

Abstract
Danshen, in particular its derivative tanshinone IIA (TS), is a promising compound in the treatment of cardiovascular diseases and has been used for many years in traditional Chinese medicine. Although many actions of TS have been researched, its vasodilator effects in pregnancy remain unknown. There have been a few studies that have shown the ability of TS to reduce blood pressure in women with hypertensive pregnancies; however, there are no studies which have examined the vascular effects of TS in the pregnant state in either normal or complicated pregnancies. Our aim was to determine the vasoactive role of TS in multiple arteries during pregnancy including: rat resistance (mesenteric and uterine) and conduit (carotid) arteries. Further, we aimed to assess the ability of TS to improve uterine blood flow in a rodent model of intrauterine growth restriction. Wire myography was used to assess vascular responses to the water-soluble derivative, sodium tanshinone IIA sulphonate (STS) or to the endothelium-dependent vasodilator, methylcholine. At mid-pregnancy, STS caused direct vasodilation of rat resistance (pEC50 mesenteric: 4.47±0.05 and uterine: 3.65±0.10) but not conduit (carotid) arteries. In late pregnancy, human myometrial arteries responded with a similar sensitivity to STS (pEC50 myometrial: 3.26±0.13). STS treatment for the last third of pregnancy in eNOS-/- mice increased uterine artery responses to methylcholine (Emax eNOS-/-: 55.2±9.2% vs. eNOS-/- treated: 75.7±8.9%, p<0.0001). The promising vascular effects, however, did not lead to improved uterine or umbilical blood flow in vivo, nor to improved fetal biometrics; body weight and crown-rump length. Further, STS treatment increased the uterine artery resistance index and decreased offspring body weight in control mice. Further research would be required to determine the safety and efficacy of use of STS in pregnancy.
Introduction
Salvia miltiorrhiza, also known as Danshen, is a promising and increasingly researched traditional Chinese herbal medicine that has been widely used for many years to treat various disorders including coronary artery disease, cerebrovascular disease, myocardial infarction, angina pectoris, stroke and atherosclerosis [1]. Danshen has many components, including approximately 30 lipophilic diterpenes and >15 hydrophilic phenolic acid derivatives [2, 3]. Of the former category, tanshinone IIA (TS) is one of the most pharmacologically active and widely investigated. Studies have uncovered an impressive array of actions of TS including, but not limited to: vasodilation, inhibition of inflammatory mediators, oxidative stress and matrix metalloproteinases-2 and -9, as well as scavenging of peroxyl radicals and reduction of cardiac hypertrophy (reviewed in [4]). Due to its traditional cardiovascular uses, much of the vascular function work performed to date has been centered on the effects of TS on the coronary vasculature or the development of atherosclerosis. TS has been shown to cause vasodilation of mouse, rat and pig coronary arteries that was endothelium-, nitric oxide (NO)-, epoxyeicosatrienoic acid (EET)- or large-conductance, calcium-activated potassium channel (BKCa)-dependent [5–9]. TS also reduced atherosclerotic plaques and lesions in the aortae of ApoE-/- mice through a reduction of superoxide production, oxidized LDL, cholesterol and pro-inflammatory cytokines [10, 11]. In addition, researchers have shown TS-induced vasodilation of thoracic aortae from Sprague Dawley rats that was NO- and estrogen-dependent [12]. Although the effect of Danshen, a preparation of the whole root, has been studied in coronary [5–9] and femoral [13, 14] arteries, the role of the isolated TS component, or its water-soluble derivative sodium tanshinone IIA sulphonate (STS), in vascular resistance has not been extensively investigated. Our own group has shown STS-induced vasodilation of mesenteric arteries from male Sprague Dawley rats that was partially mediated by small- and intermediate-conductance calcium activated potassium channels (SKCa and IKCa) but had no reliance on the NO pathway [15]. The precise mechanisms of (S)TS-induced vasodilation are, therefore, currently not fully elucidated.
To date, there have been three clinical studies which showed positive maternal effects of TS on hypertension in pregnancy [16–18]. In these studies, TS treated subjects demonstrated a reduction in morbidity, mean arterial pressure and blood viscosity, cholesterol and lipoprotein. There are no studies, however, which have examined the vascular effects of TS in the pregnant state in either normal or complicated pregnancies, particularly with regard to intrauterine growth restriction (IUGR). IUGR represents a failure of the fetus to reach its genetically determined potential size. IUGR affects up to 10% of all pregnancies worldwide [19, 20] and currently has no treatment. IUGR fetuses and the surviving infants have lifelong health issues including, among others, cardiovascular and metabolic complications (reviewed in [21, 22]). IUGR can occur when the transport mechanisms of nutrients and oxygen to the fetus are compromised. There are several up-stream causes of impaired nutrient transfer to the fetus including maternal, fetal and placental factors (reviewed in [23, 24]). Adaptations of the maternal cardiovascular system, such as an increase of uterine artery blood flow, allow for increased transfer of nutrients across the placenta, however, an increased uterine artery resistance has been associated with IUGR [25].
In light of its effects on oxidative stress, inflammatory state and vasodilation, (S)TS may provide a potential treatment strategy for compromised pregnancies which involve pathological changes in these areas. We hypothesized that STS would cause vasodilation of maternal resistance arteries, including the mesenteric and uterine vasculature, in pregnancy. Further, we hypothesized that treatment with STS would improve uterine artery vasodilation in an animal model of complicated pregnancy; the eNOS-/- mouse model of intrauterine growth restriction, via actions on non-NO mediated vasodilator pathways.
Methods
Ethical approval
All protocols were approved by the University of Alberta Health Sciences Animal Policy and Welfare Committee in accordance with the guidelines of the Canadian Council on Animal Care and the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health.
I. Direct effects of STS—rodent studies
Three month old female Sprague-Dawley rats (Charles River, Wilmington, MA) were maintained on ad libitum standard rodent chow and tap water in a 10:14 hour light:dark cycle. Females were acclimatized in-house before breeding. Gestational day (GD) 0 was determined by the presence of sperm in a vaginal smear following an overnight introduction of a male. Rats (total n = 17) were euthanized by exsanguination via excision of the superior vena cava under inhaled isoflurane anaesthesia on GD 10.9±0.2 (term 22 days); corresponding to mid-gestation—an important clinical intervention time-point. Tissue dissections were performed in ice-cold physiological saline solution (PSS), composition (in mmol/l): 10 HEPES, 5.5 glucose, 1.56 CaCl2, 4.7 KCl, 142 NaCl, 1.17 MgSO4, 1.18 KH2PO4, pH 7.5. Small mesenteric (n = 64 artery sections from n = 14 animals) and main uterine branch (n = 56 artery sections from n = 13 animals) arteries were isolated for experimental procedures. The common carotid artery (n = 16 artery sections from n = 8 animals) was also assessed as a reference conduit artery to ascertain vascular bed specificity.
