

Abstract
Alzheimer’s disease (AD) is a devastating age-related neurodegenerative disease with no specific treatment at present. The APPsw/Tg2576 mice exhibit age-related deterioration in memory and learning as well as amyloid-beta (Aβ) accumulation, and this mouse strain is considered an effective model for studying the mechanism of accelerated brain aging and senescence. The present study was aimed to investigate the beneficial effects of dietary supplements pomegranate, figs, or the dates on suppressing inflammatory cytokines in APPsw/Tg2576 mice. Changes in the plasma cytokines and Aβ, ATP, and inflammatory cytokines were investigated in the brain of transgenic mice. Significantly enhanced levels of inflammatory cytokines IL-1β, IL-2, IL-3, IL-4, IL-5, IL-6, IL-9, IL-10, TNF-α and Eotaxin activity were decreased by administration of the diet supplements containing pomegranates, figs, or dates. In addition, putative delays in the formation of senile plaques, as indicated by a decreasing tendency of brain Aβ1–40 and Aβ1–42 contents, were observed. Thus, novel results mediated by reducing inflammatory cytokines during aging may represent one mechanism by which these supplements exert their beneficial effects against neurodegenerative diseases such as AD.
Introduction
Alzheimer’s disease (AD) is a progressive neurodegenerative disorder, characterized by abnormal accumulation of amyloid plaques and neurofibrillary tangles throughout the cerebrocortical and limbic regions [1]. Further evidence suggests that initial vascular damage plays a pivotal role in functional and structural changes of neurons [2–4], and accumulation of brain amyloid-beta (Aβ) peptides is subsequent to the blood-brain barrier (BBB) dysfunction and reduction in cerebral blood flow [3,5]. The neuropathological hallmarks of the disease are amyloid plaques and neurofibrillary tangles (NFT), which progressively accumulate in the brain. These neuropathologies are closely linked with chronic inflammation and neuronal dysfunction. Proinflammatory cytokines, such as interleukin (IL)-1β, IL-6 and tumor necrosis factor (TNF-α), have been reported to be involved in the formation of neuritic plaques in AD [6–8]. IL-1β has an important neuromodulatory role in the hippocampus, acting downstream of the initial events of long term potentiation [9]. An increased endogenous IL-1β concentration in the hippocampus may be a common trigger for impairments in long-term potentiation in age and stress-induced rats [10]. Transgenic mice over expressing IL-6 exhibited a progressive age-related decline in avoidance learning performance [11]. The Aβ plaques and NFTs accumulated in the brain activate inflammatory cells (i.e. astrocytes and microglia) and tissue levels of pro- and anti-inflammatory mediators, including cytokines and chemokines, are altered. In addition to the observation that inflammatory mediators are present in AD lesions, there is also epidemiological and genetic evidence which shows that an inflammatory process contributes to AD pathology. Prospective case-cohort studies show that higher serum levels of certain acute-phase proteins are a risk factor for the development of AD [12–15]. Certain polymorphisms of cytokines, most notably interleukin (IL)-1α, seem to be a genetic risk factor for AD [16]. Moreover, epidemiological studies indicate that longstanding use of non-steroidal anti-inflammatory drugs can prevent or retard the development of AD [17,18]. In contrast to the epidemiological studies, patients with a clinical AD syndrome do not benefit from treatment with anti-inflammatory drugs [19].
More recently, the interest in the role of dietary antioxidants in human health has prompted research in the field of AD. Fruits are good sources of these bioactive components, and there are a number of commercial polyphenol-rich beverages, which base their marketing strategies on antioxidant potency. Naturally occurring compounds from plants have been offering possible their therapeutic potential for AD [20–22].
Pomegranates (Punica granatum Linn.) contain very high levels of polyphenols as compared to other fruits and vegetables. The pomegranates have been extensively used in Unani, Ayurvedic and Chinese systems of oriental medicine. The plant is used in folklore medicine for the treatment of various diseases, such as ulcer, hepatic damage, snakebite, etc. Mediterranean and Middle-East countries are the main regions of pomegranate cultivation and production [23,24]. Pomegranates are a very rich source of anthocyanins (cyanidin 3,5-di and 3-O-glucoside, delphinidin 3,5- di and 3-O-glucoside, pelargonidin 3,5-di and 3-O-glucoside), ellagic acid, punicalagin isomers, different flavanols (catechins as catechins and epicatechin, and gallocatechins as gallocatechin and epigallocatechin), etc [25–27]. Dietary supplementation of pregnant mice with pomegranate juice was shown to protect against neurodegeneration in neonatal mice subjected to hypoxic–ischemic brain injury [28].
The fig (Ficus carica L.) is a classic fruit tree associated with the beginnings of horticulture in the Mediterranean basin [29,30]. The Mediterranean region and especially the Middle Eastern countries have been the most important center of figs growth from time immemorial [31]. Compared with other common fruits and beverages, figs are an excellent source of minerals, vitamins and dietary fiber; they are fat and cholesterol free and are contain abundant amino acids [32–35]; they also contain the highest concentrations of polyphenols [36]. The fig fruit is well known for its attractive taste, nutritive value due to its antioxidant properties, and it is consumed fresh or as dried products worldwide [37–39]. In traditional medical system, figs are used for treating various ailments including cardiovascular disease, respiratory problems, ulcers, warts, etc. [40–41]. Figs have been reported to exhibit antioxidant [34], antibacterial, anti-fungal [42], antispasmodic, antiplatelet [43], antipyretic [44], anti-HSV [45], haemostatic [46], hypoglycemic [47], anticancer [48–49], hepatoprotective [50], antituberculosis [51] and hypo-lipidemic activities [52]. The leaves have been used traditionally in the treatment of jaundice [53]. Figs are an excellent source of phenolic compounds, such as pro-anthocyanidins [54]. Actually, red wine and tea, two well-publicized sources of phenolic compounds contain lower amounts of polyphenols than figs [55]. Ficus carica has been reported to excellent the radical scavenging and antioxidant [34] activities.
Fruits of the date palm (Phoenix dactylifera L. Arecaceae) are commonly consumed in several parts of the world and represent a staple food in most of the Arabian countries. The date fruit has been used in folk remedies for the treatment of various infectious diseases, cancer and immunomodulatory activity [55]. Numerous studies have also shown the antibacterial, antihyperlipidemic activity [56,57], hepatoprotective activity [58], nephroprotective activity [59], Anticancer activity [60], anti-fungal [61,62] properties and antimutagenic activity [63] of date fruits.
Dates are a good source of energy, vitamins, and important elements such as phosphorus, iron, potassium, and a significant amount of calcium [64]. Besides nutritional value, date fruits are rich in phenolic compounds with free radical scavenging and antioxidant activity. Several studies have reported such activities of date fruits cultivated in Algeria [65], Kuwait [63], Oman [66], Iran [67], Bahrain [68] and the USA [65]. These studies showed different amounts of phenolic acids in fresh and dried dates. Studies with three varieties of Omani dates have shown the presence of both free (protocatechuic acid, vanillic acid, syringic acid, and ferulic acid) and bound phenolic acids (gallic acid, protocatechuic acid, p-hydroxybenzoic acid, vanillic acid, caffeic acid, syringic acid, p-coumaric acid, ferulic acid, and coumaric acid) [66]. The potent antioxidant activity of dates is due to its phenolic compounds and flavonoid constituents [69–70]. Date varieties from different regions of Oman had different levels and patterns of phenolic acids. Nine phenolic acids (gallic, protocatechuic, p-hydroxybenzoic, vanillic, caffeic, syringic, p-coumaric, ferulic, and o-coumaric acid) have been tentatively identified. It was found that ferulic acid was the major phenolic acid for all date varieties in Oman [70].
