

Abstract
Despite worldwide dissemination of New Delhi metallo-β-lactamase1 (blaNDM-1), outbreaks remain uncommon in China. In this study, we describe the characteristics of the outbreak-related blaNDM-1-producing K. pneumonia isolates in a neonatal unit in Shandong province, China. We recovered 21 non-repetitive carbapenem-resistant K. pneumoniae isolates with a positively modified Hodge test (MHT) or EDTA synergistic test from patients and environmental samples in Shandong provincial hospital. Pulsed-field gel electrophoresis (PFGE) and multilocus sequence typing (MLST) data show K. pneumoniae isolates from 19 patients were clonally related and belong to the clonal groups ST20 and ST17. We note two outbreaks, the first caused by ST20 during August 2012 involving four patients, and the second caused by ST20 and ST17 during January 2012 and September 2013 involving fourteen patients. We found the bed railing of one patient was the source of the outbreak. We verified the presence of the blaNDM-1 gene in 21 K. pneumoniae isolates. The genes blaCTX-M-15, blaCTX-M-14, blaDHA-1, blaTEM-1 and Class I integron were also present in 18 (85.7%), 3 (14.3%), 18 (85.7%), 19 (90.5%) and 19 (90.5%) isolates, respectively. We also found an isolate with both blaNDM-1 and blaIMP-4. All of the isolates exhibited a multidrug-resistance phenotype. The β-lactam resistance of 20 isolates was transferable via conjugation. In addition, we show the resistance of 21 K. pneumoniae isolates to carbapenem is not related to lack of outer-membrane proteins OmpK35 and OmpK36 nor overpression of efflux pumps. This study provides the first report confirming blaNDM-1-producing K. pneumoniae ST20 and ST17 were associated with outbreak. Early detection of resistance genes is an effective strategy in preventing and controlling infection by limiting the dissemination of these organisms.
Introduction
K. pneumonia is a main pathogen in communities and hospitals. It causes infection in the respiratory tract, urinary tract, bloodstream and abdominal cavity. Carbapenem is the most effective antibiotic for treatment of gram negative bacilli that are resistant to β-Lactamases. However, in recent years, widespread use of carbapenem has accelerated the growth of carbapenem-resistant strains with reports in many countries [1–3].
Carbapenem resistance in enterobacteriaceae is mainly caused by two mechanisms: one by the production of carbapenemase (including class A serine carbapenemase: blaKPC, blaGES, blaSME, blaNMC, blaIMI; class B metal enzyme: blaNDM, blaIMP, blaVIM, blaSIM, blaSPM, blaGIM; class D enzyme: blaOXA); and the second by an absence or decreased expression of outer membrane proteins. In addition, mutations in the AMPC enzyme or extended-spectrum-β-lactamases (ESBL) play an important role in the drug-resistance mechanisms.
Class A and class B carbapenemases, especially blaKPC, blaIMP and blaNDM, are more commonly reported in enterobacteriaceae. Steinmann et al. reported on the hospital outbreak caused by blaKPC-2-producing K. pneumoniae in July 2010 - January 2011 in Germany that resulted in four deaths[4]. In March 2011, blaNDM-1-producing E.coli was isolated in Hong Kong, China [5]. Occurrences of carbapenem-resistant enterobacteriaceae (CRE) have been rising gradually in recent years in Mainland China, and have caused dissemination of nosocomial infection. An outbreak of 77 blaKPC-2-producing enterobacteriaceae was reported in Shanghai Huashan hospital in 2012, severely stressing the ability of hospitals and clinics to control the infection[6]. Fortunately, an outbreak of blaNDM-1-producing K.pneumoniae has not been reported in Chinese hospitals.
In this study, we collected 21 carbapenem-resistant K. pneumoniae from patients in the neonatal and cardiac surgery ward from August 2012 to August 2013. We investigated the drug-resistance and transmission mechanisms of carbapenem-resistant strains and homogeneity of strains.
Materials and Methods
Collection of isolates
Twenty-one non-duplicated carbapenem-resistant K. pneumoniae were included in this study. Twenty isolates were from neonatal and one from cardiac surgery during August 2012 to September 2013 in Shandong province, China. Swabs were collected from hands and noses of doctors and nursing staff in the neonatal unit; and environmental cultures were collected using a swab pre-moistened with sterile saline to identify other potential reservoirs for blaNDM-1 producing K. pneumoniae. Cultured sites included bed railing, stethoscope, weighing machine, milk powder, suction apparatus, wash basin, radiant warmer, washing area, electric switch, medicine tray and ventilator machine. All isolates were identified by a VITEK-2 compact system and screened for carbapenemase production by the modified Hodge test (MHT) and ertapenem-EDTA double-disc synergistic test [7]. Informed consent was obtained from the participants before the study. The participants provide written informed consent to participate in this study. The subjects’ rights and interests were protected well in the research. This study and consent procedure was approved by the Medical Ethics Committee of Shandong Provincial Hospital affiliated with Shandong University.
Susceptibility testing
Antibiotic susceptibility testing was assayed by the agar dilution method. We measured minimum inhibitory concentration (MICs) of cefotaxime, ceftazidime, cefepime, cefoxitin, piperacillin-tazobactam, trimethoprim-sulfamethoxazole, meropenem, imipenem, ertapenem, gentamicin, levofloxacin, amikacin, aztreonam, tigecycline, colisin and fosfomycin. All antibiotics, except tigecycline and colisin, were interpreted according to the approved standard of Clinical and Laboratory Standards Institute (CLSI) 2013 guidelines [8]. The 2013 European Committee on Antimicrobial Susceptibility Testing breakpoint was used (available at http://www.eucast.org/clinicalbreakpoint) for colistin and tigectcline. E.coli ATCC25922 was used for quality control.
