. doi: 10.1016/s0006-3223(02)01424-5. [DOI] [PubMed] [Google Scholar]
- 35.Kauhanen M.-L., Korpelainen J. T., Hiltunen P., et al. Poststroke depression correlates with cognitive impairment and neurological deficits. Stroke. 1999;30(9):1875–1880. doi: 10.1161/01.str.30.9.1875. [DOI] [PubMed] [Google Scholar]
- 36.Narushima K., Chan K.-L., Kosier J. T., Robinson R. G. Does cognitive recovery after treatment of poststroke depression last? A 2-year follow-up of cognitive function associated with poststroke depression. The American Journal of Psychiatry. 2003;160(6):1157–1162. doi: 10.1176/appi.ajp.160.6.1157. [DOI] [PubMed] [Google Scholar]
- 37.Tiemeier H. Biological risk factors for late life depression. European Journal of Epidemiology. 2003;18(8):745–750. doi: 10.1023/a:1025388203548. [DOI] [PubMed] [Google Scholar]
- 38.Tang W. K., Liang H., Chu W. C. W., Mok V., Ungvari G. S., Wong K. S. Association between high serum total bilirubin and post-stroke depression. Psychiatry and Clinical Neurosciences. 2013;67(4):259–264. doi: 10.1111/pcn.12051. [DOI] [PubMed] [Google Scholar]
- 39.Yang L., Zhang Z., Sun D., Xu Z., Zhang X., Li L. The serum interleukin-18 is a potential marker for development of post-stroke depression. Neurological Research. 2010;32(4):340–346. doi: 10.1179/016164110x12656393665080. [DOI] [PubMed] [Google Scholar]
- 40.Rothenburg L. S., Herrmann N., Swardfager W., et al. The relationship between inflammatory markers and post stroke cognitive impairment. Journal of Geriatric Psychiatry and Neurology. 2010;23(3):199–205. doi: 10.1177/0891988710373598. [DOI] [PubMed] [Google Scholar]
- 41.Gold A. B., Herrmann N., Swardfager W., et al. The relationship between indoleamine 2,3-dioxygenase activity and post-stroke cognitive impairment. Journal of Neuroinflammation. 2011;8, article 17 doi: 10.1186/1742-2094-8-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Huppert J., Closhen D., Croxford A., et al. Cellular mechanisms of IL-17-induced blood-brain barrier disruption. The FASEB Journal. 2010;24(4):1023–1034. doi: 10.1096/fj.09-141978. [DOI] [PubMed] [Google Scholar]
- 43.Swardfager W., Herrmann N., Andreazza A. C., et al. Poststroke neuropsychiatric symptoms: relationships with IL-17 and oxidative stress. BioMed Research International. 2014;2014:6. doi: 10.1155/2014/245210.245210 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Halliwell B. Reactive species and antioxidants. Redox biology is a fundamental theme of aerobic life. Plant Physiology. 2006;141(2):312–322. doi: 10.1104/pp.106.077073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Belmaker R. H., Agam G. Major depressive disorder. The New England Journal of Medicine. 2008;358(1):55–68. doi: 10.1056/nejmra073096. [DOI] [PubMed] [Google Scholar]
- 46.Dowlati Y., Herrmann N., Swardfager W., et al. A meta-analysis of cytokines in major depression. Biological Psychiatry. 2010;67(5):446–457. doi: 10.1016/j.biopsych.2009.09.033. [DOI] [PubMed] [Google Scholar]
- 47.Chung C. P., Schmidt D., Stein C. M., Morrow J. D., Salomon R. M. Increased oxidative stress in patients with depression and its relationship to treatment. Psychiatry Research. 2013;206(2-3):213–216. doi: 10.1016/j.psychres.2012.10.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Vogelgesang A., Dressel A. Immunological consequences of ischemic stroke: Immunosuppression and autoimmunity. Journal of Neuroimmunology. 2011;231(1-2):105–110. doi: 10.1016/j.jneuroim.2010.09.023. [DOI] [PubMed] [Google Scholar]
- 49.Pachner A. R. A Primer of Neuroimmunological Disease. Boston, Mass, USA: Springer; 2012. Neuroimmunology of degenerative diseases and stroke; pp. 155–158. [DOI] [Google Scholar]
