

Abstract
Midkine is a heparin binding growth factor with important functions in neuronal development and survival, but little is known about its function in the retina. Previous studies show that in the developing zebrafish, Midkine-a (Mdka) regulates cell cycle kinetics in retinal progenitors, and following injury to the adult zebrafish retina, mdka is strongly upregulated in Müller glia and the injury-induced photoreceptor progenitors. Here we provide the first data describing Mdka protein localization during different stages of retinal development and during the regeneration of photoreceptors in adults. We also experimentally test the role of Mdka during photoreceptor regeneration. The immuno-localization of Mdka reflects the complex spatiotemporal pattern of gene expression and also reveals the apparent secretion and extracellular trafficking of this protein. During embryonic retinal development the Mdka antibodies label all mitotically active cells, but at the onset of neuronal differentiation, immunostaining is also localized to the nascent inner plexiform layer. Starting at five days post fertilization through the juvenile stage, Mdka immunostaining labels the cytoplasm of horizontal cells and the overlying somata of rod photoreceptors. Double immunolabeling shows that in adult horizontal cells, Mdka co-localizes with markers of the Golgi complex. Together, these data are interpreted to show that Mdka is synthesized in horizontal cells and secreted into the outer nuclear layer. In adults, Mdka is also present in the end feet of Müller glia. Similar to mdka gene expression, Mdka in horizontal cells is regulated by circadian rhythms. After the light-induced death of photoreceptors, Mdka immuonolabeling is localized to Müller glia, the intrinsic stem cells of the zebrafish retina, and proliferating photoreceptor progenitors. Knockdown of Mdka during photoreceptor regeneration results in less proliferation and diminished regeneration of rod photoreceptors. These data suggest that during photoreceptor regeneration Mdka regulates aspects of injury-induced cell proliferation.
Introduction
Midkine is a heparin-binding growth factor that forms a two-member family with Pleiotrophin. Both factors are abundantly expressed during embryogenesis, with particularly high levels in the developing nervous system [1]. Beyond mid-gestation and during postnatal stages, the expression of midkine and pleiotrophin are rapidly downregulated [2–6]. Genes encoding both Midkine and Pleiotrophin are up-regulated under disease conditions, most notably those that affect the nervous system [7–11]. For example, in rodents, Midkine is upregulated after retinal damage [11], and the up-regulation of midkine and pleiotrophin coincides with cytokine activity during nervous system repair [12–15]. Throughout the nervous system Midkine is proposed to play a role in reparative mechanisms.
The retina is used extensively as a model to study brain development, injury and diseases [16]. It is comprised of a precisely patterned arrangement of six neuronal classes, that include two classes of photoreceptors (rods and cones), three classes of interneurons (horizontal cells, bipolar cells and amacrine cells), ganglion cells that serve as the output neurons and one glial cell type (Müller glia). Although both structure and function of the retina are highly conserved among vertebrates, there are vast differences between species in their ability to regenerate neurons following injury [17]. Mammals have an almost nonexistent capacity for neuronal regeneration. In stark contrast, any lesion that kills retinal neurons in zebrafish leads to complete neuronal regeneration [18–22]. This robust regenerative neurogenesis in the retina is dependent on Müller glia, which act as intrinsic stem cells and give rise to progenitors capable of replenishing each of the six neuronal cell types.
Several paradigms have been used to study neuronal regeneration in the zebrafish retina [23–29]. A photolytic lesion, which results in the selective death of photoreceptors [25,30,31], is widely used and serves as a model of human photoreceptor dystrophies [32]. The death of the photoreceptors stimulates Müller glia to re-enter the cell cycle, divide and give rise to rapidly proliferating progenitors that form radial clusters surrounding the parent Müller glia. These progenitors then migrate to ONL, exit the cell cycle and differentiate into both rod and cone photoreceptors (reviewed in [33]). Cones are regenerated prior to rods, and the first regenerated cones begin to appear at around 4 days post-lesion [34]. The depleted ONL is functionally reconstituted within 15–20 days [35].
The mechanisms leading to the de-differentiation of Müller glia and has been intensively studied. For example, dying photoreceptors signal to Müller glia by synthesizing and secreting TNF-α [36]. Several transcription factors and signaling pathways are then required for Müller glia to re-enter the cell cycle and to sustain proliferation, including Ascl1a [37], Insm1a [37], Stat3 [36], FGF [38], TGF-β [21] and Stil [39].
In zebrafish, there are two midkine paralogs, midkine-a (mdka) and midkine-b (mdkb), which share 68% of amino acid identity [40]. Both midkines are differentially regulated during brain development [40], and both are upregulated in zebrafish during regeneration of multiple tissues and organs, e.g., heart [41], fin [42] and retina [10]. mdka and mdkb were first identified in the retina by an unbiased screen for genes induced by the death of the photoreceptors and during photoreceptor regeneration [43]. The same study showed that during embryonic retinal development, mdka and mdkb have distinct cellular patterns of expression. A subsequent study of Mdka function showed that in retinal progenitors this protein governs cell cycle kinetics. Loss- and gain-of-function slows and accelerates the cell cycle, respectively [44].
The goal of the present study was to establish the patterns of Mdka protein localization during retinal development and photoreceptor regeneration and gain insight into its extracellular trafficking and function during these events. The expression of mdka in Müller glia and photoreceptor progenitors after light-induced lesion [43] and the known roles of Midkine in neural repair, lead us to experimentally test the hypothesis that Mdka has a fundamental function in governing Müller glial-based photoreceptor regeneration. The data show that during embryonic retinal development the antibodies against Mdka labels all mitotically-active cells. As progenitors exit the cell cycle and begin to differentiate, Mdka immunostaining becomes restricted to horizontal cells. Coinciding with this transition, the Mdka antibodies transiently label the inner plexiform layer, and the immunostaining of photoreceptors in the outer nuclear layer first appears. In the adult retina, the Mdka antibodies label horizontal cells, rod photoreceptors and the end feet of Müller glia. For horizontal cells, Mdka immunostaining is present in the cytosol, co-localizes with markers of the Golgi complex and is strongly regulated by the circadian rhythm. During photoreceptor regeneration, the Mdka antibodies label Müller glia and retinal progenitors. Following knockdown of Mdka, there are fewer retinal progenitors and regenerated rods, indicating that Mdka governs proliferation of injury-induced progenitors.
Methods
Animals and retinal lesions
AB strain zebrafish (Danio rerio) were purchased from the Zebrafish International Research Center (ZIRC, University of Oregon, Eugene, Oregon), propagated in-house and housed in recirculating systems at 28.5°C on a 14/10-h light/dark cycle. For the development studies, embryos were collected within 15 minutes of spawning and incubated at 28.5°C on a 14/10-h light/dark cycle. All experimental protocols were approved by the University of Michigan’s committee for the Use and Care of Animals.
Knockdown of Mdka in embryos
Morpholino oligonucleotides (MOs; Gene Tools, LLC, Cowallis, OR, USA) were used to knockdown Mdka in embryos. Previously described and validated protocols for the ATG-targeted mdka MOs were used (see [44]). Embryos were injected at the 1-cell stage with 3ng of Mdka MO (5’-CCGCATTTTGTTTTCTGTGTCGAAA-3’) or mismatch control (Mdka MM, 5’-CCGgATTTTcTTTTCTcTcTgGAAA-3’) diluted in 1x Danieau buffer [45], and analyzed at 48 hours post fertilization (hpf).
