

Abstract
Aims
Extracellular ATP is an important regulator of renal tubular transport. Recently, we found that basolateral ATP markedly inhibits Na+ and Cl− absorption in mouse medullary thick ascending limb (mTAL) via a P2X receptor. The underlying mechanism that mediates this ATP-dependent transport inhibition in mTAL is, however, unclear. The renal outer medullary K+ channel (ROMK) is sensitive to intracellular pH where a reduction leads to closing of ROMK. We speculated that P2X receptor stimulation in the TAL could lead to changes in pHi, leading to a reduction in NaCl transport.
Methods
To test this hypothesis, we measured pHi in single perfused mouse mTALs using the fluorescent ratiometric dye 2′,7′-bis-(2-carboxyethyl)-5-(and-6)-carboxyfluorescein acetoxymethylester.
Results
Interestingly, basolateral ATP (100 μm) caused a prominent, reversible intracellular alkalization of mTAL, with an average pHi increase of 0.14 ± 0.02 (n = 14). This was completely abolished by the P2X receptor antagonist periodate-oxidized ATP (50 μm). The P2X receptor-mediated intracellular alkalization required the activity of the apical Na+/H+ exchanger (NHE3). Typically, Gq-coupled receptors cause a significant acidification of tubular epithelial cells, which was confirmed in this study, by P2Y2 and Ca2+ sensing receptor stimulation.
Conclusion
This study reports that stimulation of basolateral P2X receptors causes a substantial intracellular alkalization in the isolated perfused mouse mTAL. This intracellular alkalization is mediated through an increased apical NHE3 activity, similar to what we previously observed when tubular transport is inhibited with furosemide. This increased NHE3 activity causes H+ secretion in the mTAL and provides further support that the TAL is a site of urinary acidification.
Keywords: intracellular pH, loop of Henle, Na+/H+ exchanger 3, P2 receptors, purinergic
The thick ascending limb of Henle's loop (TAL) is essential for overall body water and salt homoeostasis. The TAL reabsorbs 20–30% of the filtered NaCl load and creates the osmotic gradient in the renal medulla, thereby facilitating H2O reabsorption in the collecting duct. Na+ uptake in the TAL is mediated by the Na+ K+ Cl− cotransporter NKCC2 (Greger 1985). Na+ leaves the TAL through the basolateral Na+/K+ ATPase (Greger 1985), and Cl− exits via the basolateral Cl− channel ClCkb (Greger 1985). K+ is partially recycled into the lumen by the renal outer medullary K+ channel (ROMK). Inhibition of ROMK with 3 mm Ba2+ results in a complete block of Na+ and Cl− reabsorption in TAL (Greger & Schlatter 1983). ROMK has been shown to be sensitive to intracellular pH (pHi), where a minor cytosolic acidification results in closing of the channel (Bleich et al. 1990, Choe et al. 1997, Leipziger et al. 2000). It has never been demonstrated whether the unique pHi dependence of ROMK serves the function of regulating transport rates in the TAL, that is if agonist- or antagonist-induced changes of pHi actually alter Na+ and Cl− absorption.
The autocrine and paracrine signalling pathway through purinergic (P2) receptors plays a role in the regulation of renal epithelial transport. The P2 receptors are subdivided into G-protein-coupled receptors (P2Y) and ligand-gated ion channels (P2X). Both receptor types are abundantly expressed along the nephron and are generally involved in inhibition of transport processes (Kishore et al. 1995, Lehrmann et al. 2002, Bailey 2004, Rieg et al. 2007, Pochynyuk et al. 2008). The TAL has been shown to express functional apical P2Y2 receptors and basolateral P2Y2 and P2X receptors. Stimulation of the P2 receptors of either type results in rises in [Ca2+]i (Jensen et al. 2007, Geyti et al. 2008), and extracellular ATP is known to reduce O2 consumption in TAL suspensions, which is the likely result of Na+ and Cl− transport inhibition (Silva & Garvin 2009). Indeed, transport measurements in isolated perfused medullary TAL (mTAL) show that basolateral ATP causes an inhibition of Na+ and Cl− transport that is mediated through multiple P2X receptors, including the P2X4 receptor (Marques et al. 2012). The mechanism for this transport inhibition is not yet established. In this study, we wanted to investigate whether extracellular ATP induces changes in pHi in isolated perfused mouse mTAL. It was speculated that stimulation of P2X receptors could cause an intracellular acidification, which potentially could trigger transport inhibition. In several studies that use agonists to elevate [Ca2+]i, it has been established that a rise in [Ca2+]i is associated with intracellular acidification (Berk et al. 1987, Sage et al. 1990). This is also the case for P2X receptors, where stimulation associates with an intracellular acidification (Henriksen & Novak 2003). Thus, P2X4 receptor stimulation could potentially result in a decrease in cytosolic pH, thereby closing ROMK to cause an inhibition of Na+ and Cl− transport in the TAL.
