

Abstract
Diverse classes of RNA, ranging from small to long non-coding RNAs, have emerged as key regulators of gene expression, genome stability and defence against foreign genetic elements. Small RNAs modify chromatin structure and silence transcription by guiding Argonaute-containing complexes to complementary nascent RNA scaffolds and then mediating the recruitment of histone and DNA methyltransferases. In addition, recent advances suggest that chromatin-associated long non-coding RNA scaffolds also recruit chromatin-modifying complexes independently of small RNAs. These co-transcriptional silencing mechanisms form powerful RNA surveillance systems that detect and silence inappropriate transcription events, and provide a memory of these events via self-reinforcing epigenetic loops.
In many organisms, intergenic or antisense transcription gives rise to different classes of small RNAs and long non-coding RNAs (lncRNAs) that have emerged as key regulators of chromatin structure in eukaryotic cells1,2. In addition to their roles in RNA degradation and translational repression, small RNAs modify chromatin and target gene expression via RNA interference (RNAi) pathways3–11 (BOX 1). In many instances, nuclear RNAi pathways mediate histone or DNA methylation events that repress transcription. Studies of the mustard plant Arabidopsis thaliana6–9,12 first demonstrated that post-transcriptional gene silencing, and the accompanying DNA methylation of target loci, correlated with the production of small interfering RNAs (siRNAs) and thus linked RNA-directed DNA methylation to the RNAi pathway, which had previously been described in Caenorhabditis elegans13. Studies in the fission yeast Schizosaccharomyces pombe, the ciliate protozoa Tetrahymena thermophila, as well as in animal germline and somatic cells, then revealed a general role for RNAi and related mechanisms in heterochromatin formation or DNA methylation10,14–17 (see Supplementary information S1 (figure)).
Box 1. Small-RNA silencing pathways.
RNA interference (RNAi) is used broadly to refer to various RNA silencing pathways that use small RNAs, together with a member of the conserved Argonaute (AGO) and PIWI family of proteins, to target genes for inactivation at the post-transcriptional or transcriptional levels1,140. Classical RNAi is triggered by long double-stranded RNA (dsRNA) precursors, which are processed to 22–23-nucleotide duplex small interfering RNAs (siRNAs) with 2-nucleotide 3′ overhangs by the RNase III ribonuclease Dicer141–145. siRNA duplexes contain 5′ monophosphate and 3′ OH groups, and are loaded onto AGO family proteins, which associate with the siRNA 5′ monophosphate-containing nucleotide via their middle (MID) domain, and with the siRNA 3′ nucleotide via their PIWI–AGO–ZWILLE (PAZ) domain146–148. The arrangement and specificity of the MID and PAZ domains in AGO proteins allow them to associate with small RNAs of specific sizes with distinct termini. siRNAs guide AGO proteins and their associated complexes to complementary RNAs, which are then targeted for degradation, translational repression or transcriptional silencing3–5. In addition to the MID and PAZ domains, AGO proteins contain an RNase H-like domain that can cleave or slice target RNAs, promoting their degradation149. In some organisms, this slicer activity is also critical for the release of one of the two AGO-bound siRNA strands and the conversion of duplex siRNA to mature single-stranded siRNA150–153. In Schizosaccharomyces pombe, plants, Tetrahymena thermophila and Caenorhabditis elegans, siRNA-programmed AGO recruits an RNA-dependent RNA polymerase (RdRP), which uses the targeted RNA as a template to synthesize a dsRNA substrate for Dicer, thereby amplifying siRNAs and the RNAi response27,154–156.
The genomes of Drosophila melanogaster and mammals do not seem to encode RdRPs. However, these organisms harbour another class of small RNAs, PIWI-interacting RNAs (piRNAs), which mediate RNA degradation in the cytoplasm and DNA or histone methylation in the nucleus157–160. Metazoan piRNAs originate from single-stranded RNA precursors, and their amplification in the D. melanogaster and mammalian germ lines involves the ‘ping-pong’ cycle whereby piRNA-guided cleavage of complementary RNAs by one PIWI paralogue generates the 5′ ends of new piRNAs that are loaded onto another paralogue, and vice versa, in a process that degrades transposon mRNAs84,161,162. It is proposed that piRNA pools are initially derived from random sampling of the transcriptome and then selectively enriched by ping-pong amplification for sequences corresponding to transposons that are actively transcribed and that are able to contribute substrates to the cycle85. This idea is reminiscent of primal RNAs (priRNAs) in S. pombe. priRNAs also result from cellular RNA sampling by Ago1, but they only direct small-RNA amplification at loci where antisense RNA targets are available163. This leads to repeat-specific RNAi-dependent siRNA accumulation163.
RNA also regulates chromatin modifications and structure through pathways that do not involve RNAi; some lncRNAs, and even some mRNAs, seem to contain signals that recruit chromatin-modifying complexes independently of small RNAs18. Early examples include X inactive specific transcript (XIST), which coats the entire inactive X chromosome in female mammals, and RNA on the X 1 (roX1) and roX2, which coat the X chromosome in male flies, ultimately leading to increased transcription19–21. More recently, a large number of other lncRNAs have been shown to act at a gene-specific level, rather than at a chromosomal level, to either activate or silence transcription18,22 (see Supplementary information S1 (figure)).
A unifying mechanism by which small RNAs and lncRNAs modify chromatin structure and silence transcription is the formation of RNA scaffolds. Although the machineries that use RNA scaffolds have greatly diverged throughout evolution, a number of key similarities enable us to define the common themes and principles that are conserved in eukaryotes from fission yeast to mammals.
In this Review, we discuss recent progress in our understanding of the role of RNA in genome regulation, focusing on the roles of different classes of chromatin-bound RNAs as scaffolds for chromatin-modifying complexes. Moreover, we review recent mechanistic insights into how small-RNA amplification loops are coupled to histone or DNA methylation to form self-reinforcing positive feedback systems that maintain epigenetic states. First, we discuss how small RNAs and Argonaute (AGO) complexes are assembled and how they target specific chromatin regions for silencing, focusing on the better understood S. pombe and A. thaliana systems where distinct mechanisms have been elucidated by which siRNAs and histone or DNA methylation events form self-reinforcing epigenetic loops. Second, we review nuclear small-RNA silencing pathways in other model systems, including C. elegans, Drosophila melanogaster and mammals, highlighting conservation and divergence in the roles of these pathways in gene and genome regulation. Finally, we discuss the mechanisms by which lncRNAs and mRNAs interact with RNA processing and chromatin-modifying machineries independently of RNAi pathways.
RNAi-mediated heterochromatin assembly
RNAi-mediated transcriptional gene silencing is best understood in S. pombe, in which many basic principles of the pathway were first deciphered. S. pombe contains a single gene each for Ago (ago1+), Dicer (dcr1+) and RNA-dependent RNA polymerase (RdRP; rdp1+) (BOX 2). Deletion of any of these genes was shown to result in loss of heterochromatic gene silencing at pericentromeric DNA repeat regions and a reduction in the levels of histone H3 lysine 9 (H3K9) methylation, a conserved marker of heterochromatin in organisms ranging from yeast to plants and mammals10. Additionally, early sequencing experiments detected small RNAs that mapped to the pericentromeric repeat regions, and Rdp1 was found to associate with centromeric DNA10,11. Based on these results, it was suggested that RNAi has an important role in the initiation of heterochromatin formation and its subsequent maintenance at pericentromeric DNA repeats via the Rdp1-mediated recruitment of H3K9 methylation. However, later studies indicated that a small-RNA-programmed Ago1 complex mediates the targeting of specific chromosome regions and recruitment of H3K9 methylation23 (see below).
Box 2. Heterochromatic small RNAs.
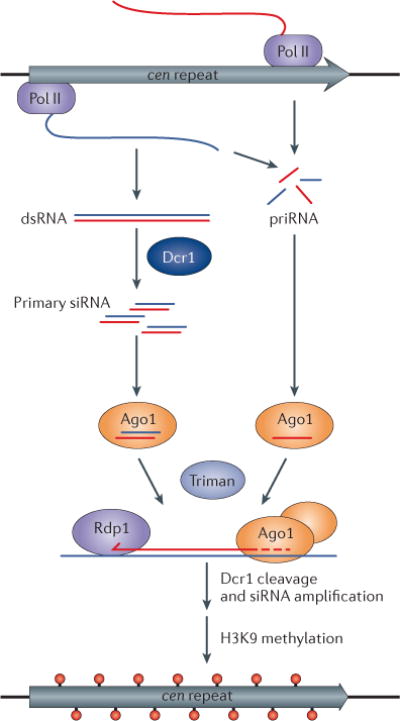
The discovery of small RNAs derived from the pericentromeric repeats of the fission yeast Schizosaccharomyces pombe marked the first example of heterochromatic small RNAs in any organism11, and studies of S. pombe have remained useful for uncovering the modes of biogenesis of this small-RNA class. Non-coding transcripts generated from pericentromeric DNA repeats (cen) act as the precursors for heterochromatic small interfering RNAs (siRNAs) in S. pombe. Initially, it was hypothesized that heterochromatic small RNAs arise from Dicer 1 (Dcr1) cleavage of double-stranded RNA (dsRNA) formed by the base-pairing of complementary pericentromeric transcripts. According to this hypothesis, the RNA-dependent RNA polymerase Rdp1 amplifies the siRNA pool by generating additional Dcr1 substrates10. In this scenario, heterochromatic small RNA levels are expected to be higher in cells lacking Rdp1, in which pericentromeric transcripts from opposite strands can still hybridize and undergo Dcr1 processing, than in cells lacking Dcr1. Initially, no differences in small RNA levels in rdp1− and dcr1− cells were detected on northern blots or in early small-RNA sequencing experiments27,163. However, recent advances in increasing the depth of sequencing have enabled the detection of a small population of Dcr1-dependent, Rdp1-independent heterochromatic small RNAs known as primary siRNAs, which are thought to arise from the base-pairing and Dcr1 processing of bidirectionally synthesized transcripts164.
Dcr1 can limit the biogenesis of primary siRNAs, as overexpression of Dcr1 not only increases primary siRNA levels but also induces siRNA production from convergently transcribed loci genome-wide164. Thus, limited Dcr1 availability is likely to represent an adaptation that restricts heterochromatic small-RNA biogenesis to a subset of loci with high levels of bidirectional transcription. The low abundance of Dcr1 and primary siRNAs is compensated for by the Rdp1-mediated generation of secondary heterochromatic siRNAs and also by the cooperation of the two enzymes — Dcr1 associates with Rdp1 and stimulates it dsRNA synthesis activity50. Consistent with an adaptive relationship between low Dcr1 abundance and Rdp1 activity, overexpression of Dcr1 suppresses, to a large extent, the requirement for Rdp1 and for Dsh1, a factor required for Dcr1–Rdp1 association, in pericentromeric silencing and histone H3 lysine 9 (H3K9) methylation164,165. These observations also rule out a critical role for Rdp1 in the direct recruitment of H3K9 methylation activity to chromatin as previously proposed10.
Finally, a class of Dcr1-independent heterochromatic small RNAs known as primal RNAs (priRNAs), which are capable of directing H3K9 methylation in an Ago1-dependent manner, has been discovered (see the figure)163. This finding, together with the detection of primary siRNAs164, suggests that self-reinforcing feedback loops involving small RNAs and chromatin modification (BOX 3) may be nucleated by their small RNA components. In support of this idea, priRNAs and siRNAs, which are both trimmed to their mature length by the 3′ exonuclease Triman, are required for the maintenance of RNAi-dependent facultative heterochromatin islands and de novo establishment of constitutive pericentromeric heterochromatin (see the figure)166.