Arteries were cleaned of all surrounding adipose and connective tissues and 2 mm sections were mounted on two 40 μm wires attached to a wire myograph (DMT, Copenhagen, SV, Denmark) to allow isometric tension recordings. Vessels were normalized through a series of stepwise increases in diameter to determine their optimal resting tension, set to 0.8 x IC100 (the internal circumference equivalent to a transmural pressure of 100 mmHg).
Following a 30-minute equilibration period, vessels were twice exposed to a single dose of phenylephrine (PE, 10 μmol/l) followed by a single dose of methylcholine (MCh, 3 μmol/l) to check functional endothelial and smooth muscle integrity. A cumulative concentration-response curve to the adrenergic agonist phenylephrine (PE, 0.1 to 30 μmol/l, mesenteric and uterine arteries) or the thromboxane mimetic U46619 (U19, 0.01 to 3 μmol/l, carotid arteries) was performed. The EC80 (the effective concentration producing 80% of the maximum response) for each vasoconstrictor/artery combination was then determined.
Vascular responses to STS (1 to 100 μmol/l) were investigated following preconstriction with the EC80 concentration of the relevant vasoconstrictor. STS responses were performed in the absence or presence of inhibitors to investigate some of the possible vasodilator mechanisms. The three main endothelium-dependent vasodilator pathways; nitric oxide, prostaglandins and endothelial derived hyperpolarisation (EDH), were investigated using inhibitors of nitric oxide synthase (NOS) (N-nitro-L-arginine methyl ester hydrochloride, L-NAME, 100 μmol/l), cyclooxygenase (meclofenamate, 1 μmol/l), or a combination of apamin (100 nmol/l) and TRAM-34 (10 μmol/l) to block SKCa and IKCa channels, respectively. The inhibitors used are well-established and doses were chosen based on published methods [26–29].
In a separate series of experiments, the endothelium of mesenteric and uterine arteries was denuded using a knotted, human hair. The lack of endothelium was confirmed by assessment of responses to MCh (3 μmol/l). Vascular responses to STS (1 to 100 μmol/l) were then assessed following preconstriction with the EC80 concentration of the relevant vasoconstrictor.
II. Direct effects of STS—human studies
Proof of principle studies were performed in a small number of human arteries (n = 12 artery sections from n = 3 patients) to determine if STS has vasodilator effects in human tissues from a relevant vascular bed. All human tissues were obtained from patients attending the Royal Alexander Hospital. All patients provided informed written consent to participate and the procedure was approved following full ethics review by the Alberta Health Services Ethics Committee, Edmonton. Signed consent forms were retained as documentation of participant consent. A myometrial biopsy from the uterus was obtained during scheduled caesarean sections of healthy pregnancies, mean gestational age 38 weeks and 6 days (st.dev. 4 days), without labour. The biopsy was immediately placed into ice-cold modified Kreb’s solution for collection and transportation. The tissue arrived in the laboratory and myometrial vessels were isolated within 60 minutes of delivery. Isolation of vessels and subsequent experimental protocols were performed in PSS as per animal vessels. Responses to the thromboxane mimetic U46619 (U19, 0.01 to 3 μmol/l) were investigated to assess the EC80 following which vascular responses to STS (1 to 100 μmol/l) were determined.
III. In vivo administration of STS in mice
eNOS-/- (strain B6.129P2-Nos3 tm1Unc/J) mice were chosen as an established model of IUGR that has been previously used in our laboratory. These mice have been shown to have increased blood pressure, decreased uterine artery blood flow and growth restricted offspring, both by ourselves and others [30–33]. C57BL/6J (control, n = 13) and eNOS-/- (Jackson Laboratories, Bar Harbor, ME, n = 13) mice were maintained on ad libitum standard rodent chow and tap water in a 10:14 hour light:dark cycle. Females were acclimatized in-house before breeding. GD 0 of pregnancy was determined by the presence of a vaginal plug following an overnight introduction of a male. A subset of control (n = 8) and eNOS-/- (n = 7) mice were treated with STS in their drinking water (approx. 27 mg/kg/day) from GD 12 to 18 (term 19 days). The dosage of STS was chosen following a literature review which determined treatment regimens of 80 mg/day in human (approx. 1.1 mg/kg/day) [34, 35] and ovine (approx. 1.8 mg/kg/day) [36, 37] trials and a median of 20 mg/kg/day in rodent studies [5, 10, 38–41]. The timing of treatment was chosen as a clinically relevant time-point during which intervention would be possible in humans.
Systolic blood pressure was measured by a validated tail-cuff system (IITC Life Science, CA, USA) on GD 17. Uterine and umbilical artery blood flow velocities were assessed in vivo in control and eNOS-/- mice under anaesthesia on GD 17 using an ultrasound biomicroscope (model Vevo 2100, VisualSonics®, ON, Canada) as previously described [42, 43]. Briefly, uterine artery Doppler waveforms were obtained from both the left and right uterine arteries, and from the umbilical arteries of at least two fetuses. Peak systolic velocity (PSV) and end diastolic velocity (EDV) were measured from at least three cardiac cycles and the results were averaged. The uterine and umbilical vascular resistance indices were calculated from PSV and EDV measures.
On GD 18, mice were euthanized by exsanguination via cardiac puncture under inhaled isoflurane anaesthesia and the pregnancy outcome was determined by weighing and measuring the pups. The main uterine branch arteries were dissected in ice-cold PSS and prepared for experimental procedures on a wire myograph as described for studies in rat vessels. Following a 30-minute equilibration period, vessels were exposed to PE (10 μmol/l) and MCh (3 μmol/l) to check functional endothelial and smooth muscle integrity. Responses to the adrenergic agonist PE (0.0001 to 10 μmol/l) were performed to determine the EC80. Following constriction with PE, responses to the endothelium-dependent vasodilator MCh (0.0001 to 10 μmol/l) or the nitric oxide donor, sodium nitroprusside (SNP, 0.0001 to 10 μmol/l) in the absence or presence of L-NAME (100 μmol/l) were investigated.
Statistical Analysis
All vascular function data were presented as mean ± standard error of the pEC50 (negative log of the effective concentration that will produce 50% of the maximum response) or the Emax (maximum response). Phenotypical parameters from ultrasound biomicroscopy, blood pressure measurements and offspring biometrics were presented as mean ± standard error. All data were normally distributed as assessed by the Kolmogorov-Smirnov test for Gaussian distribution. The significance of the difference in mean values of continuous variables between groups was determined by a one- or two-way analysis of variance (ANOVA), with Bonferroni post-test for multiple groups. A p value < 0.05 was considered statistically significant.
Results
Mechanisms of direct vascular responses to STS
In mid-gestation rats, STS caused vasodilation of mesenteric resistance arteries (Fig. 1). Uterine arteries from mid-gestation rats demonstrated a biphasic response to STS. An initial contractile response (Emax: -28.1 ± 9.4%) was followed by a vasodilator response that was significantly less sensitive than that in mesenteric arteries (p<0.0001, Fig. 1). Maximal vasodilation of mesenteric arteries and uterine arteries to STS were not significantly different. In the carotid arteries, STS caused only a vasoconstrictor response.