We have recently reported that the dietary supplementation of pomegranates, figs, or date palm fruits growing in Oman can provide benefits against behavior and oxidative stress related abnormalities in an APPSw2576 transgenic mouse model of AD [71–73]. In this study, we examined whether dietary supplementation with pomegranates, figs or date palm fruits can attenuate the levels of AD-like Aβ, inflammatory cytokines and ATP in aged APPsw/Tg2576 mice as an in vivo model for AD in comparison to wild type aged mice.
Materials and Methods
Collection of Fruits and Diet Preparation
Fresh pomegranates, figs, or date palm fruits were purchased from Al-Jabal Al-Akdhar farms, Oman. All the fruits were frozen at (-40°C) for 5 days. After that, the samples were ground into fine powder using a coffee grinder. The ground fruits were sent to USA to prepare the separate diets for the mice. The diets were prepared by mixing the powdered fruits (4% w/w)[71–76] individually with regular diet as per National Institutes of Health (NIH), USA protocol by Research Diets Inc, NJ, USA.
Animals and treatment
Sixty four transgenic females (APPsw/Tg2576) and 16 wild-type control (non-transgenic) mice (Taconic Farm, NY, USA) were used for this study. Animals were quarantined for 7 days after shipping and individually housed in plastic cages in an animal room, which was maintained at a temperature of 22±2°C, a relative humidity of 50±10%, and a 12-h light/dark automatic light cycle (light: 0800–2000 h). Tap water was offered ad libitum throughout the study. The study was approved by the Animal Care and Use Committee of the Sultan Qaboos University, Oman (SQU/AEC/2010-11/3), and all the procedures involving animals and their care were carried out in accordance with international laws and policies (EEC Council directives 86/609, OJL 358, 1 December, 12, 1987; NIH Guide for the Care and Use of Laboratory Animals, NIH Publications No. 85–23, 1985). All these animals are free from pathogens and viruses. Experimental period commenced from the age of 4 months. The animals were divided into five groups (n = 16/group): Group 1: Wild type (non-transgenic) control of the APPsw mice fed with regular diet, Group 2: AD transgenic mice also fed with regular diet, Group 3: AD mice fed with 4% pomegranates, Group 4: AD mice fed with 4% figs and Group 5: AD mice fed with 4% date fruits. The experimental and control mice were fed an 4% pomegranates, figs, dates, or a control diet for 15 months and then assessed for the effect of each diet on plasma cytokine levels (IL-1β, IL-2, IL-3, IL-4, IL-5, IL-6, IL-9, IL-10, TNF-α and Eotaxin), Aβ and ATP.
Sample collection
The day after completion of the behavioral tests (data not included), the animals were anesthetized with an intraperitoneal injection of ketamine (75 mg/kg) and xylazine (5 mg/kg), and blood (about 1.5 mL) was collected from the anterior vena cava and placed into heparinized tubes. Collected blood samples were centrifuged at 4000 RPM for 15min at 4°C to obtain the plasma. Brains of experimental and control animals were carefully removed, and homogenized in 9 volumes (1:9 w/v) of cold saline, centrifuged and collected the supernatant. The samples of the brain and plasma were stored at −80°C until measurement.
Cytokine analyses
The plasma levels of many cytokines were measured using Bio-Rad Bio-Plex kits (Bio-Rad, catalogue # 171F11181). Samples and standards were prepared using the manufacturer’s protocols with the initial concentration of standards ranging from 32 ng/ml to 1.95 pg/ml. Plasma samples were prepared for analysis by diluting 1 volume of the serum sample with three volumes of the Bio-Plex mouse sample diluent. Using the microplate readout, each cytokine level was calculated based on its own standard curve.
Aβ ELISA analysis
Hippocampal and cortex levels of soluble Aβ1–40 and Aβ1–42 were measured by enzyme-linked immunosorbent assay (ELISA). Briefly, 30 mg brain tissues were homogenized in 400 μL of RIPA buffer [100 mm Tris (pH 8.0), 150 mM NaCl, 0.5% deoxycholic acid, 1% nonyl phenoxylpolyethoxy ethanol-40, 0.2% sodium dodecyl sulfate, and 1 tablet protease inhibitor per 100 mL (S8820; Sigma, St. Louis, MO, USA)], and sonicated for 20 s on ice. Samples were then centrifuged for 30 min at 27,000 x g at 4°C, and the supernatants were transferred into new screw cap tubes. The supernatants obtained from this protocol were then stored at -80°C for determination of soluble Aβ levels using ELISA kits (KHB3482 for Aβ1–40, KHB3442 for Aβ-1–42; Invitrogen, Carlsbad, CA, USA). Standards and samples were mixed with detection antibody and loaded onto the antibody-pre-coated plate at the designated wells. After washing the unbound samples, horseradish peroxidase-conjugated antibody was added to all plates, and the substrates of were added for colorimetric reaction, which was stopped with sulfuric acid. Optical density was obtained and concentrations were calculated according to the standard curve.
Estimation of ATP
ATP contents in mouse brains were determined, as described by Tota et al. [77] using ATP colorimetric/fluorometric assay kit (Biovision, Catalog # K354-100) by following the manufacturer’s instructions.
Assessment of IL-1β, TNF-α and IL-6
Each brain section was mixed with 10 volumes of ice-cold buffer (20 mM Tris–HCl, pH 7.4) containing 0.5 mM PMSF, 0.5 mM benzamidine, 1.0 mM DTT and 1.0 mM EDTA. Total protein was mechanically dissociated from tissue using an ultrasonic cell disrupter. The sonicated samples were immediately centrifuged at 30,000 × g for 30 min at 4°C and the supernatants were removed and stored at 28°C until an ELISA was performed. Total protein concentrations of sonicated brain samples were determined by using a Bio-Rad assay kit using bovine serum albumin as the standard. The ELISAs for mouse IL-1β, TNF-α and IL-6 were performed by using the commercially available kits from Endogen (Woburn, MA, USA).
Data Analysis
Statistical analysis was performed using the software statistical package SPSS 12 (SPSS, Chicago, IL, USA). A univariate analysis of variance was performed using genotype (wild-type and transgenic), treatment (transgenic + pomegranates), and their interactions as between-individuals fixed factors. According to this, differences between treatments and genotype, or differences between transgenic and treatment were analyzed. Results are provided as mean values ± standard deviation (SD), student’s t test employed. For all tests, the level of statistical significance was set at P < 0.05.
Results
Effects of pomegranates, figs, or date palm fruits on plasma cytokine levels
As described in Material and Mmethod section, blood plasma from control and experimental animals were used to measure the levels of various cytokines (IL-2, IL-3, IL-4, IL-5, IL-9, IL-10 and Eotaxin). As shown in Fig. 1, the basal levels of these cytokines in control APPsw/Tg2576 mice were significantly increased (0.82 to 1.56 fold) in comparison to those of control wild-type mice. The levels of IL-5 and Eotaxin were almost similar in both control wild type and APPsw/Tg2576 mice. However, the elevated plasma cytokine levels in the APPsw/Tg2576 mice exposed to 4% pomegranates were significantly decreased in comparison to untreated control APPsw/Tg2576 mice (29.30 to 40.64%) followed by figs (22.27 to 30.49%) and dates (14.85 to 18.72%). Among all the cytokines, the total level of IL-10 was highest, ranging from 202.10 ± 15.47 pg/mL, in comparison to IL-2 which was 64.03 ± 4.90 pg/mL. The basal levels of plasma cytokine levels of control wild type were between 63.00 ± 4.80 to 108.02 ± 8.23 pg/mL, respectively with the exception of IL-2, which was 35.01 ± 2.67, and IL-4 which was 32.03 ± 5.03 pg/mL. Among all three fruits tested, significant decrease in plasma cytokine levels was observed in the pomegranate-supplemented diet, suggesting that pomegranate juice is more effective in decreasing pro-inflammatory cytokines and may mediate anti-inflammatory effects. This trend was consistently observed in all other cytokines measured and their amounts were significantly different from those of controls.