Pol II, RNA polymerase II.
The RITS complex and the emergence of the ‘nascent transcript’ model
A physical connection between the RNAi pathway and heterochromatin was established by the purification of Chp1, a chromodomain protein that is required for silencing the same heterochromatic regions targeted by RNAi23,24. Chp1 was found to be a component of an RNA-induced transcriptional silencing (RITS) complex that also contains Ago1 and heterochromatic small RNAs (BOX 2). The third RITS complex subunit, Tas3, is also required for silencing and contains a conserved Ago-binding GW domain23,25,26. The RITS complex associates with the RNA-dependent RNA polymerase complex (RDRC), which includes Rdp1, the helicase Hrr1 and the non-canonical poly(A) polymerase Cid12 (REF. 27). Both complexes associate not only with heterochromatin and one another, but also with non-coding pericentromeric RNAs27. The functional importance of this RNA association is made clear by the observation that ectopic tethering of the RITS complex to a euchromatic mRNA triggers H3K9 methylation at its site of transcription28. Together, these results led to the development of the ‘nascent transcript’ model for RNAi-dependent heterochromatin assembly27,28 (FIG. 1). Chromatin-associated RNAs are thought to act as scaffolds for the cooperative assembly of complex machineries that couple small-RNA-mediated recognition to chromatin modifications. This co-transcriptional gene silencing mechanism contains two primary components28–31. First, the nascent transcript is degraded by both RNAi-dependent and RNAi-independent mechanisms, with RNAi-independent mechanisms involving TRAMP and exosome complexes29. Second, RNAi-dependent H3K9 methylation leads to heterochromatin formation and transcriptional gene silencing30,31. This small-RNA-based targeting strategy for heterochromatin assembly is in contrast with that used by the distant fungal relative Saccharomyces cerevisiae, in which specificity is instead determined by site-specific DNA-binding proteins (reviewed in REF. 32).
Figure 1. The ‘nascent transcript’ model and a self-reinforcing epigenetic loop in S. pombe.
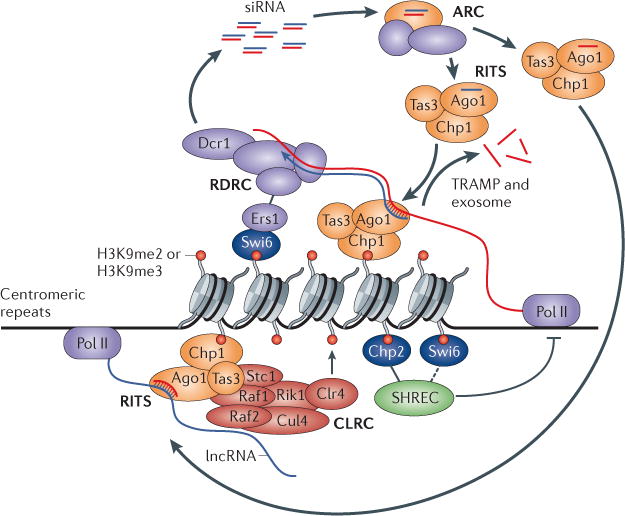
In Schizosaccharomyces pombe, the RNA-induced transcriptional silencing (RITS) complex establishes a physical connection between small interfering RNAs (siRNAs) and heterochromatin by targeting a nascent transcript, and forms the basis of a self-sustaining feedback mechanism that couples siRNA production to chromatin modification. A siRNA-targeted centromeric long non-coding (lncRNA) bound to the RITS complex becomes a template for double-stranded RNA (dsRNA) synthesis by the RNA-dependent RNA polymerase complex (RDRC, which is composed of Rdp1, Hrr1 and Cid12) and generation by Dicer 1 (Dcr1) of new siRNAs, leading to further targeting of the RITS complex after passage of Argonaute (Ago) through the ARC (Ago siRNA chaperone) complex. The Chp1 subunit of the RITS complex anchors the complex onto nucleosomes with histone H3 lysine 9 (H3K9) methylation, and the RITS complex recruits the Clr4–Rik1–Cul4 (CLRC, of which Clr4 is the methyltransferase) complex via Rik1 and Stc1 to promote the further spread of H3K9 methylation. The heterochromatin protein 1 (HP1) homologue Swi6 binds to methylated H3K9 and promotes RDRC recruitment and siRNA biogenesis via the silencing factor Ers1. Swi6, and particularly the other HP1 protein Chp2, help to restrict RNA polymerase II (Pol II) access by recruiting the Snf2–histone deacetylase repressor complex (SHREC). The TRAMP non-canonical poly(A) polymerase and the exosome also contribute to silencing. Together, the RITS complex and the nascent lncRNA transcript provide a hub for the assembly of machineries that make siRNAs, modify histones and silence gene expression. me2, dimethylation.
In addition to the RDRC and the RITS complex, the nascent transcript heterochromatin assembly platform also includes the Clr4–Rik1–Cul4 (CLRC) complex, the Clr4 subunit of which is the sole H3K9 methyltransferase in S. pombe33–37. The CLRC complex subunit Rik1 associates with both the RDRC and the RITS complex, and the peripheral CLRC member Stc1 also provides a link to the RNAi machinery via its interaction with Ago1, although it seems to be dispensable for the interaction of the CLRC complex with the RITS complex subunit Tas3 (REFS 38–40). These physical connections suggest the existence of a feedback loop in which the activities of the RITS complex, the RDRC and the CLRC complex reinforce each other (FIG. 1) (see below).
The central role of nascent RNAs in the recruitment of RNAi and histone-modifying activities has prompted investigations of the interactions between RNA-mediated silencing and other nuclear processes in S. pombe. The CLRC complex components Raf2 (also known as Cmc2) and Rik1 were recently found to associate with Cdc20 (the catalytic subunit of the leading-strand DNA polymerase-ɛ) and Mms19 (a conserved regulator of the general transcription factor TFIIH41), suggesting the coordination of DNA replication with RNAi-dependent release of RNA polymerase II (Pol II) in the inheritance of heterochromatin42. However, this Raf2–Rik1 complex seems to be distinct from the CLRC complex, and direct evidence of cooperation between the RNAi and DNA replication machineries is lacking. It has also been proposed that the splicing machinery contributes to heterochromatic silencing by directly interacting with the RDRC and the nascent transcript platform to promote siRNA biogenesis43,44. However, earlier biochemical purifications demonstrated that the RDRC subunit Cid12 forms RDRC-independent complexes with splicing factors27. Moreover, the RNAi defect found in several splicing mutants can be partially rescued by the introduction of cDNAs that encode RNAi components45, suggesting that the effects of splicing mutations on heterochromatic silencing are at least partially due to improper splicing of pre-mRNAs that encode RNAi proteins.
siRNAs form self-reinforcing epigenetic loops with DNA and histone methylation
The observation that silent states of gene expression and their associated chromatin modifications can spread along DNA in cis and persist throughout cell division32 led to the idea that positive feedback loops might form the molecular basis for epigenetic memory. Linking the recognition of signals — such as covalent histone modifications, DNA methylation or small RNAs — to the generation of new signals would ensure the stable propagation of silent states46–48 (BOX 3). Indeed, this paradigm of self-reinforcing loops has found consistent support from experimental findings in the small-RNA-dependent epigenetic pathways of both fungi and plants.
Box 3. Self-reinforcing positive feedback loops.
Self-reinforcing positive feedback loops are formed by the functional coupling of different types of signal generation events. Studies in Schizosaccharomyces pombe, Arabidopsis thaliana and mammals have uncovered the molecular basis of these coupling events by identifying the proteins that recognize either histone H3 lysine 9 dimethylation (H3K9me2) and H3K9me3, or DNA cytosine methylation (DNA-5mC), and that recruit enzymes which catalyse the other methylation events. In some organisms (for example, S. pombe and A. thaliana), the methylation events are also physically coupled to proteins that recruit small interfering RNA (siRNA) amplification loops to chromatin, thus forming self-reinforcing loops in which the histone or DNA methylation event promotes siRNA generation and siRNAs in turn promote histone or DNA methylation. Self-reinforcing loops are thought to help to ensure the maintenance of epigenetic information47,48.
In S. pombe, the discovery that pericentromeric H3K9 methylation is dependent on the RNAi machinery was quickly followed by the realization that, conversely, pericentromeric siRNA accumulation also requires the H3K9 methyltransferase Clr4 (REFS 27,49). Several physical interactions are now known to underlie this mutual dependence. First, the bivalent nature of the RITS complex — its ability to interact with nascent RNA via its siRNA-bound Ago1 subunit and with chromatin via its Chp1 subunit — forms the physical basis for a self-reinforcing positive feedback loop (FIG. 1). Methylated H3K9 stabilizes RITS complex binding via Chp1 (REF. 49), promoting siRNA amplification by the RITS complex-interacting factors RDRC and Dcr1 (REFS 27,50). Second, methylated H3K9 enhances siRNA accumulation by helping to recruit the RDRC via the heterochromatin protein 1 (HP1) homologue Swi6 in an interaction that is directly bridged by Ers1 (REFS 30,51,52). Finally, direct associations between the siRNA-programmed RITS complex and the Rik1 and Stc1 subunits of the CLRC38–40 ensure that siRNAs in turn feed back on H3K9 methylation states (FIG. 1).
The notion of self-reinforcing epigenetic states that involve multiple types of signals was first proposed by Eric Selker while working with the filamentous fungus Neurospora crassa46. Initial results revealed that DNA methylation patterns were dependent on histone deacetylation, but recent findings suggest that feedback loops that involve small RNAs also exist in N. crassa. The canonical DNA methylation pathway is RNAi-independent53 and acts downstream of, and without feeding back on, H3K9 methylation states54. By contrast, a newly uncovered DNA methylation phenomenon exhibits self-reinforcing properties and correlates with the biogenesis of small RNAs. This type of DNA methylation occurs at gene-rich and intergenic loci that are transcribed in a convergent manner and that also give rise to Dicer-independent siRNAs (disiRNAs)55,56. Remarkably, disiRNA loci DNA methylation (DLDM), unlike the well-studied DNA methylation pathway associated with repetitive sequences, is required for H3K9 trimethylation (H3K9me3) and is thus self-reinforcing through the known HP1-dependent mechanism that recruits the DNA methyltransferase DIM-2 to the regions of H3K9 methylation54,55. It was also observed that, for a given DLDM locus, most clones are not methylated; however, when methylation does occur clones are methylated extensively across the disiRNA-producing locus, suggesting that a powerful positive feedback loop leads to the spread of the epigenetic mark55. Convergent transcription is critical and, when induced artificially, is sufficient to drive this dynamic DNA methylation pattern55. The potential roles of the disiRNAs themselves, and other mechanistic aspects of DLDM, remain to be elucidated, but these results further underscore the prevalence of self-reinforcing epigenetic loops among small-RNA silencing pathways in the nucleus.