PCR and DNA sequence analysis of drug-resistance genes
We screened carbapenem-resistance genes (blaKPC, blaSME, blaIMI/blaNMC, blaGES, blaIMP, blaVIM, blaGIM, blaSIM-1, blaSPM, blaNDM-1 and blaOXA), common ESBL genes (blaCTX-M, blaTEM, and blaSHV), AMPC genes (blaMOX, blaFOX, blaDHA, blaCIT, and blaEBC), as well as integron structures (Int1, Int2.Int3) in all clinical strains using previously described primers [9–12]. The positive PCR products were screened by electrophoresis on 1.0% agarose gel and sequenced by Shanghai Majorbio Bio-Pharm Technology Co. (Shanghai, China). Nucleotide sequences were analyzed and compared using BLAST (http://www.ncbi.nlm.nih.gov/BLAST).
Resistance gene transfer experiments
Conjugation experiments were performed using azide-resistant E.coli J53 as the recipient strain. Briefly, overnight cultures of the donor strain (500 ml) and recipient strain (500 ml) were mixed with 10 ml fresh Mueller-Hinton broth and incubated for 24 h at 35°C. Mixture was inoculated on MacConkey agar plates containing sodium azide (200 ug/ml) and imipenem (0.5 ug/ml) for 24 h at 35°C. Conjugation was confirmed by indole testing. Presence of carbapenemase gene was confirmed by PCR analysis.
Plasmid profiling
Plasmid DNA of transconjugants was purified using a QuickGene plasmid kit S (Fuji, Tokyo, Japan) according to the manufacturer's instructions, followed by electrophoresis in a 0.7% agarose gel at 90 V for 3 h [13]. The sizes of the plasmids were compared by co-electrophoresis with plasmids of known sizes from E. coli (V517and 39R861). Bands were visualized with UV transilluminator after staining.
Inhibitory effects of CCCP against efflux
We determined presence of efflux pump in carbapenem-resistant enterobacteriaceae by CCCP inhibition test. MIC changes were observed in the absence or presence of CCCP (Sigma) at concentrations of 50μg/mL. An inoculum of each isolate (5 × 104 cfu/mL) was inoculated onto MH medium containing serial dilutions of imipenem and meropenem. A phenotype for positive efflux was detected after at least 4-fold dilutions of MIC for imipenem or meropenem in the absence or presence of CCCP [14].
Pulse-field gel electrophoresis (PFGE)
An overnight-grown bacterial culture was suspended in EET buffer (100 mM EDTA, 10 mM EGTA, 10 mM Tris—HCl (pH 8)) and adjusted to an optical density of 0.9 at 600 nm. The suspension was mixed with equal volumes in a 2% solution of low-melting-temperature agarose in EET buffer. After solidification, the agarose plugs were incubated for 4 h at 37°C in EET buffer containing 1 mg lysozyme and 50 ug lysostaphin per ml. The plugs were transferred to EET buffer containing 1% sodium dodecy1 sulfate and 20 mg proteinase K per ml of buffer and incubated overnight at 50°C. Plugs were washed thoroughly with TE buffer (10mM Tris—HCl, 1 mM EDTA (pH 8)) and digested overnight with XbaI restriction endonuclease (TAKARA, Shiga, Japan). DNA separation was performed in 0.5× TBE buffer in a pulsed-field electrophoresis system (CHEF MAPPER; Bio-Rad Laboratories, California, USA) with the following conditions: temperature 14°C; voltage 6 V/cm; switch angle, 120°and switch ramp of 4–40 s for 21h. Salmonella enterica serotype Braenderup H9812 was used as a marker for PFGE. The restriction patterns were analyzed and interpreted according to Tenover et al [15].
Multilocus sequence typing (MLST)
MLST of K.pneumoniae was performed using seven conserved housekeeping genes (blagapA, blainfB, blamdh, blapgi, blaphoE, blarpoB and blatonB) according to protocols available at the MLST Pasteur website (http://www.pasteur.fr/recherche/genopole/PF8/mlst/Kpneumoniae.html).
Analysis of outer membrane proteins (OMPs)
The genes of blaOMPs were screened in all clinical isolates using previously described primers by PCR [16]. PCR negative isolates were investigated for alterations in the OMPs by Sodium dodecy1 sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) as described previously [17].
Results
Clinical characteristics of 21 K. pneumoniae
Table 1 shows the clinical characteristics of the 21 carbapenem-resistant K. pneumoniae, including isolation date, specimen and ward distribution. The first strain was obtained on August 1, 2012, with three additional samples isolated from the neonatal unit. No additional isolates were observed between September and December. However, beginning in January 2013, the isolation rate significantly increased in the same ward, with 15 strains isolated from January to August. On September 10, 2013, one strain was isolated from a cardiac surgery patient with congenital heart disease. We isolated one from the bed railing of patient (H16) in September as well. Cultures from physicians, nurses and environmental samples were negative.