Systemic labeling with BrdU or EdU
Proliferating cells were labeled in adult fish with either 5-Bromo-2´-deoxyuridine (BrdU) or 5-ethynyl-2’-deoxyuridine (EdU) by swimming the fish for 24 hours in 5 mM BrdU or 250 μM EdU solution [46,47]. For BrdU staining, sections were incubated in 100°C sodium citrate buffer (10 mM sodium citrate, 0.05% Tween 20, pH 6.0) for 30 minutes to denature DNA and cooled at room temperature for 20 minutes. Sections were then subjected to standard immunolabeling as described below. EdU was visualized using the Click-it EdU kit (Invitrogen, Carlsbad, CA, USA).
Transient transgenesis
To visualize the horizontal cells in the retina, a DNA construct containing the Connexin 55.5 promoter driving expression of Enhanced Green Fluorescent Protein (Cx55.5:EGFP) in a Tol2 destination vector (a gift from Prof. Maarten Kamermans; [48]) was used to create a transient transgenic zebrafish. The DNA construct was co-injected with Tol2 transposon mRNA in 1-cell-stage wild type embryos. EGFP immunolabeling was performed as described below.
Immunohistochemistry
Whole zebrafish (30hpf to 2 months of age) or eye cups (from adults) were fixed by immersion in buffered 4% paraformaldehyde, cryoprotected by infiltration in 20% sucrose in phosphate buffer, and frozen in Optimal Cutting Temperature compound (OCT; e.g., [44]). Ten micron cryosections were mounted on glass slides and processed for immunohistochemistry or in situ hybridization combined with immunohistochemistry.
For immunostaining, sections were incubated in PBS at 37°C for 15 min, washed with PBS containing 0.5% Triton X-100 (PBST), and incubated with a blocking solution containing 5% BSA and 10% sheep serum in 20mM MgCl2 PBS. This was followed by overnight incubation at 4°C with primary antibodies. The next day, sections were washed with PBST and incubated in secondary antibodies for 1 hour at room temperature, washed again with PBST, counterstained with 1:1000 dilution of Hoechst to label nuclei and sealed with mounting media and glass coverslips.
The primary antibodies used were: rabbit anti-Mdka 1:200 (Zymed Laboratories-Invitrogen; see [49]), mouse anti-BrdU 1:100 (BD Biosciences 347580), mouse anti-GM130 1:200 (BD Biosciences 610822; [50]), mouse anti-Glutamine Synthetase 1:200 (Millipore MAB302), mouse anti-GFP 1:200 (Millipore MAB3580), mouse anti-Zpr1 1:200 (anti-Arrestin 3, ZIRC), mouse anti-Zpr3 1:200 (anti-Rhodopsin, ZIRC). Secondary antibodies (Invitrogen) were raised against in either mouse or rabbit antibodies and conjugated to Alexa Fluor 488 and 555 and diluted 1:500. Fluorescence images were captured using a Leica TCS SP5 confocal microscope (Vernon Hills, IL, USA).
In situ hybridization
In situ hybridizations on retinal sections were performed using previously published protocols (e.g., [44,51]). Briefly, sections were hybridized with digoxigenin (DIG)-labeled riboprobes rhodopsin (rho) for rod photoreceptors or phosphodiesterase 6c (pde6c) for cone photoreceptors overnight at 55°C, incubated with an alkaline-phosphatase-conjugated anti-DIG antibody overnight at 4°C and visualized using 4-nitrobluetetrazolium/5-bromo-4-chloro-3-indolyl phosphate (NBT/BCIP) as a substrate. When in situ hybridizations were combined with BrdU immunolabeling, sections were post-fixed in buffered 4% paraformaldehyde for 10 minutes and rinsed for 1 hour in several changes of PBS. Sections were then immunolabeled for BrdU as described above. Bright field and fluorescence images in double-labeled sections were captured with a Leica DM6000 microscope (Vernon Hills, IL, USA).
Western Blot Analysis
Protein samples were obtained by pooling 50 embryo heads in a lysis buffer with protease inhibitors (Complete Mini, Roche, Germany). Proteins were separated in a 12% SDS-PAGE gel and transferred to a nitrocellulose membrane (Sigma, St. Louis, MO, USA). The membrane was blocked in 5% non-fat dry milk in PBS for 2 hours and incubated with rabbit anti-Mdka antibodies (1:1,000) (Zymed Laboratories-Invitrogen, Carlsbad, CA, USA). Blots were rinsed with PBS and incubated with goat horseradish peroxidase- conjugated secondary IgG (1:5,000) for 1 hour. Bound antibodies were visualized using the enhanced chemiluminescence assay (ECL detection system, Amersham Biosciences, Arlington Heights, IL, USA). As loading controls, blots were also incubated with anti-actin (1:1,000, Calbiochem, Germany). Images were captured using the FluorChem E Imaging System (Bio-Techne Minneapolis, MN, USA).
Photolytic lesions, morpholino electroporation and cell counts
To deplete photoreceptors, adult pigmented fish were housed in complete darkness for 24 hours, then immediately exposed to high intensity light (ca. 100,000 lux) from a mercury arc lamp for 30 minutes [25,52]. Fish were then moved to a light box and exposed to constant light (ca. 30.000 lux) for 72 hours [52]. All photolytic lesions were started at the same time of day to avoid any potential influence from circadian rhythms (see S1 Fig.). Days post lesion (dpl) indicate the number days following the onset of the constant light exposure.
At 1 dpl, lissamine-tagged antisense Mdka MOs (see above) were injected (3 mM; 0.5 μl) into the vitreous chamber of one eye and electroporated into dorsal retina using methods and settings described previously [53]. Electroporation of standard control MOs (Gene Tools, LLC) served as the control. At 2 dpl, all animals were housed overnight in a solution containing 5mM BrdU to label proliferating cells. Animals were then sacrificed either at 3 dpl or returned to normal intensity and cyclical lighting and sacrificed at 6 dpl. All animals were sacrificed at the same time of day to avoid the influence of the circadian rhythms. Eyes were excluded from any further analysis if they showed damage from the electroporation, contained incomplete lesion or there was persistence of Mdka immunofluorescence. Two and three independent experiments were performed for animals at 3 and 6 dpl, respectively. For each eye, the average cell count from 3 non-adjacent cross sections spanning 500μm of the central retina was calculated. This value was averaged across 4–7 eyes from each treatment group to obtain a single value.
To count BrdU-labeled cells from retinas at 3 dpl, a confocal z-stack was collected from three non-adjacent sections through the central retina of each eye using a Leica SP5 confocal microscope (Vernon Hills, IL, USA). The number of BrdU-positive cells was counted in a 500 μm length of retina from within the lesion using Imaris Software (Bitplane, South Windsor, CT, USA), using standardized parameters for sensitivity and cell size.
Sections from the 6 dpl group were processed for in situ hybridization for either rhodopsin (rods photoreceptors) or pde6c (cone photoreceptors) and BrdU immunostaining. BrdU-labeled (regenerated) rods or cones were then counted manually in a 500 μm length of retina from within the lesion.
Statistical Analysis
Statistical significance of cell counts between control and experimental retinas was determined with a Student’s t-test using GraphPad Prism 5 (La Jolla, CA, USA). A p-value less than 0.05 was considered a statistically significant difference.