RNA-directed DNA methylation in A. thaliana
In A. thaliana, 24-nucleotide siRNAs direct de novo DNA methylation and maintenance of DNA methylation at asymmetrical CHH sites (where H represents any base except G) in a manner that depends on two plant-specific polymerase II (Pol II)-related RNA polymerases, Pol IV and Pol V57 (reviewed in REF. 58). This phenomenon exhibits many physical connections between pathway components that form the basis of a self-reinforcing epigenetic loop (FIG. 2). First, 24-nucleotide siRNAs are generated by DCL3 processing of double-stranded RNA (dsRNA) synthesized by the RdRP RDR2 using Pol IV transcripts as templates. Recent in vitro experiments have shown that RDR2 and Pol IV associate in an RNA-independent manner and that the activity of RDR2, but not of Pol IV, requires this association, which suggests an evolved mechanism for limiting siRNA biogenesis to loci occupied by Pol IV59. Loading of siRNAs onto AGO4 in the cytoplasm triggers its import into the nucleus60, where it associates directly with siRNA-complementary Pol V transcripts61,62. This sequence-specific recruitment is reinforced by direct interactions between AGO4 and the GW domains of both Pol V and the Pol V transcript-binding protein KTF1 (also known as SPT5L)25,63. While these multiple associations stabilize the localization of AGO4 along a Pol V nascent transcript, the RDM1 protein provides a physical link between AGO4 and the DRM2 DNA methyltransferase, illustrating how the instruction to silence gene expression is conveyed from small RNAs to covalent DNA modifications64. RDM1 also has an affinity for methylated DNA, which raises the possibility that it can direct the Pol V–KTF1–DRM2–AGO4 complex to pre-existing sites of DNA methylation. A recent study shows that AGO4 co-purifies with DRM2 in vivo, and that DRM2 preferentially methylates the strand of DNA that acts as a template for Pol V transcripts65. The authors propose that as AGO4 binds to a siRNA-complementary nascent Pol V transcript, it directs DRM2 specifically to the template DNA strand as it emerges from the Pol V exit channel. This study provides an unprecedented level of mechanistic detail on the feedback between small RNAs and DNA methylation.
Figure 2. A self-reinforcing loop linking siRNAs to DNA and histone methylation in A. thaliana.
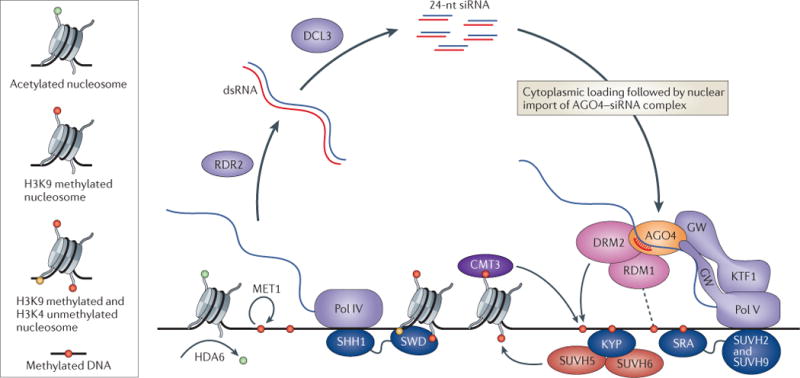
Elaborate feedback between small RNAs and DNA and histone methylation underlies a robust silencing pathway at sites of asymmetrical cytosine methylation in the Arabidopsis thaliana genome. Two plant-specific polymerases transcribe the critical RNAs. RNA polymerase IV (Pol IV) transcripts are processed by the RNA-dependent RNA polymerase RDR2 and the Dicer protein DCL3 into 24-nucleotide (nt) small interfering (siRNAs), while Pol V transcripts act as their targets. The Argonaute protein AGO4, the siRNA-dependent recruitment of which to Pol V transcripts is reinforced by interactions with the GW domains of Pol V and an associated elongation factor KTF1, in turn recruits the CHH DNA methyltransferase DRM2. RDM1 associates with the Pol V–AGO4–DRM2 complex and may link siRNA amplification to pre-existing DNA methylation. Meanwhile, another DNA methyltransferase that targets CHG sites for maintenance, CMT3, is recruited directly to methylated histone H3 lysine 9 (H3K9). Silencing by DNA methylation is augmented by H3K9 methylation, which is deposited by the enzymes KYP, SUVH5 and SUVH6. These methylation events are coupled to one another and to siRNA activity in several ways. KYP is recruited directly to methylated DNA, where it methylates neighbouring histones, and the H3K9 methylation reader SHH1 recruits Pol IV to promote siRNA generation, while the DNA methylation readers SUVH2 and SUVH9 recruit Pol V to promote AGO4 targeting and further DNA methylation. Thus, the different methylation readers, RNA polymerases and AGO4 act together to create self-reinforcing interactions between pre-existing DNA methylation and siRNA amplification. Erasure of DNA methylation by mutations in either the histone deacetylase HDA6 or the maintenance DNA methyltransferase MET1 results in loss of siRNA biogenesis, emphasizing the importance of these self-enforcing interactions. Altogether, the A. thaliana pathway for DNA methylation at asymmetrical sites is one of the most remarkable examples of a recurring theme in epigenetic regulation by small RNAs: self-reinforcing feedback loops. SRA, SET and RING finger-associated; SWD, SAWADEE domain.
Interestingly, at a subset of loci, deletion of DNA methylation by mutations in the histone deacetylase HDA6 or the maintenance cytosine methyltransferase MET1 results in loss of silent locus identity66. This observation indicates that, at these loci, silent locus identity is maintained by self-reinforcing interactions involving histone deacetylation and DNA methylation, while siRNA amplification and silencing rely on cooperation between the Pol V–KTF1–DRM2–AGO4 complex and Pol IV machineries (see above) and on pre-existing chromatin modifications66.
Embedded in the RNA-dependent DNA methylation pathway are important self-reinforcing feedback mechanisms that have been discovered only lately. For instance, regions of RNA-directed DNA methylation in A. thaliana are also enriched in H3K9 methylation58, and these two epigenetic signals mutually support one another. Three distinct H3K9 methyltransferases, KYP (also known as SUVH4), SUVH5 and SUVH6, contribute to the maintenance of DNA methylation patterns, with KYP having the primary role58. KYP is recruited to sites of DNA methylation via its SET and RING finger-associated (SRA) domain67 and, conversely, the Pol IV-associated factor SHH1 binds to nucleosomes with methylated H3K9 and unmethylated H3K4 via its SAWADEE domain, thus promoting siRNA-dependent methylation of the underlying DNA68,69. Another example of self-reinforcement involves the SUVH2 and SUVH9 proteins which, like KYP, bind to methylated DNA via their SRA domains but, unlike KYP, lack H3K9 methyltransferase activity70. Two recent studies indicate that these SRA domain-containing proteins directly recruit Pol V to sites of pre-existing DNA methylation, thus generating nascent transcript scaffolds for recruitment of AGO4–siRNA complexes and further enhancement of the DNA methylation signal71,72. This dense network of positive feedback in A. thaliana suggests that the broad evolutionary relevance of self-reinforcing siRNA-driven epigenetic loops extends beyond the S. pombe system in which they were first suggested.
Conservation and divergence of nuclear RNAi
Below, we discuss examples of small-RNA-mediated silencing in the animal germ line, including C. elegans, D. melanogaster and mammals. Nuclear RNAi in these systems differs from the examples in S. pombe or A. thaliana in one major aspect. Although animal small RNAs act as signals that trigger the modification of chromatin or DNA, it remains unclear whether small-RNA amplification involves a self-reinforcing relationship with the downstream chromatin or DNA modifications. Recent studies of the D. melanogaster Piwi-interacting RNAs (piRNAs), so called because they partner with Ago proteins of the largely germline-specific Piwi subfamily, point in this direction, but mammalian pathways seem to lack this characteristic. Nuclear RNAi-related events associated with the introduction of foreign siRNAs into animal somatic cells are not discussed here.
Small RNAs and chromatin in animal cells: the piRNA system
piRNAs were first discovered in mice and D. melanogaster, and their roles in silencing transposons in the germ line of these organisms are well established (reviewed in REF. 73). The C. elegans genome also encodes piRNAs, called 21U-RNAs, but they have only been implicated in the repression of one transposable element74,75; we discuss the roles of 21U-RNAs in epigenetic silencing in the next subsection.
In D. melanogaster, recent evidence suggests that the nuclear protein Piwi, in addition to mediating post-transcriptional gene silencing of transposons by contributing to ‘ping-pong’ amplification with its cytoplasmic counterparts Aubergine (Aub) and Ago3, also targets transposons at the transcriptional level76–80. In ovarian somatic cells, which also harbour piRNAs, most of the euchromatic H3K9 methylation islands in the genome correspond to transposon insertion sites and are Piwi-dependent79. Furthermore, artificial recruitment of Piwi to a reporter locus induces H3K9 methylation, HP1a (also known as Su(var)205) accumulation and exclusion of RNA Pol II, suggesting that piRNAs may have a direct role in guiding chromatin changes76,77. Piwi and HP1a were previously reported to interact directly in vivo81, but the reproducibility and physiological importance of this result have been debated77,80, raising the possibility that there are unidentified links that transmit instructions from piRNAs to chromatin. Mechanisms involving Piwi engagement with nascent transcripts or even the underlying DNA have been proposed76,77, but understanding the molecular details remains a major challenge for future research.
Recent findings also show that sites bearing Piwi-dependent H3K9 methylation in turn function as source loci for piRNA production in D. melanogaster ovaries. Methylated H3K9 recruits the HP1 family protein Rhino, which, together with the Cutoff protein, promotes non-canonical transcription from these loci to generate precursors to be processed into piRNAs82,83. Therefore, not unlike the plant and fungal systems discussed above, the D. melanogaster piRNA pathway seems to exhibit a positive feedback loop in which small RNAs guide histone methylation, which in turn enables biogenesis of more small RNAs.
In mice, piRNAs silence transposons in the male germ line by targeting them for de novo DNA methylation during late embryonic and early neonatal development84–87. Consistently, the nuclear PIWI protein, Piwi-like protein 4 (PIWL4; also known as MIWI2), is expressed at this time85. Thus, like plants, mammals have an RNA-directed DNA methylation pathway, although few of its mechanistic details are currently understood. Arguing against a self-reinforcing loop is the observation that piRNA levels remain high in DNA (cytosine-5-)-methyltransferase 3-like (Dnmt3l) mutants, in which transposon DNA methylation is completely compromised85. However, it is unclear to what extent H3K9 methylation at transposons, which could also contribute to piRNA generation, is affected in Dnmt3l mutants. One study has suggested that piRNA-directed DNA methylation in mice involves a system that is similar to the nascent transcript model in S. pombe88. The authors found that piRNAs mapped to a long terminal repeat (LTR) adjacent to a paternally imprinted locus, RAS protein-specific guanine nucleotide-releasing factor 1 (Rasgrf1), and that piRNA pathway components are required for methylation of Rasgrf1. They also found evidence of sequence-specific piRNA-dependent cleavage of a non-conding RNA overlapping with the LTR, supporting the nascent transcript theory88. However, it has also been shown that the endonuclease activity of PIWL4 is completely dispensable for DNA methylation of long interspersed elements (LINE1) and the intracisternal A particle (IAP) family of LTR retrotransposons89. This suggests that nascent transcript slicing is not an important event in small-RNA-dependent chromatin silencing in mice, although it has a critical role in piRNA amplification and post-transcriptional gene silencing.
