

Abstract
The transcriptional repressors Hey1 and Hey2 are primary target genes of Notch signaling in the cardiovascular system and induction of Hey gene expression is often interpreted as activation of Notch signaling. Here we report that treatment of primary human endothelial cells with serum or fresh growth medium led to a strong wave of Hey1 and Hey2 transcription lasting for approximately three hours. Transcription of other Notch target genes (Hes1, Hes5, ephrinB2, Dll4) was however not induced by serum in endothelial cells. Gamma secretase inhibition or expression of dominant-negative MAML1 did not prevent the induction of Hey genes indicating that canonical Notch signaling is dispensable. Pretreatment with soluble BMP receptor Alk1, but not Alk3, abolished Hey gene induction by serum. Consequently, the Alk1 ligand BMP9 stimulated Hey gene induction in endothelial cells. Several other cell types however did not show such a strong BMP signaling and consequently only a very mild induction of Hey genes. Taken together, the experiments revealed that bone morphogenic proteins within the serum of cell culture medium are potent inducers of endothelial Hey1 and Hey2 gene expression within the first few hours after medium change.
Introduction
Hes and Hey genes are the mammalian orthologes of hairy and enhancer-of-split genes in Drosophila.[1] They encode basic helix-loop-helix (bHLH) transcription factors that control expression of numerous loci in the human genome.[2] Both gene families are well known as the primary effector molecules of Notch signaling,[3] and both act predominantly as transcriptional repressors.[4]
The Delta-Notch cascade is a highly conserved and highly versatile signaling system that directs a multitude of binary and inductive cell fate decisions during development. Notch receptors (Notch1-Notch4) become activated upon binding of Delta-like or Jagged ligands (Jag1, Jag2, Dll1, Dll3, and Dll4) presented by adjacent cells. This leads to cleavage of the receptor and release of the Notch intracellular domain (NICD), which translocates to the nucleus. NICD interacts with Rbp-Jk (also known as CBF1 or Su(H)) that is bound to DNA and recruits MAML1 and transcriptional activators. Thereby, the Rbp-Jk complex is turned from a repressor into an activator of transcription.[3] This leads to the induction of a plethora of genes depending on the cell type and differentiation state.[4]
The most prominent Notch target genes are the Hes (Hes1-Hes7) and Hey (Hey1, Hey2, HeyL) transcription factors. At least one or two of these genes are activated by NICD in almost any cell type. Mouse knockout studies revealed that Hes genes execute Notch signals predominantly in the nervous, hematopoietic, endocrine and immune systems, while Hey genes are critically important in the cardiovascular system.[5]
Notch signaling is one of the few central regulators that guide almost every differentiation step during development, stem cell differentiation or regeneration.[6] Moreover, dysregulated Notch signaling occurs in several inherited syndromes as well as in cancer.[7, 8] Therefore, there is a great need to measure Notch signaling activity in cell culture and laboratory animals. This is not a trivial task. One well-accepted method is the detection of NICD by Western blotting,[9] which is technically challenging in cells with low Notch expression. Therefore, the indirect measurement of Notch target gene expression is most commonly used.
Expression of Hey1 and Hey2 in endothelial cells is dependent on Notch1 but both genes are still expressed in Notch1 -/- mouse tissue, although at a lower level.[10] This indicates that other signaling pathways are involved in Hey gene expression, and indeed there is a complex crosstalk between Notch, TGF-beta and BMP signaling in endothelial cells, which allows a coordinated expression of Hey genes during angiogenesis.[11] Thereby, BMP signaling via Smad1/5 synergizes with Rbp-Jk to induce Hey1 and Hey2 expression.[12–15] Recent studies indicated that oscillatory expression of BMP-Smad and Notch target genes occurs in endothelial cells in a nonsynchronized manner. Thereby, individual endothelial cells become periodically competent to adopt either the tip cell or the stalk cell phenotype during angiogenesis.[13] Tip cells can guide new vessel sprouts whereas stalk cells follow and generate the lumen. However, this is not a static state and stalk cells actively take over the position and identity of a tip cell.[16] This behavior may be due to the oscillatory Notch and BMP-Smad1/5 loops in endothelial cells.[13] Hes and Hey gene expression occurs in an oscillatory manner in the presomitic and in neuronal progenitors.[17, 18] Likewise, Hes1 expression was reported to oscillate upon stimulation with serum in several cultured cell types such as myoblastic and fibroblastic cell lines.[19] Interestingly, this process seems to be dependent of Stat and Smad signaling pathways.[20]
This study was inspired by the serendipitous observation that changing growth medium causes immense induction of Hey1 and Hey2 gene transcription in endothelial cells. The data demonstrate that at least in endothelial cells careful analysis of additional parameters is necessary to conclude that a certain treatment causes Notch signaling activation.