Results
Cellular localization of Mdka during retinal development and in the adult retina
To test the specificity of the Mdka antibodies, detection of Mdka and its absence in mdka morphants was confirmed using Western blots (see also [44]), Then the antibodies were used for immunofluorescence on retinal cryosections taken from control and morphant embryos. Western blots (S2A Fig.) show that the Mdka antibodies recognize a major band at around 13 kDa, and this band is absent in embryos injected with the mdka MOs. The source of the minor band (≈16 kDa) in the Western blot is not clear. However, the presence of this band after Mdka knockdown (S2B Fig.) suggests that the band is likely a protein that is not Mdka. In tissue sections from 48hpf embryos (S2B Fig.), the mdka morphants show the microphthalmia characteristic of Mdka knockdown [44] and a complete absence of the immunofluorescence signal. This is in marked contrast to the immunostaining for uninjected and control embryos (taken from the same clutch), which show the pattern of immunostaining characteristic of the 48hpf retina (see below). These data demonstrate that the polyclonal antibodies used here to characterize protein localization in retinal sections are specific for Mdka.
Next, to gain insights into the cellular localization of Mdka synthesis and potential location of the secreted protein, comparisons were made between in situ hybridizations using mdka probes and Mdka immunostaining. The cellular pattern of mdka expression changes markedly throughout development in the zebrafish retina ([43]; Fig. 1A-F). At 30 hpf (Fig. 1A), prior to significant neuronal differentiation, mdka is expressed throughout the neuroepithelium, with higher expression at the retinal margin, presaging the circumferential marginal zone (CMZ) of the mature retina [54]. At 48 hpf (Fig. 1B), as progenitors withdraw from the cell cycle and differentiation commences centrally, mdka is down-regulated in newly postmitotic cells, while expression persists in the still dividing cells in the periphery. The center-to-periphery pattern of neuronal differentiation continues, such that by 120 hpf (Fig. 1C), when the retina is fully differentiated, mdka expression is restricted to the CMZ and horizontal cells (see also [43]). This cellular pattern of expression remains invariant during subsequent growth and into adulthood (Fig. 1D-F).
Fig 1. mdka expression and Mdka immunostaining during retinal development and in adults.

In situ hybridization showing mdka expression at stages between 30hpf and adults (A-F). Mdka immunostaining at stages corresponding to the in situ hybridizations are also illustrated (G-H and images merged DAPI counterstain below). At 30 hpf, Mdka immunofluorescence in strongest at the basal surface of the retina (panel G, arrows). At 48hpf, Mdka antibodies transiently label the nascent inner plexiform layer (panel H, arrows). At 120hpf, when differentiated photoreceptors are present, Mdka immunostaining is present in horizontal cells (arrows) and the outer nuclear layer (brackets). Though tested numerous times, the level of Mdka immunofluorescence dramatically decreases between 7–10 dpf (panel J, J’). Note that at this time point horizontal cells express mdka (panel D). In the juvenile retina, Mdka immunostaining returns and both horizontal cells (arrows, panel K, K’) and the overlying rod photoreceptors in the ONL are intensely stained. This juvenile pattern of immunostaining is present in adults, but the overall intensity of the immunostaining is reduced, and the immunostaining of horizontal cells is consolidated to a small immunofluorescent plaque lying adjacent to each DAPI-stained nucleus (arrows, panel L’). hpf, hours post fertilizations; dpf, days post fertilization; mo, months; ONL, outer nuclear layer; INL, inner nuclear layer; IPL, inner plexiform layer; CMZ, circumferential marginal zone. Scale bar in panel J’ equals 20 μm and corresponds to images in columns A-D and G-J. Scale bar in panel L’ equals 10 μm and corresponds to images in panels E, F, K, K’, L and L’.
Similar to the developmental pattern of mdka expression, the developmental pattern of Mdka immunolabeling is also temporally and spatially dynamic. At 30 hpf (Fig. 1G, G’), the Mdka immunolabel is present throughout the retinal neuroepithelium, though the immunostaining is concentrated at the basal surface. At 48hpf (Fig. 1H, H’), there is an apparent reduction of Mdka immunostaining of dividing cells and at the basal surface, and the Mdka antibodies transiently labels the nascent inner plexiform layer. At 120 hpf (Fig. 1I, I’), reflecting the cellular pattern of gene expression, Mdka immunostaining is present in horizontal cells (arrows). However, in contrast to the mdka expression, which is restricted to horizontal cells, at 120hpf, Mdka antibodies also label photoreceptors (brackets). Unexpectedly, though repeatedly tested, between 7–10 days post fertilization (Fig. 1J, J’), Mdka immunostaining is undetectable in retinal sections. This transient absence of immunostaining is restored by 20 days post fertilization (Fig. 1K, K’), when high levels of Mdka immunostaining is present both among horizontal cells and the overlying outer nuclear layer, where the immunostaining is restricted to rod photoreceptors (S3 Fig.). In the adult retina (Fig. 1L, L’), the overall levels of Mdka immunostaining are diminished, and Mdka immunostaining in horizontal cells appears as a small plaque of fluorescence on the apical aspect of each nucleus. The intensity of the Mdka immunofluorescence surrounding each rod photoreceptor is noticeably diminished in the adult retina as well when compared to juveniles.
The immunostaining alone of the horizontal cells was insufficient to establish if the Mdka immunolabel was localized to an intracellular compartment or concentrated on the cell surface. To address this, we generated transient transgenic animals that expressed enhanced green fluorescent protein (EGFP) under the control of the promoter of connexin 55.5, a gene expressed exclusively by horizontal cells in the teleost retina [48]. Retinal sections from transgenic animals were immunostained for both EGFP, which diffusely fills the cytoplasm, and Mdka. These experiments show co-localization of the GFP and Mdka immunofluorescence, demonstrating that that in horizontal cells the Mdka immunolabel is cytosolic (Fig. 2A). Based on the size, shape and proximity to the nucleus of the intracellular plaque of Mdka immunostaining in the adult retina, we hypothesized that the Mdka immunostaining colocalizes with the Golgi complex. Double immunostaining for Mdka and the Golgi marker, GM130, showed that in each horizontal cell, the Mdka immunostaining colocalizes with markers of the Golgi complex (Fig. 2B).
Fig 2. Mdka immunostaining of horizontal cells is intracellular and colocalizes with markers of the Golgi complex.
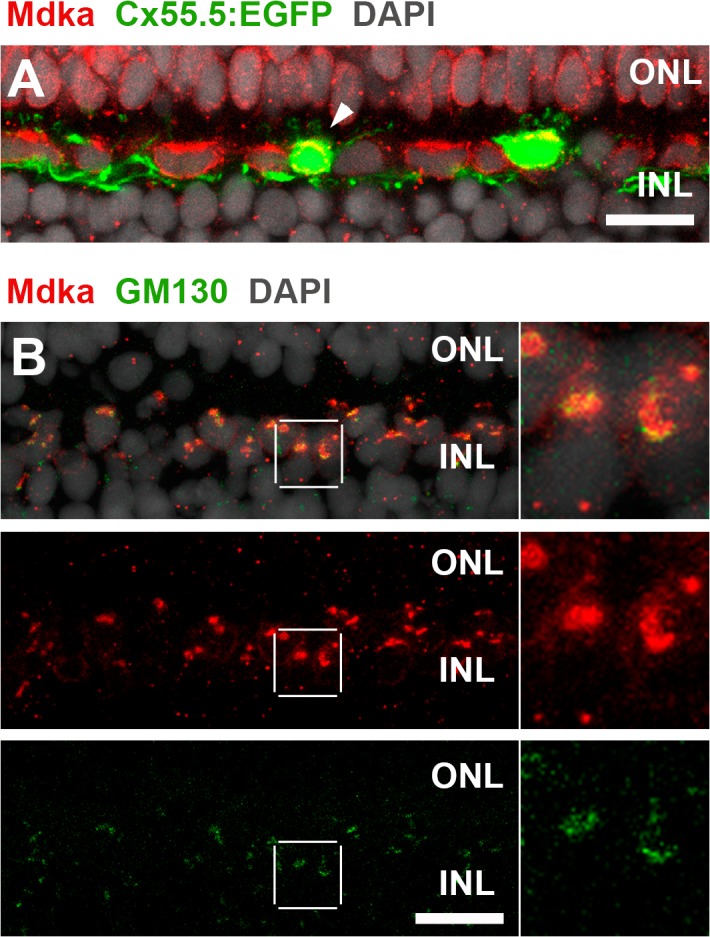
In fish injected with the Cx55.5:EGFP construct, immunolabels for both Mdka and EGFP colocalize to the cytoplasm of horizontal cells (arrowhead, panel A). In the adult retina, Mdka immunostaining colocalizes with the antibody marker of the Golgi complex (column B; and insets). ONL: outer nuclear layer; INL: inner nuclear layer. Scale bars equal 10 μm.