Finally, piRNAs also contribute to epigenetic phenomena in which they are themselves responsible for epigenetic inheritance. This idea first emerged in a landmark paper demonstrating the requirement for maternal deposition of piRNAs, in addition to the presence of genomic piRNA loci, for effective transposon silencing and fertility in the offspring15. A recent study found that in D. melanogaster, strains containing certain P-lacZ transgene insertions, but not other strains containing similar insertions at the same site, showed silencing of homologous sequences in trans90. Remarkably, if maternally inherited, the trans-silencing-competent alleles could convert the paternally inherited alleles into strong trans-silencers, which was accompanied by de novo production of piRNAs corresponding to these trans-silencing loci. The new allele also acquired the ability to convert other alleles into trans-silencers in the same manner; thus, this constitutes a case of paramutation90,91. These results indicate that maternally deposited cytoplasmic piRNAs can direct the conversion of homologous sequences into piRNA-generating loci with trans-silencing ability. This work shows how a locus can become incorporated into the piRNA repertoire and how RNA itself can act as a carrier of epigenetic information. Future discoveries regarding the molecular basis of this paramutation process will provide fundamental insights into both piRNA biology and the principles of epigenetic inheritance.
RNAi and H3K9 methylation in C. elegans
C. elegans has an array of 27 AGO homologues that associate with diverse small RNA populations and that act in distinct silencing steps92. Classical RNAi, induced by exogenous dsRNA, initially involves the AGO protein RDE-1 and primary siRNAs; however, RdRPs generate secondary siRNAs that are not loaded onto RDE-1 (REF. 92). Among other roles, secondary siRNAs can trigger RNAi in the nuclei of somatic cells by acting with the AGO protein NRDE-3. Upon small-RNA loading, NRDE-3 enters the nucleus, associates with complementary nascent transcripts and recruits the non-AGO silencing factor NRDE-2, which in turn promotes H3K9 methylation and inhibits elongation of RNA polymerases beyond the site targeted by siRNAs17,93,94 (FIG. 3). Thus, as in fungi and plants, small RNAs in C. elegans can trigger transcriptional gene silencing. However, currently, the biochemical connections remain obscure except for a direct interaction between NRDE-3 and NRDE-2 (REF. 16). More recent work has shown that additional AGO homologues are also required for chromatin targeting17. Interestingly, RNAi-induced H3K9 methylation is a transgenerational phenomenon, as it is inherited for at least two generations, in the absence of the dsRNA trigger, by a mechanism that involves germline transmission of siRNAs17,95.
Figure 3. Small-RNA-driven transcriptional silencing of gene expression in C. elegans.
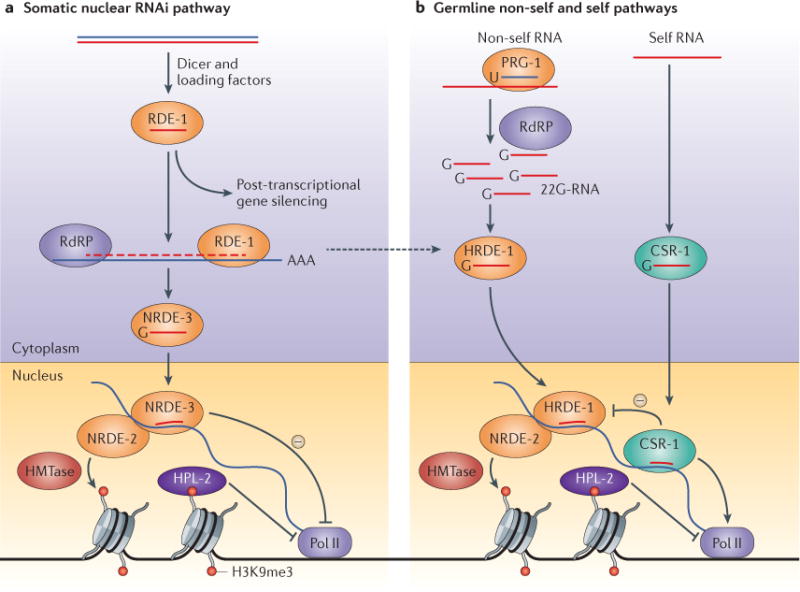
a | In Caenorhabditis elegans, exogenous double-stranded RNA (dsRNA) is processed into primary small interfering RNAs (siRNAs) that are loaded onto the Argonaute (AGO) protein RDE-1 and amplified by RNA-dependent RNA polymerases (RdRPs) to give rise to secondary siRNAs called 22G-RNAs. When loaded with 22G-RNAs, the somatic AGO protein NRDE-3 translocates to the nucleus, where it targets nascent RNA transcripts and silences corresponding genes, acting in concert with the silencing factor NRDE-2. Gene expression is halted by NRDE-2 during the elongation phase of transcription, and silencing involves histone H3 lysine 9 trimethylation (H3K9me3) and recruitment of the heterochromatin protein 1 (HP1)-like protein HPL-2. b | In the germ line, small-RNA-directed transcriptional silencing is mediated not by NRDE-3 but by a different AGO protein, HRDE-1, which also acts through NRDE-2, H3K9 methylation and HPL-2. HRDE-1 receives 22G-RNA inputs both from the pathway that responds to exogenous dsRNA and from the PIWI-interacting RNA (piRNA), or 21U-RNA, pathway that scans the transcriptome for foreign RNAs. 21U-RNA-programmed PRG-1 promotes the RdRP-dependent generation of 22G-RNAs, which are loaded onto HRDE-1. In both cases, HRDE-1 maintains a persistent, transgenerational memory of silenced genes in the germ line. Meanwhile, another AGO protein called CSR-1 binds to 22G-RNAs that represent the full complement of endogenously expressed RNAs, and protects the corresponding loci from possible silencing by HRDE-1. Thus, the 22G-RNAs bound by CSR-1 and HRDE-1 transmit a germline memory of ‘self’ and ‘non-self’ RNAs, respectively, to be appropriately licensed for expression or silenced. HMTase, histone methyltransferase; Pol II, RNA polymerase II.
In the C. elegans germ line, inheritance of H3K9 methylation induced by exogenous dsRNA is mediated by the germline-specific AGO protein HRDE-1 (also known as WAGO-9) in association with RdRP-derived siRNAs96,97. Intriguingly, both HRDE-1 and NRDE-2 are also necessary for germline immortality and fertility97. This biological requirement is reminiscent of the piRNA pathway, the widely conserved germline maintenance system discussed above and represented in C. elegans by the 21U-RNAs and the PIWI-subfamily AGO protein PRG-1 (REF. 74). Indeed, several studies have concluded concurrently that HRDE-1 is in fact the downstream effector of piRNA-induced silencing in the germ line96,98–100. Another observation was also made — while HRDE-1, NRDE-2 and the HP1 homologue HPL-2 are required to maintain a permanent memory of these silencing events at the chromatin level, PRG-1 and 21U-RNAs are only required for its initiation96,99,100 (FIG. 3). Thus, different components are responsible for the establishment and maintenance steps in this example of RNA-mediated epigenetic regulation.
In contrast to piRNAs in D. melanogaster and mammals, C. elegans piRNAs only silence one transposon family74,75. However, transgene insertions can be silenced in a PRG-1-dependent manner96,99,100, supporting a conserved function for piRNAs in recognizing ‘non-self’ nucleic acids. Interestingly, PRG-1 was found to initiate epigenetic silencing of the same single-copy transgene in some lines but not others100. Genetic crosses indicate that transgene expression states can be converted in both directions by trans-acting silencing and activating factors, leading to the proposal that HRDE-1–siRNA complexes enforce the maintenance of PRG-1-directed silencing, whereas another system prevents or even reverses its establishment100. The suggested candidate is the AGO protein CSR-1, the RdRP-dependent small RNA partners of which map not to silenced loci but instead to all mRNAs expressed in the germ line101. This was confirmed by recent experiments in which CSR-1-dependent activity was shown to counteract PRG-1-mediated silencing in a heritable manner102,103. Thus, PRG-1, associated with 21U-RNAs, is proposed to sample cellular transcripts for non-self sequences to be silenced by HRDE-1, while CSR-1 carries a repertoire of self-expressed sequences to be protected from silencing96,99,100,102,103 (FIG. 3). CSR-1 seems to have a critical role in sperm, where it promotes spermiogenic gene expression and where its associated small RNAs reflect previous germline expression patterns104.
RNA-dependent histone methylation without RNAi
In the RNAi-mediated chromatin silencing pathways discussed above, a recurring feature is the role of RNA as an assembly scaffold. Below, we focus on analogous but RNAi-independent strategies used by both mRNAs and lncRNAs to recruit histone-modifying enzymes. LncRNAs, typically defined as transcripts more than 200 nucleotides in length that do not encode proteins, have been uncovered in large numbers as a result of rapid advances in high-throughput technologies. The function of lncRNAs in genome regulation, although still largely unknown, seems to be diverse and is the subject of a growing field of study (reviewed in REF. 105). A common theme in current models is the ability of many lncRNAs to target chromatin-modifying activities to particular genomic sites. Perhaps the most extensively studied examples are those that involve dosage compensation in metazoans (reviewed in REF. 106), including the mammalian lncRNA XIST, which mediates global inactivation of a randomly chosen X chromosome in females in a process known as X chromosome inactivation (XCI). XIST has been proposed to function by directing the Polycomb repressive complex 2 (PRC2) H3K27 methyltransferase to chromatin. Below, we examine the evidence and unresolved questions regarding the recruitment of Polycomb to chromatin by XIST and other lncRNAs. The basis of specificity for the proposed protein–RNA interactions remains unknown. We also discuss a class of lncRNAs transcribed from enhancers that are thought to activate gene expression in cis. Finally, we describe a case in which the mode of specificity is better understood — the targeting of the Clr4 H3K9 methyltransferase by the mRNAs of meiotic genes to their gene loci in S. pombe.
lncRNAs and Polycomb recruitment
Polycomb proteins and H3K27me3 were first linked to XCI by genetic and cytological experiments107–109. A more direct biochemical connection was shown by the detection of XIST RNA in PRC2 immunoprecipitation experiments and a corresponding in vitro test of their interaction110. REPA — an independent internal transcript within XIST, so called because it spans the conserved A repeat region of XIST — was proposed to interact directly with and recruit the PRC2 catalytic subunit EZH2 in the earliest stages of XCI establishment. The same region of full-length XIST would then directly recruit PRC2 as silencing spreads in cis110. It was also reported that other lncRNAs—such as KCNQ1 opposite strand/antisense transcript 1 (Kcnq1ot1) and HOX transcript antisense RNA (HOTAIR)—could directly recruit PRC2 to chromatin in cis or in trans, respectively111,112. However, one study failed to detect an interaction between PRC2 and the XIST A repeats using an ultraviolet crosslinking approach113, and others have found the XIST A repeat to be dispensable for PRC2 chromatin association and H3K27 methylation in mice and humans107,114. More generally, the idea that lncRNAs mediate PRC2 targeting through direct interactions has been undermined both by doubts regarding specificity and by the existence of equally compelling alternative scenarios (reviewed in REF. 115). The number of RNAs found to be associated with PRC2 in cells is enormous116–119, highlighting the risk of interpreting individual interactions as specific. For instance, a recent study revealed that the binding affinity of PRC2 for the bacterial MBP mRNA is equal to its binding affinity for HOTAIR, and that RNA length is a far better predictor of PRC2 binding affinity than RNA sequence116. Thus, the direct interactions reported between PRC2 and lncRNAs, such as HOTAIR, RepA and Kcnq1ot1 (REFS 110–112) among others, offer only minimal insight into the regulation of methyltransferase recruitment to particular genomic loci.