This hypothesis was proven wrong and, surprisingly, basolateral ATP caused a significant, sustained and reversible intracellular alkalization through P2X receptor stimulation in perfused mouse mTALs. The ATP-induced increase in pHi is mediated by the apical Na+/H+ exchanger 3 (NHE3) and can only be observed in an actively transporting tubule. Our data imply that ATP triggers NHE3-dependent H+ secretion through an inhibition of tubular transport. The results also reflect a potential role of the TAL in urinary acid secretion.
Materials and methods
Tubule perfusion
All mouse handling of animals complied with Danish animal welfare regulations. Animals had free access to standard rodent diet and tap water. Experiments were performed on 4- to 6-week-old mice with a mixed genetic background (B6D2/SV129). Mice were sacrificed by cervical dislocation, the kidneys collected, placed in ice-cold control solution containing (in mm) 145 NaCl, 0.4 KH2PO4, 1.6 K2HPO4, 5 d-glucose, 1 MgCl2, 1.3 Ca-gluconate and 5 N-2-hydroxyethylpiperazine-N-2-ethane-sulfonic acid (HEPES) and were subsequently sliced. The slices were placed in a dissection chamber with cold (4 °C) control solution. mTALs were isolated from the inner stripe of the outer medulla (ISOM) with ultrafine forceps. The dissected mTALs were transferred to a perfusion chamber mounted on an inverted microscope (Axiovert 100 TV; Zeiss, Jena, Germany) and perfused with a concentric pipette system as described previously (Greger & Hampel 1981). TALs were stabilized on the bath bottom with a holding pipette. Tubules were bathed and perfused with control solution from one side with the tubule outflow left open. All experiments were performed at 37 °C, and agonist and antagonist solutions were prepared fresh.
Fluorescence recording
The set-up for fluorescence microscopy consisted of an inverted microscope with a 63× C-Apochromat 1.2 water (Zeiss) objective, a VisiChrome polychromator system (Visitron, Puchheim, Germany) and a digital CCD camera (Spot pursuit 1.4 monochrome; Diagnostic Instruments, Sterling Heights, MI, USA). Images were acquired, and data analysed with standard software (visiview; Visitron). Intracellular pH was measured with the ratiometric fluorescent dye 2′,7′-bis-(2-carboxyethyl)-5-(and-6)-carboxyfluorescein acetoxymethylester (BCECF AM; Invitrogen, Carlsbad, CA, USA). Tubules were incubated with 5 μm basolateral BCECF AM in control solution for 20 min at RT during continuous perfusion with control solution, followed by a 5-min washout period. The pHi was measured as the emission ratio at 490/436 nm excitation during 5-s intervals. To reduce photo damage to the tissue, the excitation speed was 50 ms at 436 nm and 25 ms at 490 nm. A 500-nm beam splitter and a 520/560 band pass were used. Experimental manipulations were carried out after a stable fluorescence signal was achieved, and fluorescence of the entire tubule was recorded for analysis. ATP, oxidized ATP (oATP) (Sigma-Aldrich) and #4167 (kindly provided by Sanofi Aventis, Germany) were freshly dissolved in H2O prior experiments. Furosemide was dissolved in DMSO to 0.1 m stock solution and dissolved further in H2O prior to each experiment.
Calibration of the BCECF signal was performed using the high K+ and nigericin method (Thomas et al. 1979) in a paired fashion. The calibration solution was used previously in similar experiments (Watts & Good 1994, Odgaard et al. 2004) and contained (in mm) 95 KCl, 15 NaCl, 0.4 NaH2PO4, 1.6 Na2HPO4, 5 glucose, 1 MgCl2, 1.3 Ca-gluconate, 25 HEPES and 20 N-Methyl-D-glucamine, supplemented with 2 μm nigericin. The solution was set at pH 6.5, 7.4 and 7.8.
Statistics
All data are presented as mean ± SEM in all series, n indicates the number of tubules used. For each series, no more than two tubules were used from a single mouse. Data were tested for normality with the Kolmogorov–Smirnov test. Differences between experimental conditions were analysed using the paired or unpaired Student's t-test or anova where necessary. In all cases, P < 0.05 was considered significant.
Results
Basolateral ATP induces an alkalization in mTAL cells
Figure1a shows a typical pHi recording of a perfused mTAL. Under resting conditions, the pHi of this tubule was 7.28. The average pHi of the summarized data from all perfused mTALs was 7.31 ± 0.05 (n = 14, Fig.1b). Application of basolateral ATP (100 μm) caused a reversible intracellular alkalization in perfused mTAL, which was sustained during the 2-min exposure to ATP and then returned to the basal pHi after a washout period of 2–5 min. The average pHi during ATP exposure was 7.45 ± 0.06, and after 5-min washout, the pHi recovered to 7.31 ± 0.05 (Fig.1b, P < 0.0001). These data reflect that ATP causes an intracellular alkalization with a magnitude of 0.14 ± 0.02 pH units. Figure2 shows the concentration–response curve of ATP in a range from 100 nm to 500 μm. The estimated EC50 was 6.74 ± 1.67 μm. Interestingly, low concentrations of ATP (1 μm) resulted in a decrease in pHi, suggesting that basolateral ATP has a dual effect on intracellular pH in mouse mTAL.