Fig 1. Protective effects of pomegranates, figs, or dates on the plasma levels of many cytokines.
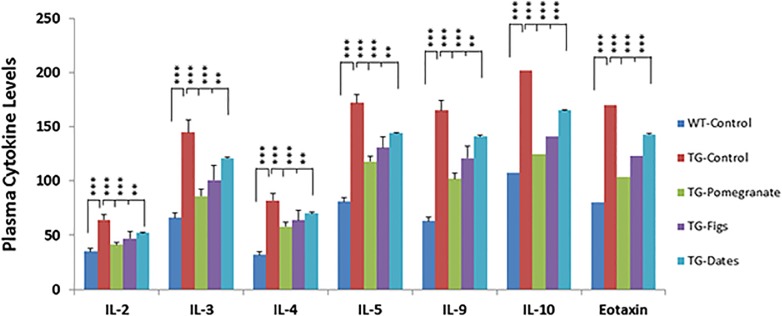
The basal plasma levels of many cytokines in WT mice and transgenic mice, fed a control diet or a diet supplemented with 4% total extracts of pomegranates, figs, or dates, as indicated, were determined by multi-plex cytokine anaylsis using the Bio-Plex kits, as described in the Materials and Methods. The representative data from at least three independent experiments are shown (** p<0.01 and *** p<0.001 vs. control TG group; n = 16/group).
Effect of pomegranates, figs, or date palm fruits on Aβ content in AD model mice
In brain, the cerebrum or the cortex is the largest portion of the brain and performs most of the brain's function. The cerebrum is divided into right and left hemispheres that are made of nerve cells which are connected by axons carrying the signals between the peripheral organs and the nerve cells. The hippocampus, an elaboration of the edge of the cerebral cortex and located in the cerebral hemisphere, is responsible for learning and memory, specifically converting temporary memories into permanent memories. These represent some regions of the brain that are susceptible to damage in neurodegenerative diseases.After measuring the levels of plasma cytokine levels, we next determined the levels of Aβ1–40 and Aβ1–42, since the accumulation of Aβ peptides activate neuro-inflammation in AD. Brain samples were collected from the untreated control or animals supplemented with pomegranates, figs or dates. The basal levels of Aβ1–40 in the cortex of control WT mice were 2105.35 ± 160.31 pg/mL and 1.36-fold higher than hippocampus (1540.26 ± 117.28 pg/mL) (Fig. 2A). Similarly, the basal levels of Aβ1–42 in the cortex of control wild-type was 1524.21 ± 95.50 pg/mL and 3.43-fold higher than hippocampus (452.08 ± 34.42 pg/mL) (Fig. 2A). The levels of Aβ1–40 in control TG (APPsw/Tg2576) were 4552.28 ± 348.45 pg/mL, which is significantly higher than that of control wild type (3906.95 ± 299.05 pg/mL) (Fig. 2A & B). The levels of Aβ1–40 in the brains of animals supplemented with the pomegranate diet decreased significantly (30.01% and 32.24%) in the cortex and hippocampus, respectively. The Aβ1–42 levels were 1956.33 ± 148.97 and 658.11 ± 50.11 pg/mL in the cortex and hippocampus, respectively. These levels in cortex and hippocampus were significantly decreased by 38.50% and 57.88% respectively (Fig. 2A & B). Supplementation of pomegranates significantly reduced the levels of Aβ1–40 and Aβ1–42 in APPsw/Tg2576 mice. This degrees of suppression of Aβ peptide levels were also observed in animals fed diets with figs or dates. The levels of Aβ1–40 in the cortex and the hippocampus in APPsw/Tg2576 mice were high and the levels of Aβ1–42 in the cortex, were ∼25–40% less than that of Aβ1–40. Interestingly, the levels of Aβ1–42 in hippocampus were estimated to be ∼60–70% less than the levels of Aβ1–40, suggesting that the Aβ1–40 peptide is the major aggregated peptide observed in AD.
Fig 2. Effects of pomegranates, figs, or dates on the Aβ and ATP levels in the cortex and hippocampus.

The levels of Aβ1–40 and Aβ1–42 in the cortex [A] and hippocampus [B] in WT and TG mice, fed a control diet or a diet supplemented with 4% total extracts of pomegranates [PGs], figs, or dates, as indicated, were determined by the ELISA as described in the Materials and Methods. [C] The % ATP levels in the cortex (Blue bars) and hippocampus (red bars) in WT or TG mice fed a control diet or a diet supplemented with pomegranates, figs, or dates are presented. The representative data from at least three independent experiments are shown (** p<0.01 and *** p<0.001 vs. control TG group; n = 16/group).
Effect of pomegranates, figs, or date palm fruits on brain ATP content of mice brains
After measuring the levels of Aβ1–40 and Aβ1–42 which serve as the biomarkers for AD, while they deplete the total ATP levels in the brain. ATP is required for numerous metabolic activities and neuronal survival. The decreased ATP levels may affect normal neuronal functions and may promote pro-inflammatory conditions by activating microglia and releasing prostanoids. Hence, to measure the ATP levels, the cortex and hippocampal regions of the brains from different groups were isolated as described in Materials and Methods. The levels of ATP in control TG mice decreased significantly (∼70%) in comparison to control wild type (Fig. 2C). However, when the animals were fed with pomegranates, figs, or dates containing diets, we observed significant recovery in ATP levels. The recovery of ATP levels ranged from 78% as observed in pomegranates followed by ∼60% (figs and ∼45% (dates), respectively. These results suggested that pomegranates could improve cerebral energy production in AD (APPsw/Tg2576) mouse model caused by aggregation of Aβ peptides (Fig. 2C).
Effect of pomegranates, figs and date palm fruits on brain IL-1β, TNF-α and IL-6 levels
After determining the levels of pro-inflammatory cytokines in the blood plasma, we measured the levels of cytokines in the brain regions, particularly in the cortex and hippocampus (Fig. 3A). Since some of the cytokines, for instance IL-6, may act as both pro- and anti-inflammatory agents, we measured the cellular levels of these cytokines including TNF-α and IL-1β. The basal levels of IL-1β in the hippocampal region were 1.40 ± 0.06 pg/mg protein the amounts in cortex were 0.60 ± 0.05 pg/mg protein, indicating about 2.33-fold greater amount in the hippocampus than that in the cortical region. However, in control APPsw (Tg2576) mice, the levels of IL-1β increased significantly, reaching 1.85 and 3.16 times greater than the basal protein levels in the cortex and hippocampus, respectively. When the animals were fed with pomegranate supplemented diet, the levels of IL-1β decreased 1.21 and 1.50 times in cortex and hippocampus, respectively, suggesting that pomegranates and other fruit supplementation could indeed reduce the levels of IL-1β and decrease neuroinflammatory activities. A similar trend was observed in the experimental animals that were fed with figs, or dates (Fig. 3A). However, the protective effects of pomegranates were more prominent, and followed by diets supplemented with figs or dates. The cellular levels of TNF-α and IL-6 in both cortex and hippocampus in Tg mice were higher as compared to those in wild control mice (Fig. 3B & C). However, the elevated TNF-α and IL-6 levels in both cortex and hippocampus in Tg mice were suppressed after the Tg mice were fed with diets supplemented with pomegranates, figs, or dates.
Fig 3. Effects of pomegranates, figs, or dates on the levels of IL-1β TNF-α and IL-6 in the cortex and hippocampus.

The levels of the three cytokinses in the WT and TG mice, as indicated, were determined by multi-plex cytokine anaylsis using the Bio-Plex kits, as described in the Materials and Methods. The representative data from at least three independent experiments are shown (** p<0.01 and *** p<0.001 vs. control TG group; n = 16/group), Pomegranates [PGs].