Although the currently prevailing models concerning Polycomb recruitment by lncRNAs remain inconclusive, the reality is likely to involve a much subtler and more complex system of sequentially and/or cooperatively assembled components, in which specific lncRNAs act as scaffolds. For example, the Jumonji family protein JARID2 was recently reported to act as an essential intermediate between Xist and PRC2 (REF. 114). While the A repeat of Xist is required for gene silencing, it does not seem to be required for the recruitment of PRC2, as previously proposed. Instead, Xist recruits JARID2 via its B and F repeats, and JARID2 is required for the localization of PRC2 and methylation of H3K27 (REF. 114). Another study has suggested that the facilitation of JARID2–PRC2 interactions may represent a more general function of lncRNAs120, although the amino acid residues that were thought to mediate JARID2 RNA binding differ between the two studies114,120. Thus, the events linking lncRNAs to the recruitment of PRC2, and other methyltransferases to chromatin, are likely to be multiple and intricate, involving the contributions of other protein factors and perhaps other histone modifications115. The increasing scepticism towards widespread assumptions is promising and should ultimately lead to a better understanding of how lncRNAs influence chromatin.
lncRNAs and enhancer function
Transcription at a large number of mammalian enhancers gives rise to lncRNAs, called enhancer RNAs (eRNAs), that seem to have major roles in the regulation of transcription. At least a subset of eRNAs seem to act as nascent transcripts that function in cis to promote the transcription of neighbouring genes. Initially, high-throughput RNA sequencing experiments identified thousands of eRNAs that are transcribed from enhancer elements in response to signals that mediate enhancer-dependent transcriptional activation121,122. RNAi-mediated knockdown of several eRNAs was then shown to result in reduced expression of proximally located target mRNAs, suggesting that eRNAs act in cis and are required for the activation of target gene transcription123,124.
Insights into the mechanism of action of eRNAs came from the identification of components of the transcription machinery that mediate eRNA function. eRNAs involved in the activation of the developmentally regulated genes T-cell acute lymphocytic leukaemia 1 (TAL1), snail family zinc-finger 1 (SNAI1) and SNAI22 require the Mediator transcription co-activator for their activity125, while HOXA distal transcript anti-sense RNA (HOTTIP), an eRNA involved in the activation of HOXA homeobox genes, functions through the WDR5–MLL (also known as KMT2A) H3K4 methyltransferase complex124. Chromosome conformation capture experiments show that the eRNA mediates looping interactions between the enhancer and promoter regions of genes. These studies have given rise to an attractive model in which eRNAs recruit co-activator complexes and promote their interaction with gene promoters to activate transcription. Consistent with this model, there is evidence that supports a role for specific enhancer sequences that encode eRNAs, which are different from enhancer sequences that form binding sites for transcription factors, in activation of target genes126. These studies provide a new function for nascent RNA transcripts as scaffolds for the recruitment of co-activator complexes that mediate chromosome looping and transcriptional activation. Large RNAs (>200 nucleotides) may be particularly suited for such architectural tasks that bring enhancer and promoter regions, usually located great distances apart, into proximity.
The identification of both the regions within eRNAs that mediate co-activator complex recruitment and the RNA-binding domains in the subunits of these complexes is required for a better understanding of how different eRNAs activate transcription. In addition to cis-acting eRNA, several trans-acting lncRNAs that activate transcription have also been identified. They are not discussed here but have recently been extensively reviewed elsewhere22,127.
Silencing of meiotic genes in S. pombe
The regulation of certain meiotic genes in S. pombe is another well-studied example in which an RNA scaffold promotes the RNAi-independent recruitment of a histone methyltransferase to chromatin. Meiosis-specific genes are silenced in vegetative cells by a post-transcriptional process that targets an RNA cis-element called determinant of selective removal (DSR)128. Remarkably, DSRs are found within the protein-coding regions of meiotic mRNAs128. Therefore, they must exhibit some degree of sequence flexibility to preserve meiotic protein structure but effectively consist of different variants of a hexanucleotide motif129. At a subset of DSR-containing genes, notably mei4+ and ssm4+, H3K9 methylation is also observed and depends on the DSR and transcription, which suggests that the mRNAs of these genes direct Clr4 to chromatin and may promote transcriptional and post-transcriptional silencing130,131.
How do DSR-containing mRNAs recruit the Clr4 methyltransferase? Much like lncRNAs and Polycomb proteins, the process seems to involve a host of factors rather than direct interactions between the RNA and the histone-modifying enzyme. In the case of meiotic mRNAs, the source of specific recognition is much clearer. DSRs are bound directly by the YTH-domain protein Mmi1, and this binding event is required for meiotic gene silencing and for H3K9 methylation of the mei4+ and ssm4+ loci128,129,131. The Mmi1-interacting zinc-finger protein Red1 is also required for silencing and H3K9 methylation at mei4+ and ssm4+ (REFS 131,132). Strikingly, Red1 associates with Clr4, which suggests that it serves as the critical link that targets histone methylation to genes producing DSR-bearing transcripts130,131. Interestingly, this Mmi1–Red1–Clr4 axis of RNA-directed H3K9 methylation also seems to operate outside the meiotic process to target context-specific genes such as pho1+, which is repressed when phosphate is available. Data from two recent studies show that an upstream lncRNA mediates Red1 chromatin binding and H3K9 methylation at the pho1+ locus in a phosphate- and Mmi1-dependent manner133,134. Importantly, the lncRNA was found to contain DSR motifs that are critical for this regulation134.
While the DSR-bearing mRNAs demonstrate RNA-mediated recruitment of histone modifications without RNAi, silencing itself seems to be independent of H3K9 methylation. This is perhaps made most evident by the observation that some DSR-containing genes are silenced by Mmi1 and Red1 without exhibiting H3K9 methylation131. DSRs inserted ectopically in other genes produce similar effects130. Most conclusively, cells lacking Clr4 show wild-type silencing of all tested meiotic genes during vegetative growth135. Instead, the critical event for silencing of these genes seems to involve degradation by the nuclear exosome through a mechanism that requires polyadenylation of the target RNAs136. Consistently, Red1 interacts with the exosome subunit Rrp6 (REF. 132), and proteomic analysis recently showed that this association is mediated by the Mtr4-like helicase Mtl1, another recently identified Red1 interactor133,135. This poses the question of what fitness advantage is conferred by H3K9 methylation at meiotic genes, if it is not necessary for silencing. The presence of H3K9 methylation might reflect an ancestral pathway that predates the contribution of the exosome and that has not yet been lost135. It is also possible that H3K9 methylation could control expression of DSR-containing genes under certain circumstances, for example, during gene induction134,135. Additional work is needed in this area not only to define the biological function of histone methylation but also to further isolate the steps that lead to Clr4 recruitment, for example, to determine whether Mmi1 and Red1 accomplish this task independently.
Studies of the meiotic mRNAs in S. pombe have provided a valuable paradigm for investigating the RNA-mediated targeting of histone methyltransferases in all its mechanistic complexity. In this regard, it is worth noting that the XIST RNA is thought to have evolved from a protein-coding gene137. This XIST precursor gene may have shared certain features of the S. pombe meiotic RNAs that enabled it to locally recruit repressive chromatin-modifying complexes before it acquired the ability to spread along the entire X chromosome.
Conclusions and perspectives
The mechanisms by which coding RNAs and non-coding RNAs regulate chromatin structure in different organisms share key similarities that allow us to note common principles which unify these ancient pathways (FIG. 4). The first is the principle of recruitment via nascent RNA. In these systems, recruitment of effector complexes that methylate histones or DNA involves the association of small-RNA-guided AGO complexes or site-specific RNA-binding proteins with nascent coding RNA or non-coding RNA scaffolds (FIG. 4b,c), rather than specific sites on DNA and DNA-binding proteins (FIG. 4a). The second major principle involves the role of RNA as a component of self-reinforcing positive feedback loops. These loops are unique to small-RNA systems that contain an amplification component and have key roles in the epigenetic inheritance of histone and DNA methylation patterns. The key event is the localization of the small-RNA amplification machineries on nascent transcripts, and their activation by the histone or DNA methylation events induced by the small RNAs themselves (BOX 2). Thus, in both S. pombe and A. thaliana, small-RNA amplification and histone or DNA methylation are co-dependent. Self-reinforcing loops have not yet been described for lncRNAs that act independently of RNAi. However, cooperative recruitment involving the association of chromatin-modifying complexes with both pre-existing chromatin modifications and DNA-binding specificity factors has been suggested to contribute to the maintenance of epigenetic states in budding yeast32. By analogy, it is possible that direct or indirect interactions between RNA-binding proteins that recognize lncRNA and also pre-existing histone modifications help to reinforce lncRNA-mediated changes in chromatin structure. Finally, small RNAs, and their association with positive feedback loops in the germ line, allow them to act as components of transgenerational inheritance mechanisms. The transmission of small RNAs through meiosis seems to act as a signal for the inheritance of internal or environmentally induced changes from parent to offspring138,139.
Figure 4. RNAs, both short and long, represent an alternative to DNA-binding proteins as specificity determinants for epigenetic regulation of gene expression.
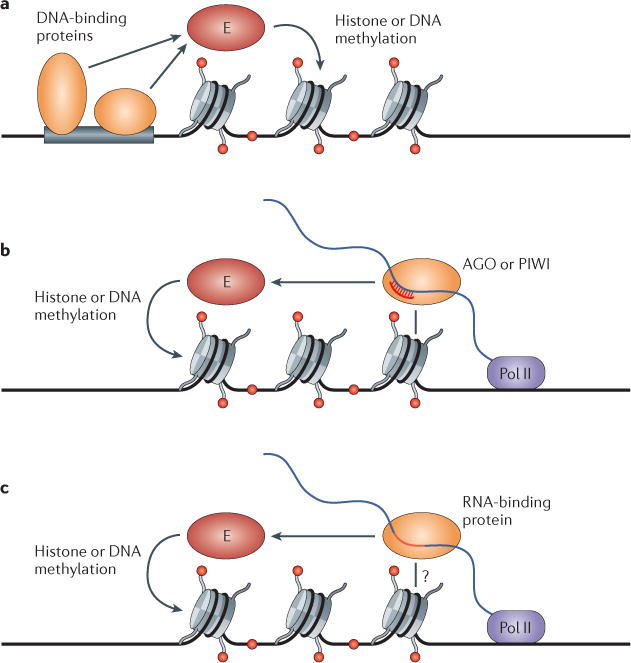
Enzymes (E) that catalyse methylation of histone tails or cytosine bases in DNA are recruited to chromatin by distinct mechanisms. a | Sequence-specific DNA-binding proteins recruit histone- or DNA-modifying enzymes to chromatin. b | Small RNAs target an Argonaute (AGO) or PIWI protein to a nascent transcript through base-pairing interactions to recruit modifying enzymes. c | Long RNAs act as scaffolds for RNA-binding proteins to recruit chromatin-modifying complexes. In all cases, the binding of the enzyme or the recruiting factors (for example, AGO–PIWI complexes in part b and RNA-binding protein complexes in part c) to chromatin may be enhanced by interactions with pre-existing modifications, which self-reinforce the epigenetic state. Pol II, RNA polymerase II.
