

Abstract
Few places bear as much historical and scientific significance as the breeding ground, the accumulation of stagnant water where disease-carrying insects lay their eggs. Since the turn of the twentieth century, when mosquitoes of the Anopheles genus were identified as the vector of malaria transmission, these aquatic habitats have been a key object of epidemiological research and public health intervention against the disease. Yet the breeding ground can be incorporated into a number of different topologies, each implying a different spatialization of malaria and a distinct imagination of what kind of mosquito control is ‘doable'. A contemporary example of malaria control in Dar es Salaam, Tanzania, illuminates an essential tension between what we characterize as territorial and bionomic approaches to the breeding ground—that is, between control strategies premised on treating all mosquito habitats within a given region, and those that prioritize certain sites on the basis of their position within ecological networks. Each topology localizes the breeding ground by reference to a distinct set of relations, and thus advances an idiosyncratic understanding of what sort of research is worthwhile conducting and what kinds of intervention are sustainable. The multiple ways in which the breeding ground can become an object of research and action clarifies the role of topology as an infra-logic of public health, and makes explicit the politics implicit in efforts to bring different orders of the local to scale.
Keywords: Topology, doability, public health, mosquitoes, malaria, Dar es Salaam
Introduction
We then investigated the conditions under which the Anopheles breed and propagate malaria. It was the rainy season and the place was full of stagnant pools. Everywhere the larvae of the dappled-winged mosquitoes were in these pools, while those of the grey and brindled mosquitoes occurred in tubs and pots. The great law of malaria—its connection with stagnant water on the ground—was explained. Moreover, simply by noting the presence of the larvae, we could tell at a glance which pools were dangerous to health and should be dealt with in the public interests (Ronald Ross, Nobel Lecture, 12 December 1902, p. 92).
The discovery that earned Ronald Ross, of the Indian Medical Service, the Nobel Prize transformed the geography of malaria. For centuries, mal-aria had been a matter of malignant atmospheres—a feverish disease of foul airs, vapours and pollutions. Though generally associated with swamps and marshes, these miasmic emanations had no precise location: they could be, at once, everywhere and nowhere; the disease was the somatic consequence of a noxious but ill-defined landscape (Nash, 2007). Identifying the role of Anopheles mosquitoes in the transmission of the Plasmodium parasite served to localize malaria in a fundamentally new way, turning the breeding ground, the locale where female anophelines lay their eggs, into the central object of scientific inquiry and medical intervention.1 The malaria parasites, Ross wrote,
do not float free in the air or water, as we supposed; they do not rise in mists and exhalations from the soil; they live encased in the bodies of mosquitoes, from which they pass into our blood at the moment when the insect inflicts her bite upon us (Ross, 1899, p. 2009).
With the realization that the parasite required an insect host, malaria acquired a different spatial configuration, one defined by the distance travelled by anophelines originating in a specific aquatic habitat. Suddenly new courses of action against the disease became imaginable (Packard, 2007). If mosquitoes were now the ‘visible enemy’ in the war against malaria (and the identification of Anopheles as the ‘vector’ of the disease did much to reformulate malaria control as a military hostility), the breeding site was the most favourable terrain for battle (Harrison, 1978). Armed with the new theory of anophelism, one could grasp the possibility of ridding a particular area of malaria, either by preventing the accumulation of stagnant water or by killing the insects when most vulnerable and immobile, in their aquatic larval stage. ‘[T]he practicability of eradicating malaria in a locality by the extermination of the dangerous mosquitoes in it', wrote Ross, ‘depends on a single question—Do these mosquitoes breed in spots sufficiently isolated and rare to be dealt with by public measures of repression?’ (Ross, 1899, p. 4).
The category of the breeding ‘spot’ or ‘site’ had very uncertain spatial and temporal boundaries—it could cover a huge variety of locales, from the small and transient footprint to the large and virtually permanent swamp. Yet, of all the elements and actors involved in malaria transmission, the accumulations of stagnant water containing the eggs of female mosquitoes seemed the least mobile and most easily localizable. The human and insect hosts were becoming increasingly mobile in the course of colonial expansion in the tropics, and the Plasmodium parasite was notoriously slippery. In contrast, for the few precious days it took the mosquito egg to become a flying adult, the vector of the disease stood confined in a body of water, stationary and fixed to the ground, ready to be uprooted if a systematic effort against its aquatic habitats could be mustered (Litsios, 1996).2
This paper explores how the conceptualization of this all-important but elusive locale, the breeding ground, continues to shape anti-malarial campaigns. The significance of the breeding ground lies, we will argue, in its power to establish a spatially explicit relationship between worthwhile research and sustainable intervention—that is, between what can be profitably investigated and what is pragmatically achievable. An object of scientific enquiry and the target of practical action, the breeding ground is also, first and foremost, a place or ‘site'. Its incorporation into one or another sort of topology will thus define which forms of malaria control are considered ‘doable'.
To develop our argument we draw on a contemporary example of mosquito larval control, the Urban Malaria Control Programme (UMCP) carried out in the city of Dar es Salaam since 2004. The history of the UMCP will allow us to bring into focus an essential tension between two alternative topologies of the breeding ground, two ways of understanding the geography of mosquito reproduction and control.
We describe as territorial those forms of action oriented towards the unambiguous delimitation of an area of intervention and the uniform application in that area of certain control measures. In this model, the breeding ground is a location, or locus, within a bounded region, and the most effective anti-malarial strategy is to identify and treat indiscriminately all the loci within that region where mosquitoes could potentially lay their eggs.
In contrast, bionomic approaches to mosquito control interpret the breeding ground as the contingent result of a series of ecological relationships within a landscape. Bionomic, a term we borrow from vector biology (e.g. Hocking and MacInnes, 1948), refers to the relationship between an organism, the development of its capacities throughout its life cycle and the features of its environment.3 In a bionomic topology, each breeding ground is a nidus in an ecological system, a node in a network of relations, rather than a locus within a territory. In a bionomic reading of space, territorial boundaries are irrelevant. The most promising strategy against malaria is one that elucidates the network of connections between mosquitoes, humans and the sites available for oviposition, and targets the most productive sites for Anopheles reproduction.
Both approaches to the breeding ground, the territorial and the bionomic, would agree with the dictum of the famous malariologist Nicolaas Swellengrebel, namely that ‘malaria is a local disease to be dealt with by local efforts’ (see Bradley, 1994). Yet, they understand ‘local’ in divergent ways: either as a location defined by its inclusion within a given territory, or as the intersection of a series of connections within a landscape. This difference in the interpretation of ‘local’ is tied to different sets of practices and instruments (Bhattacharrya, 2011; Kelly and Beisel, 2011). At the most basic level, a territorial approach demands constant and systematic walking—the methodical surveying of the control area so as to detect all the locations where water could accumulate—while a bionomic programme typically involves a great deal of waiting—the patient observation of individual breeding grounds in order to accurately ascertain their role in the distribution of mosquito reproduction.