Figure 1.
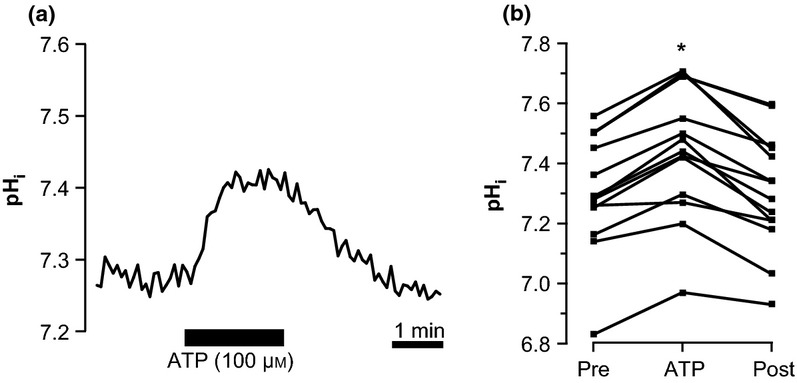
Basolateral ATP causes an intracellular alkalization in perfused medullary thick ascending limb (mTAL). (a) Representative trace of a perfused mTAL loaded with 2′,7′-bis-(2-carboxyethyl)-5-(and-6)-carboxyfluorescein acetoxymethylester (BCECF AM). ATP (100 μm) is applied to the bath for 2 min. (b) Summarized data of the experiments with 100 μm basolateral ATP, n = 14. *Indicates statistical significance (P < 0.001).
Figure 2.
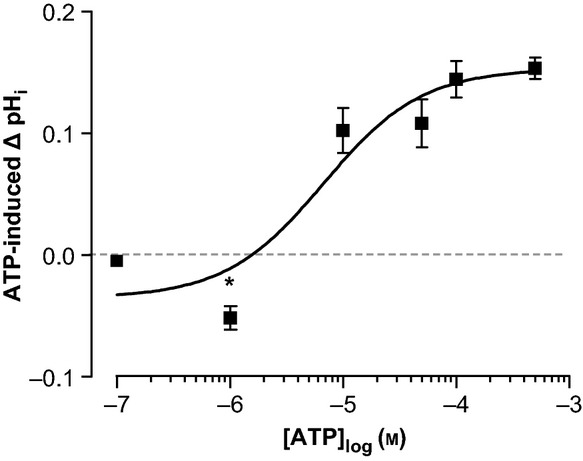
Concentration–response curve of basolateral ATP-induced alkalization in perfused medullary thick ascending limbs, n = 6–14. Values are read after 2-min ATP exposure. *Indicates statistical significance.
P2Y2 receptor stimulation causes an intracellular acidification
Our group previously demonstrated that of the P2Y receptor family, only P2Y2 and P2Y6 are expressed in murine mTAL (Marques et al. 2012). To investigate the role of P2Y receptors in the ATP-induced alkalization, we used the potent P2Y2 receptor agonist UTP that does not activate P2X receptors. Figure3a shows a typical experiment of a perfused mTAL exposed to basolateral UTP (100 μm), causing a small but significant acidification (ΔpHi 0.04 ± 0.01, n = 7, P < 0.001). UDP, a specific P2Y6 receptor agonist, did not cause any changes in pHi (results not shown). The magnitude of the UTP-induced acidification was similar to that seen when 1-μM ATP was applied to the basolateral side (ΔpHi 0.05 ± 0.01, n = 6, Fig.3b), which is sufficient to stimulate the P2Y2 receptor (Abbracchio et al. 2006). Stimulation of the Ca2+ sensing receptors (CaSR) with increased extracellular [Ca2+] (5 mm) also acidified perfused mTALs. These findings indicate that stimulation of Gq-protein-coupled receptors such as P2Y2 and CaSR in the TAL causes a small intracellular acidification.
Figure 3.
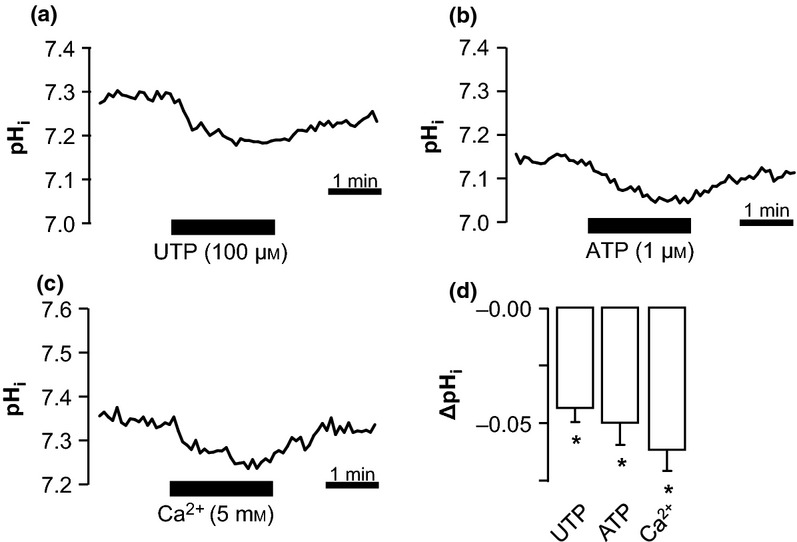
P2Y2 and Ca2+ sensing receptors (CaSR) stimulation causes an acidification in perfused medullary thick ascending limbs (mTALs). (a–c) Representative traces of perfused mTALs exposed for 2 min to basolateral UTP (100 μm), ATP (1 μm) and high Ca2+ (5 mm). (d) Summarized data of ΔpHi induced with UTP, ATP and Ca2+ (n = 4–8). *Indicates statistical significance P < 0.01.