Materials and Methods
Plasmids, chemicals
cDNA encoding amino acids 13 to 74 of MAML1,[21] which encodes a dominant-negative form (provided by Jon C. Astor, Harvard) was cloned in frame with mCherry cDNA into pENTR3c and shuttled to pAd/CMV/V5 by Gateway cloning (LifeTechnologies). Adenoviral vectors were produced in HEK293 cells, and used at a MOI of 50. The gamma secretase inhibitor DAPT (N-[(3,5-difluorophenyl)acetyl]-L-alanyl-2-phenyl]glycine-1,1-dimethylethylester; Calbiochem) was used at 25 μM. The BMP type I receptor inhibitor LDN193189 (System Biosciences) was used at 3 μM. Cells were pretreated with DAPT or LDN193189 over night or for 30 minutes respectively. Recombinant human BMP9 and TGFβ1 were purchased from R&D systems and used at 10 ng/ml. Recombinant human Alk1 and Alk3 fused to human Fc were purchased from R&D systems and medium was preincubated for 30 minutes at a concentration of 5 μg/ml before adding to the cells.
RNA isolation and qPCR analysis, Western blotting
RNA was purified with the RNeasy Mini Kit (Quiagen) and transcribed into cDNA (High Capacity cDNA Reverse Transcription Kit; Life Technologies). Real-time PCR was performed using the STEPOnePlus real-Time PCR system (Applied Biosystems). RPS29 served as a house-keeping gene for normalization.
Protein lysates were subjected to SDS-PAGE blotted to nitrocellulose and incubated with antibodies against cleaved Notch1 (Abcam ab27526), pSmad1/5 (Ser463/465; Cell Signaling 9516), VCP (Abcam ab11433) or β-tubulin (Santa Cruz sc 9104) at 4°C overnight. After washing with TBST and incubation with peroxidase-coupled secondary antibodies, bands were detected with a chemiluminescence system (BioRad).
Cell culture
HUVEC and HUAEC were grown and maintained until passage 5 in Endopan3 Growth Medium containing 3% FCS and supplements (Pan-Biotech). HBMVEC were maintained in BMEC growth media (PELO Biotech). HUASMC were cultured in high-glucose DMEM (Gibco) containing 15% FCS. HEK293, A549, and HeLa cells were cultured in DMEM with 10% FCS. FCS was from BioChrom and Sigma Aldrich, horse serum from BioChrom. HUVEC, HUAEC, HUASMC were freshly isolated, HBMVEC were purchased from PELO Biotech, Hela, A549 and HEK293T cells (ATCC numbers CCL-2, CCL-185, CRL-3216) were taken from the laboratory stock.
Promoter analysis
Genomic sequences were obtained from NCBI, exons and translational start sites from ensembl.org. “rVISTA” was used with standard settings to find the Smad1 binding motif GCCGnCGC and to assess its conservation between different species.
Statistical analysis
The results are presented as means + SD. Student’s t-test was used for pairwise comparisons between groups. p<0.05 was considered as statistically significant.
Results
Serum induces the expression of Hey1 and Hey2 in endothelial cells
Induction of Hey1 and Hey2 transcription is often considered as readout for activation of Notch signaling in endothelial cells. We treated primary human umbilical venous endothelial cells (HUVEC) with different nutrients and metabolic compounds for two hours to screen for potential effects on Notch signaling. Surprisingly, this experiment suggested that all substances would largely induce Hey1 and Hey2 transcription. More careful analysis revealed however that the induction of Hey1 and Hey2 expression was due to the exchange of the endothelial growth medium only. The addition of fresh endothelial growth medium (Endopan3 with 3% FCS and supplements) after 24 hours culture led to a more than 10-fold induction of Hey1 and Hey2 mRNA expression in HUVEC. This induction could be detected after one hour, reached a maximum between 1.5 and 2 hours and lasted until 3 to 4 hours after the addition of fresh medium (Fig. 1A).
Fig 1. Serum induces transcription of Hey1 and Hey2.

A) HUVEC were grown in full growth medium and medium was changed to fresh growth medium at t0. The chart depicts the mRNA levels of Hey1 and Hey2. B) HUVEC were grown in full growth medium and medium was changed either to fresh growth medium (GM), basal medium (BM) supplemented with 3% FCS or BM supplemented with a mix of growth factors (supplements). Transcript levels of Hey1 and Hey2 were determined after 1.5h and are shown in relation to samples with no medium change (untreated). C) HUVEC were grown in full growth medium and medium was changed to basal medium supplemented with 3% FCS purchased from two different suppliers, charcoal filtered FCS, horse serum or basal medium complemented with 3% or 10% human serum. Transcript levels of Hey1 and Hey2 are shown 1.5h after medium change. n = 3 (a,b) or one experiment shown (c). Mean and standard deviation of biological (a,b) or technical (c) replicates. Significant differences were calculated between the conditions marked with asterisks and t0 (a) or untreated control respectively.* p<0.05, ** p<0.01, *** p<0.001.