Fig 1. Direct vasodilator effects of STS in rat arteries.
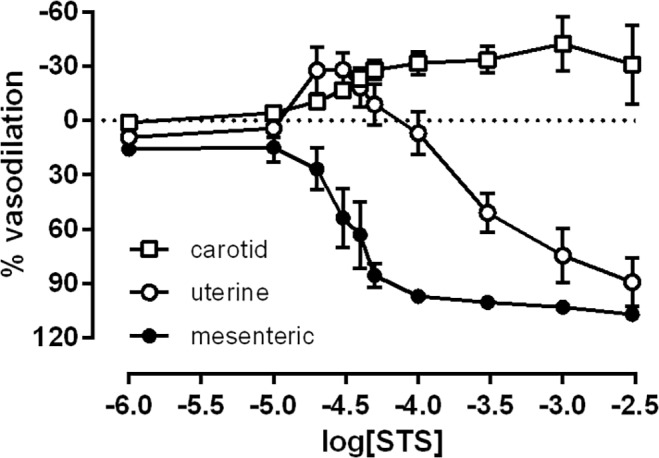
Vascular responses to STS in the carotid (open squares, n = 8), uterine (open circles, n = 10), and mesenteric arteries (closed circles, n = 8) from the pregnant rat (GD 10.9±0.2).
Inclusion of either the NOS inhibitor L-NAME or inhibition of COX using meclofenamate did not significantly affect STS-induced vasodilation of mesenteric arteries from mid-gestation rats. The presence of the potassium channel blockers apamin and TRAM-34, however, significantly inhibited STS-induced vasodilation compared to the control group (p<0.0001, Fig. 2A-B). In uterine arteries, inclusion of inhibitors for any of the three main endothelium-dependent pathways caused no significant change in STS-induced responses: including the biphasic vasoconstrictor and vasodilator components (Fig. 2C-D).
Fig 2. Involvement of endothelium-dependent vasodilator mechanisms in direct STS responses.

Mechanisms of vascular responses to STS in mesenteric (A&C) and uterine (B&D) arteries from the pregnant rat (GD 10.9±0.2). Vasodilator responses to STS in the absence (closed circles) or presence of inhibitors of nitric oxide synthase (NOS) (N-nitro-L-arginine methyl ester hydrochloride, L-NAME, 100 μmol/l, open squares), cyclooxygenase (meclofenamate, 1 μmol/l, open circles) or a combination of apamin (100 nmol/l) and TRAM-34 (10 μmol/l) which block SKCa and IKCa channels respectively (open triangles) in A: mesenteric arteries and B: uterine arteries. Sensitivity (negative log of the effective concentration producing 50% of the maximal response) of vascular responses to STS in C: mesenteric arteries or D: uterine arteries. Data analysed by one-way ANOVA with a Bonferroni post-test; ††: p<0.01 vs. control vessel. Mesenteric: control n = 8; L-NAME n = 6; Apa, TRAM n = 7; Meclo n = 4. Uterine: control n = 10; L-NAME n = 6; Apa, TRAM n = 7; Meclo n = 5.
Endothelium removal in mesenteric arteries resulted in a significant reduction in maximal vasodilator responses to MCh of 62.1 ± 7.4% (p<0.0001). Despite a 2.6-fold reduction in endothelial function, there was no significant change in sensitivity or maximal responses to STS following endothelium removal (Fig. 3A). Endothelium removal in uterine arteries also resulted in a significant reduction in maximal vasodilator responses to MCh of 80.0 ± 9.6% (p<0.0001). Despite a 5.7-fold reduction in endothelial function, there was no significant change in sensitivity or maximal responses to STS (Fig. 3B).
Fig 3. Endothelium-independent responses to STS.
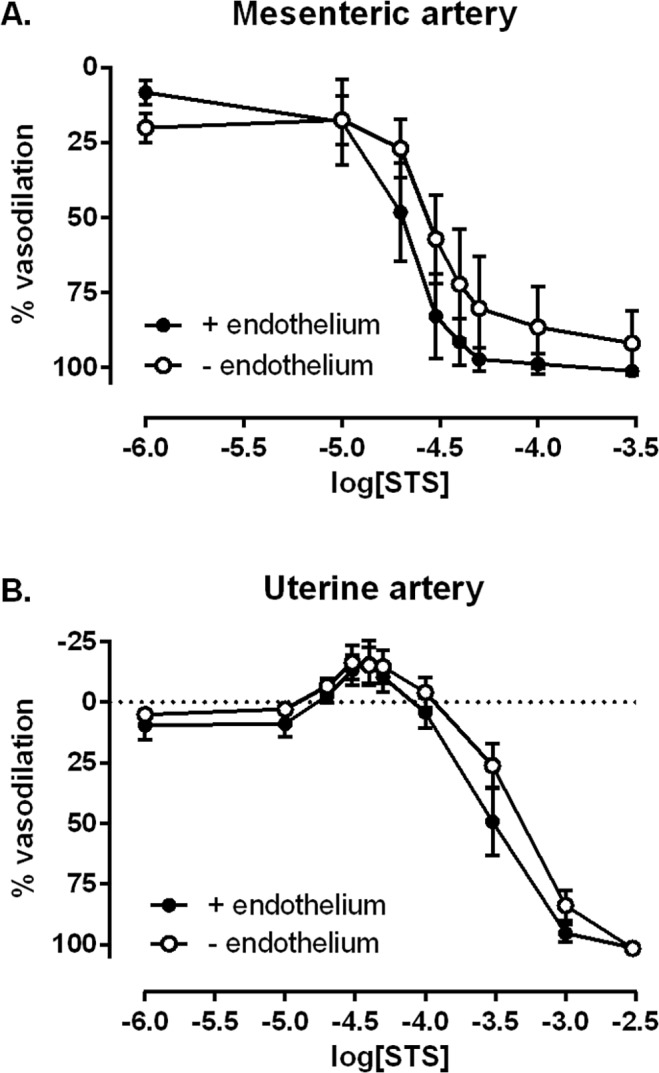
Vascular responses to STS in A: mesenteric arteries and B: uterine arteries from the pregnant rat (GD 10.9±0.2) with intact endothelium (closed circles) or following endothelial removal using a knotted human hair (open circles). Mesenteric intact n = 6; mesenteric denuded n = 5; uterine intact n = 4; uterine denuded n = 5.
Human myometrial arteries responded to STS with a vasodilator response that was of similar potency (pEC50: 3.26 ± 0.13) and efficacy (Emax: 89.07 ± 3.42%) to that observed in female pregnant rat uterine arteries. Human arteries, however, did not demonstrate the contractile component of the biphasic response observed in rat uterine arteries.