The ATP-induced alkalization is mediated through P2X receptors
We previously demonstrated that P2X1, P2X4 and P2X5 receptors are expressed in TAL of mice (Marques et al. 2012). To confirm that the ATP-induced alkalization is mediated by P2X receptor stimulation, we used the unspecific irreversible P2X receptor antagonist oATP. Figure4a shows an original trace of a time control experiment, for two consecutive applications of basolateral ATP to a perfused mTAL separated by 12-min washout. It is clear from both Figure4a and the summarized data in Figure4b that ATP induced an alkalization of comparable size in both cases. When, however, ATP was applied after the irreversible P2X receptor antagonist oATP (50 μm), the ATP-induced alkalization was completely abolished (Fig.4c). Instead, an acidification was observed, congruent with a residual P2Y2 receptor stimulation (Fig.4d). Thus, the ATP-induced alkalization is mediated through basolateral P2X receptors.
Figure 4.
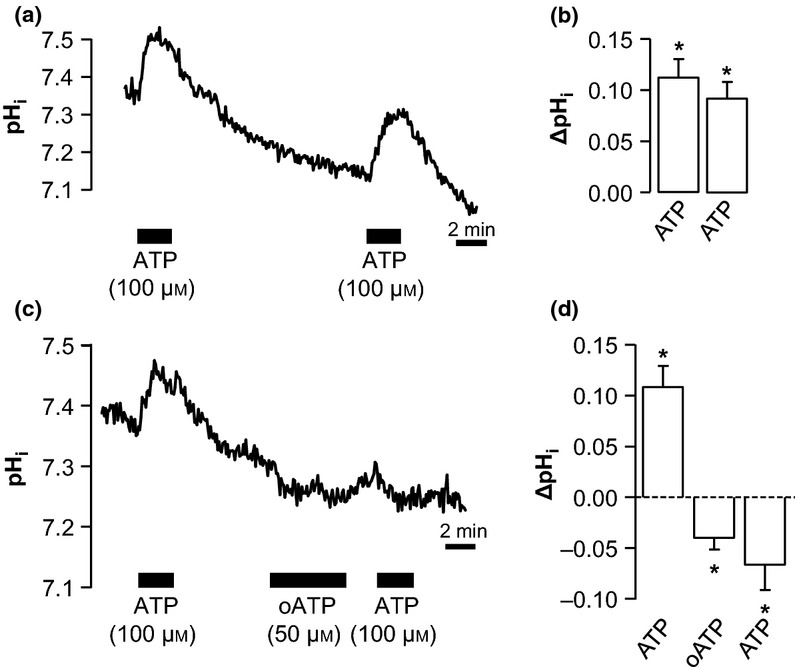
P2X receptor antagonist oxidized ATP (oATP) inhibits the ATP-induced alkalization. (a+b) Original trace and summary (ΔpHi) of time control experiments, where a perfused medullary thick ascending limb is exposed to basolateral ATP twice with a 12-min washout period in between. (c+d) Original trace and summary (ΔpHi) of experiments, where pre-incubation with oATP (50 μm, 5-min exposure) completely inhibits the ATP-induced alkalization. *Indicates statistical significance P < 0.01, n = 6.
ATP-induced alkalization is mediated through activation of apical NHE3
A recent study from our group has established that furosemide causes an intracellular alkalization in the mTAL through increased apical NHE3-mediated H+ secretion (de Bruijn et al. 2013). To investigate whether the ATP-induced intracellular alkalization occurs by the same mechanism, basolateral ATP was tested in the presence of the specific NHE3 blocker #4167 (Reuter et al. 2008). Figure5a shows that luminal #4167 (1 μm) caused a significant intracellular acidification (ΔpH −0.35 ± 0.02, n = 8, Fig.5b). During this NHE3 inhibition, the ATP-induced alkalization was completely abolished in five of eight experiments, whereas in three experiments, the alkalization was strongly attenuated. These data indicate that apical NHE3 activity is required for the ATP-induced alkalization.
Figure 5.