Despite the considerable progress outlined in this Review, several important questions about the biogenesis and function of non-coding RNAs remain unanswered. The mechanisms that distinguish between different types of transcription and that trigger the generation of different classes of small RNAs remain to be fully understood, although the available evidence indicates a major role for RNA processing events that act co-transcriptionally to determine whether a nascent transcript becomes a functional mRNA or is marked for processing by RNAi and other surveillance mechanisms. Finally, the mechanisms by which lncRNAs participate in the recruitment of Polycomb proteins and other chromatin-modifying activities, particularly the molecular basis of specificity, remain poorly defined. We can look forward to answers to these questions and, if the recent past is a guide, to more exciting and unexpected discoveries about the roles of RNA in gene regulation.
Supplementary Material
Acknowledgments
D.H. was supported by the US National Science Foundation Graduate Research Fellowship Program. Research in D.M.’s laboratory is supported by grants from the US National Institutes of Health. D.M. is an Investigator of the Howard Hughes Medical Institute.
Glossary
- RNA interference
(RNAi). Broadly refers to RNA silencing pathways that use Argonaute and PIWI proteins and small RNAs to silence gene expression
- Small interfering RNAs
(siRNAs). 22–24-nucleotide small RNAs that are generated from longer double-stranded RNA precursors by the ribonuclease Dicer
- Heterochromatin
Regions of chromatin that retain the condensed appearance of mitotic chromosomes throughout the cell cycle. Heterochromatic regions are associated with repressive histone modifications and structural proteins, and are transcriptionally silent
- Argonaute
(AGO). A family of proteins that bind to small RNAs and that are conserved in all domains of life. They mediate target recognition via base-pairing interactions between their bound small RNA and complementary coding or non-coding RNAs
- Heterochromatic gene silencing
Silencing of gene expression within heterochromatin. It was originally thought to exclusively involve transcriptional gene silencing mechanisms, but recent findings indicate that co-transcriptional degradation of nascent RNA, or co-transcriptional gene silencing, also play important parts in silencing
- Pericentromeric DNA repeat
A repeated DNA sequence that surrounds the centromeres of most eukaryotic chromosomes. These repeats are assembled into heterochromatin, which has been demonstrated to have roles in cohesin recruitment in fission yeast and mammals, and de novo centromere assembly in fission yeast
- RNA-induced transcriptional silencing
(RITS). A protein complex first identified in Schizosaccharomyces pombe. In addition to Argonaute 1, the RITS complex contains a GW domain protein, Tas3, and a chromodomain protein, Chp1, which tether the complex to the chromosome via interactions with nascent long non-coding RNAs and nucleosomes with methylated histone H3 lysine 9
- Long terminal repeat
(LTR). A DNA sequence that is repeated at the ends of retrotransposons or pro-viral DNA that is formed from retroviral RNA by reverse transcription. Plant and mammalian genomes contain thousands of LTRs
- Epigenetic phenomena
Phenomena in which changes in gene expression occur without a corresponding change in the DNA sequence; such changes are stable in the absence of initiating signals.
- Paramutation
The ability of a silent allele to convert an active allele to the silent (and paramutagenic) form. It was first described in Zea mays
Footnotes
Competing interests statement
The authors declare no competing interests.
SUPPLEMENTARY INFORMATION
See online article: S1 (figure)
References
- 1.Moazed D. Small RNAs in transcriptional gene silencing and genome defence. Nature. 2009;457:413–420. doi: 10.1038/nature07756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Cech TR, Steitz JA. The noncoding RNA revolution — trashing old rules to forge new ones. Cell. 2014;157:77–94. doi: 10.1016/j.cell.2014.03.008. [DOI] [PubMed] [Google Scholar]
- 3.Hammond SM, Boettcher S, Caudy AA, Kobayashi R, Hannon GJ. Argonaute2, a link between genetic and biochemical analyses of RNAi. Science. 2001;293:1146–1150. doi: 10.1126/science.1064023. [DOI] [PubMed] [Google Scholar]
- 4.Hutvagner G, Zamore PD. A microRNA in a multiple-turnover RNAi enzyme complex. Science. 2002;297:2056–2060. doi: 10.1126/science.1073827. [DOI] [PubMed] [Google Scholar]
- 5.Meister G, et al. Human Argonaute2 mediates RNA cleavage targeted by mi RNAs and siRNAs. Mol Cell. 2004;15:185–197. doi: 10.1016/j.molcel.2004.07.007. [DOI] [PubMed] [Google Scholar]
- 6.Hamilton AJ, Baulcombe DC. A species of small antisense RNA in posttranscriptional gene silencing in plants. Science. 1999;286:950–952. doi: 10.1126/science.286.5441.950. [DOI] [PubMed] [Google Scholar]
- 7.Dalmay T, Hamilton A, Mueller E, Baulcombe DC. Potato virus X amplicons in Arabidopsis mediate genetic and epigenetic gene silencing. Plant Cell. 2000;12:369–379. doi: 10.1105/tpc.12.3.369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Jones L, et al. RNA–DNA interactions and DNA methylation in post-transcriptional gene silencing. Plant Cell. 1999;11:2291–2301. doi: 10.1105/tpc.11.12.2291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Mette MF, van der Winden J, Matzke MA, Matzke AJ. Production of aberrant promoter transcripts contributes to methylation and silencing of unlinked homologous promoters in trans. EMBO J. 1999;18:241–248. doi: 10.1093/emboj/18.1.241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Volpe TA, et al. Regulation of heterochromatic silencing and histone H3 lysine-9 methylation by RNAi. Science. 2002;297:1833–1837. doi: 10.1126/science.1074973. [DOI] [PubMed] [Google Scholar]
- 11.Reinhart BJ, Bartel DP. Small RNAs correspond to centromere heterochromatic repeats. Science. 2002;297:1831. doi: 10.1126/science.1077183. [DOI] [PubMed] [Google Scholar]
- 12.Matzke M, Matzke AJ, Kooter JM. RNA: guiding gene silencing. Science. 2001;293:1080–1083. doi: 10.1126/science.1063051. [DOI] [PubMed] [Google Scholar]
- 13.Fire A, et al. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 1998;391:806–811. doi: 10.1038/35888. [DOI] [PubMed] [Google Scholar]
- 14.Mochizuki K, Fine NA, Fujisawa T, Gorovsky MA. Analysis of a PIWI-related gene implicates small RNAs in genome rearrangement in tetrahymena. Cell. 2002;110:689–699. doi: 10.1016/s0092-8674(02)00909-1. References 6–810, 11 and 14 provided the first evidence for the role of small RNAs in chromatin modification and transcriptional gene silencing. [DOI] [PubMed] [Google Scholar]
- 15.Brennecke J, et al. An epigenetic role for maternally inherited piRNAs in transposon silencing. Science. 2008;322:1387–1392. doi: 10.1126/science.1165171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Guang S, et al. Small regulatory RNAs inhibit RNA polymerase II during the elongation phase of transcription. Nature. 2010;465:1097–1101. doi: 10.1038/nature09095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Gu SG, et al. Amplification of siRNA in Caenorhabditis elegans generates a transgenerational sequence-targeted histone H3 lysine 9 methylation footprint. Nature Genet. 2012;44:157–164. doi: 10.1038/ng.1039. This study provides convincing evidence that classical RNAi in C. elegans results in specific H3K9 methylation at targeted loci. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Rinn JL, Chang HY. Genome regulation by long noncoding RNAs. Annu Rev Biochem. 2012;81:145–166. doi: 10.1146/annurev-biochem-051410-092902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lee JT, Bartolomei MS. X-inactivation, imprinting, and long noncoding RNAs in health and disease. Cell. 2013;152:1308–1323. doi: 10.1016/j.cell.2013.02.016. [DOI] [PubMed] [Google Scholar]
- 20.Schulz EG, Heard E. Role and control of X chromosome dosage in mammalian development. Curr Opin Genet Dev. 2013;23:109–115. doi: 10.1016/j.gde.2013.01.008. [DOI] [PubMed] [Google Scholar]
- 21.Yang PK, Kuroda MI. Noncoding RNAs and intranuclear positioning in monoallelic gene expression. Cell. 2007;128:777–786. doi: 10.1016/j.cell.2007.01.032. [DOI] [PubMed] [Google Scholar]
- 22.Bonasio R, Shiekhattar R. Regulation of transcription by long noncoding RNAs. Annu Rev Genet. 2014;48:433–455. doi: 10.1146/annurev-genet-120213-092323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Verdel A, et al. RNAi-mediated targeting of heterochromatin by the RITS complex. Science. 2004;303:672–676. doi: 10.1126/science.1093686. This study identifies a nuclear AGO complex, RITS, which uses Dicer-generated small RNAs to target specific chromosome regions for heterochromatin formation. The RITS complex provided the first direct physical link between RNAi and heterochromatin. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Partridge JF, Scott KS, Bannister AJ, Kouzarides T, Allshire RC. Cis-acting DNA from fission yeast centromeres mediates histone H3 methylation and recruitment of silencing factors and cohesin to an ectopic site. Curr Biol. 2002;12:1652–1660. doi: 10.1016/s0960-9822(02)01177-6. [DOI] [PubMed] [Google Scholar]
- 25.El-Shami M, et al. Reiterated WG–GW motifs form functionally and evolutionarily conserved Argonaute-binding platforms in RNAi-related components. Genes Dev. 2007;21:2539–2544. doi: 10.1101/gad.451207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Till S, et al. A conserved motif in Argonaute-interacting proteins mediates functional interactions through the Argonaute PIWI domain. Nature Struct Mol Biol. 2007;14:897–903. doi: 10.1038/nsmb1302. [DOI] [PubMed] [Google Scholar]
- 27.Motamedi MR, et al. Two RNAi complexes, RITS and RDRC, physically interact and localize to noncoding centromeric RNAs. Cell. 2004;119:789–802. doi: 10.1016/j.cell.2004.11.034. This study reports the first usage of RNA immunoprecipitation experiments for a chromatin-associated protein. [DOI] [PubMed] [Google Scholar]
- 28.Buhler M, Verdel A, Moazed D. Tethering RITS to a nascent transcript initiates RNAi- and heterochromatin-dependent gene silencing. Cell. 2006;125:873–886. doi: 10.1016/j.cell.2006.04.025. [DOI] [PubMed] [Google Scholar]
- 29.Buhler M, Haas W, Gygi SP, Moazed D. RNAi-dependent and -independent RNA turnover mechanisms contribute to heterochromatic gene silencing. Cell. 2007;129:707–721. doi: 10.1016/j.cell.2007.03.038. This was the first study to demonstrate a major role for co-transcriptional RNAi-, and TRAMP- and exosome-dependent RNA degradation in heterochromatic gene silencing. Together with references 27 and 28, it provides evidence for nascent non-coding RNAs as scaffolds for the assembly of RITS and other complexes that mediate heterochromatin formation. [DOI] [PubMed] [Google Scholar]
- 30.Motamedi MR, et al. HP1 proteins form distinct complexes and mediate heterochromatic gene silencing by nonoverlapping mechanisms. Mol Cell. 2008;32:778–790. doi: 10.1016/j.molcel.2008.10.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sugiyama T, et al. SHREC, an effector complex for heterochromatic transcriptional silencing. Cell. 2007;128:491–504. doi: 10.1016/j.cell.2006.12.035. [DOI] [PubMed] [Google Scholar]