We will use this contrast between walking and waiting to highlight the fact that these alternative topologies offer different answers to the two fundamental questions of vector control: what is worth knowing about mosquito behaviour, and what kinds of actions against Anopheles have a better chance of succeeding across space and over time. Before we discuss these issues in relation to the Dar es Salaam Urban Malaria Control Programme, we will outline some analytical resources that will help us examine the topological infra-logics of research programmes, and how different spatializations sustain alternative visions of doability and scalability in public health interventions.
Topology and the ‘Doability’ of Problems
In science studies there is a long tradition of research into how certain problems—particular objects of research and modalities of scientific work—emerge as ‘doable’ in relation to particular spaces of action. In her famous account of the organization of cancer research, Fujimura explained the constitution of ‘doable problems’ as the result of an alignment of three levels of work organization: experiment, laboratory and social world. By articulating tasks across these three levels, Fujimura argued, scientists are able to carve out pragmatically achievable matters of inquiry. Furthermore, Fujimura describes two conditions that facilitate the construction of doable problems: the degree of modularity of problem structure and the ability to produce packages of standardized tasks. ‘With work organized as a system of modular tasks, researchers can cut off pieces of a problem, work on each piece separately, and then plug the results back in at either the original site or another site’ (Fujimura, 1987, p. 277).
As the quote suggests, there is a spatial dimension in Fujimura's analysis; it is because problems can be understood in topological terms that one can shift results from one ‘site’ to another. This relationship between doability and topology has been elaborated by science studies scholars in at least two directions. A substantial body of work—indeed, one could argue, a central current of inquiry within science studies—explores what Shapin described as ‘the siting of knowledge production’ (1988, p. 373): how specific physical locales enable the conduct of particular forms of research and facilitate the stabilization of the resulting facts and artefacts (see also Hannaway, 1986; Livingstone, 2003; Gieryn, 2006; Henke and Gieryn, 2008). Lynch qualified the relationship between the location of scientific work and the organization of knowledge production by introducing the notion of ‘topical contexture’ (Lynch, 1991, p. 74). Rather than being contained by a particular architecture or material setting (as in Shapin's ‘house of experiment’ or Gieryn's ‘truth spots'), ‘the “place” of scientific work', Lynch argues, ‘is defined by locally organized topical contextures', that is, by ‘spatial grammars [that] are topically tied to complexes of action and equipment’ (pp. 51, 53). Spatiality, in other words, is not reducible to the features of the physical locale in which research takes place, but should be understood in relation to the efficacy of particular instrumental complexes and the kind of spatial reach those complexes enable.
The conceptualization of the space, or contexture, of research as distinct from its mere location within a physical setting opens the door to a properly topological consideration of research practices and research problems. Topology, as Mol and Law argue, ‘articulates different rules for localizing in a variety of coordinate systems’ (1994, p. 643; emphasis in original). In their triptych of topologies of anaemia, Mol and Law identified three such sets of rules: regions, networks and fluid spatialities. As we will see below, the categories of region and network overlap significantly with our distinction between territorial and bionomic imaginaries of the malaria breeding ground. Regions are spaces defined by boundaries and relations of metric proximity. Networks, on the other hand, are spaces ‘in which distance is a function of the relations between the elements and difference a matter of relational variety’ (Mol and Law, 1994, p. 641).4 Thus a disease [in this case anaemia, but similarly atherosclerosis in Mol's later work (see Mol, 2002)] is understood and researched differently—it is in fact a radically different entity—in each topological configuration.
Medical entomology provides a fruitful site for exploring spatial ontologies further. Its object of inquiry, the insect ‘vector', is characterized by its ability to shift scale, to interconnect multiple levels of description and their respective politics (see Hinchliffe, 2008; Shaw et al., 2010; Lezaun, 2011; Kelly, 2012). In the specific context of public health, perennial discussions about the scalability of interventions tend to be resolved by the ascendancy of a particular topological imaginary. If the choice of topology determines the understanding of the ‘local', it follows that scale will have a very different meaning depending on the topology in play. ‘Scaling up’ an intervention, or escalating a particular research programme, is never simply a matter of enlarging or extending a particular geographical area. If only because shifts in scale imply changes in the number and type of actors involved, and in the quality of their relations, an intervention is hardly ever kept constant as it is transposed across different contextures of action; each reconfiguration of the intervention area must contend with, and be shaped by, a different ecology of actors and practices. As Bloom and Ainsworth (2010) argue, ‘becoming larger', in the sense of widening the geographical coverage of a certain mode of action, is only one possible meaning of ‘scaling up’ in contemporary public health. Increasing or expanding impact (even, and especially, when the action remains tied to the same intervention area), or intensifying the degree of grassroots mobilization (intensifying local forms of collective action) are alternative interpretations of the process. In other words, scaling up, or escalating, is not merely a technical exercise but a political process, involving decisions over what constitutes impact and for whom, and, ultimately, which actors and institutions will be empowered and held accountable. As we will see below, decisions about whether, when and how to ‘scale up’ a project like the Dar es Salaam malaria control programme served to bring into focus alternative topological imaginaries of the object of intervention, of the doability of different modes of action, and of the relevant constituencies and lines of authority.
Larval Control in Dar es Salaam
Launched in 2004 by the City Medical Office of Health, the Dar es Salaam Urban Malaria Control Programme (UMCP) was conceived as a five-year pilot project designed to test the efficacy and cost-effectiveness of mosquito abatement, and specifically larval control, across large sections of the city. The programme received funding from a variety of national and international organizations, while the Ifakara Health Institute (IHI), a Tanzanian institution in regular collaboration with foreign academic groups, coordinated the different research activities in support of the programme's objectives.5
The UMCP extended over 15 of the city's 75 wards, encompassing a total area of about 55 square kilometres that is home to a population of more than 600,000 people. This intervention area encompassed a variety of urban terrains, including some of the most densely populated neighbourhoods in the city and sizable areas dedicated to agricultural and industrial activities. In these 15 wards the UMCP sought to identify and record all the locations where Anopheles mosquitoes could potentially breed—that is, every open body of water, however small or transient, whether water-filled or presently dry, where mosquito eggs could spend the few days they need to become flying adults (Dar es Salaam Urban Malaria Control Programme, 2005). In the three ‘intervention wards’ of Buguruni, Mikocheni and Kurasini, the surveying of breeding grounds was complemented by weekly applications of a microbial pesticide, Bacillus thuringiensis var. israelensis. All these tasks—finding, mapping and treating larval habitats—were carried out by local citizens recruited neighbourhood by neighbourhood and known as Community-Owned Resource Persons, or CORPs.