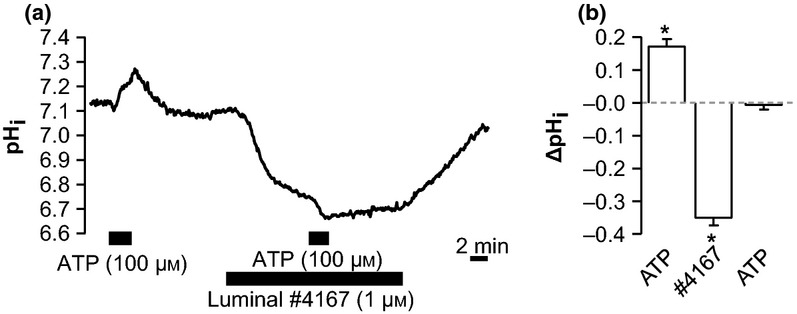
Effect of Na+/H+ exchanger (NHE3) inhibition with #4167 on ATP-induced alkalization. (a) Original experiment of perfused medullary thick ascending limb exposed to luminal #4167 (1 μm) and basolateral ATP (100 μm). (b) Summary (ΔpHi) of the series. *Indicates statistical significance P < 0.01, n = 8.
ATP-induced alkalization is abolished in the presence of furosemide
Furosemide and ATP both cause significant intracellular alkalizations in mTAL that require NHE3 activity (this study and de Bruijn et al. 2013), and they are known to reduce tubular transport in this segment (Greger 1985, Marques et al. 2012). It was therefore interesting to study whether the pHi effects of the two substances are additive. Figure6a illustrates that furosemide causes a marked intracellular alkalization from pH 7.25 ± 0.08 to pH 7.60 ± 0.02 (n = 6, Fig.6b). When basolateral ATP (100 μm) was added during the continuous presence of luminal furosemide, pHi remained unchanged. These results do not support an additive effect of these two alkalizing stimuli and thus are consistent with the notion of a common underlying cause.
Figure 6.
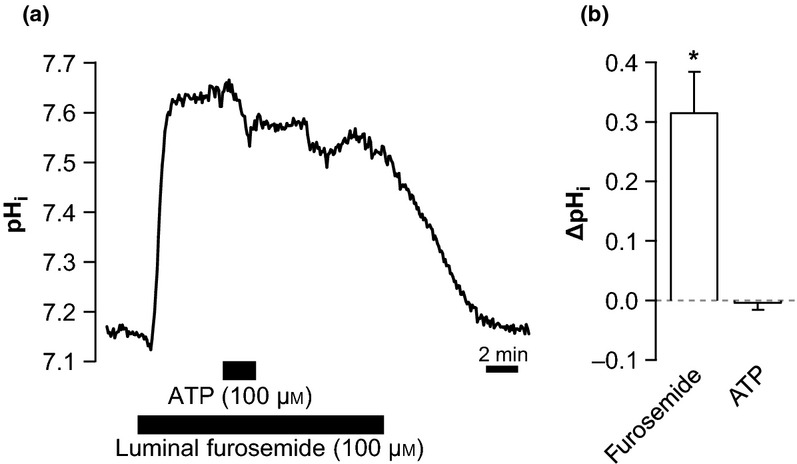
Luminal furosemide inhibits the basolateral ATP-induced alkalization. (a) Original trace of an experiment, where ATP (100 μm) is added to the bath in the presence of luminal furosemide (100 μm). (b) Summary (ΔpHi) of the data, n = 6. *Indicates statistical significance, P < 0.001.
Discussion
In this study, we describe the surprising finding that basolateral P2X receptor stimulation triggers a marked alkalization in the isolated perfused mouse mTAL. It is well established that GPCRs and ligand-gated ion channels in a variety of tissues trigger a significant intracellular acidification (Berk et al. 1987, Sage et al. 1990, Henriksen & Novak 2003). We were therefore curious to define the mechanism for the pronounced ATP-triggered alkalization. As a first step, we confirmed that P2Y2 receptor and CaSR stimulation triggered the expected acidification in the mTAL. We further showed that the ATP-stimulated alkalization occurred via basolateral P2X receptors. As the experiments were conducted in HEPES buffer, the ATP-stimulated alkalization must reflect the removal of protons from the cytosol. Finally, we demonstrated that the P2X receptor-mediated alkalization was completely inhibited by blocking the apical NHE3. In summary, our results are consistent with an ATP-activated H+ secretion into the tubular lumen via the Na+-dependent H+ exchanger NHE3 in mTAL from mice.
Clues to a mechanism of how ATP activates NHE3 can be found in a parallel report from our group (de Bruijn et al. 2013). In this work, we demonstrated that furosemide causes a prominent intracellular alkalization, which results from a sudden drop in [Na+]i that apparently leads to an increased activity of the apical NHE3 by a more favourable chemical Na+ gradient. The marked drop of [Na+]i is best explained by acute inhibition of Na+ influx via the apical NKCC2 cotransporter during continuous basolateral Na+ efflux via the Na+/K+ ATPase. Moreover, it was shown that furosemide stimulated H+ secretion into the mTAL lumen by directly measuring luminal pH. One main conclusion of this parallel work was that transport inhibition with loop diuretics triggers a marked H+ secretion and therefore urinary acidification.