Effect of in vivo STS treatment on vascular responses
Isolated mouse uterine arteries from GD 18 demonstrated reduced PE-induced vasoconstriction in eNOS-/- mice that was increased following six days of STS treatment (Emax control untreated: 2.04 ± 0.31 mN/mm vs. treated: 2.94 ± 0.15 mN/mm; eNOS-/- untreated 1.49 ± 0.13 mN/mm vs. treated 1.88 ± 0.19 mN/mm; group genotype effect p<0.001, group treatment effect p<0.01). MCh-induced vasodilation was also significantly reduced in eNOS-/- mice compared to control mice (p<0.0001, Fig. 4A-B). Treatment with STS increased uterine arteries responses to MCh in both control (p<0.001) and eNOS-/- mice (p<0.0001) (Fig. 4A-C). The increase in MCh-induced vasodilation was shown to be NO-independent using the NOS inhibitor, L-NAME (data not shown). In addition, SNP-induced vasodilation (pEC50 control: 7.36 ± 0.18 vs. eNOS-/-: 7.70 ± 0.13) was not altered in eNOS-/- compared to control mice nor was it affected by STS treatment (pEC50 control treated: 7.50 ± 0.08; eNOS-/- treated: 7.89 ± 0.11).
Fig 4. Effect of STS treatment on vasodilator responses in mouse arteries.
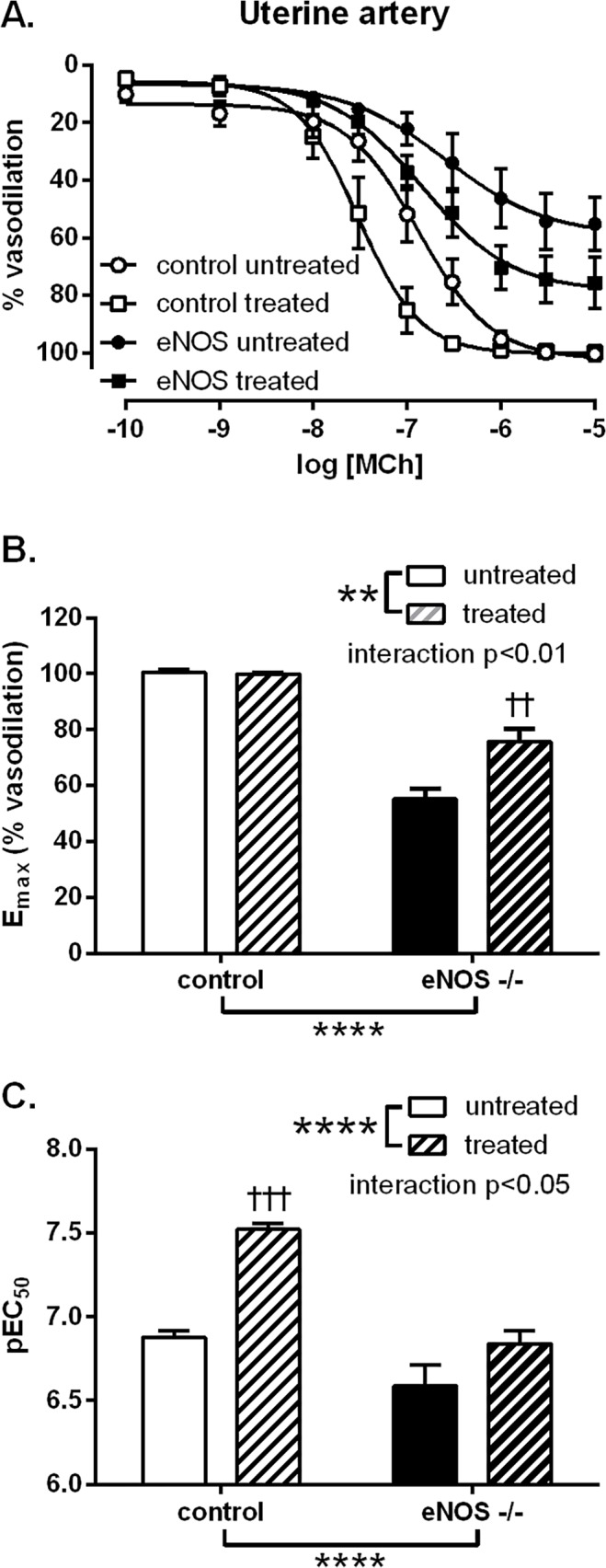
A: Vascular responses of mouse uterine arteries to MCh from control (circles) and eNOS-/- (squares) late pregnant (GD 18) mice, untreated (closed symbols) or treated (open symbols) with STS from GD 12 to 18. STS was given in drinking water at a dose of approx. 27 mg/kg/day. Summary data of B: maximal responses and C: sensitivity (negative log of the effective concentration producing 50% of the maximal response) to MCh. Data analysed by two-way ANOVA with a Bonferroni post-test; **: p<0.01 group treatment effect, ****: p<0.0001 group genotype effect, ††: p<0.01 vs. eNOS-/- untreated, †††: p<0.001 vs. control untreated. Control untreated n = 5; control treated n = 5; eNOS-/- untreated n = 6; eNOS-/- treated n = 4.
Following the observation of direct STS-induced vasodilation and STS-mediated upregulation of MCh-induced vasodilation of the uterine vasculature, the ability of STS to improve uterine and umbilical artery blood flow and the outcomes of compromised pregnancy in an animal model of IUGR was tested. In vivo, the eNOS-/- mice had a reduced umbilical artery end diastolic velocity (p<0.001, Fig. 5A) and peak systolic velocity (p<0.001, Fig. 5B), and a corresponding increase in the resistance index (p<0.01, Fig. 5C) for this vascular bed compared with control mice. Treatment with STS did not improve umbilical artery blood flow velocities in control or eNOS-/- groups. In the uterine vasculature, these measures of blood flow were unaffected by the eNOS-/- phenotype (peak diastolic velocity control: 525.2 ± 111.0 mm/s vs. eNOS-/-: 395.4 ± 28.5 mm/s; end diastolic velocity control: 283.8 ± 62.1 mm/s vs. eNOS-/-: 226.9 ± 20.3 mm/s). Contrary to our hypothesis, STS treatment led to a slight increase in the uterine artery resistance index (control untreated: 0.46 ± 0.02 vs. treated: 0.51 ± 0.02; eNOS-/- control 0.43 ± 0.02 vs. treated 0.51 ± 0.04; group treatment effect p<0.05).
Fig 5. Effect of STS treatment on umbilical artery blood flow velocity in mice.