VEGF enhances Hey gene expression through induction of Dll4-Notch signaling.[22] VEGF is together with EGF, FGF, IGF, ascorbic acid, hydrocortisone and heparin a component of the endothelial growth medium supplement. However, VEGF alone or in combination with the other supplements was not responsible for the induction of Hey gene expression (Fig. 1B). Addition of basal medium (lacking FCS and supplements) did not alter Hey gene expression. However, the addition of 3% FCS led to almost the same induction of Hey gene transcription as the complete growth medium (Fig. 1B). The filtering of FCS with charcoal, which reduces the amount of lipophilic substances, did not significantly decrease the induction of Hey1 and Hey2 compared to non-filtered FCS (Fig. 1C). The importance of serum was further verified by treating HUVEC with other FCS preparations, as well as horse serum and serum from healthy adult human volunteers (Fig. 1C). As such, a hydrophilic serum compound was responsible for a short-term induction of Hey gene expression upon medium exchange.
Hey genes encode for transcriptional repressors, which orchestrate the expression of many other genes. In endothelial cells, VEGFR2 and VEGF are important established target genes of Hey proteins. Given the time for Hey protein synthesis, we tested the expression of these genes four hours after adding serum to HUVEC. At this time point, mRNA levels of VEGFR2 and VEGF were significantly down regulated to 67±13% and 75±11% respectively (n = 3; p < 0.05). This indicates that serum does not only induce Hey genes but indeed affects the regulatory network downstream of these transcriptional repressors as well.
Serum-induced Hey1 gene expression is independent of Notch signaling
Hey genes are regarded as the primary Notch target genes in endothelial cells.[10] Therefore, we tested if inhibition of Notch signaling activity would abolish the induction of Hey1 and Hey2 expression upon treatment with serum. HUVEC were pretreated with the gamma-secretase inhibitor DAPT overnight to inhibit Notch receptor cleavage and were then stimulated with growth medium or 3% FCS. DAPT treatment significantly decreased basal Hey1 and Hey2 mRNA levels. However, the addition of growth medium or 3% FCS still led to a considerable induction of Hey gene expression irrespectively if DAPT was present or not (Fig. 2A). This indicated that induction of Hey gene expression was independent of Notch receptor cleavage.
Fig 2. Induction of Hey genes by serum does not depend on Notch signaling.

A) HUVEC were grown in growth medium containing 25μM DAPT or an equivalent volume of DMSO. Medium was unchanged (DMSO, DAPT) or changed to fresh growth medium (GM) or basal medium (BM) with 3% FCS containing DAPT or DMSO respectively. Transcript levels of Hey1 and Hey2 were measured 1.5h after medium change and are shown in relation to the sample incubated with DMSO, whose medium was not changed. B) HUVEC were transduced with adenoviral vectors expressing GFP or dnMAML. Cells were grown in growth medium. The medium was either unchanged (GFP, dnMAML) or replaced with fresh growth medium or basal medium containing 3% FCS. Transcript levels were determined after 1.5h and are normalized to the cells transduced with GFP and unchanged medium. C) HUAEC were grown in growth medium (untreated) or the medium was substituted with fresh growth medium or basal medium containing 3% FCS. The western blot is showing the cleaved intracellular domain of Notch1 (NICD) and β-tubulin 30 minutes after medium change. The chart summarizes four independent experiments and depicts the relative levels of NICD normalized for β-tubulin. D) HUVEC were left untreated or the medium was changed to fresh growth medium or basal medium supplemented with 3% FCS. After incubating 1.5h, the cells were harvested and the mRNA levels of several established Notch target genes (Hey1, Hey2, Hes1, Hes5, Dll4 EFNB2) were determined. Fold changes and significances were calculated to the untreated control. Mean and standard deviation, n = 3, * p<0.05, ** p<0.01, *** p<0.001, n.s. not significant.
We repeated this experiment with a different kind of Notch inhibitor. Dominant negative Mastermind-like 1 protein (dnMAML) is a well-established tool to interfere with the nuclear Notch protein complex at the promoter of target genes.[21] dnMAML1 forms an inactive complex with NICD to inhibit the transcription of direct Notch target genes like Hey1 and Hey2 without affecting Rbp-Jk repressor function. Adenoviral transduction of HUVEC with dnMAML reduced the amount of Hey1 and Hey2 mRNA. However, dnMAML1 could not prevent the induction of Hey2 transcription by serum, whereas the induction of Hey1 expression was inhibited but not prevented by dnMAML1 (Fig. 2B).
The gold standard to detect Notch activity in endothelial cells is to detect the levels of cleaved and thus active Notch1 (NICD). The levels of NICD and Dll4 in HUVEC are low and difficult to detect with antibodies.[23] Therefore, we employed arterial endothelial cells (HUAEC), which have higher NICD levels. Hey1 and Hey2 expression was induced in these cells to a similar extent as in HUVEC (see below, and Fig. 3D). The addition of growth medium or 3% FCS did however only slightly increase the rate of Notch1 cleavage by approximately 15% (Fig. 2C).