In its chosen course of action the UMCP offered an almost perfect example of territorial intervention. It sought, above all, to achieve comprehensiveness and uniformity of action within the designated intervention area by identifying all potential breeding sites. Yet prior to the launch of the programme, researchers at IHI had explored the possibility of a more selective strategy, one that would concentrate on a particular subset of breeding grounds, namely those with the highest productivity of adult anophelines. Such a ‘targeted', or, as we prefer to call it, bionomic, approach required the discovery of which particular habitats, among the multitude of bodies of stagnant water scattered across the city, provided the best reproductive conditions for mosquito larvae.6
Anopheles gambiae, the most prolific vector of malaria in Dar es Salaam, is known to favour relatively small bodies of water exposed to sunlight, but otherwise the pattern of their oviposition choices is hardly evident.7 The effort of the IHI researchers to develop criteria for the selective targeting of habitats failed to discover any obvious distribution of mosquito breeding in the city. Larvae of Anopheles were found in a bewildering variety of aquatic locations across Dar es Salaam, including many that had been considered inimical to anopheline reproduction. This precluded the possibility of a discriminating approach, and suggested instead the need to act widely and universally, to treat each and every accumulation of water without exception. ‘Clear ecological characteristics of the breeding requirements of Anopheles sp. larvae', the report of this initial survey noted, ‘could not be identified in this setting. Hence, every stagnant open water body, including very polluted ones, have to be considered as potential malaria vector breeding sites’ (Sattler et al., 2005, abstract).
As the result of this exploratory research, and of their involvement in malaria control programmes elsewhere in sub-Saharan Africa, the researchers who designed the UMCP saw Anopheles mosquitoes, and specifically those of the gambiae species complex, as ‘notoriously opportunistic’ (Killeen et al., 2006, p. 517), capable of breeding in virtually any receptacle of water and quick to adapt to changes in their material environment. There might be factors influencing where they laid their eggs, but the available entomological expertise was too limited, and changes in the urban environment too rapid and haphazard, to allow a reasonable prediction of what particular locations in the city would offer the most attractive places for breeding. In this context, the most reasonable course of action was an indiscriminate but systematic effort, based on ‘rigorous searches on foot’ (Mukabana et al., 2006, para 2), to locate every single potential habitat in the city. Puddles and tyre tracks, swampy areas, mangrove forests, drains and ditches, but also construction pits and any other man-made hole, open water storage containers, rice paddies, matuta (agricultural ridges and furrows), ponds, streams and river beds, abandoned tyres, in sum, every site where water could accumulate for a few days, ought to be identified and surveyed for the presence of larvae. By 2008, four years into the programme, the CORPs recruited by the UMCP had recorded about 65,000 potential breeding locations in their areas of responsibility (Fillinger et al., 2008).8
In the territorial topology of the UMCP there was thus an explicit relationship between the sort of actor Anopheles was thought to be and the nature of the control activities adopted by the programme. In an urban terrain as large, diverse and rapidly shifting as that of Dar es Salaam, the mosquito's predilection for ‘habitats that often occur transiently and unpredictably’ (Fillinger et al., 2008, table 2) set clear limits to any attempt to target or prioritize certain locations, and recommended instead an approach premised on the exhaustive coverage of the intervention area. The opportunism of Anopheles, its resourcefulness in turning the most unlikely places into breeding grounds, made the constant walking of the CORPs indispensable; the insect's capricious nomadism was to be met by the regimented mobility of the surveillance teams. Extensive and methodical walking, that ‘prime way of exploring the unpredictable and the incalculable’ (Solnit, 2001, p. 10), was the UMCP's response to the erratic distribution of breeding opportunities.
Proponents of a bionomic approach to malaria control, in contrast, advance a different interpretation of mosquito behaviour, and of the value and limits of entomological knowledge. The flying behaviour of an adult Anopheles might well include an opportunistic dimension, but the insect ‘switches to directional, confirmatory flight when cues originating from a resource are perceived’ (Gu and Novak, 2009). As a result, mosquitoes are ultimately predictable actors, provided one dedicates sufficient time and effort to studying the distribution of the resources—fundamentally blood, nectar and water—that determine the spatial pattern of their foraging. As we will see, this confidence in the promise of entomological knowledge implies an important difference in the tempo and material practice of control.
Mapping the Territory
The daily walks of the CORPs in search of mosquito breeding grounds brought an element of regularity to a city notorious for its degree of ‘informal’ organization and opaqueness to administrative oversight. The 15 wards of the UMCP encompassed a Y-shaped transect of the city connecting Temeke, Ilala and Kinondoni, the three municipalities that make up the city of Dar es Salaam. This territory reached from the relatively affluent area of Mikocheni in the North to the port facilities along Kurasini Creek in the South, and encompassed ‘as wide a variety of malariological situations as possible’ (Fillinger et al., 2008, para 7).
Each of the CORPs employed in the surveillance effort was responsible for an area small enough to be traversed on foot weekly—on average about 0.6 km2 of territory.9 This area of individual responsibility typically included a handful of Ten-Cell-Units (TCUs), the smallest section of territorial organization in Dar es Salaam. Introduced in the 1960s by the ruling party TANU (the Tanganyika African National Union, currently in power as CCM, or ‘Party of the Revolution'), TCUs were a means of mobilizing grassroots support for party and state initiatives. They can encompass anything from the nominal 10 to several dozen households, and are headed by an elected leader, or mjumbe, who for several decades played the double function of party representative and municipal administrator (Brennan et al., 2007). While TCUs and their leaders lost their official administrative role with the abolition of the one-party system in 1992, they proved essential in enabling the UMCP to complete one of its central tasks: the production of a detailed cartography of the territory of intervention.
Each of the breeding sites discovered by the larval inspectors had to be described, assigned a unique identifier, and plotted on a map for the benefit of subsequent inspections and pesticide application. Yet municipal authorities possessed no map of Dar es Salaam at a sufficiently small scale, or sufficiently up-to-date, to allow the proper inscription of the breeding grounds that the CORPs were discovering during their walks. In order to produce a new representation of the intervention area from the bottom up, CORPs were instructed to draw a sketch map of every TCU within their area of responsibility (Figure 1). These maps should, first of all, delineate with accuracy the boundaries of every TCU. For this purpose, the CORPs would take their first tour of the area in the company of the respective TCU leader, along with the leaders of all the adjacent units. That way disputes over the exact limit of each unit would be adjudicated in situ, thus ensuring that different areas of responsibility were effectively contiguous and that the sketch maps left no land unsurveyed—a problem particularly pressing in open areas, such as swamps, river valleys or mangrove forests, where TCU boundaries were contentious or simply inexistent. ‘Explain to the 10-cell unit leaders', the CORPs mapping manual suggests, ‘that unless the boundaries are correctly and mutually agreed upon, mosquitoes will breed in these boundary areas and fly into the 10-cell units’ (Dar es Salaam Urban Malaria Control Programme, date unknown).10 Once those boundaries had been clearly delimited, the hand-drawn maps were superimposed on satellite images of the city and the limits of each TCU adjusted with the help of GIS technology, until a complete cartography of the intervention area and of its different areas of individual responsibility was composed (Dongus et al., 2007).
Figure 1.
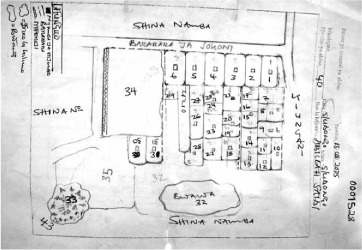
Ten-cell unit sketch map. Source: Dongus et al. (2007).