- 50.Pae C.-U., Yu H.-S., Kim T.-S., et al. Monocyte chemoattractant protein-1 (MCP1) promoter-2518 polymorphism may confer a susceptibility to major depressive disorder in the Korean population. Psychiatry Research. 2004;127(3):279–281. doi: 10.1016/j.psychres.2004.04.004. [DOI] [PubMed] [Google Scholar]
- 51.Pae C.-U., Marks D. M., Han C., Patkar A. A., Steffens D. Does neurotropin-3 have a therapeutic implication in major depression? International Journal of Neuroscience. 2008;118(11):1515–1522. doi: 10.1080/00207450802174589. [DOI] [PubMed] [Google Scholar]
- 52.Berg A., Lönnqvist J., Palomäki H., Kaste M. Assessment of depression after stroke a comparison of different screening instruments. Stroke. 2009;40(2):523–529. doi: 10.1161/STROKEAHA.108.527705. [DOI] [PubMed] [Google Scholar]
- 53.Stadtman E. R., Levine R. L. Free radical-mediated oxidation of free amino acids and amino acid residues in proteins. Amino Acids. 2003;25(3-4):207–218. doi: 10.1007/s00726-003-0011-2. [DOI] [PubMed] [Google Scholar]
- 54.Safwen K., Selima S., Mohamed E., et al. Protective effect of grape seed and skin extract on cerebral ischemia in rat: implication of transition metals. International Journal of Stroke. 2014 doi: 10.1111/ijs.12391. [DOI] [PubMed] [Google Scholar]
- 55.Chen H.-Z., Guo S., Li Z.-Z., et al. A critical role for interferon regulatory factor 9 in cerebral ischemic stroke. Journal of Neuroscience. 2014;34(36):11897–11912. doi: 10.1523/jneurosci.1545-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Abd-Elsameea A. A., Moustaf A. A., Mohamed A. M. Modulation of the oxidative stress by metformin in the cerebrum of rats exposed to global cerebral ischemia and ischemia/reperfusion. European Review for Medical and Pharmacological Sciences. 2014;18(16):2387–2392. [PubMed] [Google Scholar]
- 57.Lee J.-C., Won M.-H. Neuroprotection of antioxidant enzymes against transient global cerebral ischemia in gerbils. Anatomy & Cell Biology. 2014;47(3):149–156. doi: 10.5115/acb.2014.47.3.149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Domański L., Dołegowska B., Safranow K., et al. Early phase of reperfusion of human kidney allograft does not affect an erythrocyte anti-oxidative system. Nephrology. 2006;11(5):467–470. doi: 10.1111/j.1440-1797.2006.00672.x. [DOI] [PubMed] [Google Scholar]
- 59.Broughton B. R. S., Reutens D. C., Sobey C. G. Apoptotic mechanisms after cerebral ischemia. Stroke. 2009;40(5):e331–e339. doi: 10.1161/STROKEAHA.108.531632. [DOI] [PubMed] [Google Scholar]
- 60.Dalle-Donne I., Rossi R., Giustarini D., Milzani A., Colombo R. Protein carbonyl groups as biomarkers of oxidative stress. Clinica Chimica Acta. 2003;329(1-2):23–38. doi: 10.1016/S0009-8981(03)00003-2. [DOI] [PubMed] [Google Scholar]
- 61.Dalle-Donne I., Giustarini D., Colombo R., Rossi R., Milzani A. Protein carbonylation in human diseases. Trends in Molecular Medicine. 2003;9(4):169–176. doi: 10.1016/S1471-4914(03)00031-5. [DOI] [PubMed] [Google Scholar]
- 62.Wang Z., Liu T., Gan L., et al. Shikonin protects mouse brain against cerebral ischemia/reperfusion injury through its antioxidant activity. European Journal of Pharmacology. 2010;643(2-3):211–217. doi: 10.1016/j.ejphar.2010.06.027. [DOI] [PubMed] [Google Scholar]
- 63.Groitl B., Jakob U. Thiol-based redox switches. Biochimica et Biophysica Acta—Proteins and Proteomics. 2014;1844(8):1335–1343. doi: 10.1016/j.bbapap.2014.03.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Tsai N.-W., Chang Y.-T., Huang C.-R., et al. Association between oxidative stress and outcome in different subtypes of acute ischemic stroke. BioMed Research International. 2014;2014:7. doi: 10.1155/2014/256879.256879 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Wang F., Liang W., Lei C., et al. Combination of HBO and memantine in focal cerebral ischemia: is there a synergistic effect? Molecular Neurobiology. 2014 doi: 10.1007/s12035-014-8949-5. [DOI] [PubMed] [Google Scholar]