Table 1. Characteristics of 21 K. pneumoniae isolates with NDM-1.
| Isolate No. | Sex | Patient age | Ward | Isolate date | Specimen | HTM | EDTA | PFGE pattern | Sequence type | Resistance gene | Class I integrons | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Carbapenemase | ESBL | AMPC | |||||||||||
| H1 | M | 30d | neonatal unit | 2012/8/1 | sputum | - | + | cluster1 | 20 | blaNDM-1 | blaTEM-1,blaCTX-M-15 | blaDHA-1 | + |
| H2 | M | 21d | neonatal unit | 2012/8/23 | sputum | - | + | cluster1 | 20 | blaNDM-1 | blaTEM-1,blaCTX-M-15 | blaDHA-1 | + |
| H3 | M | 1d | neonatal unit | 2012/8/29 | sputum | ± | - | cluster1 | 20 | blaNDM-1 | blaTEM-1,blaCTX-M-15 | blaDHA-1 | + |
| H4 | M | 13d | neonatal unit | 2012/8/30 | sputum | - | + | cluster1 | 20 | blaNDM-1 | blaTEM-1,blaCTX-M-15 | blaDHA-1 | + |
| H5 | M | 60d | neonatal unit | 2013/1/1 | sputum | - | + | cluster1 | 20 | blaNDM-1 | blaTEM-1,blaCTX-M-15 | blaDHA-1 | + |
| H6 | F | 22d | neonatal unit | 2013/1/29 | sputum | - | + | cluster1 | 20 | blaNDM-1 | blaTEM-1,blaCTX-M-15 | blaDHA-1 | + |
| H7 | M | 30d | neonatal unit | 2013/2/25 | sputum | - | + | cluster1 | 20 | blaNDM-1 | blaTEM-1,blaCTX-M-15 | blaDHA-1 | + |
| H8 | M | 56d | neonatal unit | 2013/3/5 | sputum | ± | + | cluster1 | 20 | blaNDM-1 | blaCTX-M-15 | blaDHA-1 | + |
| H9 | M | 28d | neonatal unit | 2013/3/6 | sputum | - | + | cluster1 | 20 | blaNDM-1 | blaTEM-1,blaCTX-M-15 | blaDHA-1 | + |
| H10 | F | 25d | neonatal unit | 2013/3/14 | sputum | - | + | cluster1 | 20 | blaNDM-1 | blaTEM-1,blaCTX-M-15 | blaDHA-1 | + |
| H11 | M | 32d | neonatal unit | 2013/3/19 | sputum | - | + | cluster1 | 20 | blaNDM-1 | blaCTX-M-15 | blaDHA-1 | + |
| H12 | M | 34d | neonatal unit | 2013/4/19 | sputum | - | - | cluster1 | 20 | blaNDM-1 | blaTEM-1,blaCTX-M-15 | blaDHA-1 | + |
| H13 | M | 13d | neonatal unit | 2013/4/23 | umbilicus | + | - | cluster4 | 54 | blaNDM-1 | blaTEM-1,blaCTX-M-15 | blaDHA-1 | + |
| H14 | F | 15d | neonatal unit | 2013/7/10 | sputum | - | - | cluster3 | 20 | blaNDM-1 | blaTEM-1,blaCTX-M-15 | blaDHA-1 | + |
| H15 | M | 9d | neonatal unit | 2013/7/13 | sputum | - | - | cluster1 | 20 | blaNDM-1 | blaTEM-1,blaCTX-M-15 | blaDHA-1 | + |
| H16 | M | 35d | neonatal unit | 2013/7/21 | sputum | + | - | cluster1 | 20 | blaNDM-1 | blaTEM-1,blaCTX-M-15 | blaDHA-1 | + |
| J17 | neonatal unit | 2013/7/24 | bed railing | - | + | cluster1 | 20 | blaNDM-1 | blaTEM-1,blaCTX-M-15 | blaDHA-1 | + | ||
| H18 | M | 1d | neonatal unit | 2013/7/27 | sputum | + | + | cluster2 | 17 | blaNDM-1 | blaTEM-1,blaCTX-M-14 | - | - |
| H19 | F | 26d | neonatal unit | 2013/7/29 | blood | ± | + | cluster1 | 20 | blaNDM-1 | blaTEM-1,blaCTX-M-14 | blaDHA-1 | + |
| H20 | F | 2d | neonatal unit | 2013/8/19 | sputum | - | + | cluster2 | 17 | blaNDM-1 | blaTEM-1,blaCTX-M-15 | - | - |
| H21 | F | 1y | cardiac surgery | 2013/9/10 | sputum | ± | + | cluster2 | 17 | blaNDM-1,blaIMP-4 | blaTEM-1,blaCTX-M-14 | - | dfrA1-orfC |
Kpn: Klebsiella pneumoniae, M: male, F: female, EDTA: EDTA synergistic test, HTM: modified Hodge test, ESBL: extended-spectrum-β-lactam
19 isolates were from newborns in the neonatal unit, including 14 males and 5 females. Eighteen newborns had pneumonia and one had umbilical infection. All were premature with low birth weight. Nine had intrauterine infection at birth. All were previously treated with various antimicrobial agents including cloxacillin, cefathiamidine and ceftizoxime. Eight were treated with imipenem or meropenem. The patient from cardiac surgery had respiratory tract infection and fever five days after operation.
Susceptibility results
The MICs of each antibiotic are shown in Table 2. 21 K. pneumoniae isolates carrying blaNDM-1 exhibited resistance to cefotaxime, ceftazidime, cefepime, cefoxitin, piperacillin-tazobactam, meropenem, imipenem and ertapenem. Tigecycline, colistin, levofloxacin and amikacin showed strong activity against carbapenem-resistant K. pneumoniae with a susceptibility rate of 100%. The antibiotic resistant rates of gentamicin, aztreonam, trimethoprim-sulfamethoxazole and fosfomycin were 14.28% (3/21), 95.24% (20/21), 23.81% (5/21), and 4.76% (1/21), respectively. It is worth noting that based on MLST results, these gentamicin-resistant K. pneumoniae all belong to ST17, of which two were derived from neonatal unit and one from cardiac surgery.