In this study, we identify that basolateral P2X receptor stimulation leads to the activation of the apical NHE3. However, the cellular mechanism could not be resolved. It is well established that NHEs are activated by cell shrinkage and thus involved in regulatory volume increase (RVI, Hoffmann et al. 2009). However, NHE3 has been demonstrated to be activated by cell swelling rather than shrinkage in rat TAL (Watts & Good 1999, Good et al. 2000). Activation of the P2X receptors results in the influx of cations and, therefore, could result in cell swelling. Preliminary data confirm that ATP causes cell volume increases in perfused mTAL and show that hyposmolality result in an intracellular alkalization (unpublished data). It is thus likely that the observed intracellular alkalization caused by P2X receptor stimulation is a consequence of cell swelling-induced NHE3 activation. However, Watts and Good have shown that this swelling-induced activation of NHE3 still takes place in the presence of furosemide (Watts & Good 1999). Our data with furosemide and ATP show that NHE3 can apparently not be further stimulated by P2X receptor activation (Fig.6). More detailed studies on the activation of NHE3 in both cases are required to fully understand its regulation.
Basolaterally applied ATP is established to inhibit Na+ and Cl− absorption substantially (approx. 25%) via P2X receptors (Marques et al. 2012). The current results indicate that inhibition of transport, irrespective of the mode of induction, associates with an intracellular alkalization caused by increased H+ secretion. Taken together, these results indicate that ATP, much similar to furosemide, increases the driving force for luminal H+ exit via the NHE3 (Fig.7 for model). It is worth to note that partial transport inhibition as seen under P2X receptor stimulation causes a moderate alkalization as compared to a massive pH effect when Na+ and Cl− absorption was fully inhibited with furosemide. These results indicate that the rate of Na+ and Cl− absorption inversely correlates with the rate of H+ secretion via apical NHE3. Indeed, it has been shown that AVP, which stimulates NKCC2 activity (Welker et al. 2008, Marques et al. 2013), reduces  reabsorption, consistent with a decrease in NHE3 activity (Good 1990).
reabsorption, consistent with a decrease in NHE3 activity (Good 1990).
Figure 7.
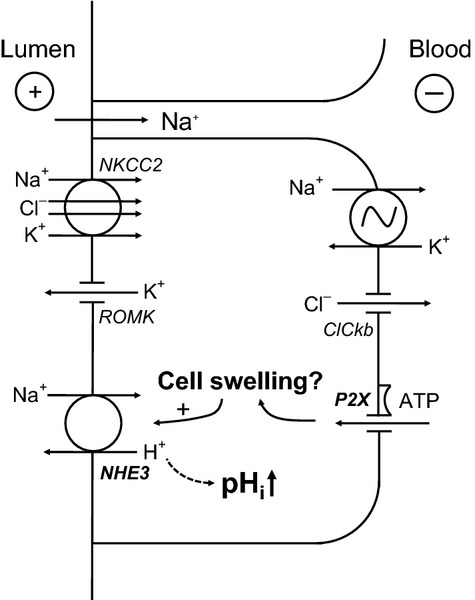
Model of P2X receptor-mediated intracellular alkalization in medullary thick ascending limb. Stimulation of the P2X receptor causes an influx of cations, which should lead to cell swelling. This in turn may stimulate the apical Na+/H+ exchanger (NHE3), leading to the observed intracellular alkalization.
The main motivation for this study was to investigate the underlying signalling mechanism that explains how basolateral P2X receptor stimulation inhibits Na+ and Cl− transport in the TAL. Currently, it was unresolved whether alterations of pHi could matter in this signalling cascade. The current through the ROMK channel is known to be significantly reduced by intracellular acidification, which potentially could lead to transport inhibition, much similar to that observed with luminal Ba2+. Clearly, our results show that P2X receptor stimulation does not cause an intracellular acidification and therefore argues against this hypothesis.
Several experimental observations argue against a critical role of intracellular acidification as important modulator of Na+ and Cl− absorption in the TAL. We show here that P2Y2 receptor and CaSR stimulation resulted in a small intracellular acidification. It is, however, unlikely that this acidification results in ROMK closure and transport inhibition, as it has been shown that acute basolateral P2Y2 receptor stimulation has no effect on Na+ and Cl− reabsorption in the TAL (Marques et al. 2013). In accordance with this, inhibition of apical Na+/H+ exchange with amiloride, which is known to cause marked intracellular acidifications in the TAL, does not change the transepithelial voltage in perfused mouse and rat mTAL (Good 1985). These results again suggest that the pHi sensitivity of ROMK may not play an important role in regulation of the transepithelial transport in TAL.