Discussion
Many experimental animal models for AD are available to study the pathogenesis mechanisms and translational research. For instance, the proposed model for neurodegeneration in AD brains is based on free radical mediated oxidative stress associated with Aβ1–40 and Aβ1–42 accumulation [78–79]. The role of Met-35 as a mediator of the toxicity of Aβ is more likely to involve an oxidative event at the sulfur atom, leading to lipid peroxidation and protein oxidation in neuronal membranes. However, the event that initiates the oxidation of Met-35 is not yet clear. The increased levels of Aβ in AD have been shown [80–81]. Furthermore, in an experimental mouse model of AD,greater amounts of Aβ1–42 and Aβ1–40 are started to be secreted after a few months of age and then accumulated in Tg2576 mice than their wild type control litter mates throughout their lives [82–83]. By using the Tg2576 mouse model for AD, we aimed to study the beneficial effects of antioxidants present in pomegranates, figs, or dates on a few neuro-inflammatory markers in blood plasma and brain regions. For this purpose, we specifically selected the fruits that are grown in Oman. Many studies suggest that different species from various geographical areas have diverse micronutrients and other bioactive components that may prevent or alleviate pathophysiological conditions. Our results showed significant increase in Aβ1–40 and Aβ1–42 levels both in the cortex and hippocampus. These results, consistent with the already published reports, suggest that the increased Aβ1–40 and Aβ1–42 accumulation is likely to promote oxidative stress responsible for the progression of neurodegeneration. Extended supplementation with pomegranates, figs, or dates (for 15 months) indeed decreased the Aβ1–40 and Aβ1–42 levels in the brain of Tg2576 mice in comparison to control diet-fed mice. Despite being small but significant, the observed decreases are promising, when considering the dynamic nature of Aβ present in plasma samples. Our previous studies demonstrated that dietary supplementation with pomegranates attenuates cognitive and behavioral deficits in a transgenic mouse model of AD [84].
The roles of pro-inflammatory cytokines in mediating a number of metabolic and neurological diseases are well documented. Our current results showed that the levels of pro-inflammatory cytokines, particularly IL-1β, TNF-α and IL-6 in the brains of experimental animals increased in APPsw (Tg2576) mice. The levels of IL-1β, TNF-α and IL-6 in the cerebral cortex and the hippocampus were decreased in the brains of Tg2576 mice fed diets supplemented with pomegranates, figs or dates. IL-1β, a critical cytokine in the orchestration of the complex immune response to infection and injury [85], was originally described as a peripheral immune cell mediator. This cytokine has also been reported to be synthesized in the brain by glial cells and certain neurons; and IL-1β receptors have been found in different regions of the brain, with the highest abundance in the hippocampus [86–88]. Proinflammatory cytokines including IL-1β, TNF-α and IL-6 have been reported to be significantly elevated in the cerebro-spinal fluid or plasma of AD patients [89–90]. In addition, a important role of inflammation in AD is well-supported through the inverse relationship between anti-inflammatory drug therapy and the onset and symptoms [91–92]. Griffin et al [93], reported the expression of IL-1β in different plaque types in AD, indicating that an inflammatory response plays a central role in plaque development and dystrophic neurite formation. IL-6 was present during the early stages of plaque formation and expression of this cytokine was correlated with clinical dementia [94]. IL-1β augments Aβ-peptide cytotoxicity in rat pheochromocytoma cells [95]. It has been reported that β-amyloid proteins and interferon (IFN)-δ activate microglia to produce neurotoxic TNF-α and reactive nitrogen intermediates and these events may play a role in the pathogenesis of neuronal degeneration observed in aging and AD [96].
The mechanism of the reduction of IL-1β, TNF-α and IL-6 by pomegranates is uncertain, since its multiple active components such as anthocyanins, ascorbic acid, ellagic acid, gallic acid, fumaric acid, caffeic acid, catechin, EGCG, quercetin, rutin, tannins, alkaloids and flavanoids, have multifunctional action, thus making it pharmacologically complex. Our current results, in agreement with previous reports, suggest that pomegranates in diet indeed decreased the cytokine levels [97–102]. However, the antioxidant properties of pomegranates have been well-documented. These properties include free radical scavenging and inhibition of lipid peroxidation as well as enhancement of antioxidant status [103–105] and neuroprotection [28,106–108].
Similarly, the date palm fruits also contain flavonoid glycosides of luteolin, quercetin, and apigenin. Recent research studies suggest that apigenin exhibits some mild sedative effects with anti-inflammatory properties [109–110] and neuroprotection.
Many fruits, when compared to vegetables and cereals, have very high anti-oxidant values, which are measured in terms of their "Oxygen Radical Absorbent Capacity" or ORAC. These compounds have potent anti-oxidant properties that help remove free radicals from the body, and thus provide protection against cancers, aging, and neurodegeneration. All these compounds help the body prevent or at least prolong the natural changes of aging by protecting from damage and rejuvenating cells, tissues, and organs. Including fruits in the daily diet protects from minor ailments like wrinkling of skin, hair-fall, and memory loss to major ailments like age-related macular degeneration of the retina in the eyes, neurodegenerative diseases including AD, cancers, osteoporosis [111]. Research supporting the beneficial roles of phytochemicals against cancers, coronary heart disease, diabetes, high blood pressure, inflammation, microbial, viral and parasitic infections, psychotic diseases, spasmodic conditions, ulcers, etc is based on the chemical mechanisms using in vitro cell culture systems, various disease states in animals and the epidemiology of humans [112].
Conclusion
Natural fruits, nuts, herbs and vegetables constitute a wide array of biologically active compounds including ferulic acid, anthocyanins, ellagic acid, punicagins, flavonols, phenolic acids and very important micronutrients such as phosphorus, iron, potassium and calcium, that are found abundantly in the plant kingdom. They are gaining interest due to their beneficial properties and with minimum side effects. Some of these natural products are effective in treating or preventing the majority of cardiovascular, metabolic and neurodegenerative diseases. Antioxidant activity is the key factor of all flavonoids by which they mediate the beneficial effects against the majority of many different diseases. The actions of dietary flavonoids involve a number of effects within the brain, such as modulation of neuronal signaling and the protection against neuronal losses. An extensive study on structure-function relationships of flavonoid activities provides valuable information for rationale drug designs of future pharmaceuticals in the prevention and treatment of several life-threatening diseases.
In conclusion, the pomegranates, figs, and date palm fruits grown in Oman provide possible protection against the inflammation in Tg2576 AD mouse brain and the mechanisms of protection may be related to their antioxidant activities of phenolic constituents (Fig. 4). Based on the in vivo experimental studies and the active ingredient profiles, it can be concluded that these fruits showed promising therapeutic potential against neurodegenerative diseases including AD, that areassociated with elevated inflammation. However, these results warrant further investigation of the mechanisms by which anti-inflammatory properties of these fruits can exert such beneficial effects on the brain in AD-like models.
Fig 4. Schematic diagram.

The conclusive figure showing the inflammatory signaling pathways in Alzheimer’s disease mouse model (APPsw/Tg 2576) and protection by dietary supplementation of pomegranates, figs and dates as potential complementary and alternative medicine for the neurodegenerative diseases.