Fig 3. Transcriptional activation of Hey1 and Hey2 by serum is caused by BMP-Smad-signaling.
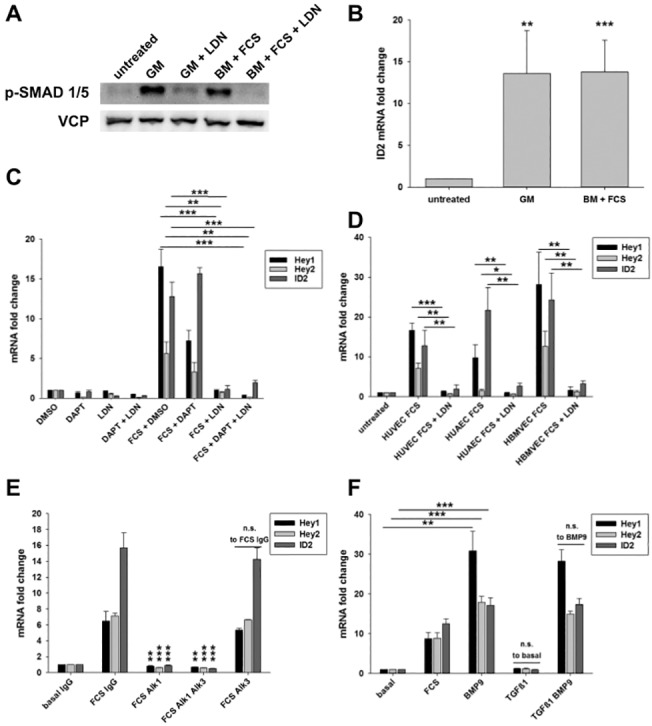
A) HUVEC were cultured in growth medium and were pretreated for 30 min with 3μM LDN193189 or a corresponding volume of DMSO. The medium was left on the cells or it was replaced with fresh growth medium (GM) or basal medium (BM) with 3% FCS, both of which were containing LDN193189 or DMSO respectively. The western blot is showing phosphorylated Smad 1/5 (Ser463/465) and VCP in cells lysed 1.5h after medium change. B) HUVEC were cultured in growth medium, which was replaced with fresh growth medium or basal medium containing 3% FCS. Relative ID2 mRNA levels are shown 1.5 h after medium change normalized to samples not subjected to medium change (untreated). C) HUVEC were cultured in growth medium and were pretreated over night with 25μM DAPT, 30 min with 3μM LDN193189 or the corresponding volumes of DMSO. Medium was not replaced (DMSO, DAPT, LDN, DAPT + LDN) or changed to basal medium with 3% FCS complemented with the inhibitors or DMSO respectively. The transcript levels of Hey1, Hey2 and ID2 were measured 1.5h after changing the medium. D) HUVEC, HUAEC and HBMVEC were cultured in growth medium and were pretreated 30 min with 3μM LDN193189 or DMSO. The medium remained unchanged (untreated) or was replaced with basal medium with 3% FCS containing LDN193189 or DMSO. The transcript levels of Hey1, Hey2 and ID2 were measured 1.5h after changing the medium. E) HUVEC were treated with medium containing no or 3% FCS and either soluble Alk1-Fc, Alk3-Fc or control IgG (5 μg/ml) for 1.5h. Transcript levels of Hey1,Hey2 or Id2 and significances are calculated to the basal medium IgG control. F) Cells were treated with basal medium containing 3% FCS, 10 ng/ml recombinant human BMP9 or 10 ng/ml recombinant human TGFβ1 for 1.5h. Transcript levels of Hey1, Hey2 and Id2 are calculated to the sample treated with basal medium. Mean and standard deviation, n≥3, * p<0.05, ** p<0.01, *** p<0.001.
To finally clarify this issue we analyzed the expression of other “classical” Notch target genes in endothelial cells upon treatment with growth medium or 3% FCS. Hes1 expression was unaltered, while Hes5, Dll4 and ephrinB2 were only very slightly changed (Fig. 2D). This regulation fits quite well to the slightly increased cleavage rate of Notch1. However, the exaggerated Hey1 and Hey2 expression rates must be due to activation of another signaling pathway.
BMP-Smad1/5 induces Hey1 and Hey2 gene expression in endothelial cells
It was shown that bone morphogenic protein signaling via Smad1/5/8 synergizes with Notch signaling to induce Hey1 and Hey2 transcription in endothelial cells,[11] and that BMP9 and BMP10 stimulate expression of Notch target genes.[14, 15] Since various BMPs are components of FCS,[15, 24] we tested BMP-Smad1/5 signaling as the potential inducer of Hey1 and Hey2 expression. Indeed, treatment of HUVEC with growth medium or 3% FCS led to a strong activation of Smad1/5 as determined by Western blotting with phospho-specific antibodies (Fig. 3A). The prototypical Smad1/5 target gene Id2 was also induced more than 15-fold one and a half hours after treatment (Fig. 3B).