In summary, this study reports the novel finding that stimulation of basolateral P2X receptors causes a substantial intracellular alkalization in the isolated perfused mouse mTAL. Together with previous studies (Good 1990, de Bruijn et al. 2013), the cellular mechanism is described and highlights that the intracellular alkalization is mediated through an increased apical NHE3 activity. This increased NHE3 activity causes H+ secretion in the mTAL and provides further support that the TAL is a site of urinary acidification.
Conflicts of interest
None.
We greatly appreciate the expert technical assistance from Edith B. Møller and Helle Jakobsen. Funding was provided by the Danish Medical Research Council.
References
- Abbracchio MP, Burnstock G, Boeynaems JM, Barnard EA, Boyer JL, Kennedy C, Knight GE, Fumagalli M, Gachet C, Jacobson KA. Weisman GA. International Union of Pharmacology LVIII: update on the P2Y G protein-coupled nucleotide receptors: from molecular mechanisms and pathophysiology to therapy. Pharmacol Rev. 2006;58:281–341. doi: 10.1124/pr.58.3.3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bailey MA. Inhibition of bicarbonate reabsorption in the rat proximal tubule by activation of luminal P2Y1 receptors. Am J Physiol Renal Physiol. 2004;287:F789–F796. doi: 10.1152/ajprenal.00033.2004. [DOI] [PubMed] [Google Scholar]
- Berk BC, Brock TA, Gimbrone MA., Jr Alexander RW. Early agonist-mediated ionic events in cultured vascular smooth muscle cells. Calcium mobilization is associated with intracellular acidification. J Biol Chem. 1987;262:5065–5072. [PubMed] [Google Scholar]
- Bleich M, Schlatter E. Greger R. The luminal K+ channel of the thick ascending limb of Henle's loop. Pflugers Arch. 1990;415:449–460. doi: 10.1007/BF00373623. [DOI] [PubMed] [Google Scholar]
- de Bruijn PIA, Bleich M, Himmerkus N, Praetorius HA. Leipziger J. The thick ascending limb is an important site of furosemide-induced urinary acidification. J Am Soc Nephrol. 2013;24 &, TH-PO 632 (Abstract) [Google Scholar]
- Choe H, Zhou H, Palmer LG. Sackin H. A conserved cytoplasmic region of ROMK modulates pH sensitivity, conductance, and gating. Am J Physiol. 1997;273:F516–F529. doi: 10.1152/ajprenal.1997.273.4.F516. [DOI] [PubMed] [Google Scholar]
- Geyti CS, Odgaard E, Overgaard MT, Jensen MEJ, Leipziger J. Praetorius HA. Slow spontaneous [Ca2+]i oscillations reflect nucleotide release from renal epithelia. Pflugers Arch. 2008;455:1105–1117. doi: 10.1007/s00424-007-0366-4. [DOI] [PubMed] [Google Scholar]
- Good DW. Sodium-dependent bicarbonate absorption by cortical thick ascending limb of rat kidney. Am J Physiol. 1985;248:F821–F829. doi: 10.1152/ajprenal.1985.248.6.F821. [DOI] [PubMed] [Google Scholar]
- Good DW. Inhibition of bicarbonate absorption by peptide hormones and cyclic adenosine monophosphate in rat medullary thick ascending limb. J Clin Invest. 1990;85:1006–1013. doi: 10.1172/JCI114530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Good DW, Di Mari JF. Watts BA., III Hyposmolality stimulates Na(+)/H(+) exchange and HCO(3)(−) absorption in thick ascending limb via PI 3-kinase. Am J Physiol Cell Physiol. 2000;279:C1443–C1454. doi: 10.1152/ajpcell.2000.279.5.C1443. [DOI] [PubMed] [Google Scholar]
- Greger R. Ion-transport mechanisms in thick ascending limb of Henle's loop of mammalian nephron. Physiol Rev. 1985;65:760–797. doi: 10.1152/physrev.1985.65.3.760. [DOI] [PubMed] [Google Scholar]
- Greger R. Hampel W. A modified system for in vitro perfusion of isolated renal tubules. Pflugers Arch. 1981;389:175–176. doi: 10.1007/BF00582110. [DOI] [PubMed] [Google Scholar]
- Greger R. Schlatter E. Properties of the lumen membrane of the cortical thick ascending limb of Henle's loop of rabbit kidney. Pflugers Arch. 1983;396:315–324. doi: 10.1007/BF01063937. [DOI] [PubMed] [Google Scholar]
- Henriksen KL. Novak I. Effect of ATP on intracellular pH in pancreatic ducts involves P2X7 receptors. Cell Physiol Biochem. 2003;13:93–102. doi: 10.1159/000070253. [DOI] [PubMed] [Google Scholar]
- Hoffmann EK, Lambert IH. Pedersen SF. Physiology of cell volume regulation in vertebrates. Physiol Rev. 2009;89:193–277. doi: 10.1152/physrev.00037.2007. [DOI] [PubMed] [Google Scholar]