- 32.Moazed D. Mechanisms for the inheritance of chromatin states. Cell. 2011;146:510–518. doi: 10.1016/j.cell.2011.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Nakayama J, Rice JC, Strahl BD, Allis CD, Grewal SI. Role of histone H3 lysine 9 methylation in epigenetic control of heterochromatin assembly. Science. 2001;292:110–113. doi: 10.1126/science.1060118. [DOI] [PubMed] [Google Scholar]
- 34.Hong EJ, Villen J, Gerace EL, Gygi SP, Moazed D. A cullin E3 ubiquitin ligase complex associates with Rik1 and the Clr4 histone H3-K9 methyltransferase and is required for RNAi-mediated heterochromatin formation. RNA Biol. 2005;2:106–111. doi: 10.4161/rna.2.3.2131. [DOI] [PubMed] [Google Scholar]
- 35.Horn PJ, Bastie JN, Peterson CL. A Rik1-associated, cullin-dependent E3 ubiquitin ligase is essential for heterochromatin formation. Genes Dev. 2005;19:1705–1714. doi: 10.1101/gad.1328005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Jia S, Kobayashi R, Grewal SI. Ubiquitin ligase component Cul4 associates with Clr4 histone methyltransferase to assemble heterochromatin. Nature Cell Biol. 2005;7:1007–1013. doi: 10.1038/ncb1300. [DOI] [PubMed] [Google Scholar]
- 37.Li F, et al. Two novel proteins, dos1 and dos2, interact with rik1 to regulate heterochromatic RNA interference and histone modification. Curr Biol. 2005;15:1448–1457. doi: 10.1016/j.cub.2005.07.021. [DOI] [PubMed] [Google Scholar]
- 38.Bayne EH, et al. Stc1: a critical link between RNAi and chromatin modification required for heterochromatin integrity. Cell. 2010;140:666–677. doi: 10.1016/j.cell.2010.01.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Gerace EL, Halic M, Moazed D. The methyltransferase activity of Clr4Suv39h triggers RNAi independently of histone H3K9 methylation. Mol Cell. 2010;39:360–372. doi: 10.1016/j.molcel.2010.07.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zhang K, Mosch K, Fischle W, Grewal SI. Roles of the Clr4 methyltransferase complex in nucleation, spreading and maintenance of heterochromatin. Nature Struct Mol Biol. 2008 doi: 10.1038/nsmb.1406. [DOI] [PubMed] [Google Scholar]
- 41.Li F, Martienssen R, Cande WZ. Coordination of DNA replication and histone modification by the Rik1–Dos2 complex. Nature. 2011;475:244–248. doi: 10.1038/nature10161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Zaratiegui M, et al. RNAi promotes heterochromatic silencing through replication-coupled release of RNA Pol II. Nature. 2011;479:135–138. doi: 10.1038/nature10501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Bayne EH, et al. Splicing factors facilitate RNAi-directed silencing in fission yeast. Science. 2008;322:602–606. doi: 10.1126/science.1164029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Chinen M, Morita M, Fukumura K, Tani T. Involvement of the spliceosomal U4 small nuclear RNA in heterochromatic gene silencing at fission yeast centromeres. J Biol Chem. 2010;285:5630–5638. doi: 10.1074/jbc.M109.074393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kallgren SP, et al. The proper splicing of RNAi factors is critical for pericentric heterochromatin assembly in fission yeast. PLoS Genet. 2014;10:e1004334. doi: 10.1371/journal.pgen.1004334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Selker EU. Trichostatin A causes selective loss of DNA methylation in Neurospora. Proc Natl Acad Sci USA. 1998;95:9430–9435. doi: 10.1073/pnas.95.16.9430. This paper was the first to propose self-reinforcing relationships between different types of chromatin modifications: DNA methylation and histone deacetylation. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Selker EU. Gene silencing: repeats that count. Cell. 1999;97:157–160. doi: 10.1016/s0092-8674(00)80725-4. [DOI] [PubMed] [Google Scholar]
- 48.Richards EJ, Elgin SC. Epigenetic codes for heterochromatin formation and silencing: rounding up the usual suspects. Cell. 2002;108:489–500. doi: 10.1016/s0092-8674(02)00644-x. [DOI] [PubMed] [Google Scholar]
- 49.Noma K, et al. RITS acts in cis to promote RNA interference-mediated transcriptional and post-transcriptional silencing. Nature Genet. 2004;36:1174–1180. doi: 10.1038/ng1452. [DOI] [PubMed] [Google Scholar]
- 50.Colmenares SU, Buker SM, Buhler M, Dlakic M, Moazed D. Coupling of double-stranded RNA synthesis and siRNA generation in fission yeast RNAi. Mol Cell. 2007;27:449–461. doi: 10.1016/j.molcel.2007.07.007. [DOI] [PubMed] [Google Scholar]
- 51.Hayashi A, et al. Heterochromatin protein 1 homologue Swi6 acts in concert with Ers1 to regulate RNAi-directed heterochromatin assembly. Proc Natl Acad Sci USA. 2012;109:6159–6164. doi: 10.1073/pnas.1116972109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Rougemaille M, et al. Ers1 links HP1 to RNAi. Proc Natl Acad Sci USA. 2012;109:11258–11263. doi: 10.1073/pnas.1204947109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Freitag M, et al. DNA methylation is independent of RNA interference in Neurospora. Science. 2004;304:1939. doi: 10.1126/science.1099709. [DOI] [PubMed] [Google Scholar]
- 54.Honda S, Selker EU. Direct interaction between DNA methyltransferase DIM-2 and HP1 is required for DNA methylation in Neurospora crassa. Mol Cell Biol. 2008;28:6044–6055. doi: 10.1128/MCB.00823-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Dang Y, Li L, Guo W, Xue Z, Liu Y. Convergent transcription induces dynamic DNA methylation at disiRNA loci. PLoS Genet. 2013;9:e1003761. doi: 10.1371/journal.pgen.1003761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Lee HC, et al. The DNA/RNA-dependent RNA polymerase QDE-1 generates aberrant RNA and dsRNA for RNAi in a process requiring replication protein A and a DNA helicase. PLoS Biol. 2010;8:e1000496. doi: 10.1371/journal.pbio.1000496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Ream TS, et al. Subunit compositions of the RNA-silencing enzymes Pol IV and Pol V reveal their origins as specialized forms of RNA polymerase II. Mol Cell. 2009;33:192–203. doi: 10.1016/j.molcel.2008.12.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Matzke MA, Mosher RA. RNA-directed DNA methylation: an epigenetic pathway of increasing complexity. Nature Rev Genet. 2014;15:394–408. doi: 10.1038/nrg3683. [DOI] [PubMed] [Google Scholar]
- 59.Haag JR, et al. In vitro transcription activities of Pol IV, Pol V, and RDR2 reveal coupling of Pol IV and RDR2 for dsRNA synthesis in plant RNA silencing. Mol Cell. 2012;48:811–818. doi: 10.1016/j.molcel.2012.09.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Ye R, et al. Cytoplasmic assembly and selective nuclear import of Arabidopsis Argonaute4/siRNA complexes. Mol Cell. 2012;46:859–870. doi: 10.1016/j.molcel.2012.04.013. [DOI] [PubMed] [Google Scholar]
- 61.Wierzbicki AT, Haag JR, Pikaard CS. Noncoding transcription by RNA polymerase Pol IVb/Pol V mediates transcriptional silencing of overlapping and adjacent genes. Cell. 2008;135:635–648. doi: 10.1016/j.cell.2008.09.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Wierzbicki AT, Ream TS, Haag JR, Pikaard CS. RNA polymerase V transcription guides ARGONAUTE4 to chromatin. Nature Genet. 2009;41:630–634. doi: 10.1038/ng.365. This paper provided the first evidence that nascent Pol V transcripts act as scaffolds for the recruitment of AGO4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.He XJ, et al. NRPD4, a protein related to the RPB4 subunit of RNA polymerase II, is a component of RNA polymerases IV and V and is required for RNA-directed DNA methylation. Genes Dev. 2009;23:318–330. doi: 10.1101/gad.1765209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Gao Z, et al. An RNA polymerase II- and AGO4-associated protein acts in RNA-directed DNA methylation. Nature. 2010;465:106–109. doi: 10.1038/nature09025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Zhong X, et al. Molecular mechanism of action of plant DRM de novo DNA methyltransferases. Cell. 2014;157:1050–1060. doi: 10.1016/j.cell.2014.03.056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Blevins T, et al. A two-step process for epigenetic inheritance in Arabidopsis. Mol Cell. 2014;54:30–42. doi: 10.1016/j.molcel.2014.02.019. This paper shows that histone deacetylation and DNA methylation mark a locus for epigenetic inheritance and siRNA-mediated silencing. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Johnson LM, et al. The SRA methyl-cytosine-binding domain links DNA and histone methylation. Curr Biol. 2007;17:379–384. doi: 10.1016/j.cub.2007.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Law JA, et al. Polymerase IV occupancy at RNA-directed DNA methylation sites requires SHH1. Nature. 2013;498:385–389. doi: 10.1038/nature12178. This paper shows that SHH1 recognizes H3 tails with unmodified lysine 4 and methylated lysine 9 residues, and recruits Pol IV to chromatin to promote siRNA generation. This provides a clear basis for self-reinforcement between histone modification and small RNA biogenesis in plants. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Zhang H, et al. DTF1 is a core component of RNA-directed DNA methylation and may assist in the recruitment of Pol IV. Proc Natl Acad Sci USA. 2013;110:8290–8295. doi: 10.1073/pnas.1300585110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Johnson LM, Law JA, Khattar A, Henderson IR, Jacobsen SE. SRA-domain proteins required for DRM2-mediated de novo DNA methylation. PLoS Genet. 2008;4:e1000280. doi: 10.1371/journal.pgen.1000280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Johnson LM, et al. SRA- and SET-domain-containing proteins link RNA polymerase V occupancy to DNA methylation. Nature. 2014;507:124–128. doi: 10.1038/nature12931. This study shows that the putative methyl DNA-binding proteins SUVH2 and SUVH9 recruit Pol V to sites of pre-existing DNA methylation. By promoting further RNA-directed DNA methylation, SUVH2 and SUVH9 thus act as key members of a self-reinforcing loop. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Liu ZW, et al. The SET domain proteins SUVH2 and SUVH9 are required for Pol V occupancy at RNA-directed DNA methylation loci. PLoS Genet. 2014;10:e1003948. doi: 10.1371/journal.pgen.1003948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Malone CD, Hannon GJ. Small RNAs as guardians of the genome. Cell. 2009;136:656–668. doi: 10.1016/j.cell.2009.01.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Batista PJ, et al. PRG-1 and 21U-RNAs interact to form the piRNA complex required for fertility in C. elegans. Mol Cell. 2008;31:67–78. doi: 10.1016/j.molcel.2008.06.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Das PP, et al. PIWI and piRNAs act upstream of an endogenous siRNA pathway to suppress Tc3 transposon mobility in the Caenorhabditis elegans germline. Mol Cell. 2008;31:79–90. doi: 10.1016/j.molcel.2008.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Huang XA, et al. A major epigenetic programming mechanism guided by piRNAs. Dev Cell. 2013;24:502–516. doi: 10.1016/j.devcel.2013.01.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Le Thomas A, et al. Piwi induces piRNA-guided transcriptional silencing and establishment of a repressive chromatin state. Genes Dev. 2013;27:390–399. doi: 10.1101/gad.209841.112. Together with reference 76, this paper demonstrates that ectopic targeting of Piwi in D. melanogaster using artificially generated piRNAs leads to H3K9 methylation and transcriptional silencing at target loci. This suggests that piRNA-progammed Piwi can interact with nascent transcripts to induce chromatin changes. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Rozhkov NV, Hammell M, Hannon GJ. Multiple roles for Piwi in silencing Drosophila transposons. Genes Dev. 2013;27:400–412. doi: 10.1101/gad.209767.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Sienski G, Donertas D, Brennecke J. Transcriptional silencing of transposons by PIWI and maelstrom and its impact on chromatin state and gene expression. Cell. 2012;151:964–980. doi: 10.1016/j.cell.2012.10.040. This study was the first to show that, in D. melanogaster, Piwi and piRNAs silence transposons at the transcriptional level genome-wide. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Wang SH, Elgin SC. Drosophila Piwi functions downstream of piRNA production mediating a chromatin-based transposon silencing mechanism in female germ line. Proc Natl Acad Sci USA. 2011;108:21164–21169. doi: 10.1073/pnas.1107892109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Brower-Toland B, et al. Drosophila Piwi associates with chromatin and interacts directly with HP1a. Genes Dev. 2007;21:2300–2311. doi: 10.1101/gad.1564307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Mohn F, Sienski G, Handler D, Brennecke J. The Rhino–Deadlock–Cutoff complex licenses noncanonical transcription of dual-strand piRNA clusters in Drosophila. Cell. 2014;157:1364–1379. doi: 10.1016/j.cell.2014.04.031. [DOI] [PubMed] [Google Scholar]