In addition to tracing the boundaries of TCUs, CORPs were required to delineate every single plot of land in each unit, and to identify its owner or main occupant—the person who could grant access to the premises for surveillance or larviciding purposes. Here again TCU leaders proved to be the key mediators. Not only were they in a position to identify individual plots and the name of their occupants, they were also indispensable in helping the CORPs—members of the local community with little power and no legal mandate to enter private premises—gain access to walled compounds and properties whose occupants were reluctant to allow inspection (Chaki et al., 2011).
TCU leaders were in fact so instrumental that their own houses were used as the reference point in the map-making effort. As the UMCP mapping handbook indicates (Figure 2), the leaders’ houses, which often fly the green flag of the ruling CCM party and can be readily identified by local residents, provided the most visible landmark in an often confusing urban geography.
Figure 2.
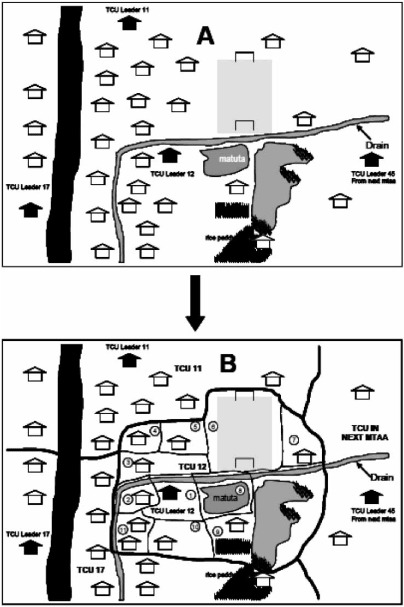
Guidelines for ten-cell-unit mapping. Source: Dar es Salaam's UMCP, Guidelines (date unknown).
Diagram A in Figure 2, the manual notes,
represents how a part of Dar es Salaam City would look like to any other person who is not interested in 10-cell units mapping whereas diagram B [at the bottom] represents what we would like to achieve in our 10-cell unit mapping exercise. (Dar es Salaam UMCP, date unknown, p. 6)
The black houses represent the homes of TCU leaders in the area. They serve as the primary reference point for drawing the unit's boundaries: each TCU should be mapped around the residence of the respective mjumbe, and should contain only one such residence. When in doubt about whether a given section of territory belonged to one or another TCU, CORPs should simply ask local residents to identify the local mjumbe.
The details of this cartographic effort illuminate a central aspect of territorial interventions: how, by their emphasis on the precise demarcation of geographical jurisdictions, they are unavoidably entangled with the political actors and constituencies that operate in the control area and as a result they become indirectly implicated in questions of political legitimacy. The UMCP was formally embedded within the official municipal bureaucracies (City Council, municipalities, wards), but it was also reliant on constituencies of a more ambiguous character, like TCUs and their leaders, or the ‘street chairman’ who assisted in the recruitment of the CORPs. The relevance of TCUs and their leaders, in particular, stems from the fact that they represented a system of territorial jurisdictions that was immediately recognized—or at least recognizable—by actors on the ground. By using TCUs as the unit of surveillance, rather than superimposing a raster grid over the urban space, the programme was able to mobilize local expertise and authority in the benefit of an exhaustive coverage of the intervention area. The advantage of the UMCP was, in the words of the researchers, that ‘it considers user-definable boundaries that can be agreed in a participatory manner on the ground and that can be readily recognized by community-based staff without access to, or the necessary education to use, GIS technology’ (Dongus et al., 2007, para 5). This recognizability of the TCU leader, the expectation that local residents would be able to identify who the mjumbe in each and every piece of territory was and where he lived, was a grassroots, everyday political reality that underpinned the entomological activities of the UMCP. These leaders might have lacked a formal role in the governance of the city, but they were in a position to adjudicate the micrological issues at stake in the production of the sort of fine-grained urban cartography on which a territorial approach like the UMCP depended.
Bionomic Counter-points
In a critique of malaria vector control campaigns that attempt to deal simultaneously with all the breeding grounds in a particular area, Gu and Novak question the practical value of the sort of extensive walking and territorial mapping characteristic of programmes like the Dar es Salaam UMCP. ‘It is unnecessary', they write, ‘to manage all aquatic habitats to obtain significant reductions in incidence and prevalence of malaria in situations of low and intermediate levels of transmission’ (2005, p. 546). Instead, it is possible to reduce the rate of transmission more efficiently by concentrating the larviciding effort on the subset of breeding grounds that accounts for the largest proportion of the adult mosquito population. In the particular context of sub-Saharan Africa, moreover, campaigns that attempt to tackle all potential larval habitats are bound to fail, they argue, as scarce resources are dissipated in a futile attempt to discover and treat every accumulation of stagnant water. In their view, the reason many African governments and international donors show little interest in sponsoring larval control programmes is that ‘the interventions are typically built around “all-out” campaigns of blanket treatment of all aquatic habitats, which is clearly beyond the reach of most resource-deprived communities in sub-Saharan Africa’ (Gu et al., 2008, p. 2).
As we have indicated, the bionomic alternative rests on a different interpretation of the spatiality of malaria and of the topology of its breeding grounds. In any ecological system, the bionomic argument goes, the resources necessary for mosquito reproduction will be unevenly distributed across space. As a result, a small proportion of all potential breeding sites will in effect account for a disproportionate number of adult mosquitoes. In fact, each breeding ground, as the physical node of a series of ecological relationships, will have a specific degree of productivity. This productivity will depend on a series of features idiosyncratic to that particular topos (depth, temperature and salinity of the water, the presence or absence of organic matter and predators, position in sun or shade, etc.), and on its location within a network of mosquito foraging and oviposition resources. The design of an effective mosquito and malaria control programme must take this fact into account, and elucidate the meshwork of biological relations in order to identify the ‘focal points’ of mosquito breeding. In a bionomic interpretation of malaria transmission, then, it is the system of proximities and distances between humans, mosquitoes and pools of stagnant water, not the boundaries of irrelevant territorial jurisdictions that provide the most practical topology for a successful anti-malarial campaign.
Crucially, a bionomic topology is tied to a very different set of practices and equipments. The paradigmatic instrument of a territorial programme like the UMCP, which depends on extensive walking, is the larval dipper. A plastic or wooden stick with a small container at the end, the dipper is carried by the inspector on his surveillance walks and used to sample bodies of water. The CORPs enrolled by the UMCP were instructed to dip up to 10 times in every body of water, depending on its size. If no larvae were captured in any of the dips, the site was characterized as containing ‘no larvae', if the average was one larva per dip or less that habitat would be registered as ‘low density', and if the CORP was able to find more than one larva per dip on average the location would be described as ‘high density'. These rough classifications were updated weekly, ‘to keep up with the rapidly changing field situation’ (Fillinger et al., 2008, overview, para 3), and served to guide the larviciding effort.