Dorsomorphin and LDN19318, a derivative of it, selectively inhibit BMP type I receptors such as Alk1, Alk2 and Alk3 with high affinity. [25, 26] LDN19318 abolished Smad1/5 phosphorylation upon treatment with serum (Fig. 3A). We pretreated HUVEC for 30 minutes with LDN193189 before the addition of fresh growth medium or 3% FCS. The inhibition of BMP-Smad1/5 signaling abrogated any induction of Hey1, Hey2 or Id2 (Fig. 3C). The same behavior was also observed in HUAEC and primary human microvascular brain endothelial cells (HBMVEC, Fig. 3D). This indicates that the induction of Hey gene expression in different primary endothelial cell cultures is mediated via BMP type I receptor signaling.
To further clarify, which of the BMP type I receptors is involved in the induction of Hey-genes by serum, we employed soluble, recombinant human Alk1 and Alk3 receptors fused to Fc. Blocking Alk3 signaling did not inhibit the induction of Hey-genes and Id2 by serum, whereas blocking Alk1 completely reversed these effects (Fig. 3E). Treating HUVEC with recombinant human BMP9, which is an activator of Alk1, did elicit transcription of Hey1, Hey2 and Id2 similar to serum (Fig. 3F) further confirming the role of Alk1. Alk1 can also be activated by TGFβ.[27] However, when treating HUVEC with recombinant human TGFβ1, there was no induction of Hey1 and Hey2 after 1.5 hours. Likewise, TGFβ1 could not further enhance the induction of Hey genes by BMP9 (Fig. 3F). These experiments show BMPs such as BMP9, which are components of serum, signal through Alk1 to induce transcription of Hey1 and Hey2.
Smad1/5 can activate the transcription of Hey1 by binding to a cluster of GCCGnCGC motifs located within 1 kb upstream of the first exon.[28] Hey1 and Hey2 have redundant roles in endothelial cells and have comparably similar promoter regions. We analyzed the Hey2 and Hey1 promoters for such Smad1-binding motifs in silico (Fig. 4) and found two of these motifs in the proximal promoter of human Hey2 (within 200bp upstream of the translational start site). These sites were conserved between several mammalian species, which indicates that these motifs have indeed a regulatory function and might be implicated in the induction of Hey2 transcription by Smad signaling.
Fig 4. Smad1 binding sites in the promoter regions of Hey1 and Hey2.

Schematic alignment of the Hey1 and Hey2 promoters of Homo sapiens (human), Pan troglodytes (chimpanzee), Bos Taurus (cow) and Rattus norvegicus (rat). Potential Smad1 binding sites (GCCGnCGC) are depicted in red. For a clearer visualization, recognition sites, which are separated by less than 10 bp, are only depicted by one red bar.
Lastly, we tested other cell types in the same experimental settings as described for endothelial cells. However, we could not observe such an induction of Hey1, Hey2 or Id2 in primary human vascular smooth muscle cells from the umbilical artery, HEK293, HeLa or A549 adenocarcinoma cells (Fig. 5).This indicates that endothelial cells react much stronger to BMPs in the serum than other cell types and cell lines.
Fig 5. Marginal activation of Hey1, Hey2 and ID2 in non-endothelial cells.
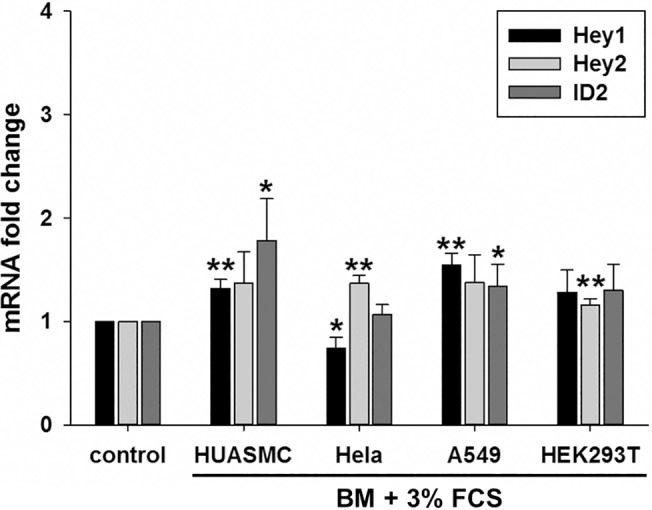
HUASMC, Hela, A549 and HEK293T cells were cultured in their standard medium, before they were left untreated or the medium was changed to basal medium containing 3% FCS. The transcript levels of Hey1, Hey2 and ID2 were determined 1.5h after changing the medium and were normalized to the samples, whose medium was not changed. Mean and standard deviation, n = 3, * p<0.05, ** p<0.01.
Taken together, the experiments revealed that bone morphogenic proteins such as BMP9 act through Alk1 on endothelial cells to stronglyinduce the expression of Hey1 and Hey2 genes in the first few hours upon medium exchange. Given that Hey gene induction is frequently used as readout for Notch activity, this should be carefully taken into account.