Table 2. The antibiotic susceptibility of blaNDM-producing k. pneumonia (μg/ml).
| isolate No. | IMP | MEM | ETP | CN | AK | TZP | CTX | CAZ | FEP | FOX | LEV | SXT | TGC | CO | FOS | ATM |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H1 | 8 | 8 | >32 | 0.5 | 2 | >256 | >256 | >256 | >256 | >256 | 0.5 | >32 | 0.5 | 0.125 | 32 | >256 |
| H2 | 24 | 8 | >32 | 0.5 | 4 | >256 | >256 | >256 | >256 | >256 | 0.5 | 0.125 | 0.5 | 0.125 | 48 | >256 |
| H3 | >32 | >32 | >32 | 0.5 | 2 | >256 | >256 | >256 | >256 | >256 | 1 | 0.125 | 0.5 | 0.125 | 48 | >256 |
| H4 | >32 | >32 | >32 | 1 | 4 | >256 | >256 | >256 | >256 | >256 | 1 | 0.125 | 0.5 | 0.125 | 32 | >256 |
| H5 | >32 | >32 | >32 | 1 | 4 | >256 | >256 | >256 | >256 | >256 | 0.5 | 0.25 | 0.5 | 0.125 | 16 | >256 |
| H6 | >32 | >32 | >32 | 1 | 2 | >256 | >256 | >256 | >256 | >256 | 1 | 0.125 | 0.5 | 0.125 | 32 | >256 |
| H7 | >32 | >32 | >32 | 0.5 | 2 | >256 | >256 | >256 | >256 | >256 | 1 | 0.25 | 0.5 | 0.125 | 16 | >256 |
| H8 | >32 | >32 | >32 | 0.5 | 2 | >256 | >256 | >256 | >256 | >256 | 1 | 0.25 | 0.25 | 0.125 | 48 | >256 |
| H9 | 32 | 32 | 32 | 0.5 | 4 | >256 | >256 | >256 | >256 | >256 | 1 | 0.25 | 1 | 0.125 | 24 | >256 |
| H10 | >32 | >32 | >32 | 0.5 | 2 | >256 | >256 | >256 | >256 | >256 | 1 | 0.25 | 0.5 | 0.032 | 48 | >256 |
| H11 | >32 | >32 | >32 | 0.5 | 2 | >256 | >256 | >256 | >256 | >256 | 1 | 0.25 | 0.5 | 0.032 | 32 | >256 |
| H12 | >32 | >32 | >32 | 1 | 2 | >256 | >256 | >256 | >256 | >256 | 1 | 0.125 | 0.5 | 0.125 | 48 | >256 |
| H13 | 4 | 32 | 32 | 0.5 | 4 | >256 | >256 | >256 | >256 | >256 | 1 | >256 | 1 | 0.125 | >256 | 1 |
| H14 | 32 | 32 | >32 | 0.5 | 2 | >256 | >256 | >256 | >256 | >256 | 1 | 0.125 | 1 | 0.25 | 48 | >256 |
| H15 | >32 | >32 | >32 | 0.5 | 4 | >256 | >256 | >256 | >256 | >256 | 1 | 0.125 | 0.5 | 0.25 | 48 | >256 |
| H16 | >32 | >32 | >32 | 0.5 | 4 | >256 | >256 | >256 | >256 | >256 | 0.5 | 0.5 | 1 | 0.25 | 16 | >256 |
| J17 | >32 | >32 | >32 | 0.5 | 4 | >256 | >256 | >256 | >256 | >256 | 0.5 | 0.5 | 0.5 | 0.25 | 32 | >256 |
| H18 | >32 | 8 | >32 | 32 | 2 | >256 | >256 | >256 | >256 | >256 | 0.5 | >32 | 1 | 0.25 | 16 | >256 |
| H19 | >32 | >32 | >32 | 0.5 | 4 | >256 | >256 | >256 | >256 | >256 | 0.5 | 0.5 | 1 | 0.25 | 48 | >256 |
| H20 | >32 | 4 | 32 | 32 | 2 | >256 | >256 | >256 | >256 | >256 | 1 | >32 | 0.5 | 0.125 | 16 | >256 |
| H21 | >32 | >32 | >32 | 16 | 2 | >256 | >256 | >256 | >256 | >256 | 0.5 | >32 | 1 | 0.125 | 16 | >256 |
MIC: Minimum inhibitory concentration, CTX: cefotaxime, CAZ: ceftazidime, FEP: cefepime, FOX: cefoxitin, TZP: piperacillin-tazobactam, SXT: trimethoprim-sulfamethoxazole, MEM: meropenem, IMP: imipenem, EPT: ertapenem, CN: gentamicin, LEV: levofloxacin, AK: amikacin, ATM: aztreonam, TGC: tigecycline, CO: colisin FOS: fosfomycin; EC: E.coli
Characterization of resistance genes, OMPs and efflux analysis
All isolates resistant to ertapenem (MICs > 8 mg/L) were positive in EDTA synergistic test, and seven of them were positive or weakly positive in the modified Hodge test (MHT). We confirmed the presence and production of metallo-β-lactamas (MBL) by sequencing. All 21 isolates carried blaNDM-1, and one isolate from cardiac surgery also carried blaIMP-4. In addition to MBL, we examined other types of β-lactamase, including ESBL and AMPC. The distributions of resistance genes in these strains are listed in Table 1. We identified the ESBL genes as blaCTX-M-15, blaTEM-1, and blaCTX-M-14 in 18 (85.7%), 19 (90.5%) and 3 (14.3%) isolates, respectively. Genotyping results of AMPC genes confirmed blaDHA-1 in 18 (85.7%) isolates. Moreover, 19 strains carried class 1 integron. Sequence data from the variable region showed the class 1 integron of all 17 isolates belonging to ST20 had no resistant gene cassettes. Only isolate H21, belonging to ST 17, carried two different gene cassettes (dfrA1 gene cassette encoding resistance determinants to trimethoprim and orfC gene cassette).