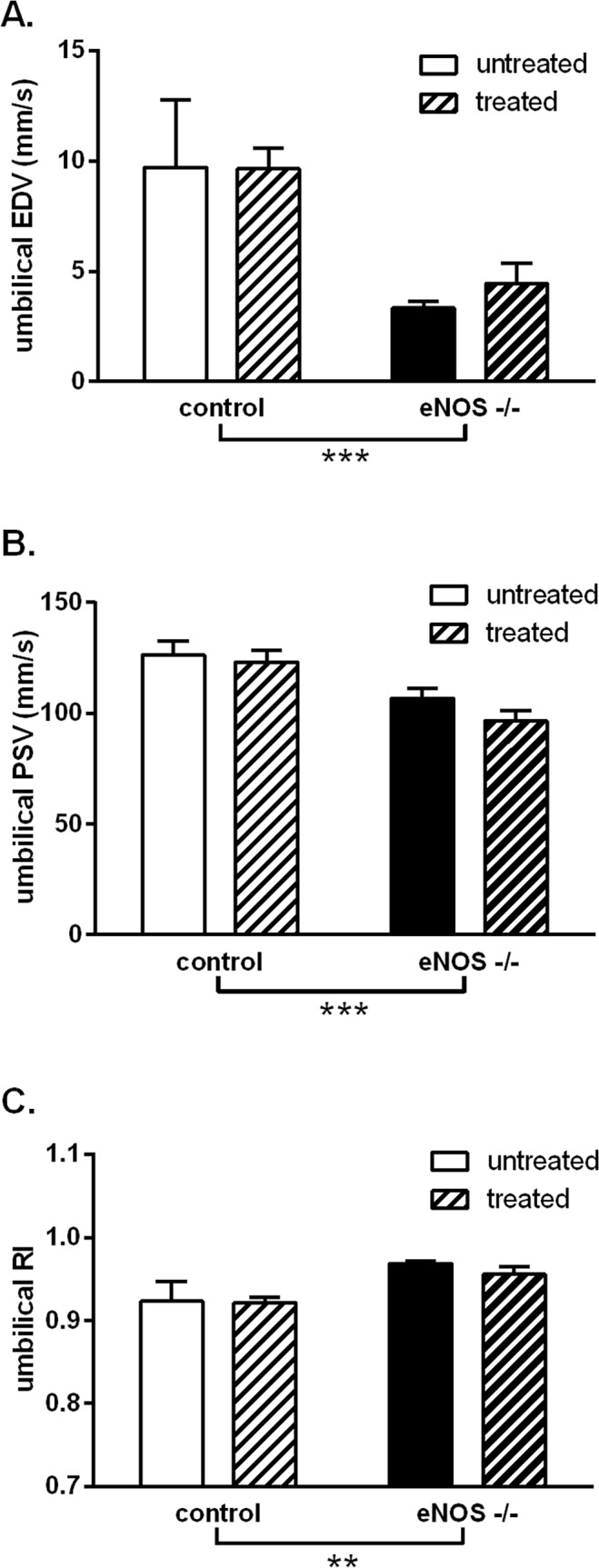
Echocardiographic parameters of umbilical blood flow from control (white bars) and eNOS-/- (black bars) late pregnant (GD 18) mice, untreated (open bars) or treated (hatched bars) with STS from GD 12 to 18. STS was given in drinking water at a concentration of approx. 27 mg/kg/day. A: end diastolic velocity (EDV), B: peak systolic velocity (PSV) and C: calculated resistance index (RI). Data analysed by two-way ANOVA with a Bonferroni post-test; **: p<0.01, ***: p<0.001, group genotype effect. Control untreated n = 4; control treated n = 8; eNOS-/- untreated n = 6; eNOS-/- treated n = 8.
In terms of pregnancy outcomes, the eNOS-/- genotype caused a reduction in offspring birth weight (p<0.0001, Fig. 6A) and crown to rump length (p<0.0001, Fig. 6B) compared to control pregnancies. There was no change in maternal systolic blood pressure (Fig. 6C) or placental weight (Fig. 6D) with mouse genotype. Treatment with STS did not improve fetal growth in eNOS-/- mice and, contrary to our hypothesis, STS treatment significantly reduced birth weight in control mice. STS treatment had no effect on maternal blood pressure or placental weight.
Fig 6. Effect of STS treatment on pregnancy outcomes in mice.

Pregnancy outcomes in control (white bars) and eNOS-/- (black bars) late pregnant (GD 18) mice, untreated (open bars) or treated (hatched bars) with STS from GD 12 to 18. STS was given in drinking water at a concentration of approx. 27 mg/kg/day. A: pup weight, B: pup crown to rump length, C: maternal systolic blood pressure and D: placental weight. Data analysed by two-way ANOVA with a Bonferroni post-test; **: p<0.01, ***: p<0.001, group genotype or treatment effects; ††: p<0.01 vs. control untreated. Control untreated n = 5(33); control treated n = 8(60); eNOS-/- untreated n = 6(30); eNOS-/- treated n = 7(43); where n = dams(pups).
Discussion
This study demonstrated a direct vasodilator effect of STS in vascular resistance arteries from pregnant rats. We have also demonstrated that STS was effective not only in dilating uterine arteries from the pregnant rat but also similarly dilates human myometrial arteries; providing proof of principle that STS may be effective in human pathology. Further, we showed that STS was able to improve the vasodilator capacity of uterine arteries when given in vivo to pregnant mice. The effect of STS on the vasculature, however, did not lead to improved uterine or umbilical blood flow in vivo or improved pregnancy outcomes.
STS caused non-NO dependent vasodilation of rat resistance (mesenteric and uterine) but not conduit (carotid) arteries. This was in line with our own studies in male rat mesenteric arteries [44] but in contrast to previous studies in the thoracic aorta and coronary circulations [8, 9, 12]; which demonstrated partially NO-mediated and endothelium-dependent vasodilation. In our study, STS-induced vasodilation in the mesenteric arteries was shown to be partly due to the small- and/or intermediate-conductance calcium-activated potassium channels (SKCa and IKCa respectively). Since the STS response was also shown to be endothelium-independent, these potassium channels were likely to be located on the smooth muscle. Interestingly, responses to STS in the mesenteric resistance vasculature from pregnant rats were consistent in efficacy, potency and sensitivity to apamin and TRAM-34 with our previous studies demonstrating vasodilator responses to STS in young male rats [44], suggesting that STS has sex-independent effects on the vasculature.
The effect of STS on the uterine vasculature during the pregnant state was of greater interest. Responses of rat uterine arteries to STS had a distinctly biphasic character that has also been observed in a previous study of rat pulmonary arteries [45]. In contrast to the mesenteric arteries, but in line with the pulmonary study, STS-induced responses were non-SKCa and—IKCa, non-NO and non-prostanoid dependent; leaving their mechanism of action unknown. Consistent with the mesenteric arteries, responses in uterine arteries were also endothelium-independent. As anticipated following the observation of direct STS-induced vasodilation of rat uterine arteries, STS treatment was also shown to improve MCh-induced vasodilation of mouse uterine arteries. Improvement of vasodilation was observed in both the control and eNOS-/- strains; providing additional support to the conclusion that STS-mediated effects on vasodilation are NO-independent.