Finally, also in the adult retina, Mdka protein is present in the end feet of the Müller glia (Fig. 3D-F; S4 Fig.). Double immunostaining for Mdka and glutamine synthetase (GS), which is specific for Müller glia [23,55,56], shows that the thin ribbon of Mdka immunostaining at the basal surface of the retina colocalizes with GS immunostaining that marks the end feet of Müller glia (Fig. 4A). Further, there was no systematic change in the thickness of this label, from retinal periphery to the optic nerve head, allowing us to conclude that the antibodies do not stain ganglion cell axons in the optic fiber layer (data not shown).
Fig 3. Circadian regulation of Mdka immunostaining in horizontal cells.

At the onset of the subjective day, Mdka immunostaining of horizontal cells is maximum (column A), decreases throughout the subjective day (column B) and reaches a minimum during the subjective night (column C). In contrast, there is no corresponding substantial change in the intensity of immunostaining within the endfeet of the Müller glia (D-F). ONL: outer nuclear layer; INL: inner nuclear layer; GCL: ganglion cell layer; NFL: nerve fiber layer. Scale bar = 10 μm.
Fig 4. Mdka protein localization following photoreceptor ablation.

In unlesioned retinas, Mdka immunostaining is localized to the horizontal cells and endfeet of glutamine synthetase (GS)-positive Müller glia (row A). At 4 dpl, Mdka antibodies label the radial processes of Müller glia (row B, arrows). Note the increased Mdka immunostaining in the endfeet of the Müller glia in lesioned retinas (cf. rows A and B). Also at 4 dpl, Mdka immunostaining is localized to each of the EdU-positive nuclei in both the INL and ONL (row C, arrowheads). ONL: outer nuclear layer; INL: inner nuclear layer; dpl: days post lesion. Scale bars = 25 μm.
Mdka synthesis is regulated by circadian rhythms
It was shown previously that in the retina of adult zebrafish, mdka expression is strongly regulated by the circadian clock. mdka levels are highest early in the subjective day, then decrease during the day and into the initial hours of the subjective night [49]. To determine if Mdka protein synthesis and/or its putative secretion are regulated similarly, we repeated the experimental rearing conditions used previously and immunostained retinal sections with the Mdka antibodies at various points during the circadian cycle. This experiment showed that Mdka immunostaining follows a circadian pattern that reflects the mRNA (Fig. 3A-C). Mdka immunofluorescence in horizontal cells and the overlying rod photoreceptors is highest at the onset of the subjective day and decreases into the early hours of the subjective night. In contrast to this circadian pattern, Mdka protein levels remain unchanged (Fig. 3D-F).
Mdka regulates injury-induced cell proliferation and the regeneration of rod photoreceptors
After a photolytic lesion, mdka is expressed in Müller glia and the injury-induced photoreceptor progenitors, suggesting Mdka has a functional role during photoreceptor regeneration [43]. As a prelude to testing the function of Mdka, we established the cellular pattern of Mdka immunostaining in the retinas of animals at 4 dpl, which corresponds to the peak of the injury-induced cell proliferation following the photolytic death of photoreceptors [57]. At this time point, Mdka immunostaining is distributed along the radial processes of the Müller glia throughout the thickness of the retina. Particularly dense Mdka immunostaining is present in the glial endfeet at the basal surface of the retina (Fig. 4B). Mdka immunolabel also surrounds each proliferative, EdU-positive nucleus in the ONL and INL (Fig. 4C).
To test the functional role of Mdka during photoreceptor regeneration, Mdka knockdown was performed by electroporating ATG-targeted or control MOs at 1dpl (S1 Fig.). The effectiveness of the knockdown was validated by the absence of Mdka immunofluorescence in sections adjacent to those used for cell counts (S4 Fig.). In control retinas, the BrdU immunostaining reflected the well-described pattern of cell proliferation in response to photoreceptor death (e.g., [57]). Beneath the depleted photoreceptor layer, progenitors form radial clusters within the INL, surrounding each parent Müller glial stem cell, and accumulate within the nascent ONL (Fig. 5A). Following knockdown of Mdka, the average number of BrdU-positive cells per 500 μm was significantly less in underlying the lesion (239.0 ± 9.9 vs. 154.7 ± 17.7; p-value <0.01), and the proliferating cells appeared more scattered among the retinal layers (Fig. 5B,C).
Fig 5. Mdka knockdown reduces the number of injury-induced progenitors.

Progenitors were labeled with BrdU in the retinas of control (panel A) and experimental fish (panel B) at 3 dpl. Note the radial clusters of progenitors in the control retinas (arrows in A) and their relative absence in experimental retinas (panel B). BrdU-positive cells were counted, and this is graphically represented in panel C. INL: inner nuclear layer; GCL: ganglion cell layer; BrdU: Bromodeoxyuridine. Scale bar = 25 μm. ** p<0.01.
The effect of Mdka knockdown on the injury-induced proliferation led us to hypothesize that Mdka may also impact the regeneration of rod and/or cone photoreceptors. For retinas at 6 dpl, in situ hybridizations specific for rod or cone genes were used to separately label these cells, and regenerated rods and cones were quantified. BrdU immunostaining was combined with the in situ labels to exclude photoreceptors that may have survived the photolytic lesions and to insure that only regenerated rods and cones were counted. Mdka knockdown was confirmed by immunostaining sample sections from each eye (data not shown). Compared to control retinas, Mdka knockdown resulted in statistically less regenerated rods in experimental retinas than controls (22.17 ± 5.5 vs. 9.631 ± 1.3; Fig. 6A). In contrast, no significant differences were found in the number of regenerated cones (Fig. 6B).
Fig 6. Knockdown of Mdka reduces the number of regenerated rod photoreceptors.
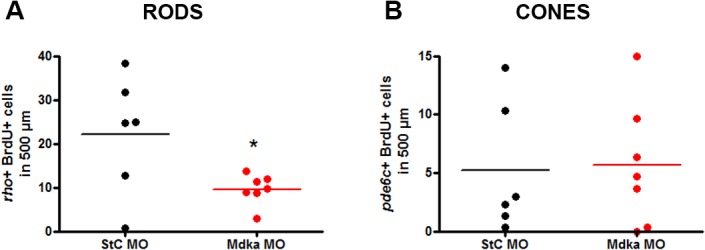
Graphs in panels A and B represent the number of regenerated rods and cones, respectively, counted within the photolytic lesions of experimental and control fish. Bromodeoxyuridine; rho: rhodopsin; pde6c: phosphodiesterase6c. *p-value < 0.05.