We detected the genes of outer membrane proteins OmpK35 and OmpK36 in all 21 isolates. The base deletions of OmpK36 genes were observed in H21 and H15. OMPs of H21 and H15 were detected by SDS-PAGE. Compared with K. pneumoniae ATCC13883, OmpK35 and OmpK36 remained in H21 and H15 with molecular weights of 35–40 kDa (Fig. 1). We found no change in MIC for imipenem or meropenem in the absence or presence of CCCP.
Fig 1. SDS-PAGE of outer membrane proteins. M: Marker; 13883: ATCC13883; B1: strain with OmpK36 deficiency.

Association among sequence typing, resistance genes expression and antibiotic susceptibility
All 21 K. pneumoniae with blaNDM-1 were susceptible to levofloxacin, amikacin, tigecycline, and colisin. Among them, 17 isolates of ST20 carried blaCTX-M-15, blaDHA-1 and class 1 integron; and 15 isolates of ST20 also carried blaTEM-1. 17 isolates of ST20 were also susceptible to gentamicin and fosfomycin. Only isolate H1 was resistant to trimethoprim-sulfamethoxazole. Three K. pneumoniae of ST17 carried blaTEM-1 and blaCTX-M-14 in addition to blaNDM-1. Isolate H21 also carried blaIMP-4 and class 1 integron containing gene cassettes. Compared with ST20, K. pneumoniae ST17 did not carry blaDHA-1; and all ST17 were resistant to gentamicin and trimethoprim-sulfamethoxazole. Although one K. pneumoniae of ST54 produced the same β-lactamase gene as ST20, it was resistant to trimethoprim-sulfamethoxazole and fosfomycin. It is interesting to note that only K. pneumoniae ST54 was sensitive to aztreonam.
Transfer of carbapenem resistance
Our results show that β-lactam resistance was successfully transferred to E. coli J53 from K. pneumonia in 20 isolates. Compared to the recipient strain E.coli J53, all E. coli transconjugants exhibited significantly reduced carbapenem susceptibility. MIC of imipenem, meropenem and etarpenem increased 8–64 fold, 32–512 fold and 256–2048 fold, respectively (Table 3). The sensitivities of cefotaxime, ceftazidime, cefepime, cefoxitin, piperacillin-tazobactam and aztreonam were similar to donor strains. All E. coli transconjugants showed positive EDTA synergistic tests with the same carbapenemase genes as the donor. Plasmid profiling revealed that transconjugants acquired single high-molecular-weight plasmids of approximately 336 kb.
Table 3. The results of antibiotic susceptibility testing of E.coli J53 transconjugant strains derived from blaNDM-producing k. pneumonia transconjugants (μg/ml).
| isolate No. | IMP | MEM | ETP | CN | AK | TZP | CTX | CAZ | FEP | FOX | LEV | SXT | TGC | CO | FOS | ATM |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H1-J53 | 2 | 2 | >32 | <1 | <2 | 64 | >256 | >256 | 8 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | 32 |
| H2-J53 | 2 | 2 | >32 | <1 | <2 | 64 | >256 | >256 | >256 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | >256 |
| H3-J53 | 2 | 4 | 4 | <1 | <2 | 64 | >256 | >256 | 8 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | >256 |
| H4-J53 | >32 | >32 | >32 | <1 | <2 | >256 | >256 | >256 | >256 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | >256 |
| H5-J53 | >32 | >32 | >32 | <1 | <2 | >256 | >256 | >256 | >256 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | >256 |
| H6-J53 | 2 | 2 | 16 | <1 | <2 | >256 | >256 | >256 | 8 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | >256 |
| H7-J53 | 2 | 2 | 4 | <1 | <2 | >256 | >256 | >256 | >256 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | >256 |
| H8-J53 | 4 | 4 | 4 | <1 | <2 | >256 | >256 | >256 | >256 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | >256 |
| H9-J53 | 32 | 32 | 32 | <1 | <2 | >256 | >256 | >256 | >256 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | >256 |
| H10-J53 | 2 | 4 | >32 | <1 | <2 | >256 | >256 | >256 | 8 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | >256 |
| H11-J53 | >32 | >33 | >33 | <1 | <2 | >256 | >256 | >256 | >256 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | >256 |
| H12-J53 | >32 | >32 | >32 | <1 | <2 | >256 | >256 | >256 | >256 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | >256 |
| H13-J53 | 4 | 32 | 32 | <1 | <2 | >256 | >256 | >256 | >256 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | 1 |
| H14-J53 | 32 | 32 | >32 | <1 | <2 | >256 | >256 | >256 | >256 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | >256 |