The effect of STS was also assessed in the closest available human equivalent to the rodent uterine artery, the myometrial artery. Excitingly, not only were vasodilator responses observed, these responses in the myometrial artery were consistent in efficacy and potency to those observed in the rat uterine artery suggesting that the effects of STS were translatable from rat to human arteries.
A vasodilator of the uterine vasculature during pregnancy has potential clinical applicability, particularly for disorders that involve a decrease in placental and fetal perfusion. In particular, the endothelium-independence of STS responses provides a potential therapeutic approach in conditions characterized by endothelial dysfunction, such as preeclampsia or IUGR. Therefore, we next tested the ability of in vivo STS treatment to improve vascular function in a rodent model of IUGR. While there was increased uterine artery vasodilation following STS treatment of control and eNOS-/- mice, we did not observe improved blood flow in the uterine or umbilical arteries during pregnancy nor did the vascular effects lead to improved fetal outcomes in either genotype. While the effect of in vivo STS treatment on isolated uterine artery function provides evidence that the treatment regimen was effective systemically, the multiple actions of STS and an integrated response of the tissues in vivo may have affected the outcome which was observed.
Several concerns regarding the use of STS in pregnancy were raised in regards to its effect on the control animals. STS treatment caused a biphasic response in rat uterine arteries which included almost 30% constriction prior to the vasodilator response. Further, adrenergic-mediated vasoconstriction in mouse uterine arteries was increased by STS treatment. These observations might suggest a risk for maternal hypertension or decreased uterine blood flow as a result of a hyper-constrictive phenotype. The increased adrenergic constriction following STS treatment, however, normalized the reduced PE constriction observed in untreated eNOS-/- mice compared with untreated control mice. In addition, in our in vivo model, neither maternal systolic blood pressure nor uterine blood flow velocity were affected in either group of mice treated with STS. A previous study in pregnant sheep has demonstrated no maternal cardiovascular changes but a slight, significant increase in fetal systolic blood pressure in response to maternal treatment [46]; suggesting that the fetal effects of STS treatment need to be further explored. In our study, treatment with STS did cause a slight increase in the uterine resistance index in control and eNOS-/- mice, however, the physiological significance of the magnitude of this change is questionable. Reassuringly, constriction to STS was not observed in human myometrial arteries and, therefore, might be a species specific effect. In our study, STS treatment caused a significant reduction in the body weight of control offspring, raising a further concern for its use during pregnancy. Further characterization of STS responses in human vessels and in vivo studies of the effect of STS in pregnancy and on the offspring would be necessary to determine the safety and efficacy of the in vivo use of STS during pregnancy.
In summary, we have demonstrated vasodilation to the Danshen derivative, STS, in resistance arteries that was consistent between rodent and human arteries. The in vivo effect of STS during healthy and complicated pregnancies, however, warrants further investigation. As discussed above, the non-NO, non-endothelium dependent mechanisms of STS-induced vasodilation observed in the current study may be of benefit in conditions which are known to be associated with endothelial dysfunction; such as preeclampsia and IUGR. Further research, however, is necessary to fully elucidate the effect of STS on uterine and feto-placental blood flow; particularly in regard to its effect on fetal outcomes. Given the current lack of treatments for pregnancy complications such as IUGR, the use of a novel compound such as STS that has been historically employed for other pathological conditions could provide a solution in an area of critical need.
Data Availability
All relevant data are within the paper.
Funding Statement
S. T. Davidge is a Canada Research Chair (CRC) in Maternal and Perinatal Cardiovascular Health. The Davidge laboratory receives funding from the Canadian Institutes of Health Research (CIHR, Grant numbers: 118160540; 0020054) and the Women and Children’s Health Research Institute through the generous support of the Stollery Children's Hospital Foundation and the Royal Alexandra Hospital Foundation. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Chen J, Chen T. Dan Shen (Radix Salviae Miltiorrhizae) In: Crampton L, editor. Chinese Medical Herbology and Pharmacology: Art of Medicine Press; 2003. [Google Scholar]
- 2. Wang X, Morris-Natschke SL, Lee K-H. New developments in the chemistry and biology of the bioactive constituents of tanshen. Medicinal Research Reviews. 2007;27(1):133–48. 10.1002/med.20077 [DOI] [PubMed] [Google Scholar]
- 3. Zhou L, Zuo Z, Chow MSS. Danshen: An Overview of Its Chemistry, Pharmacology, Pharmacokinetics, and Clinical Use. The Journal of Clinical Pharmacology. 2005;45(12):1345–59. 10.1177/0091270005282630 [DOI] [PubMed] [Google Scholar]
- 4.Gao S, Liu Z, Li H, Little PJ, Liu P, Xu S. Cardiovascular actions and therapeutic potential of tanshinone IIA. Atherosclerosis.10.1016/j.atherosclerosis.2011.06.041(0). 10.1016/j.atherosclerosis.2011.06.041 [DOI] [PubMed]
- 5. Pan C, Lou L, Huo Y, Singh G, Chen M, Zhang D, et al. Salvianolic acid B and Tanshinone IIA attenuate myocardial ischemia injury in mice by NO production through multiple pathways. Therapeutic Advances in Cardiovascular Disease. 2011;5(2):99–111. 10.1177/1753944710396538 [DOI] [PubMed] [Google Scholar]
- 6. Tan X, Yang Y, Cheng J, Li P, Inoue I, Zeng X. Unique action of sodium tanshinone II-A sulfonate (DS-201) on the Ca2+ dependent BKCa activation in mouse cerebral arterial smooth muscle cells. European journal of pharmacology. 2011;656(1–3):27–32. 10.1016/j.ejphar.2011.01.028 [DOI] [PubMed] [Google Scholar]