The dipper turned the walks of the CORPs into scientific expeditions. Crucially, this scientific instrument interrupted their walking as little as possible. With a minimal pause in his perambulation, the inspector was able to survey his territory and produce a rough estimate of the larval population in his area of responsibility. Furthermore, the CORPs were taught to distinguish the larvae of Anopheles mosquitoes macroscopically, without the need of any further apparatus or trip to the laboratory, by their habit of floating parallel to the water surface (as opposed to hanging down from the surface, as members of the Culex genus tend to do).
The dipper of a territorial intervention is, however, ill-suited to the requirements of a bionomic programme. Larval density is a poor predictor of the number of adult mosquitoes that will eventually emerge from an aquatic habitat. The most accurate way of measuring the productivity of a breeding ground is through ‘emergence sampling', that is, by effectively capturing and counting every adult mosquito as it flies away from the body of water. Most bionomic programmes will, however, settle for the less accurate but faster method of sampling the population of pupae. Given the relatively low rate of pupal mortality, this count will produce a better estimate of the number of adults that will eventually rise from any given habitat.
Dipping is not an adequate means to sample the population of pupae, however, pupae tend to be more concentrated than larvae, and therefore are easier to miss even after repeated dipping. They are also increasingly alert to disturbances in their environment and thus more likely to elude the dipper by swimming to the bottom of the breeding ground. The instrument of choice for the collection of pupae is the area sampler—an open ended plastic or metallic cylinder that is placed firmly in the ground and serves to capture all the mosquitoes in the pupal stage contained in that portion of the habitat (e.g. Mutuku et al., 2006; Fillinger et al., 2009).
However, installing and handling the area sampler takes considerable time. This fact points to the particular relationship between the mode of action, the preferred instrumental complex, and the temporal organization of the intervention. In short, a bionomic reading of the landscape of mosquito breeding demands a considerable upfront investment in research time. Mosquitoes have to be patiently observed to discern the distribution of their oviposition, and once the selection of breeding grounds to be sampled has been made, the process of capturing the full population of pupae in those sites takes time (in the extreme case of ‘emergency sampling', researchers will have to wait days until the traps placed over the water surface have captured all the adult mosquitoes coming out of the particular body of water). This clearly slows down the intervention, or at least delays the start of the larviciding effort. The pay-off for that initial stillness is, the bionomic argument goes, a more precise mosquito killing operation.
The choice between dipper and area sampler, or between territorial and bionomic programmes more generally, is thus a choice over how much knowledge about the vector is worth acquiring at what cost in time and effort. As we have seen, this choice presupposes a judgment about the relative strengths and weaknesses of humans vis-à-vis mosquitoes. The dipper is designed to facilitate the mobility of a walking inspector whose duty is to comprehensively survey a certain region of territory and detect all the breeding locations of a highly opportunistic actor. The area sampler is designed to produce a more accurate elucidation of the geography of mosquito reproduction, at the expense of a more static stance but with the hope that a more detailed (and prolonged) scientific investigation will result in a more precise larval intervention—an intervention that will require much less walking, once the parameters of mosquito behaviour have become clear. In a territorial topology, the circulation of the surveyor, not the movement of the mosquito, is the measure of all things; walking is both the organizing principle in the demarcation of control areas and the most important tool of larval control. Patient observation is the hallmark of a bionomic approach, for it serves to clarify the network of ecological connections that structure mosquito reproduction, and thus permits the concentration of scarce resources on those locations that are most likely to make a difference to the overall density of the mosquito population.
The unbridled mobility of their control practices gives territorial approaches a special political quality. In her history of walking, Rebeca Solnit notes how ‘on foot everything stays connected, for while walking one occupies the spaces between’ (2001, p. 9). The constant perambulation required by a programme like the UMCP entangles malaria control practices in myriad urban realities, such as the adjudication of contested administrative boundaries or the recognition of ‘informal’ political authorities at the street level. The travails of the CORPs as they zigzag through the city in search of bodies of stagnant water express this ability of the urban walker to trespass on different aspects of the city's political and material reality.
In sum, territorial and bionomic topologies of vector control imply much more than contrasting imaginations of the spaces of mosquito habitation and malaria transmission. The contrast between walking and waiting, which we have used as a sort of shorthand to describe alternative forms of operating in the world of mosquitoes, captures an opposition between two different sets of bodily and instrumental stances, two starkly different ‘taskscapes’ of malaria control (Ingold, 1993). Each topology articulates an idiosyncratic notion of space, and of the way of treating mosquitoes in it, whether that space is the bounded region of the territorial approach, or the network of ecological relations of a bionomic landscape.
Shifting Scales
As the UMCP neared the end of its planned five-year development, the question of its future as a permanent and city-wide municipal programme emerged with new force. The UMCP had been designed as a pilot project intended to test the feasibility and cost-effectiveness of larval control, and it was now time to decide whether what had been achieved in the 15 wards of the programme, and particularly in the three ‘intervention wards’ where weekly larviciding had been implemented, could be extended to the city as a whole and become a routine component of the administration of Dar es Salaam.
With the question of the scalability of the UMCP, the choice of topological imaginary came again into focus. Shifts in the spatial and temporal scale of the programme—how to extend the original larval control practices geographically, and how to make them sustainable over a longer period of time—were bound to open up once again the question of what sort of spatialization of the breeding ground would best guarantee the doability of mosquito abatement. Were the sort of territorial, labour-intensive practices of the UMCP sustainable across a city sprawling over an area of 1,800 square kilometres? Could the systematic walking of the CORPs, and the thorough mapping and record-keeping that guaranteed exhaustive coverage of the territory, be extended across an enlarged intervention area? And how would a temporary effort like the UMCP fare as it became a routine component of the municipal bureaucracy?
Fred Soper, whose 1930s campaign against Anopheles in Brazil was often credited as an inspiration for the UMCP's territorial philosophy, had famously established a straightforward geometry of mosquito control. ‘There is no law of diminishing returns and no indestructibility of a biological entity', he noted.
The mathematics of eradication is simple; what can be done in one square metre can be done in two square metres; what can be done in two square metres can be done in four. Thus, by geometrical progression the world is soon covered (Soper, 1962, quoted in Shaw et al., 2010 p. 380).
Things were less straightforward, or geometrical, in Dar es Salaam. In fact, the UMCP itself had laid bare some of the difficulties in achieving comprehensive coverage (Chaki et al., 2009), difficulties that would only be compounded with the geographical extension of the area of intervention: hundreds of CORPs would need to be supervised, hundreds of thousands of breeding sites would need to be visited regularly. In the face of such administrative challenges, some participants in the UMCP argued in favour of a more explicit targeting of certain kinds of habitats; a bionomic shift, in other words, that would see the programme focus on those locations with the highest mosquito productivity.
Discussions about the imminent scalability of the UMCP thus reintroduced a tension between alternative topologies of the breeding ground, and brought to the forefront different views about the value of entomological knowledge and the urgency of intervention. Underlying the territorial approach of the UMCP there had always been a degree of scepticism about the possibility of translating greater and more precise scientific knowledge into more effective larval control on the ground. First, because it is unclear that a sufficiently accurate knowledge of the behaviour of mosquitoes could ever be produced. ‘Despite our long experience with sampling Anopheles larvae in a variety of sites across Africa', argued a group of experts in malaria control that included leading members of the UMCP research tem in response to calls for a more targeted approach, ‘we are unable to reach consensus about what the most consistently productive habitat types are or where we would look for them on a given day in a given setting’ (Killeen et al., 2006, p. 517). In a constantly changing urban terrain like that of Dar es Salaam ‘the distribution of habitats themselves is highly dynamic and represents a moving target’ (p. 517), and this limited the value of highly complex but ultimately static representations of the malariological landscape.