| H15-J53 | >32 | >32 | >32 | <1 | <2 | >256 | >256 | >256 | >256 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | >256 |
| H16-J53 | >32 | >32 | >32 | <1 | <2 | >256 | >256 | >256 | >256 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | >256 |
| J17-J53 | 2 | >32 | >32 | <1 | <2 | >256 | >256 | >256 | 32 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | >256 |
| H18-J53 | >32 | 8 | >32 | <1 | <2 | >256 | >256 | >256 | >256 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | >256 |
| H19-J53 | >32 | >32 | >32 | <1 | <2 | >256 | >256 | >256 | >256 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | >256 |
| H20-J53 | >32 | 4 | 32 | <1 | <2 | >256 | >256 | >256 | >256 | >256 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | >256 |
| EC J53 | <1 | <1 | <0.5 | <1 | <2 | <0.5 | <1 | <1 | <1 | <1 | <0.25 | <0.5 | 0.25 | 0.5 | 2 | <1 |
MIC: Minimum inhibitory concentration, CTX: cefotaxime, CAZ: ceftazidime, FEP: cefepime, FOX: cefoxitin, TZP: piperacillin-tazobactam, SXT: trimethoprim-sulfamethoxazole, MEM: meropenem, IMP: imipenem, EPT: ertapenem, CN: gentamicin, LEV: levofloxacin, AK: amikacin, ATM: aztreonam, TGC: tigecycline, CO: colisin FOS: fosfomycin; EC J53: recipient
Molecular epidemiology of 21 K. pneumoniae
PFGE revealed 4 distinct clusters among the 21 K. pneumoniae isolates (Fig. 2). Strain H13, isolated from umbilicus, belonged to ST54, which was a different clone from the strains isolated from sputum and blood. Isolates H18, H20 and H21 belonged to the same clone, sharing the same sequence type as ST17. Isolate H21 was from cardiac surgery. Compared with PFGE that divided other strains into 2 different clusters, MLST assigned them into the same ST20 cluster. Our results show two clones of K. pneumoniae spread in the neonatal unit. Two outbreaks caused by K. pneumoniae (ST20) were noted, the first during August 2012 involving four patients and the second beginning in January 2013 involving twelve patients. During the second outbreak involving three patients (two from neonatal unit and one from cardiac surgery), the clone K. pneumoniae (ST17) was isolated. K. pneumoniae ST17 and ST20 caused newborn pneumonia. ST17 spread from the neonatal unit to the cardiac surgery unit. Fortunately, K. pneumoniae ST54, cause of neonatal umbilical infection, did not disseminate. Ultimately, only one patient died and all other patients were treated and discharged. Results show the source strain (J17) K. pneumoniae, belonging to ST20, was from the bed railing of patient (H16).
Fig 2. Dendrogram analysis.
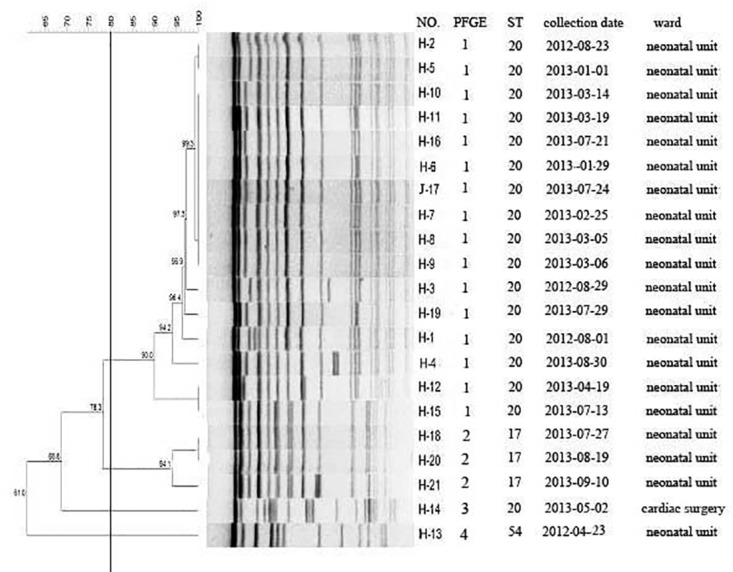
Dendrogram generated with the Fingerprinting II Informatix software package (Bio-Rad Laboratories, Hercules, CA) showing the relatedness of fingerprints (XbaI-PFGE) for 21 K. pneumoniae. The phylogenetic tree was constructed using the Dice coefficient and UPGMA clustering. A genetic similarity index scale is shown in the left of the dendrogram. PFGE types, strain number, collection data and wards distribution information are included along each PFGE lane. No., number
Discussion
Carbapenem antibiotics are the primary treatment of serious infections caused by ESBL-producing gram-negative bacteria. Once carbapenem-resistant strains emerge, no drugs are available for treatment, with limited therapeutic choices including aminglycosides, polymxins, tigecycline, fosfomycin and temocillin. Therefore, the issue of carbapenem—resistance in enterobacteriaceae deserves special attention. In our hospital, K. pneumoniae producing blaNDM-1 was mainly derived from neonatal wards. The prematurity, very low body weight and poor immunity of newborns greatly increased chances of infection. The investigation of the surrounding environment revealed that the source of the outbreak was the bed railing of a patient who carried carbapenem-resistant K. pneumoniae, underscoring the importance of strict disinfection.
The use of antimicrobial drugs is a risk factor for infection of carbapenem—resistant enterobacteriaceae (CRE). Clinical data show that two weeks before patients were treated with cephalosporins, six patients used carbapenem antibiotics. Currently, no effective antimicrobials are available to treat CRE infection in newborns. CRE infections have a high mortality rate, approximately 70% in patients with bacteremia [18]. In this study, one patient with bloodstream infection died.