- 7. Wan AKS, Leung SWS, Zhu D-Y, Man RYK. Vascular Effects of Different Lipophilic Components of “Danshen”, a Traditional Chinese Medicine, in the Isolated Porcine Coronary Artery. Journal of Natural Products. 2008;71(11):1825–8. 10.1021/np800119k [DOI] [PubMed] [Google Scholar]
- 8. Wu G-b, Zhou E-x, Qing D-x. Tanshinone IIA elicited vasodilation in rat coronary arteriole: Roles of nitric oxide and potassium channels. European journal of pharmacology. 2009;617(1–3):102–7. 10.1016/j.ejphar.2009.06.046 [DOI] [PubMed] [Google Scholar]
- 9. Yang Y, Cai F, Li P-Y, Li M-L, Chen J, Chen G-L, et al. Activation of high conductance Ca2+-activated K+ channels by sodium tanshinoneII-A sulfonate (DS-201) in porcine coronary artery smooth muscle cells. European journal of pharmacology. 2008;598(1–3):9–15. 10.1016/j.ejphar.2008.09.013 [DOI] [PubMed] [Google Scholar]
- 10. Tang F-T, Cao Y, Wang T-Q, Wang L-J, Guo J, Zhou X-S, et al. Tanshinone IIA attenuates atherosclerosis in ApoE−/− mice through down-regulation of scavenger receptor expression. European journal of pharmacology. 2011;650(1):275–84. 10.1016/j.ejphar.2010.07.038. 10.1016/j.ejphar.2010.07.038 [DOI] [PubMed] [Google Scholar]
- 11. Xu S, Little PJ, Lan T, Huang Y, Le K, Wu X, et al. Tanshinone II-A attenuates and stabilizes atherosclerotic plaques in Apolipoprotein-E knockout mice fed a high cholesterol diet. Archives of Biochemistry and Biophysics. 2011;515(1–2):72–9. 10.1016/j.abb.2011.08.006 [DOI] [PubMed] [Google Scholar]
- 12. Fan G, Zhu Y, Guo H, Wang X, Wang H, Gao X. Direct Vasorelaxation by a Novel Phytoestrogen Tanshinone IIA Is Mediated by Nongenomic Action of Estrogen Receptor Through Endothelial Nitric Oxide Synthase Activation and Calcium Mobilization. Journal of Cardiovascular Pharmacology. 2011;57(3):340–7 10.1097/FJC.0b013e31820a0da1 Available: http://journals.lww.com/cardiovascularpharm/Fulltext/2011/03000/Direct_Vasorelaxation_by_a_Novel_Phytoestrogen.10.aspx. [DOI] [PubMed] [Google Scholar]
- 13. Lam FFY, Yeung JHK, Cheung JHY. Mechanisms of the Dilator Action of Danshen (Salvia miltiorrhiza) on Rat Isolated Femoral Artery. Journal of Cardiovascular Pharmacology. 2005;46(3):361–8. Available: http://journals.lww.com/cardiovascularpharm/Fulltext/2005/09000/Mechanisms_of_the_Dilator_Action_of_Danshen.18.aspx. [DOI] [PubMed] [Google Scholar]
- 14. Lam FFY, Yeung JHK, Cheung JHY, Or PMY. Pharmacological Evidence for Calcium Channel Inhibition by Danshen (Salvia miltiorrhiza) on Rat Isolated Femoral Artery. Journal of Cardiovascular Pharmacology. 2006;47(1):139–45 10.1097/01.fjc.0000197540.12685.ce Available: http://journals.lww.com/cardiovascularpharm/Fulltext/2006/01000/Pharmacological_Evidence_for_Calcium_Channel.20.aspx. [DOI] [PubMed] [Google Scholar]
- 15. Liu J, Morton J, Miedzyblocki M, Lee TF, Bigam DL, Fok TF, et al. Sodium tanshinone IIA sulfonate increased intestinal hemodynamics without systemic circulatory changes in healthy newborn piglets. American Journal of Physiology—Heart and Circulatory Physiology. 2009;297(4):H1217–H24. 10.1152/ajpheart.00477.2009 [DOI] [PubMed] [Google Scholar]
- 16. Lin JH, Yang YK, Liu H, Lin QD, Zhang WY, Cooperation Group on Special Project "Study on the prevention and treatment for hypertension disorders and hematopexis related complications in pregnancy". Effect of antioxidants on amelioration of high-risk factors inducing hypertensive disorders in pregnancy. Chinese medical journal. 2010;123(18):2548–54. [PubMed] [Google Scholar]
- 17. Liu SY, Xu YY, Zhu JY. The effects of Salvia miltiorrhizae Bge and Ligustrazine on thromboxane A2 and prostacyclin in pregnancy induced hypertension. Zhonghua fu chan ke za zhi. 1994;29(11):648–50. [PubMed] [Google Scholar]
- 18. Wang XF, Zhao MQ. Ligustrazine and Salvia miltiorrhiza injection solution in complementary therapy of pregnancy-induced hypertension: clinical analysis of 60 cases. Di Yi Jun Yi Da Xue Xue Bao. 2003;23(9):969–71. [PubMed] [Google Scholar]
- 19. Romo A, Carceller R, Tobajas J. Intrauterine growth retardation (IUGR): epidemiology and etiology. Pediatric endocrinology reviews: PER. 2009;6 Suppl 3:332–6. Epub 2009/05/16. PubMed . [PubMed] [Google Scholar]
- 20. Saleem T, Sajjad N, Fatima S, Habib N, Ali SR, Qadir M. Intrauterine growth retardation—small events, big consequences. Italian Journal of Pediatrics. 2011;37:41-. 10.1186/1824-7288-37-41 PubMed . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Visentin S, Grumolato F, Nardelli GB, Di Camillo B, Grisan E, Cosmi E. Early origins of adult disease: Low birth weight and vascular remodeling. Atherosclerosis. 2014;237(2):391–9. 10.1016/j.atherosclerosis.2014.09.027. 10.1016/j.atherosclerosis.2014.09.027 [DOI] [PubMed] [Google Scholar]
- 22. Giussani DA, Davidge ST. Developmental programming of cardiovascular disease by prenatal hypoxia. J Dev Orig Health Dis. 2013;4(05):328–37. 10.1017/S204017441300010X [DOI] [PubMed] [Google Scholar]
- 23. Kingdom JC, Kaufmann P. Oxygen and placental villous development: origins of fetal hypoxia. Placenta. 1997;18(8):613–21; discussion 23–6. Epub 1997/11/19. PubMed . [DOI] [PubMed] [Google Scholar]
- 24. Lager S, Powell TL. Regulation of nutrient transport across the placenta. Journal of pregnancy. 2012;2012:179827 Epub 2013/01/11. 10.1155/2012/179827 PubMed ; PubMed Central PMCID: PMCPmc3523549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Carbillon L. First trimester uterine artery Doppler for the prediction of preeclampsia and foetal growth restriction. Journal of Maternal-Fetal and Neonatal Medicine. 2012;25(7):877–83. 10.3109/14767058.2011.601364 PubMed . [DOI] [PubMed] [Google Scholar]
- 26. Rees DD, Palmer RM, Schulz R, Hodson HF, Moncada S. Characterization of three inhibitors of endothelial nitric oxide synthase in vitro and in vivo. British Journal of Pharmacology. 1990;101(3):746–52. Available: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1917753/. PubMed . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Boctor AM, Eickholt M, Pugsley TA. Meclofenamate sodium is an inhibitor of both the 5-lipoxygenase and cyclooxygenase pathways of the arachidonic acid cascade in vitro. Prostaglandins, Leukotrienes and Medicine. 1986;23(2–3):229–38. 10.1016/0262-1746(86)90190-3. [DOI] [PubMed] [Google Scholar]