But even if a more accurate representation of Anopheles behaviour were to be achieved, the most important question was whether that augmented body of knowledge would result in more efficient mosquito control. Proponents of territorial approaches contend that greater reliance on scientific expertise is detrimental to programmes, like the UMCP, that rely on modestly paid ‘volunteers’ systematically completing menial tasks. ‘Communities', write the organizers of the UMCP, ‘represent the greatest and least exploited resource available for malaria control in Africa today’ (Mukabana et al., 2006, para 3). The adequate mobilization of that resource requires a programme that makes modest demands on the level of technical expertise of those administering and implementing its day-today activities. ‘Even if reliable targeting criteria could be identified, the successful application of such elegant but technically complex criteria by community-based personnel in the most disadvantaged countries in the world is difficult to envisage in the near future’ (Vanek et al., 2006, concl. para 2). This trade-off between greater scientific precision and practical feasibility was often stated by members of the UMCP in defence of the programme's territorial orthodoxy:
Targeting of the most productive habitats could improve the cost-effectiveness of mosquito larval control in Africa. (…) However, the operational challenges of a large-scale programme and the lack of scientific evidence of its feasibility and effectiveness suggest the need for exhaustive coverage and very simple implementation protocols that can be implemented by community-level staff with minimal education (Dongus et al., 2007, p. 2).
In a context like Dar es Salaam, success in the elimination of mosquitoes does not depend on the further scientific elucidation of mosquito behaviour, the organizers of the UMCP argued, but on the availability of ‘simple and readily verifiable targets [to be] provided to field workers at the sharp end of the mosquito control spear: to kill all mosquitoes'. The policy most likely to succeed, they noted, is ‘shoot first, ask questions later'—a philosophy that ‘may seem crude in the modern era, but [whose] success remains unrivalled, and to implement it today is a challenge in itself’ (Killeen et al., 2006, p. 518).
One hears in these remarks echoes of Ronald Ross's warning against those tempted to make the effort against mosquitoes conditional upon the resolution of complex scientific questions:
Amateurs are fond of advising that practical measures should be postponed pending the carrying out of detailed researches upon the habits of Anophelines, the parasite rate of localities, the effect of minor works, and so on. In my opinion this is a fundamental mistake. It implies the sacrifice of life and health on a large scale, while researches which may have little real value, and which may be continued indefinitely are being attempted. (Ross, 1910, pp. 229–330)
For Ross, the difficulty of the enterprise recommended prompt, indiscriminate and comprehensive interventions. A contemporary exemplar of this philosophy, the UMCP was built around ‘practical procedures that rely on minimal technology’ (Fillinger et al., 2008, abstract), and saw in the simplification of the task at hand the most critical variable of success. In African cities, the work of locating, mapping and treating breeding grounds is likely to be conducted by low-paid local citizens, with little prior training and, as in the case of the CORPs, employed casually and outside the formal municipal bureaucracy. A research-intensive effort would make the programme too dependent on technical expertise, and would require a degree of scientific training and monitoring at odds with local ‘ownership’ of the programme.
Conclusion
In this paper we have tried to show that every effort to intervene against malaria—or any other insect-borne disease, for that matter—at the level of the breeding ground will adopt a certain topological orientation, and in so doing will establish a particular relationship between what sort of knowledge is considered worth acquiring and what forms of action are seen as pragmatically sustainable.
We have differentiated two possible topologies of the breeding ground, a territorial and a bionomic one. This distinction is best understood as posing a set of choices regarding the selection of targets and the manner of implementing malaria control strategies. Each topology is tied to a different understanding of the insect vector, and is articulated in a different instrumental complex. The mosquito that inhabits an ecological network of bionomic productivity is a very different actor from the insect that populates the control area of a territorial campaign. If the territorial topology is a response to the opportunism and adaptability of Anopheles in a fast-changing urban terrain, the bionomic effort to ascribe a measurement of productivity to individual breeding sites rests on the assumption that a complex but decipherable pattern underpins the mosquito's choice of reproductive locations, and that the explication of such a pattern represents a worthwhile investment. Territorial and bionomic perspectives thus put forward contrasting views on the practical value of gaining increasingly precise knowledge about the behaviour of mosquitoes. They offer, in other words, differing views on the relationship between entomological research and mosquito control.
In its pilot stage, the Dar es Salaam UMCP was a resolutely territorial programme: it sought to identify every single aquatic habitat within clearly delimited areas of control, seeking comprehensiveness and uniformity of action. The genealogy of territorial topologies in the history of malaria control goes back to the advice of Ronald Ross not to make mosquito abatement dependent on the resolution of difficult scientific questions, and to rely instead on the systematic and relentless application, within a well-characterized area, of what is already known, on the thorough implementation of what is doable from the very start.
The great thing is to make a beginning: not to form counsels of perfection, not to measure means with ends, but simply to set to work with whatever force there is available, however small it may be. A single private citizen can eradicate malaria from a whole town. In an enterprise of this nature, the means grow as the work proceeds. (Ross, 1902, p. 30)
At critical points in the life of the UMCP—when the nature of the intervention was first being considered, and when the time came to extend the initial programme to the whole city—the possibility of tempering its territorial emphasis with a measure of bionomic ‘targeting’ was seriously considered. Particularly at the end of the five-year pilot programme, when the choice had to be made whether to extend the programme geographically and how to transform it into a permanent feature of municipal administration, the tension between these two approaches became most evident.
That tension makes clear the role of topological infra-logics in construing a particular kind of intervention as ‘doable'. In making the case for one or another form of larval control, proponents and critics of the sort of territorial action the UMCP exemplifies mobilized arguments about the behaviour of mosquitoes, the reliability of particular control techniques (and of the people who will use them), the practical value of entomological knowledge, and the proper distribution of roles between researchers and administrators. The breeding ground was the nexus of both territorial and bionomic programmes of action, but that seemingly unambiguous ‘place’ could be configured within very different spatial grammars. The choice made different forms of larviciding more or less ‘doable', and construed malaria control as either a matter of constant and rigorous walking, or of patient and attentive waiting.
Acknowledgements
Our first thanks go to the Public Health Entomology Team at Ifakara Health Institute in Dar es Salaam, and in particular to Prosper Chaki, Stefan Dongus, and Gerry Killeen. This paper draws from research conducted under the auspices of the project ‘Understanding the Dynamics of Urban Flexibility and Reconstruction', sponsored by the Oxford Programme for the Future of Cities. We would like to thank our colleagues in the research group, Idalina Baptista, Matilde Cordoba, Michael Guggenheim, Ella MacPherson and Fernando Dominguez Rubio for their collaboration and advice. Amy Hinterberger and Natalie Porter provided useful comments on an early version of the paper. Preliminary ethnographic work by Ann Kelly received funding from the Wellcome Trust (Bioethics Grant #2173); further research by Javier Lezaun received funding from the European Research Council under the European Community's Seventh Framework Programme (FP7/2007–2013)/ERC grant agreement no. 263447 (BioProperty).