Discussion
Induction of Hey gene transcription has been interpreted as activation of Notch signaling in hundreds of published studies. This work however suggests that careful analysis is necessary to draw such conclusions. We found that in particular endothelial cells react with a single wave of strong Hey1 and Hey2 expression to fresh serum. This wave lasted for three to four hours and was not dependent on Notch receptor cleavage but on Alk1 signaling. Therefore, this study revealed that bone morphogenic proteins (BMP) like BMP9, which are present in fetal serum, induce the expression of Hey genes, which play critical roles for endothelial cell biology.
The analysis of Notch signaling is complicated by the fact that four receptors and several ligands are present in the human genome.[3] One of the most reliable methods is the detection of cleaved Notch receptors and quantification of the intracellular domain (NICD).[9] However, this method relies on Western blotting and requires a substantial amount of cells. With immunocytochemistry one can detect nuclear NICD as an indicator of active Notch, however small differences between groups are difficult to measure. Alternatively, one can use reporter assays based on Rbp-Jk responsive DNA elements [29]. These have however the disadvantage that primary cells like HUVEC cannot be transfected in a sufficient degree. Therefore, the most convenient way to measure Notch activity in primary cells is the quantification of “Notch target genes”.
There is no doubt that Hes and Hey transcription factors are the primary target genes of Notch. However, one needs to consider that other signaling pathways may also control their expression. At least for Hes1 there is evidence that hedgehog, FGF, Wnt, and STAT signaling co-regulate transcription of the Hes1 gene or the activity of the Hes1 protein.[11, 20, 30] In endothelial cells BMP-Smad cooperates with NICD to potentiate Hes1 and Hey1 expression.[12–15, 31] Our experiments showed that the gamma-secretase inhibitor DAPT which blocks Notch receptor cleavage was unable to interfere with serum-induced Hey gene induction in endothelial cells, whereas dominant-negative MAML1 (dnMAMAL1) inhibited the induction of Hey1. This may be due to the fact that BMP-Smad1/5 signaling acts through Rbp-Jk at the promoter elements of Hes and Hey genes.[12, 13, 32] dnMAML1 interferes with the Notch/Rbp-Jk complex,[21] and this may also disturb the action of Smad1/5 on Rbp-Jk. Taken together, the presented data suggest that the analysis of Notch activation in endothelial cells should be evaluated by the additional analysis of other Notch targets like ephrinB2 and Dll4, which were not induced by serum,[23, 33] and if possible Western blotting against NICD.
This study revealed that Hey gene induction was dependent of Alk1 and the effects of serum could be phenocopied by supplying recombinant BMP9 to the cells. This protein activates Alk1 and is present in serum at comparably high concentrations.[15] Consistently, BMP9 was previously reported to induce Hey1 expression in mesenchymal stem cells.[34]
Hey1 and Hey2 gene induction lasted only for about three hours after serum treatment. This may be due to the short half-life of bone morphogenic proteins, but also due to the fact that Hey1 and Hey2 proteins repress the expression of their own gene promoter.[34] Kagayama and colleagues have demonstrated nonsynchronized oscillatory gene expression of Hes1 in several cell types.[18–20, 35] Synchronized cycling of Hes and Hey gene expression was also detected in the presomitic mesoderm in mice and zebrafish and this oscillatory gene expression is critical for the formation of somites.[30] In cell culture one can synchronize the cells by starvation followed by serum treatment. This leads to a three to four-fold gene induction of Hes1 expression within 1–2 hours.[19] Fig. 5 shows that we observed such a weak wave of Hey1 and Hey2 gene expression in non-endothelial cells. However the induction by Alk1 signaling in endothelial cells was considerably stronger and thus probably masked this effect. Taken together, in the tested endothelial cells serum induces one large wave of Hey expression, whereas in other cell types serum seems to elicit a much weaker but oscillatory induction.[19, 20] The reason for this difference is unknown but could involve differential expression of BMP receptors or components of Smad signaling. For instance, the expression of Alk1, the receptor triggering the Hey induction described in this report, is mainly limited to endothelial cells.[36] The absence of Alk1 in non-endothelial cells would explain why serum fails to induce Hey expression in these systems.
We do not know if the presented findings have any implication for endothelial cells within an intact blood vessel. Nevertheless, BMP4, -9, and -10 circulate in high concentrations in serum,[15, 24] while BMP2, -4, -6, and -7 can be produced by injured or hypoxic tissue to promote angiogenesis.[11] As such we speculate that in situations like reperfusion of occluded vessels the contact with blood serum may stimulate a wave of Hey gene expression, which could have consequences for endothelial cell behavior, e.g. impaired sprouting angiogenesis.
Acknowledgments
We thank all members of the Fischer laboratory for critical discussion.
Data Availability
All relevant data are within the paper.