- 66.Bijak M., Ponczek M., Saluk J., Chabielska M., Stepniak J., Nowak P. Peroxynitrite a strong biological oxidant. Chemical News. 2012;66 [Google Scholar]
- 67.Kooy N. W., Royall J. A., Ye Y. Z., Kelly D. R., Beckman J. S. Evidence for in vivo peroxynitrite production in human acute lung injury. American Journal of Respiratory and Critical Care Medicine. 1995;151(4):1250–1254. doi: 10.1164/ajrccm/151.4.1250. [DOI] [PubMed] [Google Scholar]
- 68.Fukuyama N., Takebayashi Y., Hida M., Ishida H., Ichimori K., Nakazawa H. Clinical evidence of peroxynitrite formation in chronic renal failure patients with septic shock. Free Radical Biology and Medicine. 1997;22(5):771–774. doi: 10.1016/S0891-5849(96)00401-7. [DOI] [PubMed] [Google Scholar]
- 69.Hayashi Y., Sawa Y., Nishimura M., et al. Peroxynitrite, a product between nitric oxide and superoxide anion, plays a cytotoxic role in the development of post-bypass systemic inflammatory response. European Journal of Cardio-Thoracic Surgery. 2004;26(2):276–280. doi: 10.1016/j.ejcts.2004.03.033. [DOI] [PubMed] [Google Scholar]
- 70.Salvemini D., Cuzzocrea S. Oxidative stress in septic shock and disseminated intravascular coagulation. Free Radical Biology and Medicine. 2002;33(9):1173–1185. doi: 10.1016/S0891-5849(02)00961-9. [DOI] [PubMed] [Google Scholar]
- 71.Szabó C. Multiple pathways of peroxynitrite cytotoxicity. Toxicology Letters. 2003;140-141:105–112. doi: 10.1016/S0378-4274(02)00507-6. [DOI] [PubMed] [Google Scholar]
- 72.Hoffman M. Alterations of fibrinogen structure in human disease. Cardiovascular and Hematological Agents in Medicinal Chemistry. 2008;6(3):206–211. doi: 10.2174/187152508784871981. [DOI] [PubMed] [Google Scholar]
- 73.Vadseth C., Souza J. M., Thomson L., et al. Pro-thrombotic state induced by post-translational modification of fibrinogen by reactive nitrogen species. The Journal of Biological Chemistry. 2004;279(10):8820–8826. doi: 10.1074/jbc.m306101200. [DOI] [PubMed] [Google Scholar]
- 74.Bas D. F., Topcuoglu M. A., Gursoy-Ozdemir Y., Saatci I., Bodur E., Dalkara T. Plasma 3-nitrotyrosine estimates the reperfusion-induced cerebrovascular stress, whereas matrix metalloproteinases mainly reflect plasma activity: a study in patients treated with thrombolysis or endovascular recanalization. Journal of Neurochemistry. 2012;123(2):138–147. doi: 10.1111/j.1471-4159.2012.07952.x. [DOI] [PubMed] [Google Scholar]
- 75.Coucha M., Li W., Johnson M. H., Fagan S. C., Ergul A. Protein nitration impairs the myogenic tone of rat middle cerebral arteries in both ischemic and nonischemic hemispheres after ischemic stroke. The American Journal of Physiology—Heart and Circulatory Physiology. 2013;305(12):H1726–H1735. doi: 10.1152/ajpheart.00535.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Tajes M., Ill-Raga G., Palomer E., et al. Nitro-oxidative stress after neuronal ischemia induces protein nitrotyrosination and cell death. Oxidative Medicine and Cellular Longevity. 2013;2013:9. doi: 10.1155/2013/826143.826143 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Venarucci D., Venarucci V., Vallese A., et al. Free radicals: important cause of pathologies refer to ageing. Panminerva Medica. 1999;41:335–339. [PubMed] [Google Scholar]
- 78.Miller E., Mrowicka M., Malinowska K., Mrowicki J., Saluk-Juszczak J., Kedziora J. Effects of whole-body cryotherapy on a total antioxidative status and activities of antioxidative enzymes in blood of depressive multiple sclerosis patients. World Journal of Biological Psychiatry. 2011;12(3):223–227. doi: 10.3109/15622975.2010.518626. [DOI] [PubMed] [Google Scholar]