Supporting Information
(XLSX)
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by a Research grant from the Research Council of Oman (RC/AGR/FOOD/11/01) to MME. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Coyle JT, Price DL, Delong MR. Alzheimer’s disease: a disorder of central cholinergic innervation. Science. 1983;219:1184–1190. [DOI] [PubMed] [Google Scholar]
- 2. Zlokovic BV. Neurovascular mechanisms of Alzheimer’s neurodegeneration. Trends Neurosci. 2005; 28, 202–208. [DOI] [PubMed] [Google Scholar]
- 3. Bell RD, Winkler EA, Sagare AP, Singh I, LaRue B, Deane R, et al. Pericytes control key neurovascular functions and neuronal phenotype in the adult brain and during brain aging. Neuron. 2010;68:409–427. 10.1016/j.neuron.2010.09.043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Marchesi VT. Alzheimer’s dementia begins as a disease of small blood vessels, damaged by oxidative-induced inflammation and dysregulated amyloid metabolism: implications for early detection and therapy. FASEB J. 2011;25:5–13. 10.1096/fj.11-0102ufm [DOI] [PubMed] [Google Scholar]
- 5. Zlokovic BV. Neurovascular pathways to neurodegeneration in Alzheimer’s disease and other disorders. Nat. Rev. Neurosci. 2011;12:723–738. 10.1038/nrn3114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Strauss S J. Bauer J, Ganter U, Jonas M, Berger B Volk BB. Detection of interleukin-6 and 2-macroglobulin immunoreactivity in cortex and hippocampus of Alzheimer’s disease patients, Lab. Invest. 1992; 66:223–230. [PubMed] [Google Scholar]
- 7. Griffin WST, Sheng JG Roberts GW, Mark RE. Interleukin-1 expression in different plaque types in Alzheimer’s disease: signifcance in plaque evolution, J. Neuropathol. Exp. Neurol. 1995; 54:276–281. [DOI] [PubMed] [Google Scholar]
- 8. Patterson PH. (1995). Cytokines in Alzheimer’s disease and multiple sclerosis, Curr. Opin. Neurobiol. 1995; 5:642–646. [DOI] [PubMed] [Google Scholar]
- 9. Schneider H, Pitossi F, Balschun D, Wagner A, del Rey A, Besedovsky HO. A neuromodulatory role of interleukin-1b in the hippocampus, Physiology. 1998;95:7778–7783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Murray CA, Lynch MA. Evidence that increased hippocampal expression of the cytokine interleukin-1b is a common trigger for age- and stress-induced impairments in long-term potentiation, J. Neurosci. 1998;18: 2974–2981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Heyser CJ, Masliah E, Samimi A, Campbell IL, Gold LH. Progressive decline in avoidance learning paralleled by inflammatory neurodegeneration in transgenic mice expressing interleukin 6 in the brain, Proc. Natl. Acad. Sci. USA 1997;94:1500–1505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Schmidt R, Schmidt H, Curb JD, Masaki K, White LR, Launer LJ. Early inflammation and dementia: a 25-year follow-up of the Honolulu-Asia Aging Study. Ann. Neurol. 2002;52:168–174. [DOI] [PubMed] [Google Scholar]
- 13. Yaffe K, Lindquist K, Penninx BW, Simonsick EM, Pahor M, Kritchevsky S, et al. Inflammatory markers and cognition in well-functioning African–American and white elders. Neurology. 2003; 61:76–80. [DOI] [PubMed] [Google Scholar]
- 14. Engelhart MJ, Geerlings MI, Meijer J, Kiliaan A, Ruitenberg A, van Swieten JC, et al. Inflammatory proteins in plasma and the risk of dementia: the rotterdam study. Arch. Neurol. 2004; 61:668–672. [DOI] [PubMed] [Google Scholar]
- 15. Dik MG, Jonker C, Hack C.E, Smit JH, Comijs HC, Eikelenboom P. Serum inflammatory proteins and cognitive decline in older persons. Neurology. 2005;64:1371–1377. [DOI] [PubMed] [Google Scholar]
- 16. Rainero I, Bo M, Ferrero M, Valfre W, Vaula G, Pinessi L. Association between the interleukin-1alpha gene and Alzheimer’s disease: a meta-analysis. Neurobiol. Aging. 2004;25:1293–1298. [DOI] [PubMed] [Google Scholar]
- 17. McGeer PL, Schulzer M, McGeer EG. Arthritis and anti-inflammatory agents as possible protective factors for Alzheimer’s disease: a review of 17 epidemiologic studies. Neurology. 1996;47:425–432. [DOI] [PubMed] [Google Scholar]
- 18. in t’Veld BA, Ruitenberg A, Hofman A, Launer LJ, van Duijn CM, Stijnen T, et al. Nonsteroidal anti-inflammatory drugs and the risk of Alzheimer’s disease. N. Engl. J. Med. 2001;345:1515–1521. [DOI] [PubMed] [Google Scholar]
- 19. van Gool WA, Aisen PS, Eikelenboom P. Anti-inflammatory therapy in Alzheimer’s disease: is hope still alive? J. Neurol. 2003;250:788–792. [DOI] [PubMed] [Google Scholar]
- 20. Muthaiyah B, Essa MM, Chauhan V, Chauhan A. Protective effects of walnut extract against amyloid beta peptide-induced cell death and oxidative stress in PC12 cells. Neurochem Res. 2011; 36(11):2096–103. 10.1007/s11064-011-0533-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Essa MM, Guillemin GJ, Al-Adawi S, Al-Asmi A, Vaishnav R, Ramachandiran N, et al. Anti amyloidogenic effect of dates with reference to their protection against Alzheimer’s disease The Dates—Genous Phoneix” Eds: Manickavasagan A, Essa MM, Sukumar E, CRC press, UK: 2010. a;397–403. [Google Scholar]
- 22. Essa MM, Vijayan RK, Castellano-Gonzalez G, Memon MA, Braidy N, Guillemin GJ. Neuroprotective Effect of Natural Products against Alzheimer's Disease. Neurochem Res. 2010. b;37(9):1829–42. [DOI] [PubMed] [Google Scholar]
- 23. Jabir R, Hasnaoui N, Mars M, Marrakchi M, Trifi M. Characterization of Tunisian pomegranate (Punica granatum L.) cultivars using amplified fragment length polymorphism analysis. Sci Hortic. 2008;115:231–237. [Google Scholar]
- 24. Melgarejo P, Martínez JJ, Hernández Fca, Martínez R, Legua P, Oncina R, et al. Cultivar identification using 18S–18S rDNA intergenic spacer-RFLP in pomegranate (Punica granatum L.). Sci. Hortic. 2009; 120:500–503. [Google Scholar]
- 25. Kelawala NS, Ananthanarayan L. Antioxidant activity of selected foodstuffs. Int J Food Sci Nutr. 2004; 55:511–516. [DOI] [PubMed] [Google Scholar]
- 26. Xu J, Guo CJ, Yang JJ, Wei JY, Li YF, Pang W, et al. Intervention of antioxidant system function of aged rats by giving fruit juices with different antioxidant capacities. Zhonghua Yu Fang Yi Xue Za Zhi. 2005;39:80–83. [PubMed] [Google Scholar]
- 27. González-Molina E, Moreno DA, García-Viguera C. A new drink rich in healthy bioactives combining lemon and pomegranate juices. Food Chemistry. 2009;115:1364–1372. [Google Scholar]
- 28. Loren DJ, Seeram NP, Schulman RN, Holtzman DM. Maternal Dietary Supplementation with Pomegranate Juice Is Neuroprotective in an Animal Model of Neonatal Hypoxic-Ischemic Brain Injury. Pediatr Res 2005;57(6):858–64. [DOI] [PubMed] [Google Scholar]
- 29. Kislev ME, Hartmann A, Bar-Yosef O. Early domesticated fig in the Jordan Valley”, Science, 2006. a312:1372–1374 [DOI] [PubMed] [Google Scholar]
- 30. Kislev ME, Hartmann A, Bar-Yosef O. Response to Comment on ‘‘Early Domesticated Fig in the Jordan Valley., Science, 2006. b;314:1683 [DOI] [PubMed] [Google Scholar]
- 31. Aljane F, Ferchichi A. Assessment of Genetic Diversity among Some Southern Tunisian Fig (Ficus carica L.) Cultivars Based on Morphological Descriptors. Jordan Journal of Agricultural Sciences. 2009;5(1): 1–16. [Google Scholar]