Funding Statement
This work was supported by grants from the Deutsche Forschungsgemeinschaft (SFB-TR23), the Deutsche Krebshilfe and the Helmholtz Association. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Leimeister C, Externbrink A, Klamt B, Gessler M. Hey genes: a novel subfamily of hairy- and Enhancer of split related genes specifically expressed during mouse embryogenesis. Mechanisms of development. 1999;85(1–2):173–7. [DOI] [PubMed] [Google Scholar]
- 2. Heisig J, Weber D, Englberger E, Winkler A, Kneitz S, Sung WK, et al. Target gene analysis by microarrays and chromatin immunoprecipitation identifies HEY proteins as highly redundant bHLH repressors. PLoS genetics. 2012;8(5):e1002728 10.1371/journal.pgen.1002728 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Kopan R, Ilagan MX. The canonical Notch signaling pathway: unfolding the activation mechanism. Cell. 2009;137(2):216–33. 10.1016/j.cell.2009.03.045 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Fischer A, Gessler M. Delta-Notch—and then? Protein interactions and proposed modes of repression by Hes and Hey bHLH factors. Nucleic Acids Res. 2007;35(14):4583–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Wiese C, Heisig J, Gessler M. Hey bHLH factors in cardiovascular development. Pediatric cardiology. 2010;31(3):363–70. 10.1007/s00246-009-9609-9 [DOI] [PubMed] [Google Scholar]
- 6. Guruharsha KG, Kankel MW, Artavanis-Tsakonas S. The Notch signalling system: recent insights into the complexity of a conserved pathway. Nature reviews Genetics. 2012;13(9):654–66. 10.1038/nrg3272 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Ntziachristos P, Lim JS, Sage J, Aifantis I. From fly wings to targeted cancer therapies: a centennial for notch signaling. Cancer Cell. 2014;25(3):318–34. 10.1016/j.ccr.2014.02.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Penton AL, Leonard LD, Spinner NB. Notch signaling in human development and disease. Seminars in cell & developmental biology. 2012;23(4):450–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Nakajima M, Shimizu T, Shirasawa T. Notch-1 activation by familial Alzheimer's disease (FAD)-linked mutant forms of presenilin-1. Journal of neuroscience research. 2000;62(2):311–7. [DOI] [PubMed] [Google Scholar]
- 10. Fischer A, Schumacher N, Maier M, Sendtner M, Gessler M. The Notch target genes Hey1 and Hey2 are required for embryonic vascular development. Genes Dev. 2004;18(8):901–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Beets K, Huylebroeck D, Moya IM, Umans L, Zwijsen A. Robustness in angiogenesis: Notch and BMP shaping waves. Trends in genetics: TIG. 2013;29(3):140–9. 10.1016/j.tig.2012.11.008 [DOI] [PubMed] [Google Scholar]
- 12. Itoh F, Itoh S, Goumans MJ, Valdimarsdottir G, Iso T, Dotto GP, et al. Synergy and antagonism between Notch and BMP receptor signaling pathways in endothelial cells. EMBO J. 2004;23(3):541–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Moya IM, Umans L, Maas E, Pereira PN, Beets K, Francis A, et al. Stalk cell phenotype depends on integration of Notch and Smad1/5 signaling cascades. Dev Cell. 2012;22(3):501–14. 10.1016/j.devcel.2012.01.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Larrivee B, Prahst C, Gordon E, del Toro R, Mathivet T, Duarte A, et al. ALK1 signaling inhibits angiogenesis by cooperating with the Notch pathway. Dev Cell. 2012;22(3):489–500. 10.1016/j.devcel.2012.02.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Ricard N, Ciais D, Levet S, Subileau M, Mallet C, Zimmers TA, et al. BMP9 and BMP10 are critical for postnatal retinal vascular remodeling. Blood. 2012;119(25):6162–71. 10.1182/blood-2012-01-407593 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Jakobsson L, Franco CA, Bentley K, Collins RT, Ponsioen B, Aspalter IM, et al. Endothelial cells dynamically compete for the tip cell position during angiogenic sprouting. Nat Cell Biol. 2010;12(10):943–53. 10.1038/ncb2103 [DOI] [PubMed] [Google Scholar]
- 17. Leimeister C, Dale K, Fischer A, Klamt B, Hrabe de Angelis M, Radtke F, et al. Oscillating expression of c-Hey2 in the presomitic mesoderm suggests that the segmentation clock may use combinatorial signaling through multiple interacting bHLH factors. Dev Biol. 2000;227(1):91–103. [DOI] [PubMed] [Google Scholar]
- 18. Imayoshi I, Isomura A, Harima Y, Kawaguchi K, Kori H, Miyachi H, et al. Oscillatory control of factors determining multipotency and fate in mouse neural progenitors. Science. 2013;342(6163):1203–8. 10.1126/science.1242366 [DOI] [PubMed] [Google Scholar]