- 28. Hsieh Y-C, Chang P-C, Hsueh C-H, Lee YS, Shen C, Weiss JN, et al. Apamin-Sensitive Potassium Current Modulates Action Potential Duration Restitution and Arrhythmogenesis of Failing Rabbit Ventricles. Circulation: Arrhythmia and Electrophysiology. 2013;6(2):410–8. 10.1161/circep.111.000152 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Wulff H, Miller MJ, Hänsel W, Grissmer S, Cahalan MD, Chandy KG. Design of a potent and selective inhibitor of the intermediate-conductance Ca2+-activated K+ channel, IKCa1: A potential immunosuppressant. Proceedings of the National Academy of Sciences. 2000;97(14):8151–6. 10.1073/pnas.97.14.8151 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Hefler LA, Tempfer CB, Moreno RM, O'Brien WE, Gregg AR. Endothelial-derived nitric oxide and angiotensinogen: blood pressure and metabolism during mouse pregnancy. American Journal of Physiology Regulatory, Integrative and Comparative Physiology. 2001;280(1):R174–R82. Available: http://ajpregu.physiology.org/ajpregu/280/1/R174.full.pdf. [DOI] [PubMed] [Google Scholar]
- 31. Kulandavelu S, Whiteley KJ, Qu D, Mu J, Bainbridge SA, Adamson SL. Endothelial Nitric Oxide Synthase Deficiency Reduces Uterine Blood Flow, Spiral Artery Elongation, and Placental Oxygenation in Pregnant Mice. Hypertension. 2012;60(1):231–8. 10.1161/hypertensionaha.111.187559 [DOI] [PubMed] [Google Scholar]
- 32. Poudel R, Stanley JL, Rueda-Clausen CF, Andersson IJ, Sibley CP, Davidge ST, et al. Effects of resveratrol in pregnancy using murine models with reduced blood supply to the uterus. PLOS one. 2013;8(5):e64401 Epub 2013/05/15. 10.1371/journal.pone.0064401 PubMed [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Rueda-Clausen CF, Stanley JL, Thambiraj DF, Poudel R, Davidge ST, Baker PN. Effect of Prenatal Hypoxia in Transgenic Mouse Models of Preeclampsia and Fetal Growth Restriction. Reproduction Science. 2013;10.1177/1933719113503401. Epub 2013/10/03. 10.1177/1933719113503401 PubMed 24084523. [DOI] [PMC free article] [PubMed]
- 34. Li CF, Gou WL, Han Z. [Investigation on treatment of fetal growth restriction by salvia injection combined with composite amino acid]. Zhongguo Zhong xi yi jie he za zhi Zhongguo Zhongxiyi jiehe zazhi = Chinese journal of integrated traditional and Western medicine / Zhongguo Zhong xi yi jie he xue hui, Zhongguo Zhong yi yan jiu yuan zhu ban. 2009;29(1):68–71. Epub 2009/04/03. PubMed . [PubMed] [Google Scholar]
- 35. Lin JH, Yang YK, Liu H, Lin QD, Zhang WY. Effect of antioxidants on amelioration of high-risk factors inducing hypertensive disorders in pregnancy. Chinese medical journal. 2010;123(18):2548–54. Epub 2010/11/03. PubMed . [PubMed] [Google Scholar]
- 36. Mao C, Zhang Y, Zhang Y, Cao L, Shao H, Wang L, et al. The effect of tanshinone IIA on the cardiovascular system in ovine fetus in utero. Am J Chin Med. 2009;37(6):1031–44. Epub 2009/11/26. 10.1142/s0192415x09007478 PubMed . [DOI] [PubMed] [Google Scholar]
- 37. Zhang Y, Can R, Mao C, Zhang Y, Zhu L, Shao H, et al. The effect of tanshinone IIA on renal and liver functions in ovine fetuses in utero. Drug and chemical toxicology. 2009;32(4):362–71. Epub 2009/10/02. 10.1080/01480540903176594 PubMed . [DOI] [PubMed] [Google Scholar]
- 38. Wu W-y, Yan H, Wang X-b, Gui Y-z, Gao F, Tang X-l, et al. Sodium Tanshinone IIA Silate Inhibits High Glucose-Induced Vascular Smooth Muscle Cell Proliferation and Migration through Activation of AMP-Activated Protein Kinase. PloS one. 2014;9(4):e94957 10.1371/journal.pone.0094957 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Xu S, Liu Z, Huang Y, Chen J, Chen S, Shen X, et al. Effectiveness of combination therapy of atorvastatin and non lipid-modifying tanshinone IIA from Danshen in a mouse model of atherosclerosis. International journal of cardiology. 2014;174(3):878–80. 10.1016/j.ijcard.2014.04.226. 10.1016/j.ijcard.2014.04.226 [DOI] [PubMed] [Google Scholar]
- 40. Xu Y, Feng D, Wang Y, Lin S, Xu L. Sodium Tanshinone IIA Sulfonate Protects Mice From ConA-Induced Hepatitis via Inhibiting NF-κB and IFN-γ/STAT1 Pathways. J Clin Immunol. 2008;28(5):512–9. 10.1007/s10875-008-9206-3 [DOI] [PubMed] [Google Scholar]
- 41. Zhou LH, Hu Q, Sui H, Ci SJ, Wang Y, Liu X, et al. Tanshinone II—a inhibits angiogenesis through down regulation of COX-2 in human colorectal cancer. Asian Pacific journal of cancer prevention: APJCP. 2012;13(9):4453–8. Epub 2012/11/22. PubMed . [DOI] [PubMed] [Google Scholar]
- 42. Kulandavelu S, Qu D, Adamson SL. Cardiovascular Function in Mice During Normal Pregnancy and in the Absence of Endothelial NO Synthase. Hypertension. 2006;47(6):1175–82. 10.1161/01.HYP.0000218440.71846.db [DOI] [PubMed] [Google Scholar]
- 43. Stanley JL, Andersson IJ, Poudel R, Rueda-Clausen CF, Sibley CP, Davidge ST, et al. Sildenafil Citrate Rescues Fetal Growth in the Catechol-O-Methyl Transferase Knockout Mouse Model. Hypertension. 2012;59(5):1021–8. 10.1161/hypertensionaha.111.186270 [DOI] [PubMed] [Google Scholar]
- 44. Liu J, Morton J, Miedzyblocki M, Lee TF, Bigam DL, Fok TF, et al. Sodium tanshinone IIA sulfonate increased intestinal hemodynamics without systemic circulatory changes in healthy newborn piglets. American Journal of Physiology Regulatory, Integrative and Comparative Physiology. 2009;297(4):H1217–H24. 10.1152/ajpheart.00477.2009 [DOI] [PubMed] [Google Scholar]
- 45. Wang J, Dong M-Q, Liu M-L, Xu D-Q, Luo Y, Zhang B, et al. Tanshinone IIA modulates pulmonary vascular response to agonist and hypoxia primarily via inhibiting Ca2+ influx and release in normal and hypoxic pulmonary hypertension rats. European journal of pharmacology. 2010;640(1–3):129–38. 10.1016/j.ejphar.2010.04.047 [DOI] [PubMed] [Google Scholar]
- 46. Mao C, Zhang Y, Zhang Y, Cao L, Shao H, Wang L, et al. The effect of tanshinone IIA on the cardiovascular system in ovine fetus in utero. Am J Chin Med. 2009;37(6):1031–44. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the paper.