In our study, homology analysis indicated that CRE easily spread to different patients in the same ward, with a tendency to spread among multiple wards. Epidemic clones in this study were K. pneumoniae ST20 and ST17. Both have been previously identified among newborns affected by ESBL-K. pneumoniae, which produced blaCTXM-15 in Spain and Canada [19,20]. Here, we provide the first report confirming blaNDM-1-producing K. pneumoniae ST20 and ST17 caused an infection outbreak. Comparing ST20 and ST17, only the infB allele was different, with a base variant at 279 (T-C). Dortet L found that Acinetobacter sp. plays a key role in spreading blaNDM genes into enterobacteriaceae [21]. Our conjugation data indicates blaNDM-1 was transferred on a 336kb plasmid. Furthermore, two MBLs, blaIMP-4 and blaNDM-1, were initially co-expressed in a single carbapenem-resistant pathogen (K. pneumoniae ST17). However, the isolate carrying two MBLs and class 1 integron containing gene cassettes failed to transfer. We also found 3 strains of K. pneumoniae ST17 were resistant to gentamicin, and one strain was resistant to fosfomycin. Several gene cassettes encoding extended-spectrum beta-lactamases or carbapenemases have been described [22]. Most isolates with the class 1 integron in this study had no resistance gene cassettes. The spread of resistance genes was not due to class 1 integron.
Previous studies suggest OmpK plays an important role in the resistance or reduced susceptibility to carbapenems in K. pneumoniae isolates producing AmpC, ESBL or broad-spectrum β-lactamase [23, 24]. OmpK deficiency also contributes to the high-level carbapenem resistance in K. pneumoniae carrying blaKPC or blaIMP[25, 26]. In this study, although there was a high frequency of ESBL, AMPC and blaNDM-1 in 21 K. pneumonia isolates with high carbapenems MICs, we did not find a lack of outer-membrane proteins. We show base deletions of blaOmpK36 in H21 and H15, but we found no loss of protein. Shi et al suggests mutation or base deletions of protein-coding genes lead to alterations in the open reading frame, amino acid sequences and protein configuration [27]. Thus, the pore size of protein OmpK35 or OmpK36 can be affected and the accessibility of drugs can be impeded. In this study, no MIC changes of imipenem or meropenem were observed when CCCP was added. The results indicate that the resistance to carbapenems seems to be unrelated to efflux.
In conclusion, the production of blaNDM-1 coupled with blaDHA-1,blaCTX-M and blaTEM-1 β-lactamase genes plays an important role in conferring resistance of K. pneumoniae strains to carbapenems. The outbreak caused by blaNDM-1-producing K. pneumoniae highlights an urgent need to develop effective strategies for the prevention and control of infections. Our results show that strict disinfection of the environment and hand hygiene may be the most effective strategy in preventing CRE spreading. Limited application of antimicrobials, especially for carbapenems and cephalosporins, may reduce the appearance of CRE.
Acknowledgments
We thank Dr. Austin Cape for careful reading and feedback.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This study was supported by the Shandong province of Science and Technology Development Program of China (grant 2012G0021844) whose role was the study design and decision to publish; the National Natural Science Foundation of China (no.81401696) whose role was data collection and analysis; the Shandong Provincial Natural Science Foundation of China (ZR2011HM019) whose role was data collection and analysis; the Shandong Provincial Awards Foundation for Young Scientists (BS2011SW035) whose role was data collection and analysis; and the Construction Program of Shandong key clinical specialty whose role was the contribution of equipment.
References
- 1. Yong D, Toleman MA, Giske CG, Cho HS, Sundman K, Lee K, et al. Characterization of a new metallo-β-Lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob Agents Chemother. 2009; 53: 5046–5054. 10.1128/AAC.00774-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Centers for Disease Control and Prevention (CDC). Detection of Enterobacteriaceae isolates carrying metallo-beta-lactamase-United States. MMWR Morb Mortal Wkly Rep. 2010; 59: 750 [PubMed] [Google Scholar]
- 3. Struelens MJ, Monnet DL, Magiorakos AP, Santos O'Connor F, Giesecke J, European NDM-1 Survey Participants. New Delhi metallo-beta-lactamase 1–producing Enterobacteriaceae: emergence and response in Europe. Euro Surveill. 2010; 15 pii: 19716 [DOI] [PubMed] [Google Scholar]
- 4. Steinmann J, Kaase M, Gatermann S, Popp W, Steinmann E, Damman M, et al. Outbreak due to a Klebsiella pneumoniae strain harbouring KPC-2 and VIM-1 in a German university hospital, July 2010 to January. Euro Surveill. 2011; 16 pii: 19944 [PubMed] [Google Scholar]
- 5. Ho PL, Lo WU, Yeung MK, Lin CH, Chow KH, Ang I, et al. Complete sequencing of pNDM-HK encoding NDM-1carbapenemase from a multidrug-resistant Escherichia coli strain isolated in Hong Kong. PLoS One. 2011; 6: e17989 10.1371/journal.pone.0017989 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Hu F, Chen S, Xu X, Guo Y, Liu Y, Zhu D, et al. Emergence of carbapenem-resistant clinical Enterobacteriaceae isolates from a teaching hospital in Shanghai, China. J Med Microbiol. 2012; 61: 132–136. 10.1099/jmm.0.036483-0 [DOI] [PubMed] [Google Scholar]