Discussion
Developmental neurogenesis in the zebrafish retina is amazingly rapid [58], and, as in all vertebrates, is controlled by complex signaling events [59]. The zebrafish retina is formed from a pool of mitotic progenitors that begin exiting the cell cycle around 28–32 hpf [60,61] and thereafter continue to exit the cell cycle in a complex spatiotemporal pattern [62]. The cellular expression of mdka reflects this pattern of neurogenesis. mdka is expressed in all mitotic progenitors and downregulated as cells exit the cell cycle [43]. Upon retinal differentiation, mdka is then expressed in a single type of mature neuron, the horizontal cell, which resides in a monolayer between the inner retinal neurons, the outer plexiform layer and photoreceptors. Interestingly, the expression of mdka in horizontal cells is strongly regulated by the circadian rhythm [49]. Loss- and gain-of-function studies demonstrated that during retinal neurogenesis, Mdka governs cell cycle kinetics [44], perhaps, functioning in an autocrine manner. In the present study, we use immunofluorescence with polyclonal antibodies specific for Mdka to document immunolocalization in both the developing and adult retinas and during photoreceptor regeneration. In the developing retina, cellular pattern of Mdka immunostaining varies with developmental stage, reflecting the dynamic spatiotemporal pattern of the gene expression. Antibodies against Mdka stain progenitors in the embryonic retina and horizontal cells in adults. However, in both embryonic and adult retinas, the immunostaining only partially overlaps with cellular expression of mdka, leading us to infer that Mdka is secreted and may have paracrine functions as well. Finally, we also tested the function of Mdka during photoreceptor regeneration, and these experiments showed that Mdka governs injury-induced cell proliferation and this leads to a deficit in the regeneration of rod photoreceptors.
The mismatch in the cellular pattern of immunostaining and gene expression suggests that Mdka may play multiple roles. In the early neuroepithelium, between 24 and 36hpf, the Mdka immunostaining labels each retinal progenitor, but the densest staining is at the basal surface of the retina. Apicobasal polarity of growth factors in the retinal neuroepithelium is a well-documented mechanism for governing interkinetic nuclear migration, cell proliferation and cell fate determination [63,64]. Following knockdown of Mdka, retinal progenitors undergo interkinetic nuclear migration, but their transit from basal to apical surface is slowed and the proportion of cells that enter mitosis is diminished [44]. We speculate that the accumulation of Mdka at the basal surface of the retinal neuroepithelium contributes to apicobasal polarity and functions to govern phases of the cell cycle among progenitors in proximity to the basal surface (see also [65]). At 48hpf, when rapid cellular differentiation is happening, Mdka immunostaining at the basal surface is lost, and the immunolabel transiently accumulates in the nascent inner plexiform layer. The immunostaining of this synaptic layer is the first evidence that Mdka may be secreted and trafficked to retinal compartments separate from the mdka-expressing cells. In the central nervous system, Mdka plays a role promoting neurite outgrowth (reviewed in [15]; see also, [66]). Similarly, the apparently transient localization of Mdka in the developing inner plexiform layer may play a role in directing the outgrowth of immature dendritic processes and/or establishing the initial synaptic contacts made there.
The early, spatially dynamic pattern of Mdka immunostaining stabilizes as the cellular differentiation of the retina becomes complete. At and after 120 hpf, the Mdka antibodies label horizontal cells, corresponding to the unique cellular pattern of mdka expression. The Mdka immunolabel in horizontal cells is initially diffuse within the cytoplasm, but in adults, Mdka immunostaining is consolidated to a small plaque, which precisely colocalizes with markers of the Golgi complex. The Mdka antibodies also label rod photoreceptors in the outer nuclear layer. Numerous in situ hybridizations over several years and multiple investigators [43,44] indicate that rods do not express mdka. The colocalization of Mdka immunostaining with the Golgi complex of horizontal cells and the immunostaining of rods suggests that Mdka is synthesized by the horizontal cells, secreted in a circadian manner and sequestered by overlying rod photoreceptors. We do not know the molecular mechanisms by which Mdka associates with rod photoreceptors nor the function it mediates, however, the data provided here is, to the best of our knowledge, the first evidence of a bona fide growth factor released by horizontal cells into the outer nuclear layer.
In the adult retina, the normal pattern of Mdka immunostaining changes dramatically following the death of the photoreceptors (see also [43]). Mdka immunostaining in the horizontal cells beneath the depleted ONL is lost, and Müller glia and their basal endfeet are strongly labeled. Mdka antibodies also label the injury-induced retinal progenitors. These observations, and our previous data [44], led us to hypothesize that Mdka may govern aspects of photoreceptor regeneration. Using specific morpholinos to knockdown Mdka, we tested this hypothesis. Compared to controls, following Mdka knockdown, there were less proliferating retinal progenitors. The electroporation was performed near the time point when Müller glia enter the cell cycle following the death of photoreceptors [57], and, therefore, the loss of Mdka may either limit the ability of Müller glia to enter the cell cycle or, via inheritance of the morpholinos from parent Müller glia, inhibit proliferation of the Müller glia-derived progenitors. The reduced number progenitors following the knockdown of Midkine was accompanied by the absence of the characteristic neurogenic clusters that form around each Müller glia, and the progenitors that were present were dispersed throughout the retina. Further, following Mdka knockdown, there was a deficit in the regeneration of rod photoreceptors. We interpret this to show that decreased Mdka, similar to what was observed in the developing retina [44], slows the cell cycle, leading to a reduced number of progenitors and, by extension, a deficit in the number of regenerated rod photoreceptors. This suggests that during photoreceptor regeneration, Mdka functions to govern cell cycle kinetics, similar to what was demonstrated for this protein in the developing retina [44].
The results presented here are consistent with multiple studies implicating Midkine in the repair of the mammalian nervous system (for a review see [15]). For example, Midkine promotes functional recovery after spinal cord injury in rats [67], and in Mdk knockout mice, the time course of recovery from cerebral ischemia or peripheral nerve injury is delayed [13,68]. Similarly, Mdk knockout mice show early preclinical features of Parkinson's disease [69] and a higher vulnerability to the neurotoxic effects induced by amphetamine [70]. There is a rapidly growing body of knowledge about molecular mechanisms that underlie neuronal regeneration in the zebrafish retina (reviewed in [33,71]), and our data demonstrate that Mdka and the signaling pathways it governs can be added mechanisms that govern photoreceptor genesis. The role of Mdka in this stem cell-based regeneration opens the possibility of leveraging the Midkine signaling pathways as a potential therapeutic approach to treating degenerative disease affecting tissues within the mammalian central nervous system.
Supporting Information
Adult zebrafish were dark-adapted for 24 h, and photolytic lesions consisted of a 30 min exposure to high-intensity light, by three days of constant bright light. At one day post-lesion (dpl), morpholinos were injected into the vitreous cavity and electroporated into the right eye of experimental and control animals. At two dpl, fish were immersed in BrdU solution for 24 hrs. Animals were then sacrificed at three dpl for quantification of BrdU+ cells, or returned to normal light conditions and sacrificed at six dpl for quantification of regenerated rod and cone photoreceptors. BrdU: Bromodeoxyuridine; rho: rhodopsin; pde6c: phosphodiesterase 6c.