- Jensen MEJ, Odgaard E, Christensen MH, Praetorius HA. Leipziger J. Flow-induced [Ca2+]i increase depends on nucleotide release and subsequent purinergic signaling in the intact nephron. J Am Soc Nephrol. 2007;18:2062–2070. doi: 10.1681/ASN.2006070700. [DOI] [PubMed] [Google Scholar]
- Kishore BK, Chou CL. Knepper MA. Extracellular nucleotide receptor inhibits AVP-stimulated water permeability in inner medullary collecting duct. Am J Physiol. 1995;269:F863–F869. doi: 10.1152/ajprenal.1995.269.6.F863. [DOI] [PubMed] [Google Scholar]
- Lehrmann H, Thomas J, Kim SJ, Jacobi C. Leipziger J. Luminal P2Y(2) receptor-mediated inhibition of Na+ absorption in isolated perfused mouse CCD. J Am Soc Nephrol. 2002;13:10–18. doi: 10.1681/ASN.V13110. [DOI] [PubMed] [Google Scholar]
- Leipziger J, MacGregor GG, Cooper GJ, Xu J, Hebert SC. Giebisch G. PKA site mutations of ROMK2 channels shift the pH dependence to more alkaline values. Am J Physiol Renal Physiol. 2000;279:F919–F926. doi: 10.1152/ajprenal.2000.279.5.F919. [DOI] [PubMed] [Google Scholar]
- Marques RD, de Bruijn PIA, Sorensen MV, Bleich M, Praetorius HA. Leipziger J. Basolateral P2X receptors mediate inhibition of NaCl transport in mouse medullary thick ascending limb (mTAL) Am J Physiol Renal Physiol. 2012;302:F487–F494. doi: 10.1152/ajprenal.00570.2011. [DOI] [PubMed] [Google Scholar]
- Marques RD, Praetorius HA. Leipziger J. P2Y2 receptor knock-out mice display normal NaCl absorption in medullary thick ascending limb. Front Physiol. 2013;4:280. doi: 10.3389/fphys.2013.00280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Odgaard E, Jakobsen JK, Frische S, Praetorius J, Nielsen S, Aalkjaer C. Leipziger J. Basolateral Na+-dependent HCO3− transporter NBCn1-mediated HCO3− influx in rat medullary thick ascending limb. J Physiol. 2004;555:205–218. doi: 10.1113/jphysiol.2003.046474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pochynyuk O, Bugaj V, Rieg T, Insel PA, Mironova E, Vallon V. Stockand JD. Paracrine regulation of the epithelial Na+ channel in the mammalian collecting duct by purinergic P2Y2 receptor tone. J Biol Chem. 2008;283:36599–36607. doi: 10.1074/jbc.M807129200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reuter S, Velic A, Edemir B, Schroter R, Pavenstadt H, Gabriels G, Bleich M. Schlatter E. Protective role of NHE-3 inhibition in rat renal transplantation undergoing acute rejection. Pflugers Arch. 2008;456:1075–1084. doi: 10.1007/s00424-008-0484-7. [DOI] [PubMed] [Google Scholar]
- Rieg T, Bundey RA, Chen Y, Deschenes G, Junger W, Insel PA. Vallon V. Mice lacking P2Y2 receptors have salt-resistant hypertension and facilitated renal Na+ and water reabsorption. FASEB J. 2007;21:3717–3726. doi: 10.1096/fj.07-8807com. [DOI] [PubMed] [Google Scholar]
- Sage SO, Jobson TM. Rink TJ. Agonist-evoked changes in cytosolic pH and calcium concentration in human platelets: studies in physiological bicarbonate. J Physiol. 1990;420:31–45. doi: 10.1113/jphysiol.1990.sp017900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silva GB. Garvin JL. Extracellular ATP inhibits transport in medullary thick ascending limbs: role of P2X receptors. Am J Physiol Renal Physiol. 2009;297:F1168–F1173. doi: 10.1152/ajprenal.00325.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas JA, Buchsbaum RN, Zimniak A. Racker E. Intracellular pH measurements in Ehrlich ascites tumor cells utilizing spectroscopic probes generated in situ. Biochemistry. 1979;18:2210–2218. doi: 10.1021/bi00578a012. [DOI] [PubMed] [Google Scholar]
- Watts BA., III Good DW. Apical membrane Na+/H+ exchange in rat medullary thick ascending limb. pH-dependence and inhibition by hyperosmolality. J Biol Chem. 1994;269:20250–20255. [PubMed] [Google Scholar]
- Watts BA., III Good DW. Hyposmolality stimulates apical membrane Na(+)/H(+) exchange and HCO(3)(−) absorption in renal thick ascending limb. J Clin Invest. 1999;104:1593–1602. doi: 10.1172/JCI7332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Welker P, Bohlick A, Mutig K, Salanova M, Kahl T, Schluter H, Blottner D, Ponce-Coria J, Gamba G. Bachmann S. Renal Na+-K+-Cl− cotransporter activity and vasopressin-induced trafficking are lipid raft-dependent. Am J Physiol Renal Physiol. 2008;295:F789–F802. doi: 10.1152/ajprenal.90227.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]