- 7. Lee K Chong Y, Sllin HB, Kim YA, Yong D, Yum JH. Modged Hodge and EDTA disk synergy tests to screen metallo-β-lactamase producing strains of Pseudomonas and Acinetobacter specie. C1in Microbiol Infection. 2001; 7: 88–91. [DOI] [PubMed] [Google Scholar]
- 8. Clinical and Laboratory Standards Institute. Performance standards for antimicrobial susceptibility testing twentieth informational supplement, 2013, M100-S23 Wayne, PA: CLSI. [Google Scholar]
- 9. Queenan AM, Bush K. Carbapenemases: the Versatile β-Lactamases. Clin Microbiol Rev. 2007; 20:440–458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Fang H, Ataker F, Hedin G, Dornbusch K. Molecular epidermiology of Extended-Spectrum β-Lactamases among Escherichia coli isolates collected in a Swedish hospital and its associated health care facilities from 2001 to 2006. J Clin Microbio. 2008; 46:707–712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Perez-Perez FJ, Hanson ND. Detection of plasmid-mediated AmpC beta-lactamase genes in clinical isolates by using multiplex PCR. J Clin Microbio. 2002; 40: 2153–2162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Goldstein C, Lee MD, Sanchez S, Hudson C, Phillips B, Register B et al. Incidence of class 1 and 2 integrases in clinical and commensal bacteria from livestock, companion animals, and exotics. Antimicrob Agents Chemother. 2001; 45:723–726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Kado CI, Liu ST. Rapid procedure for detection and isolation of large and small plasmids. J Bacteriol. 1981; 145: 1365–1373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Pakzad I, Zayyen Karin M, Taherikalani M, Boustanshenas M, Lari AR. Contribution of AcrAB efflux pump to ciprofloxacin resistance in Klebsiella pneumoniae isolated from burn patients. GMS Hyg Infect Control. 2013; 8: Doc15 10.3205/dgkh000215 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Tenover FC, Arbeit RD, Goering RV, Mickelsen PA, Murray BE, Persing DH, et al. Interpreting chromosomal DNA restriction patterns produced by pulsed—field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol. 1995; 33: 2233–2239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Doumith M, E11ington MJ, Livennore DM, Woodford N. Molecular mechanisms disrupting porin expression in ertapenem-resistant K1ebsiella and Enterobacter spp. clinical isolates from the UK. J Antimicrob Chemother. 2009; 63: 659–667. 10.1093/jac/dkp029 [DOI] [PubMed] [Google Scholar]
- 17. Hemandez-Alles S, Alberti S, Alvarez D, Doménech-Sánchez A, Martínez-Martínez L, Gil J, et al. Pofin expression in clinical isolates of Klebsiella pneumoniae. Microbiology. 1999; 145: 673–679. [DOI] [PubMed] [Google Scholar]
- 18. Borer A, Saidel-Odes L, Riesenberg K, Eskira S, Peled N, Nativ R, et al. Attributable mortality rate for carbapenem-resistant Klebsiella pneumoniae bacteremia. Infect Control Hosp Epidemiol. 2009; 30: 972–976. 10.1086/605922 [DOI] [PubMed] [Google Scholar]
- 19. Peirano GL, Sang JH, Pitondo-Silva A, Laupl KB, Pitout JD. Molecular epidemiology of extended-spectrum-β-lactamase-producing Klebsiella pneumoniae over a 10 year period in Calgary, Canada. J Antimicrob Chemother. 2012; 67: 1114–1120. 10.1093/jac/dks026 [DOI] [PubMed] [Google Scholar]
- 20. Mavroidi A, Liakopoulos A, Gounaris A, Goudesidou M, Gaitana K, Miriagou V, et al. Successful control of a neonatal outbreak caused mainly by ST20 multidrug-resistant SHV-5-producing Klebsiella pneumoniae, Greece. BMC Pediatr. 2014;14: 105 10.1186/1471-2431-14-105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Dortet L, Poirel L, Nordmann P. Worldwide dissemination of the NDM-Type carbapenemases in gram-negative bacteria. Biomed Res Int. 2014; 2014: 249856 10.1155/2014/249856 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Fluit AC1, Schmitz FJ. Class 1 integrons, gene cassettes, mobility, and epidemiology. Eur J Clin Microbiol Infect Dis. 1999; 18: 761–770. [DOI] [PubMed] [Google Scholar]
- 23. Domenech-Sanchez A, Hemandez-Alles S, Martinez-Martinez L, Benedí VJ, Albertí S. Identification and characterization of a new porin gene of Klebsiella pneumoniae: its role in beta-lactam antibiotic resistance. J Bacteriol. 1999; 181: 2726–2732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Wang XD, Cai JC, Zhou HW, Zhang R, Chen GX. Reduced susceptibility to carbapenems in Klebsiella pneumoniae clinical isolates associated with plasmid-mediated beta-lactamase production and OmpK36 porin deficiency.J Med Microbiol.2009; 58: 1196–202. 10.1099/jmm.0.008094-0 [DOI] [PubMed] [Google Scholar]
- 25. Landman D1, Bratu S, Quale J. Contribution of OmpK36 to carbapenem susceptibility in KPC-producing Klebsiella pneumoniae. J Med Microbiol. 2009; 58: 1303–1308. 10.1099/jmm.0.012575-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Chen LR, Zhou HW, Cai JC, Zhang R, Chen GX. Combination of IMP-4 metallo-beta-lactamase production and porin deficiency causes carbapenem resistance in a Klebsiella oxytoca clinical isolate. Diagn Microbiol Infect Dis. 2009; 65:163–167. 10.1016/j.diagmicrobio.2009.07.002 [DOI] [PubMed] [Google Scholar]
- 27. Shi W, Li K, Ji Y, Jiang Q, Wang Y, Shi M, et al. Carbapenem and cefoxitin resistance of Klebsiella pneumoniae strains associated with porinOmpK36 loss and DHA-1 β-lactamase production. Braz J Microbiol. 2013; 44:435–442. 10.1590/S1517-83822013000200015 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.