(TIF)
Specificity of the anti-Mdka antibodies was determined by the selective loss of Mdka immunolabeling in Western blots (panel A) and retinal sections (panel B) from morphant embryos at 48 hpf. (A) For Western blots, β-actin served as the loading control. (B) Mdka MM—embryos injected with control morpholinos; Mdka MO—embryos injected with ATG-targeted morpholinos. Scale bar = 20 μm.
(TIF)
Sections were stained with antibodies against Mdka and the rod marker Zpr3 (panel A) or the red-green cones marker Zpr1 (panel B). Note the Mdka immunostaining of the rod photoreceptor nuclei and co-localizations with rod inner segments (arrows, panel A). Zpr1 labels the cell surface of red-green cones and this marker does not colocalize with the Mdka immunostaining (panel B) ONL: outer nuclear layer; INL: inner nuclear layer. Scale bar equals 10 μm.
(TIF)
Sections from retinas electroporated with control morpholinos show robust Mdka immunostaining (column A). In contrast, retinas electroporated with ATG-targeted morpholinos show a clear knockdown of Mdka immunostaining (column B). INL: inner nuclear layer; GCL: ganglion cell layer. Scale bar equals 50μm.
(TIF)
Acknowledgments
The authors thank Laura Kakuk-Atkins and Dilip Pawar for technical assistance and Prof. Maarten Kamermans, who provided the Cx55.5:EGFP construct. The authors would also like to thank Dimitris Grivas and Dr. José Luis De La Pompa Mínguez (CNIC, Madrid) for the immunohistochemistry protocol used to detect Mdka in zebrafish tissue.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by the National Institutes of Health (National Eye Institute) R01 EY07060. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Winkler C, Yao S. The midkine family of growth factors: Diverse roles in nervous system formation and maintenance. Br J Pharmacol. 2013. [DOI] [PMC free article] [PubMed]
- 2. Qi M, Ikematsu S, Ichihara-Tanaka K, Sakuma S, Muramatsu T, Kadomatsu K. Midkine rescues Wilms' tumor cells from cisplatin-induced apoptosis: regulation of Bcl-2 expression by Midkine. Journal of biochemistry. 2000; 127: 269–277. [DOI] [PubMed] [Google Scholar]
- 3. Taravini IRE, Ferrario JE, Delbe J, Ginestet L, Debeir T, Courty J, et al. Immunodetection of heparin-binding growth associated molecule (pleiotrophin) in striatal interneurons. Brain research. 2005; 1066: 196–200. [DOI] [PubMed] [Google Scholar]
- 4. Bloch B, Normand E, Kovesdi I, Böhlen P. Expression of the HBNF (heparin-binding neurite-promoting factor) gene in the brain of fetal, neonatal and adult rat: an in situ hybridization study. Brain Res Dev Brain Res. 1992; 70: 267–278. [DOI] [PubMed] [Google Scholar]
- 5. Vanderwinden JM, Mailleux P, Schiffmann SN, Vanderhaeghen JJ. Cellular distribution of the new growth factor pleiotrophin (HB-GAM) mRNA in developing and adult rat tissues. Anat Embryol. 1992; 186: 387–406. [DOI] [PubMed] [Google Scholar]
- 6. Hoggard N, Cruickshank M, Moar K-M, Bashir S, Mayer C-D. Using gene expression to predict differences in the secretome of human omental vs. subcutaneous adipose tissue. Obesity (Silver Spring). 2012; 20: 1158–1167. 10.1038/oby.2012.14 [DOI] [PubMed] [Google Scholar]
- 7. Marchionini DM, Lehrmann E, Chu Y, He B, Sortwell CE, Becker KG, et al. Role of heparin binding growth factors in nigrostriatal dopamine system development and Parkinson's disease. Brain research. 2007; 1147: 77–88. [DOI] [PubMed] [Google Scholar]
- 8. Ezquerra L, Pérez-García C, Garrido E, Díez-Fernández C, Deuel TF, Alguacil LF, et al. Morphine and yohimbine regulate midkine gene expression in the rat hippocampus. Eur J Pharmacol. 2007; 557: 147–150. [DOI] [PubMed] [Google Scholar]
- 9. Flatscher-Bader T, Wilce PA. Impact of alcohol abuse on protein expression of midkine and excitatory amino acid transporter 1 in the human prefrontal cortex. Alcohol Clin Exp Res. 2008; 32: 1849–1858. 10.1111/j.1530-0277.2008.00754.x [DOI] [PubMed] [Google Scholar]
- 10. Craig SEL, Calinescu A-A, Hitchcock PF. Identification of the molecular signatures integral to regenerating photoreceptors in the retina of the zebra fish. J Ocul Biol Dis Infor. 2008; 1: 73–84. 10.1007/s12177-008-9011-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Miyashiro M, Kadomatsu K, Ogata N, Yamamoto C, Takahashi K, Uyama M, et al. Midkine expression in transient retinal ischemia in the rat. Curr Eye Res. 1998; 17: 9–13. [DOI] [PubMed] [Google Scholar]
- 12. Muramatsu T. Midkine, a heparin-binding cytokine with multiple roles in development, repair and diseases. Proceedings of the Japan Academy Series B, Physical and biological sciences. 2010; 86: 410–425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Sakakima H, Yoshida Y, Yamazaki Y, Matsuda F, Ikutomo M, Ijiri K, et al. Disruption of the midkine gene (Mdk) delays degeneration and regeneration in injured peripheral nerve. J Neurosci Res. 2009; 87: 2908–2915. 10.1002/jnr.22127 [DOI] [PubMed] [Google Scholar]
- 14. Herradón G, Pérez-García C. Targeting midkine and pleiotrophin signalling pathways in addiction and neurodegenerative disorders: recent progress and perspectives. Br J Pharmacol. 2014; 171: 837–848. 10.1111/bph.12312 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Yoshida Y, Sakakima H, Matsuda F, Ikutomo M. Midkine in repair of the injured nervous system. Br J Pharmacol. 2014; 171: 924–930. 10.1111/bph.12497 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. London A, Benhar I, Schwartz M. The retina as a window to the brain-from eye research to CNS disorders. Nat Rev Neurol. 2013; 9: 44–53. 10.1038/nrneurol.2012.227 [DOI] [PubMed] [Google Scholar]
- 17. Karl MO, Reh TA. Regenerative medicine for retinal diseases: activating endogenous repair mechanisms. Trends Mol Med. 2010; 16: 193–202. 10.1016/j.molmed.2010.02.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Hitchcock PF, Raymond PA. The teleost retina as a model for developmental and regeneration biology. Zebrafish. 2004; 1: 257–271. 10.1089/zeb.2004.1.257 [DOI] [PubMed] [Google Scholar]
- 19. Thummel R, Kassen SC, Enright JM, Nelson CM, Montgomery JE, Hyde DR. Characterization of Müller glia and neuronal progenitors during adult zebrafish retinal regeneration. Experimental eye research. 2008; 87: 433–444. 10.1016/j.exer.2008.07.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Craig SEL, Thummel R, Ahmed H, Vasta GR, Hyde DR, Hitchcock PF. The zebrafish galectin Drgal1-l2 is expressed by proliferating Müller glia and photoreceptor progenitors and regulates the regeneration of rod photoreceptors. Invest Ophthalmol Vis Sci. 2010; 51: 3244–3252. 10.1167/iovs.09-4879 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Lenkowski JR, Qin Z, Sifuentes CJ, Thummel R, Soto CM, Moens CB, et al. Retinal regeneration in adult zebrafish requires regulation of TGFbeta signaling. Glia. 2013; 61: 1687–1697. 10.1002/glia.22549 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Gemberling M, Bailey TJ, Hyde DR, Poss KD. The zebrafish as a model for complex tissue regeneration. Trends in genetics: TIG. 2013; 1–10. [DOI] [PMC free article] [PubMed]