- 32. Lianju W, Weibin J, Kai M, Zhifeng L Yelin W. The production and research of fig (Ficus carica L.) in China. Acta Horticulturae 2003; 605, pp. 191–196. [Google Scholar]
- 33. Vinson JA, Zubik L, Bose P, Samman N. Dried fruits: excellent in vitro and in vivo antioxidants. J Am Coll Nutr 2005; 24, pp. 44–50. [DOI] [PubMed] [Google Scholar]
- 34. Solomon A, Golubowicz S, Z. Yablowicz Z, Grossman S, Bergman M, Gottlieb HE, et al. “Antioxidant activities and anthocyanin content of fresh fruits of common fig (Ficus carica L.),” Journal of Agricultural and Food Chemistry 2006; 54, 20, pp. 7717–7723. [DOI] [PubMed] [Google Scholar]
- 35. Veberic R, Colaric M, Stampar F. Phenolic acids and flavonoids of fig fruit (Ficus carica L.) in the northern Mediterranean region. Food Chemistry 2008; 106,. 1, pp. 153–157. [Google Scholar]
- 36. Vinson JA. The functional properties of figs. Cereal Foods World, 1999;44:82–87. [Google Scholar]
- 37. Kritikar KD, Basu BD. In: Blatter E., Caius I.F. (Eds.), Indian Medicinal Plants. second ed. International Book Distributors, Dehradun, 1986;2329–2331. [Google Scholar]
- 38. Solomon A, Golubowicz S, Yablowicz Z, Bergman M, Grossman S, Altman A, et al. EPR studies of O2 •−, OH, and 1O2 scavenging and prevention of glutathione depletion in fibroblast cells by cyanidin-3-rhamnoglucoside isolated from fig (Ficus carica L.) fruits. J. Agric. Food Chem. 2010. a;58:7158–7165. 10.1021/jf100153z [DOI] [PubMed] [Google Scholar]
- 39. Solomon A, Golubowicz S, Yablowicz Z, Bergman M, Grossman S, Altman A, et al. Protection of fibroblasts (NIH-3T3) against oxidative damage by cyanidin-3-rhamnoglucoside isolated from fig fruits (Ficus carica L.). J. Agric. Food Chem, 2010. b;58: 6660–6665. 10.1021/jf100122a [DOI] [PubMed] [Google Scholar]
- 40. Gilani AH, Mehmood MH, Janbaz KH, Khan A, Saeed SA. Ethnopharmacological studies on antispasmodic and antiplatelet activities of Ficus carica. J. of Ethnopharmacology 2008;119:1–5. [DOI] [PubMed] [Google Scholar]
- 41. Patil VV, Bhangale SC, Patil VR. Evaluation of anti-pyretic potential of Ficus carica leaves. Int. J. of Pharm. Sci. Rev & Res. 2010; 2 15458825 [Google Scholar]
- 42. Aref HL, Salah KBH, Chaumont JP, Fekih A, Aouni M, Said K. In vitro Antimicrobial activity of four Ficus carica latex fractions against resistant human pathogens (antimicrobial activity of Ficus carica latex). Pak J Pharm Sci. 2010; 23(1):53–58. [PubMed] [Google Scholar]
- 43. Mohamad S, Zin NM, Wahab HA, Ibrahim P, Sulaiman SF, Zahariluddin AS, et al. Antituberculosis potential of some ethnobotanically selected Malaysian plants. J Ethnopharmacol.2010;133(3):1021–1026. 10.1016/j.jep.2010.11.037 [DOI] [PubMed] [Google Scholar]
- 44. Patil VV, Bhangale SC, Patil VR. Evaluation of antipyretic potential of Ficus carica leaves. International Journal of Pharmaceutical Sciences Review and Research. 2010; 2(2):48–50. [Google Scholar]
- 45. Wang G, Wang H, Song Y, Jia C, Wang Z, Xu H. Studies on anti-HSV effect of Ficus carica leaves. Journal of Chinese medicinal materials. 2004;27:754–756. [PubMed] [Google Scholar]
- 46. Richter G, Schwarz HP, Dorner F, Peter L. Activation and inactivation of human factor X by proteases derived from Ficus carica . British Journal of Haematology.2002;119:1042–1051. [DOI] [PubMed] [Google Scholar]
- 47. Canal JR, Torres MD, Romero A, Perez CA. Chloroform extract obtained from a decoction of Ficus carica leaves improves the cholesterolaemic status of rats with streptozotocin-induced diabetes. Acta Physiologica Hungarica. 2000; 87:71–76. [DOI] [PubMed] [Google Scholar]
- 48. Rubnov S, Kashman Y, Rabinowitz R, Schlesinger M, Mechoulam R. Suppressors of cancer cell proliferation from fig (Ficus carica) resin: isolation and structure elucidation. J Nat Prod. 2001; 64(7):993–996. [DOI] [PubMed] [Google Scholar]
- 49. Yancheva SD, Golubowicz S, Yablowicz Z, Perl A, Flaishman MA. Efficient agrobacterium-mediated transformation and recovery of transgenic fig (Ficus carica L.) plants. Plant Science.2005;168(6):1433–1441. [Google Scholar]
- 50. Gond NY, Khadabadi SS. Hepatoprotective activity of Ficus carica leaf extract on rifampicin-induced hepatic damage in rats. Indian J Pharm Sci.2008;70(3):364–366. 10.4103/0250-474X.43003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Khadabadi SS, Gond NY, Ghiware NB, Shendarkar GR. Hepatoprotective effect of Ficus carica leaf in chronic hepatitis. Indian Drugs.2007; 44(1):54–57. 17277439 [Google Scholar]
- 52. Perez C, Canal JR, Campillo JE, Romero A, Torres MD. Hypotriglyceridaemic activity of Ficus carica leaves in experimental hypertriglyceridaemic rats. Phytotherapy Research.1999; 13:188–191. [DOI] [PubMed] [Google Scholar]
- 53. KrishnaMohan G, Pallavi E, RaviKumar B, Ramesh M, Venkatesh S. “Hepatoprotective activity of Ficus carica Linn leaf extract against carbon tetrachloride-induced hepatotoxicity in rats”, DARU, 2007;15: 3:162–166. [Google Scholar]
- 54. Vinson JA, Hao Y, Zubik L. Phenol antioxidant quantity and quality in foods: Vegetables. Journal of Agricultural and Food Chemistry. 1998;46:3630–3634. [Google Scholar]
- 55. Puri A, Sahai R, Singh KL, Saxena RP, Tandon JS, Saxena KC. Immunostimulant activity of dry fruits and plant materials used in Indian traditional medical system for mothers after child birth and invalids. Journal of Ethnopharmacology. 2000; 71: 89–92. [DOI] [PubMed] [Google Scholar]
- 56. Al-Maiman SA. Effect of date palm (Phoenix dactylifera) seed fibers on plasma lipids in rats”, Journal of King Saud University.2005; 17:117–123. [Google Scholar]
- 57. El-Mougy SA, Abdel-Aziz SA, Al-Shanawany M, Omar A. The gonadotropic activity of Palmae in mature male rats. Alexandria Journal of Pharmaceutical Sciences. 1991; 5:156–159. [Google Scholar]
- 58. Saafi EB, Louedi M, Elfeki A, Zakhama A, Najjar MF, Hammami M, et al. Protective effect of date palm fruit extract (Phoenix dactylifera L.) on dimethoate induced-oxidative stress in rat liver. Exp Toxicol Pathol.2011;63(5):433–441. 10.1016/j.etp.2010.03.002 [DOI] [PubMed] [Google Scholar]
- 59. Al Qarawi AA, Abdel-Rahman H, Mousa HM, Ali BH, El-Mougy SA. Nephroprotective Action of Phoenix dactylifera in gentamicin-induced nephrotoxicity. Pharmaceutical Biology.2008;46:227–230. [Google Scholar]
- 60. Ishurda O, John FK. The anti-cancer activity of polysaccharide prepared from Libyan dates (Phoenix dactylifera L.). Carbohydrate Polymers.2005; 59:531–535. [Google Scholar]
- 61. Sallal AK, Ashkenani A. Effect of date extract on growth and spore germination of Bacillus subtilis. Microbios. 1989;59:203–210. [PubMed] [Google Scholar]
- 62. Shraideh ZA, Abu-El-Teen KH, Sallal AKJ. Ultrastructural effects of date extract on Candida albicans. Mycopathologia.1998;142:119–123. [DOI] [PubMed] [Google Scholar]
- 63. Vayalil PK. Antioxidant and antimutagenic properties of aqueous extract of date fruit (Phoenix dactylifera L. arecaceae). Journal of Agricultural and Food Chemistry. 2002;50:610–617. [DOI] [PubMed] [Google Scholar]
- 64.Gamil-Abdel-Hafez M, Fouad-Shalaby A, Akhal I. Chemical composition of 15 varieties of dates grown in Saudi Arabia. Fourth Symposium on biological aspects of Saudi Arabia.1980;89–91.