Notes
Ross's observations consolidated an emerging picture of the aetiology of malaria. Alphonse Laveran's detection of Plasmodia parasites in the blood of malaria patients in 1880 gave the disease a new pathogenic profile, and by the end of the century discoveries by Patrick Manson, Ronald Ross, Giovani Grassi and others had identified mosquitoes of the genus Anopheles as the hinge in the life cycle of the parasite.
‘[I]t is hopeless to attempt their destruction when they [the mosquitoes] are winged’, Ross wrote. ‘[F]ortunately, there is a very vulnerable stage in their career, before they reach their winged condition; that is, when they are water insects—the little wriggling larvae found in pots and tubs of water, and in stagnant puddles … if we could make arrangements to empty out once a week all the tubs of water, the ditches, puddles, and wells within a given area, we should be able to exterminate the larvae within that area, at least for a time—and therefore also we should be able to exterminate, at least to a great extent, the adult mosquitoes’ (Ross, 1899, p. 2).
A common term in entomology up to the 1950s, ‘bionomic’ lost its definitional power in favour of the more contemporaneous ‘ecological'. Yet it still captures well the regulative (nomos) dimension of life's unfolding (bios).
A fluid spatiality is neither a region nor a network—and that is probably the most important thing that can be said about it. In fluid spaces continuity is neither a matter of physical contiguity, nor the result of holding relations constant; identity is never clear-cut, a question of gradients, rather than cut-off points. While one could imagine describing malaria (as Mol and Law do for anaemia) in terms of a fluid space, it is difficult to do the same for breeding grounds—defined, if by nothing else, by the interruption of flow and the emergence of stagnation. In a later contribution (2001), Law and Mol add a fourth topological possibility: fire.
Two comparable mosquito control campaigns preceded the UMCP. Between 1987 and 1996 the Japan International Cooperation Agency sponsored a programme of ‘integrated vector management’ that included an extensive larviciding effort (De Castro et al., 2004). More recently, in 2002, the municipality of Ilala, one of the three that compose Dar es Salaam, undertook the weekly surveying and treatment of Anopheles breeding grounds within its territory (Mukabana et al., 2006).
Proponents of bionomic programmes of mosquito control often describe their preferred mode of action as ‘targeted', in opposition to the ‘blanket’ approach of territorial interventions (see for instance Gu et al., 2008). Yet, ‘targeted’ is here a misleading term. A territorial programme like the UMCP is also ‘targeted’ in a number of important ways. For instance, the selection of the UMCP's ‘intervention wards’ was based on the informed assumption that municipal personnel in those three districts had a better-than-usual grasp of the administrative requirements of larval control. Similarly, although they were instructed to locate and treat all breeding grounds, de facto the CORPs would not bother with habitats that were exceedingly small—they rarely recorded the presence of footprints, for instance—or mobile—flower pots, watering cans, small buckets, etc. Otherwise their task would have been literally infinite, and the treatment of breeding grounds would be more appropriately accomplished by ‘dusting’ from an airplane, rather than by the manual application of pesticide.
Anopheles gambiae describes in fact a series of sibling species that are morphologically indistinguishable but might very well have different (and changing) preferences when it comes to the choice of breeding ground.
The UMCP's emphasis on achieving exhaustive territorial coverage was influenced by a series of historical precedents, most notably the eradication campaign against Anopheles gambiae directed by Fred Soper in north-eastern Brazil in the late 1930s. For Soper, the challenge was primarily administrative; the subtleties of vector biology took a back seat to the deployment of abundant labour, close supervision and ‘sheer will and determination’ (Killeen, 2003) in order not to miss any single breeding ground. ‘The real secret behind the success [of Soper's campaign]', wrote one of the senior entomologists in the UMCP (Killeen et al., 2002, p. 621), ‘was in the clearly defined and rigorous organization of its activities', in particular the unambiguous division of the infested area into individual zones of responsibility, and the precise delegation and supervisions of larval control tasks.
The inspiration was again Fred Soper's work in Brazil. ‘An antilarval zone', Soper and his colleague David Bruce Wilson (of the East Africa Malaria Unit) wrote, ‘represented the area in which one man could apply larvicide to all potential foci during one week’ (Soper and Wilson, Anopheles gambiae in Brazil, 1930 to 1940; quoted in Killeen et al., 2002, p. 621).
Whenever the wards included substantial areas of industrial or agricultural land, new TCUs were created from scratch to create unambiguous spaces of surveillance assigned to individual CORPs.
References
- Bhattacharrya N. The logic of location: Malaria research in colonial India, Darjeeling and Duars, 1900-30. Medical History. 2011;55(2):183–202. doi: 10.1017/s0025727300005755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bloom G., Ainsworth P. Beyond Scaling Up: Pathways to Universal Access to Health Services. 2010. Available at www.steps-centre.org/publications.
- Bradley D. J. Watson, Swellengrebel and species sanitation: Environmental and ecological aspects. Parassitologia. 1994;36(1–2):137–147. [PubMed] [Google Scholar]
- Brennan J., Burton A., Lawi Y., editors. Dar es Salaam: Histories from an Emerging African Metropolis. Dar es Salaam/Nairobi: Mkuki na Nyota Publishers/BIEA; 2007. [Google Scholar]
- Chaki P., et al. Achieving high coverage of larval-stage mosquito surveillance: Challenges for a community-based mosquito control programme in urban Dar es Salaam. Tanzania, Malaria Journal. 2009;8 doi: 10.1186/1475-2875-8-311. Article no. 311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaki P., et al. Community Owned Resource Persons for malaria vector control: Enabling factors and challenges in an operational programme in Dar es Salaam. Tanzania, Human Resources for Health. 2011;9:21. doi: 10.1186/1478-4491-921. (accessed 3 September 2012) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dar es Salaam Urban Malaria Control Programme. Guidelines to Searching for Mosquito Breeding Habitats (Stagnant Water) and Conducting Larval Survey (revised version) City Medical Office of Health, Dar es Salaam City Council; 2005. [Google Scholar]
- Dar es Salaam Urban Malaria Control Programme (date unknown) Guidelines for 10-cell Unit Mapping to be Carried Out by the Community Owned Resource Persons and the Ward Malaria Vector Control Supervisors, City Medical Office of Health, Dar es Salaam City Council.