- 23. Fimbel SM, Montgomery JE, Burket CT, Hyde DR. Regeneration of inner retinal neurons after intravitreal injection of ouabain in zebrafish. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2007; 27: 1712–1724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Ramachandran R, Zhao X-F, Goldman D. Ascl1a/Dkk/beta-catenin signaling pathway is necessary and glycogen synthase kinase-3beta inhibition is sufficient for zebrafish retina regeneration. Proc Natl Acad Sci USA. 2011; 108: 15858–15863. 10.1073/pnas.1107220108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Taylor S, Chen J, Luo J, Hitchcock P. Light-induced photoreceptor degeneration in the retina of the zebrafish. Methods in molecular biology. 2012; 884: 247–254. 10.1007/978-1-61779-848-1_17 [DOI] [PubMed] [Google Scholar]
- 26. Montgomery JE, Parsons MJ, Hyde DR. A novel model of retinal ablation demonstrates that the extent of rod cell death regulates the origin of the regenerated zebrafish rod photoreceptors. The Journal of comparative neurology. 2010; 518: 800–814. 10.1002/cne.22243 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Morris AC, Schroeter EH, Bilotta J, Wong ROL, Fadool JM. Cone survival despite rod degeneration in XOPS-mCFP transgenic zebrafish. Invest Ophthalmol Vis Sci. 2005; 46: 4762–4771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Mathias JR, Zhang Z, Saxena MT, Mumm JS. Enhanced cell-specific ablation in zebrafish using a triple mutant of Escherichia coli nitroreductase. Zebrafish. 2014; 11: 85–97. 10.1089/zeb.2013.0937 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Fraser B, DuVal MG, Wang H, Allison WT. Regeneration of cone photoreceptors when cell ablation is primarily restricted to a particular cone subtype. PLoS ONE. 2013; 8: e55410 10.1371/journal.pone.0055410 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Vihtelic TS, Soverly JE, Kassen SC, Hyde DR. Retinal regional differences in photoreceptor cell death and regeneration in light-lesioned albino zebrafish. Experimental eye research. 2006; 82: 558–575. [DOI] [PubMed] [Google Scholar]
- 31. Thomas JL, Nelson CM, Luo X, Hyde DR, Thummel R. Characterization of multiple light damage paradigms reveals regional differences in photoreceptor loss. Experimental eye research. 2012; 97: 105–116. 10.1016/j.exer.2012.02.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Bibliowicz J, Tittle RK, Gross JM. Toward a better understanding of human eye disease insights from the zebrafish, Danio rerio. Prog Mol Biol Transl Sci. 2011; 100: 287–330. 10.1016/B978-0-12-384878-9.00007-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lenkowski JR, Raymond PA. Müller glia: Stem cells for generation and regeneration of retinal neurons in teleost fish. Prog Retin Eye Res. 2014. [DOI] [PMC free article] [PubMed]
- 34. Bernardos RL, Barthel LK, Meyers JR, Raymond PA. Late-stage neuronal progenitors in the retina are radial Müller glia that function as retinal stem cells. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2007; 27: 7028–7040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Weber A, Hochmann S, Cimalla P, Gärtner M, Kuscha V, Hans S, et al. Characterization of light lesion paradigms and optical coherence tomography as tools to study adult retina regeneration in zebrafish. PLoS ONE. 2013; 8: e80483 10.1371/journal.pone.0080483 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Nelson CM, Gorsuch RA, Bailey TJ, Ackerman KM, Kassen SC, Hyde DR. Stat3 defines three populations of Müller glia and is required for initiating maximal müller glia proliferation in the regenerating zebrafish retina. The Journal of comparative neurology. 2012; 520: 4294–4311. 10.1002/cne.23213 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Ramachandran R, Zhao X-F, Goldman D. Insm1a-mediated gene repression is essential for the formation and differentiation of Müller glia-derived progenitors in the injured retina. Nat Cell Biol. 2012; 14: 1013–1023. 10.1038/ncb2586 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Qin Z, Kidd AR, Thomas JL, Poss KD, Hyde DR, Raymond PA, et al. FGF signaling regulates rod photoreceptor cell maintenance and regeneration in zebrafish. Experimental eye research. 2011; 93: 726–734. 10.1016/j.exer.2011.09.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sun L, Li P, Carr AL, Gorsuch R, Yarka C, Li J, et al. Transcription of the SCL/TAL1 interrupting locus (Stil) is required for cell proliferation in adult zebrafish retinas. The Journal of biological chemistry. 2014. [DOI] [PMC free article] [PubMed]
- 40. Winkler C, Schafer M, Duschl J, Schartl M, Volff JN. Functional divergence of two zebrafish midkine growth factors following fish-specific gene duplication. Genome research. 2003; 13: 1067–1081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Lien C-L, Schebesta M, Makino S, Weber GJ, Keating MT. Gene expression analysis of zebrafish heart regeneration. PLoS Biol. 2006; 4: e260 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Schebesta M, Lien C-L, Engel FB, Keating MT. Transcriptional profiling of caudal fin regeneration in zebrafish. ScientificWorldJournal. 2006; 6 Suppl 1: 38–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Calinescu A-A, Vihtelic TS, Hyde DR, Hitchcock PF. Cellular expression of midkine-a and midkine-b during retinal development and photoreceptor regeneration in zebrafish. The Journal of comparative neurology. 2009; 514: 1–10. 10.1002/cne.21999 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Luo J, Uribe RA, Hayton S, Calinescu AA, Gross JM, Hitchcock PF. Midkine-A functions upstream of Id2a to regulate cell cycle kinetics in the developing vertebrate retina. Neural development. 2012; 7: 1–16. 10.1186/1749-8104-7-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Nasevicius A, Ekker SC. Effective targeted gene “knockdown” in zebrafish. Nat Genet. 2000; 26: 216–220. [DOI] [PubMed] [Google Scholar]
- 46. Otteson DC, Cirenza PF, Hitchcock PF. Persistent neurogenesis in the teleost retina: evidence for regulation by the growth-hormone/insulin-like growth factor-I axis. Mechanisms of development. 2002; 117: 137–149. [DOI] [PubMed] [Google Scholar]
- 47. Nagashima M, Barthel LK, Raymond PA. A self-renewing division of zebrafish Müller glial cells generates neuronal progenitors that require N-cadherin to regenerate retinal neurons. Development. 2013; 140: 4510–4521. 10.1242/dev.090738 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Shields CR, Klooster J, Claassen Y, Ul-Hussain M, Zoidl G, Dermietzel R, et al. Retinal horizontal cell-specific promoter activity and protein expression of zebrafish connexin 52.6 and connexin 55.5. The Journal of comparative neurology. 2007; 501: 765–779. [DOI] [PubMed] [Google Scholar]