- 65. Mansouri A, Embarek G, Kokkalou E, Kefalas P. Phenolic profile and antioxidant activity of the Algerian ripe date palm fruit (Phoenix dactylifera). Food Chemistry.2005;89:411–420. [Google Scholar]
- 66. Al-Farsi M, Alasalvar C, Morris A, Baron M, Shahidi F. Compositional and sensory characteristics of three native sun-dried date (Phoenix dactylifera L.) varieties grown in Oman. Journal of Agricultural and Food Chemistry. 2005;53:7586–7591. [DOI] [PubMed] [Google Scholar]
- 67. Biglari F, AlKarkhi Abbas FM, Easa AM. Antioxidant activity and phenolic content of various date palm (Phoenix dactylifera) Fruits from Iran. Food Chemistry.2008;107:1636–1641. [Google Scholar]
- 68. Allaith AAA. Antioxidant activity of Bahraini date palm (Phoenix dactylifera L.) fruit of various cultivars. International Journal of Food Science & Technology.43(6):1033–1040. [Google Scholar]
- 69. Vinson JA, Zubik L, Bose P, Samman N, Proch J. Dried fruits: Excellent in vitro and in vivo antioxidants. Journal of the American College of Nutrition. 2005;24:44–50. [DOI] [PubMed] [Google Scholar]
- 70.Al-Farsi M, Morris A, Baron M. Functional properties of Omani dates (Phoenix dactylifera). The Third International Date Palm Conference. Abu Dhabi, United Arab Emirates.2006;19–21.
- 71.Subash M, Essa MM, Al-Asmi A, Al-Adawi S, Vaishnav R, Guillemin GJ. Effect of dietary supplementation of dates in Alzheimer's disease APPsw/2576 transgenic mice on oxidative stress and antioxidant status. Nutritional neuroscience. 2014. [DOI] [PubMed]
- 72.Subash S, Essa MM, Braidy N, Al-Jabri A, Vaishnav R, Al-Adawi S, et al. Consumption of fig fruits grown in Oman can improve memory, anxiety, and learning skills in a transgenic mice model of Alzheimer's disease. Nutritional neuroscience. 2014. [DOI] [PubMed]
- 73.Subash S, Essa MM, Al-Asmi A, Al-Adawi S, Vaishnav R. Chronic Dietary Supplementation of 4% Figs on the Modification of Oxidative Stress in Alzheimer’s Disease Transgenic Mouse Model,” BioMed Research International; 2014, 8 pages. [DOI] [PMC free article] [PubMed]
- 74. Subash S, Essa MM, Al-Asmi A, Al-Adawi S, Vaishnav R, Braidy N, et al. Pomegranate from Oman Alleviates the Brain Oxidative Damage in Transgenic Mouse Model of Alzheimer's disease. Journal of Traditional and Complementary Medicine. 2014;4(4):232 10.4103/2225-4110.139107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Subash S, Essa MM, Guillemin G, Al-Adawi S, Al-Asmi A, Vaishnav A. Dietary supplementation of pomegranate reduces the brain oxidative stress in transgenic tg2576 mouse model of Alzheimer disease (1025.5) The FASEB Journal. 2013; 28: (1 Supplement), 1025.5. [Google Scholar]
- 76. Subash S, Essa MM, Al-Adawi S, Al-Asmi A, Vaishnav R, Guillemin GJ. Nine months of dietary supplementation of Omani fruits (pomegranate, figs and dates) improves the memory, anxiety and learning skills in Alzheimer's disease transgenic mice (845.1) The FASEB Journal. 2014;28:(1 Supplement), 845.1. [Google Scholar]
- 77. Tota S, Awasthi H, Kamat PK, Nath C, Hanif K. Protective effect of quercetin against intracerebral streptozotocin induced reduction in cerebral blood flow and impairment of memory in mice. Behav.BrainRes. 2010;209:73–79. 10.1016/j.bbr.2010.01.017 [DOI] [PubMed] [Google Scholar]
- 78. Varadarajan S, Yatin S, Aksenova M, Butterfield DA. Review: Alzheimer’s amyloid beta-peptide-associated free radical oxidative stress and neurotoxicity, J. Struct. Biol. 2000; 130:184–208. [DOI] [PubMed] [Google Scholar]
- 79. Butterfield DA, Castegna A, Lauderback CM, Drake J. Evidence that amyloid beta-peptide-induced lipid peroxidation and its sequelae in Alzheimer’s disease brain contribute to neuronal death, Neurobiol. Aging 2002;23:655–664. [DOI] [PubMed] [Google Scholar]
- 80. Cosentino SA, Stern Y, Sokolov E, Scarmeas N, Manly JJ, Tang MX, et al. Plasma {beta}-amyloid and cognitive decline. Arch Neurol.2010; 67:1485–90. 10.1001/archneurol.2010.189 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Laske C, Sopova K, Gkotsis C, Eschweiler GW, Straten G, Gawaz M, et al. Amyloid-beta peptides in plasma and cognitive decline after 1 year follow-up in Alzheimer’s disease patients. J Alzheimer’s Dis.2010;21:1263–9. [DOI] [PubMed] [Google Scholar]
- 82. Kuo YM, Kokjohn TA, Beach TG, Sue LI, Brune D, Lopez JC. Comparative analysis of amyloid-beta chemical structure and amyloid plaque morphology of transgenic mouse and Alzheimer’s disease brains. J Biol Chem. 2001; 276:12991–8. [DOI] [PubMed] [Google Scholar]
- 83. Kawarabayashi T, Younkin LH, Saido TC, Shoji M, Ashe KH, Younkin SG. Age-dependent changes in brain, CSF, and plasma amyloid (beta) protein in the Tg2576 transgenic mouse model of Alzheimer’s disease. J Neurosci. 2001;21:372–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Subash S, Essa MM, Al-Asmi A, Al-Adawi S, Vaishnav R, Guillemin GJ. Effect of dietary supplementation of dates in Alzheimer's disease APPsw/2576 transgenic mice on oxidative stress and antioxidant status. Nutritional Neuroscience.2014; (in press). [DOI] [PubMed]
- 85. Dinarello CA. Biologic basis for interleukin-1 in disease, Blood. 1996;87: 2095–2147. [PubMed] [Google Scholar]
- 86. Rothwell J, Hopkins SJ. Cytokines and the nervous system II: Actions and mechanisms of action, Trends Neurosci. 1995;18: 130–136. [DOI] [PubMed] [Google Scholar]
- 87. Besedovsky HO, del Rey A. Immune–neuro-endocrine interactions: facts and hypotheses, Endocr. Rev. 1996;17: 64–102. [DOI] [PubMed] [Google Scholar]
- 88. Haas HS, Schauenstein K. Neuroimmunomodulation via limbic structures. The neuroanatomy of psychoimmunology, Prog. Neuro biol. 1997;51:195–222. [DOI] [PubMed] [Google Scholar]
- 89. Fillit H, Ding WH, Buee L, Kalman J, Altstiel L, Lawlor B, et al. Elevated circulating TNF levels in Alzheimer’s disease, Neurosci. Lett. 1991;129:318–320. [DOI] [PubMed] [Google Scholar]