- De Castro M. C., et al. Integrated urban malaria control: A case study in Dar es Salaam, Tanzania. The American Journal of Tropical Medicine and Hygiene. 2004;71(2):103–117. [PubMed] [Google Scholar]
- Dongus S., et al. Participatory mapping of target areas to enable operational larval source management to suppress malaria vector mosquitoes in Dar es Salaam. Tanzania, International Journal of Health Geographics. 2007;6(1):37. doi: 10.1186/1476072X-6-37. (accessed 3 September 2012) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fillinger U., et al. A tool box for operational mosquito larval control: Preliminary results and early lessons from the Urban Malaria Control Programme in Dar es Salaam. Tanzania, Malaria Journal. 2008;7(1):20. doi: 10.1186/1475-2875-7-20. (accessed 3 September 2012) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fillinger U., et al. Identifying the most productive breeding sites for malaria mosquitoes in The Gambia. Malaria Journal. 2009;8(1):62. doi: 10.1186/1475-2875-7-20. (accessed 3 September 2012) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujimura J. H. Constructing do-able' problems in cancer research: Articulating alignment. Social Studies of Science. 1987;17(2):257–293. [Google Scholar]
- Gieryn T. City as truth-spot: Laboratories and field-sites in urban studies. Social Studies of Science. 2006;36(1):5–38. [Google Scholar]
- Gu W., Novak R. J. Habitat-based modeling of impacts of mosquito larval interventions on entomological inoculation rates, incidence, and prevalence of malaria. The American Journal of Tropical Medicine and Hygiene. 2005;73(3):546–552. [PubMed] [Google Scholar]
- Gu W., Novak R. J. Agent-based modelling of mosquito foraging behaviour for malaria control. Transactions of the Royal Society of Tropical Medicine and Hygiene. 2009;103(11):1105–1112. doi: 10.1016/j.trstmh.2009.01.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gu W., Utzinger J., Novak R. J. Habitat-based larval interventions: A new perspective for malaria control. The American Journal of Tropical Medicine and Hygiene. 2008;78(1):2–6. [PubMed] [Google Scholar]
- Hannaway O. Laboratory design and the aim of science: Andreas Libavius versus Tycho Brahe. Isis. 1986;77(4):585–610. [Google Scholar]
- Harrison G. Mosquitoes, Malaria and Man: A History of the Hostilities Since 1880. New York: Dutton; 1978. [Google Scholar]
- Henke C., Gieryn T. Sites of scientific practice: The enduring importance of place. In: Hackett E. J., Amsterdamska O., Lynch M., Wajcman J., editors. Handbook of STS. Third Edition. Cambridge, MA: MIT Press; 2008. pp. 353–376. [Google Scholar]
- Hinchliffe S. Reconstituting nature conservation: Towards a careful political ecology. Geoforum. 2008;39(1):88–97. [Google Scholar]
- Hocking K. S., MacInnes D. G. Notes on the Bionomics of Anopheles gambiae and A. funestus in East Africa. Bulletin of Entomological Research. 1948;39(3):453–465. doi: 10.1017/s0007485300022550. [DOI] [PubMed] [Google Scholar]
- Ingold T. The temporality of the landscape. World Archaeology. 1993;25:152–174. [Google Scholar]
- Kelly A. H. The experimental hut: Hospitable vectors. Journal of the Royal Anthropology Institute. 2012;8(issue supplement 1):145–160. [Google Scholar]
- Kelly A. H., Beisel U. Neglected malarias: The frontlines and back alleys of global health. Biosocieties. 2011;4:71–87. [Google Scholar]
- Killeen G. F. Following in Soper's footsteps: Northeast Brazil 63 years after eradication of Anopheles gambiae. Lancet Infect Diseases. 2003;3:663–666. doi: 10.1016/s1473-3099(03)00776-x. [DOI] [PubMed] [Google Scholar]
- Killeen G. F., et al. Eradication of Anopheles gambiae from Brazil: Lessons for malaria control in Africa? Lancet Infectious Diseases. 2002;2:618–627. doi: 10.1016/s1473-3099(02)00397-3. [DOI] [PubMed] [Google Scholar]
- Killeen G. F., et al. Habitat targeting for controlling aquatic stages of malaria vectors in Africa. The American Journal of Tropical Medicine and Hygiene. 2006;74(4):517–518. [PubMed] [Google Scholar]
- Lezaun J. Bees, beekeepers, and bureaucrats: Parasitism and the politics of transgenic life. Environment and Planning D: Society and Space. 2011;29(4):738–756. [Google Scholar]
- Litsios S. The Tomorrow of Malaria. Wellington, New Zealand: Pacific Press; 1996. [Google Scholar]
- Livingstone D. N. Putting Science in its Place: Geographies of Scientific Knowledge. Chicago: University of Chicago Press; 2003. [Google Scholar]
- Lynch M. Laboratory space and the technological complex: An investigation of topical contextures. Science in Context. 1991;4(1):51–78. [Google Scholar]
- Mol A. The Body Multiple: Atherosclerosis in Practice. Durham: Duke University Press; 2002. [Google Scholar]
- Mol A., Law J. Regions, networks and fluids: Anaemia and social topology. Social Studies of Science. 1994;24(4):641–671. doi: 10.1177/030631279402400402. [DOI] [PubMed] [Google Scholar]
- Mukabana W. R., et al. Ecologists can enable communities to implement malaria vector control in Africa. Malaria Journal. 2006;5:9. doi: 10.1186/14752875-5-9. (accessed 3 September 2012) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mutuku F. M., et al. Pupal habitat productivity of Anopheles gambiae complex mosquitoes in a rural village in western Kenya. The American Journal of Tropical Medicine and Hygiene. 2006;74(1):54–61. [PubMed] [Google Scholar]
- Nash L. Inescapable Ecologies: A History of Environment, Disease, and Knowledge. Berkeley: University of California Press; 2007. [Google Scholar]
- Packard R. M. The Making of a Tropical Disease: A Short History of Malaria. Baltimore: Johns Hopkins University Press; 2007. [Google Scholar]
- Ross R. Inaugural lecture on the possibility of extirpating malaria from certain localities by a new method. British Medical Journal. 1899;2:1–4. doi: 10.1136/bmj.2.2009.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ross R. Mosquito Brigades and How to Organize Them. Milton Keynes: General Books Ltd; 1902 [2009]. [Google Scholar]
- Ross R. The Prevention of Malaria. London: John Murray; 1910. [Google Scholar]
- Sattler M. A., et al. Habitat characterization and spatial distribution of Anopheles sp. mosquito larvae in Dar es Salaam (Tanzania) during an extended dry period. Malaria Journal. 2005;4:4–4. doi: 10.1186/1475-2875-4-4. (accessed 3 September 2012) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shapin S. The house of experiment in seventeenth-century England. Isis. 1988;79(3):373–404. [Google Scholar]
- Shaw I., Robbins P., Jones J. P., III A bug's life: Spatial ontologies of mosquito management. Annals of the Association of American Geographers. 2010;100(2):373–392. [Google Scholar]
- Solnit R. Wanderlust: A History of Walking. New York: Penguin; 2001. [Google Scholar]
- Vanek M. J., et al. Community-based surveillance of malaria vector larval habitats: A baseline study in urban Dar es Salaam. Tanzania, BMC Public Health. 2006;6:154. doi: 10.1186/1471-2458-6-154. (accessed 3 September 2012). [DOI] [PMC free article] [PubMed] [Google Scholar]