- 49. Calinescu A-A, Raymond PA, Hitchcock PF. Midkine expression is regulated by the circadian clock in the retina of the zebrafish. Visual neuroscience. 2009; 26: 495–501. 10.1017/S0952523809990204 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Park J-Y, Ikeda H, Ikenaga T, Ono F. Acetylcholine receptors enable the transport of rapsyn from the Golgi complex to the plasma membrane. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2012; 32: 7356–7363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Ochocinska MJ, Hitchcock PF. Dynamic expression of the basic helix-loop-helix transcription factor neuroD in the rod and cone photoreceptor lineages in the retina of the embryonic and larval zebrafish. The Journal of comparative neurology. 2007; 501: 1–12. [DOI] [PubMed] [Google Scholar]
- 52.Thomas JL, Thummel R. A novel light damage paradigm for use in retinal regeneration studies in adult zebrafish. J Vis Exp. 2013; e51017. [DOI] [PMC free article] [PubMed]
- 53.Thummel R, Bailey TJ, Hyde DR. In vivo electroporation of morpholinos into the adult zebrafish retina. J Vis Exp. 2011; e3603. [DOI] [PMC free article] [PubMed]
- 54. Raymond PA, Barthel LK, Bernardos RL, Perkowski JJ. Molecular characterization of retinal stem cells and their niches in adult zebrafish. BMC Dev Biol. 2006; 6: 36 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Linser P, Moscona AA. Induction of glutamine synthetase in embryonic neural retina: localization in Müller fibers and dependence on cell interactions. Proc Natl Acad Sci USA. 1979; 76: 6476–6480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Zhao X-F, Wan J, Powell C, Ramachandran R, Myers MG, Goldman D. Leptin and IL-6 family cytokines synergize to stimulate müller glia reprogramming and retina regeneration. Cell Rep. 2014; 9: 272–284. 10.1016/j.celrep.2014.08.047 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Vihtelic TS, Hyde DR. Light-induced rod and cone cell death and regeneration in the adult albino zebrafish (Danio rerio) retina. J Neurobiol. 2000; 44: 289–307. [DOI] [PubMed] [Google Scholar]
- 58. Stenkamp DL. Neurogenesis in the fish retina. Int Rev Cytol. 2007; 259: 173–224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Agathocleous M, Harris WA. From progenitors to differentiated cells in the vertebrate retina. Annual review of cell and developmental biology. 2009; 25: 45–69. 10.1146/annurev.cellbio.042308.113259 [DOI] [PubMed] [Google Scholar]
- 60. Schmitt EA, Dowling JE. Comparison of topographical patterns of ganglion and photoreceptor cell differentiation in the retina of the zebrafish, Danio rerio. The Journal of comparative neurology. 1996; 371: 222–234. [DOI] [PubMed] [Google Scholar]
- 61. Hu M, Easter SS. Retinal neurogenesis: the formation of the initial central patch of postmitotic cells. Developmental biology. 1999; 207: 309–321. [DOI] [PubMed] [Google Scholar]
- 62. Easter SS, Malicki JJ. The zebrafish eye: developmental and genetic analysis. Results Probl Cell Differ. 2002; 40: 346–370. [DOI] [PubMed] [Google Scholar]
- 63. Baye LM, Link BA. Nuclear migration during retinal development. Brain research. 2008; 1192: 29–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Clark BS, Cui S, Miesfeld JB, Klezovitch O, Vasioukhin V, Link BA. Loss of Llgl1 in retinal neuroepithelia reveals links between apical domain size, Notch activity and neurogenesis. Development. 2012; 139: 1599–1610. 10.1242/dev.078097 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Halfter W, Dong S, Balasubramani M, Bier ME. Temporary disruption of the retinal basal lamina and its effect on retinal histogenesis. Developmental biology. 2001; 238: 79–96. [DOI] [PubMed] [Google Scholar]
- 66. Zhou H, Muramatsu T, Halfter W, Tsim KW, Peng HB. A role of midkine in the development of the neuromuscular junction. Mol Cell Neurosci. 1997; 10: 56–70. [DOI] [PubMed] [Google Scholar]
- 67. Muramoto A, Imagama S, Natori T, Wakao N, Ando K, Tauchi R, et al. Midkine overcomes neurite outgrowth inhibition of chondroitin sulfate proteoglycan without glial activation and promotes functional recovery after spinal cord injury. Neurosci Lett. 2013; 550: 150–155. 10.1016/j.neulet.2013.06.025 [DOI] [PubMed] [Google Scholar]
- 68. Yoshida Y, Ikematsu S, Moritoyo T, Goto M, Tsutsui J, Sakuma S, et al. Intraventricular administration of the neurotrophic factor midkine ameliorates hippocampal delayed neuronal death following transient forebrain ischemia in gerbils. Brain research. 2001; 894: 46–55. [DOI] [PubMed] [Google Scholar]
- 69. Prediger RDS, Rojas-Mayorquin AE, Aguiar AS, Chevarin C, Mongeau R, Hamon M, et al. Mice with genetic deletion of the heparin-binding growth factor midkine exhibit early preclinical features of Parkinson's disease. J Neural Transm. 2011; 118: 1215–1225. 10.1007/s00702-010-0568-3 [DOI] [PubMed] [Google Scholar]
- 70. Gramage E, Martín YB, Ramanah P, Pérez-García C, Herradón G. Midkine regulates amphetamine-induced astrocytosis in striatum but has no effects on amphetamine-induced striatal dopaminergic denervation and addictive effects: functional differences between pleiotrophin and midkine. Neuroscience. 2011; 190: 307–317. 10.1016/j.neuroscience.2011.06.014 [DOI] [PubMed] [Google Scholar]
- 71. Goldman D. Müller glial cell reprogramming and retina regeneration Nature Publishing Group; 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Adult zebrafish were dark-adapted for 24 h, and photolytic lesions consisted of a 30 min exposure to high-intensity light, by three days of constant bright light. At one day post-lesion (dpl), morpholinos were injected into the vitreous cavity and electroporated into the right eye of experimental and control animals. At two dpl, fish were immersed in BrdU solution for 24 hrs. Animals were then sacrificed at three dpl for quantification of BrdU+ cells, or returned to normal light conditions and sacrificed at six dpl for quantification of regenerated rod and cone photoreceptors. BrdU: Bromodeoxyuridine; rho: rhodopsin; pde6c: phosphodiesterase 6c.
(TIF)
Specificity of the anti-Mdka antibodies was determined by the selective loss of Mdka immunolabeling in Western blots (panel A) and retinal sections (panel B) from morphant embryos at 48 hpf. (A) For Western blots, β-actin served as the loading control. (B) Mdka MM—embryos injected with control morpholinos; Mdka MO—embryos injected with ATG-targeted morpholinos. Scale bar = 20 μm.
(TIF)
Sections were stained with antibodies against Mdka and the rod marker Zpr3 (panel A) or the red-green cones marker Zpr1 (panel B). Note the Mdka immunostaining of the rod photoreceptor nuclei and co-localizations with rod inner segments (arrows, panel A). Zpr1 labels the cell surface of red-green cones and this marker does not colocalize with the Mdka immunostaining (panel B) ONL: outer nuclear layer; INL: inner nuclear layer. Scale bar equals 10 μm.
(TIF)
Sections from retinas electroporated with control morpholinos show robust Mdka immunostaining (column A). In contrast, retinas electroporated with ATG-targeted morpholinos show a clear knockdown of Mdka immunostaining (column B). INL: inner nuclear layer; GCL: ganglion cell layer. Scale bar equals 50μm.
(TIF)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.