- 90. Blum-Degen D, Muller T, Kuhn W, Gerlach M, Przuntek H, Riederer P. IL-1βand IL-6 are elevated in the cerebrospinal fluid of Alzheimer’s and de novo Parkinson’s disease patients, Neurosci. Lett. 1995;202:17–20. [DOI] [PubMed] [Google Scholar]
- 91. Rogers J, Kirby LC, Hempelman SR, Berry DL, McGeer PL, Kaszniak AW, et al. Clinical trail of indomethacin in Alzheimer’s disease, Neurology. 1993;43:1609–1611. [DOI] [PubMed] [Google Scholar]
- 92. Breitner JCS, Gau BA, Welsh KA, Plassman BL, McDonald WM, Helms MJ, et al. Inverse association of antiinflammatory treatments and Alzheimer’s disease: initial results of a co-twin control study, Neurology. 1994; 44:; 227–232. [DOI] [PubMed] [Google Scholar]
- 93. Griffin WST, Sheng JG, Roberts GW, Mark RE. Interleukin-1 expression in different plaque types in Alzheimer’s disease: significance in plaque evolution, J. Neuropathol. Exp. Neurol. 1995;54:276–281. [DOI] [PubMed] [Google Scholar]
- 94. Huell M, Stauss S, Volk B, Berger M, Bauer J. Interleukin-6 is present in early stages of plaque formation and is restricted to the brains of Alzheimer’s disease patients, Acta Neuropathol. 1995;89:544–551. [DOI] [PubMed] [Google Scholar]
- 95. Fagarasan MO, Aisen PS. IL-1 and anti-inflammatory drugs modulate Ab cytotoxicity in PC12 cells, Brain Res. 1996;723;231–234. [DOI] [PubMed] [Google Scholar]
- 96. Meda L, Cassatella MA, Szendrei GI, Otvos LJ, Baron P, Villalba M, et al. Activation of microglial cells by b-amyloid protein and interferon-g, Nature. 1995;374: 647–650. [DOI] [PubMed] [Google Scholar]
- 97. Shukla M, Gupta K, Rasheed Z, Khan KA, Haqqi TM. Consumption of hydrolyzable tannins-rich pomegranate extract suppresses inflammation and joint damage in rheumatoid arthritis. Nutrition. 2008;24:733–743. 10.1016/j.nut.2008.03.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98. Neyrinck AM, Van-Hee VF, Bindels LB, De Backer F, Cani PD, Delzenne NM. Polyphenol-rich extract of pomegranate peel alleviates tissue inflammation and hypercholesterolaemia in high-fat diet-induced obese mice: potential implication of the gut microbiota. British Journal of Nutrition.2013;109: 802–809. 10.1017/S0007114512002206 [DOI] [PubMed] [Google Scholar]
- 99. Celik F, Gocmez C, Bozkurt M, I. Kaplan I, Kamasak K, Akil E, et al. Neuroprotective effects of carvacrol and pomegranate againstmethotrexateinduced toxicity in rats. European Review for Medical and Pharmacological Sciences.2013;17: 2988–2993. [PubMed] [Google Scholar]
- 100. de Oliveira de JFF, Garreto DV, da Silva MCP, Fortes TS, de Oliveira RB, Nascimento FRF, et al. Therapeutic potential of biodegradable microparticles containing Punica granatum L. (pomegranate) in murine model of asthma. Inflamm. Res. 2013; 62:971–980. 10.1007/s00011-013-0659-3 [DOI] [PubMed] [Google Scholar]
- 101. Asgary S, Sahebkar A, Afshani MR, Keshvari M, Haghjooyjavanmard S, Rafieian-Kopaei M. Clinical Evaluation of Blood Pressure Lowering, Endothelial Function Improving, Hypolipidemic and Anti-Inflammatory Effects of Pomegranate Juice in Hypertensive Subjects. Phytother. Res. 2014;28: 193–199. 10.1002/ptr.4977 [DOI] [PubMed] [Google Scholar]
- 102. Winand J, Schneider YJ. The anti-inflammatory effect of a pomegranate husk extract on inflamed adipocytes and macrophages cultivated independently, but not on the inflammatory vicious cycle between adipocytes and macrophages. Food Funct. 2014;5: 310 10.1039/c3fo60443h [DOI] [PubMed] [Google Scholar]
- 103. Tezcan F, Gultekin-Ozguven M, Diken T, Ozcelik B, Erim FB. Antioxidant activity and total phenolic, organic acid and sugar content in commercial pomegranate juices. Food Chemistry.2009;115:873–7. [Google Scholar]
- 104. Zhang L, Fu Q, Zhang Y. Composition of anthocyanins in pomegranate flowers and their antioxidant activity. Food Chemistry 2011;127:1444–49. [Google Scholar]
- 105. Jing P, Ye T, Shi H. Antioxidant properties and phytochemical composition of China-grown pomegranate seeds. Food Chemistry. 2012;132:1457–1464. [DOI] [PubMed] [Google Scholar]
- 106. Hartman RE, Shah A, Fagan AM, Schwetye KE, Parsadanian M, Schulman RN, et al. Pomegranate juice decreases amyloid load and improves behavior in a mouse model of Alzheimer’s disease. Neurobiol Dis 2006;24:506–15 [DOI] [PubMed] [Google Scholar]
- 107.Braidy N, Subash S, Essa MM, Vaishnav R, Al-Adawi S, Al-Asmi A, Al-Senawi H, et al. Neuroprotective effects of a variety of Pomegranate Juice extracts (PJE) against MPTP-induced cytotoxicity and oxidative stress in human primary neurons. Oxid Med Cell Longev 2013;685909. [DOI] [PMC free article] [PubMed]
- 108. Rojanathammanee L, Puig KL, Combs CK. Pomegranate polyphenols and extract inhibit nuclear factor of activated T-cell activity and microglial activation in vitro and in a transgenic mouse model of Alzheimer disease.J Nutr 2013;143(5):597–605. 10.3945/jn.112.169516 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109. Funakoshi-Tago M, Nakamura K, Tago K, Mashino T, Kasahara T. Anti-inflammatory activity of structurally related flavonoids, Apigenin, Luteolin and Fisetin Department of Biochemistry, Keio University, 1-5-30 Shibakoen, Minato-ku, Tokyo 105–8512. [DOI] [PubMed] [Google Scholar]
- 110. Patel D, Shukla S, Gupta S. Apigenin and cancer chemoprevention: progress, potential and promise Department of Urology, Case Western Reserve University, Cleveland, OH 44106, USA. [Google Scholar]
- 111. Alturki SM, Shehata WF, Mohammed I. Aldaej MI. Influence of Nutrient Medium on Antioxidants Production of Date Palm (Phoenix dactylifera L.) Cultivars in vitro . Asian Journal of Plant Sciences. 2013;12: 119–127. [Google Scholar]
- 112. Dillard CJ, German JB. Phytochemicals: nutraceuticals and human health. J. Sci. Food Agric. 2000;80: 1744–1756. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(XLSX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.