- 19. Hirata H, Yoshiura S, Ohtsuka T, Bessho Y, Harada T, Yoshikawa K, et al. Oscillatory expression of the bHLH factor Hes1 regulated by a negative feedback loop. Science. 2002;298(5594):840–3. [DOI] [PubMed] [Google Scholar]
- 20. Yoshiura S, Ohtsuka T, Takenaka Y, Nagahara H, Yoshikawa K, Kageyama R. Ultradian oscillations of Stat, Smad, and Hes1 expression in response to serum. Proc Natl Acad Sci U S A. 2007;104(27):11292–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Weng AP, Nam Y, Wolfe MS, Pear WS, Griffin JD, Blacklow SC, et al. Growth suppression of pre-T acute lymphoblastic leukemia cells by inhibition of notch signaling. Mol Cell Biol. 2003;23(2):655–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Phng LK, Gerhardt H. Angiogenesis: a team effort coordinated by notch. Dev Cell. 2009;16(2):196–208. 10.1016/j.devcel.2009.01.015 [DOI] [PubMed] [Google Scholar]
- 23. Adam MG, Berger C, Feldner A, Yang WJ, Wustehube-Lausch J, Herberich SE, et al. Synaptojanin-2 binding protein stabilizes the Notch ligands DLL1 and DLL4 and inhibits sprouting angiogenesis. Circ Res. 2013;113(11):1206–18. 10.1161/CIRCRESAHA.113.301686 [DOI] [PubMed] [Google Scholar]
- 24. Herrera B, Inman GJ. A rapid and sensitive bioassay for the simultaneous measurement of multiple bone morphogenetic proteins. Identification and quantification of BMP4, BMP6 and BMP9 in bovine and human serum. BMC cell biology. 2009;10:20 10.1186/1471-2121-10-20 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Yu PB, Deng DY, Lai CS, Hong CC, Cuny GD, Bouxsein ML, et al. BMP type I receptor inhibition reduces heterotopic [corrected] ossification. Nat Med. 2008;14(12):1363–9. 10.1038/nm.1888 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Wrighton KH, Lin X, Yu PB, Feng XH. Transforming Growth Factor {beta} Can Stimulate Smad1 Phosphorylation Independently of Bone Morphogenic Protein Receptors. J Biol Chem. 2009;284(15):9755–63. 10.1074/jbc.M809223200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Oh SP, Seki T, Goss KA, Imamura T, Yi Y, Donahoe PK, et al. Activin receptor-like kinase 1 modulates transforming growth factor-beta 1 signaling in the regulation of angiogenesis. Proc Natl Acad Sci U S A. 2000;97(6):2626–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Dahlqvist C, Blokzijl A, Chapman G, Falk A, Dannaeus K, Ibanez CF, et al. Functional Notch signaling is required for BMP4-induced inhibition of myogenic differentiation. Development. 2003;130(24):6089–99. [DOI] [PubMed] [Google Scholar]
- 29. Lu FM, Lux SE. Constitutively active human Notch1 binds to the transcription factor CBF1 and stimulates transcription through a promoter containing a CBF1-responsive element. Proc Natl Acad Sci U S A. 1996;93(11):5663–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Pourquie O. Vertebrate segmentation: from cyclic gene networks to scoliosis. Cell. 2011;145(5):650–63. 10.1016/j.cell.2011.05.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Li F, Lan Y, Wang Y, Wang J, Yang G, Meng F, et al. Endothelial Smad4 maintains cerebrovascular integrity by activating N-cadherin through cooperation with Notch. Dev Cell. 2011;20(3):291–302. 10.1016/j.devcel.2011.01.011 [DOI] [PubMed] [Google Scholar]
- 32. Morikawa M, Koinuma D, Tsutsumi S, Vasilaki E, Kanki Y, Heldin CH, et al. ChIP-seq reveals cell type-specific binding patterns of BMP-specific Smads and a novel binding motif. Nucleic Acids Res. 2011;39(20):8712–27. 10.1093/nar/gkr572 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Iso T, Maeno T, Oike Y, Yamazaki M, Doi H, Arai M, et al. Dll4-selective Notch signaling induces ephrinB2 gene expression in endothelial cells. Biochem Biophys Res Commun. 2006;341(3):708–14. [DOI] [PubMed] [Google Scholar]
- 34. Nakagawa O, McFadden DG, Nakagawa M, Yanagisawa H, Hu T, Srivastava D, et al. Members of the HRT family of basic helix-loop-helix proteins act as transcriptional repressors downstream of Notch signaling. Proc Natl Acad Sci U S A. 2000;97(25):13655–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Kobayashi T, Mizuno H, Imayoshi I, Furusawa C, Shirahige K, Kageyama R. The cyclic gene Hes1 contributes to diverse differentiation responses of embryonic stem cells. Genes Dev. 2009;23(16):1870–5. 10.1101/gad.1823109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Miyazono K, Kamiya Y, Morikawa M. Bone morphogenetic protein receptors and signal transduction. Journal of biochemistry. 2010;147(1):35–51. 10.1093/jb/mvp148 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the paper.