- 83.Le Thomas A, et al. Transgenerationally inherited piRNAs trigger piRNA biogenesis by changing the chromatin of piRNA clusters and inducing precursor processing. Genes Dev. 2014;28:1667–1680. doi: 10.1101/gad.245514.114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Aravin AA, Hannon GJ, Brennecke J. The Piwi–piRNA pathway provides an adaptive defense in the transposon arms race. Science. 2007;318:761–764. doi: 10.1126/science.1146484. [DOI] [PubMed] [Google Scholar]
- 85.Aravin AA, et al. A piRNA pathway primed by individual transposons is linked to de novo DNA methylation in mice. Mol Cell. 2008;31:785–799. doi: 10.1016/j.molcel.2008.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Carmell MA, et al. MIWI2 is essential for spermatogenesis and repression of transposons in the mouse male germline. Dev Cell. 2007;12:503–514. doi: 10.1016/j.devcel.2007.03.001. [DOI] [PubMed] [Google Scholar]
- 87.Kuramochi-Miyagawa S, et al. DNA methylation of retrotransposon genes is regulated by PIWI family members MILI and MIWI2 in murine fetal testes. Genes Dev. 2008;22:908–917. doi: 10.1101/gad.1640708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Watanabe T, et al. Role for piRNAs and noncoding RNA in de novo DNA methylation of the imprinted mouse Rasgrf1 locus. Science. 2011;332:848–852. doi: 10.1126/science.1203919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.De Fazio S, et al. The endonuclease activity of Mili fuels piRNA amplification that silences LINE1 elements. Nature. 2011;480:259–263. doi: 10.1038/nature10547. [DOI] [PubMed] [Google Scholar]
- 90.de Vanssay A, et al. Paramutation in Drosophila linked to emergence of a piRNA-producing locus. Nature. 2012;490:112–115. doi: 10.1038/nature11416. This study reports the first example of paramutation in animals and describes how a locus can become included in the piRNA repertoire. Together with reference 16, it illustrates how small RNAs can serve as the carriers of epigenetic signals. [DOI] [PubMed] [Google Scholar]
- 91.Chandler VL. Paramutation’s properties and puzzles. Science. 2010;330:628–629. doi: 10.1126/science.1191044. [DOI] [PubMed] [Google Scholar]
- 92.Yigit E, et al. Analysis of the C. elegans Argonaute family reveals that distinct Argonautes act sequentially during RNAi. Cell. 2006;127:747–757. doi: 10.1016/j.cell.2006.09.033. [DOI] [PubMed] [Google Scholar]
- 93.Guang S, et al. An Argonaute transports siRNAs from the cytoplasm to the nucleus. Science. 2008;321:537–541. doi: 10.1126/science.1157647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Burkhart KB, et al. A pre-mRNA-associating factor links endogenous siRNAs to chromatin regulation. PLoS Genet. 2011;7:e1002249. doi: 10.1371/journal.pgen.1002249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Burton NO, Burkhart KB, Kennedy S. Nuclear RNAi maintains heritable gene silencing in Caenorhabditis elegans. Proc Natl Acad Sci USA. 2011;108:19683–19688. doi: 10.1073/pnas.1113310108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Ashe A, et al. piRNAs can trigger a multigenerational epigenetic memory in the germline of C. elegans. Cell. 2012;150:88–99. doi: 10.1016/j.cell.2012.06.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Buckley BA, et al. A nuclear Argonaute promotes multigenerational epigenetic inheritance and germline immortality. Nature. 2012;489:447–451. doi: 10.1038/nature11352. This study shows that HRDE-1 is the AGO protein responsible for transgenerational inheritance of classical RNAi-mediated silencing in C. elegans and that this involves H3K9 methylation of silenced genes in the germ line. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Bagijn MP, et al. Function, targets, and evolution of Caenorhabditis elegans piRNAs. Science. 2012;337:574–578. doi: 10.1126/science.1220952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Lee HC, et al. C. elegans piRNAs mediate the genome-wide surveillance of germline transcripts. Cell. 2012;150:78–87. doi: 10.1016/j.cell.2012.06.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Shirayama M, et al. piRNAs initiate an epigenetic memory of nonself RNA in the C. elegans germline. Cell. 2012;150:65–77. doi: 10.1016/j.cell.2012.06.015. Together with references 96, 98 and 99, this paper defines the mechanism by which C. elegans piRNAs recognize and target non-self RNAs for silencing, a memory of which is then maintained by secondary siRNAs and the AGO protein HRDE-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Claycomb JM, et al. The Argonaute CSR-1 and its 22G-RNA cofactors are required for holocentric chromosome segregation. Cell. 2009;139:123–134. doi: 10.1016/j.cell.2009.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Seth M, et al. The C. elegans CSR-1 argonaute pathway counteracts epigenetic silencing to promote germline gene expression. Dev Cell. 2013;27:656–663. doi: 10.1016/j.devcel.2013.11.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Wedeles CJ, Wu MZ, Claycomb JM. Protection of germline gene expression by the C. elegans Argonaute CSR-1. Dev Cell. 2013;27:664–671. doi: 10.1016/j.devcel.2013.11.016. References 102 and 103 demonstrate that the AGO protein CSR-1 protects its targets from repression by the PRG-1–HRDE-1 pathway. Thus, C. elegans uses small RNAs not only to mark non-self RNAs but also to designate expressed sequences as self. [DOI] [PubMed] [Google Scholar]
- 104.Conine CC, et al. Argonautes promote male fertility and provide a paternal memory of germline gene expression in C. elegans. Cell. 2013;155:1532–1544. doi: 10.1016/j.cell.2013.11.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Yang L, Froberg JE, Lee JT. Long noncoding RNAs: fresh perspectives into the RNA world. Trends Biochem Sci. 2014;39:35–43. doi: 10.1016/j.tibs.2013.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Ferrari F, Alekseyenko AA, Park PJ, Kuroda MI. Transcriptional control of a whole chromosome: emerging models for dosage compensation. Nature Struct Mol Biol. 2014;21:118–125. doi: 10.1038/nsmb.2763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Kohlmaier A, et al. A chromosomal memory triggered by Xist regulates histone methylation in X inactivation. PLoS Biol. 2004;2:E171. doi: 10.1371/journal.pbio.0020171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Plath K, et al. Role of histone H3 lysine 27 methylation in X inactivation. Science. 2003;300:131–135. doi: 10.1126/science.1084274. [DOI] [PubMed] [Google Scholar]
- 109.Silva J, et al. Establishment of histone H3 methylation on the inactive X chromosome requires transient recruitment of Eed–Enx1 polycomb group complexes. Dev Cell. 2003;4:481–495. doi: 10.1016/s1534-5807(03)00068-6. [DOI] [PubMed] [Google Scholar]
- 110.Zhao J, Sun BK, Erwin JA, Song JJ, Lee JT. Polycomb proteins targeted by a short repeat RNA to the mouse X chromosome. Science. 2008;322:750–756. doi: 10.1126/science.1163045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Rinn JL, et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell. 2007;129:1311–1323. doi: 10.1016/j.cell.2007.05.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Pandey RR, et al. Kcnq1ot1 antisense noncoding RNA mediates lineage-specific transcriptional silencing through chromatin-level regulation. Mol Cell. 2008;32:232–246. doi: 10.1016/j.molcel.2008.08.022. [DOI] [PubMed] [Google Scholar]
- 113.Maenner S, et al. 2D structure of the A region of Xist RNA and its implication for PRC2 association. PLoS Biol. 2010;8:e1000276. doi: 10.1371/journal.pbio.1000276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.da Rocha ST, et al. Jarid2 is implicated in the initial Xist-induced targeting of PRC2 to the inactive X chromosome. Mol Cell. 2014;53:301–316. doi: 10.1016/j.molcel.2014.01.002. [DOI] [PubMed] [Google Scholar]
- 115.Brockdorff N. Noncoding RNA and Polycomb recruitment. RNA. 2013;19:429–442. doi: 10.1261/rna.037598.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Davidovich C, Zheng L, Goodrich KJ, Cech TR. Promiscuous RNA binding by Polycomb repressive complex 2. Nature Struct Mol Biol. 2013;20:1250–1257. doi: 10.1038/nsmb.2679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Kaneko S, Son J, Shen SS, Reinberg D, Bonasio R. PRC2 binds active promoters and contacts nascent RNAs in embryonic stem cells. Nature Struct Mol Biol. 2013;20:1258–1264. doi: 10.1038/nsmb.2700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Kanhere A, et al. Short RNAs are transcribed from repressed Polycomb target genes and interact with Polycomb repressive complex-2. Mol Cell. 2010;38:675–688. doi: 10.1016/j.molcel.2010.03.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Zhao J, et al. Genome-wide identification of Polycomb-associated RNAs by RIP-seq. Mol Cell. 2010;40:939–953. doi: 10.1016/j.molcel.2010.12.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Kaneko S, et al. Interactions between JARID2 and noncoding RNAs regulate PRC2 recruitment to chromatin. Mol Cell. 2014;53:290–300. doi: 10.1016/j.molcel.2013.11.012. References 114 and 120 show that Jarid2 acts as an essential intermediate in Xist-mediated recruitment of PRC2 to chromatin, and that Xist repeat A is dispensable for PRC2 targeting. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Kim TK, et al. Widespread transcription at neuronal activity-regulated enhancers. Nature. 2010;465:182–187. doi: 10.1038/nature09033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.De Santa F, et al. A large fraction of extragenic RNA Pol II transcription sites overlap enhancers. PLoS Biol. 2010;8:e1000384. doi: 10.1371/journal.pbio.1000384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Orom UA, et al. Long noncoding RNAs with enhancer-like function in human cells. Cell. 2010;143:46–58. doi: 10.1016/j.cell.2010.09.001. This study reports the discovery of eRNAs, which are required to activate gene expression in cis. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Wang KC, et al. A long noncoding RNA maintains active chromatin to coordinate homeotic gene expression. Nature. 2011;472:120–124. doi: 10.1038/nature09819. This paper reports an eRNA that induces chromosomal looping and recruits H3K4 methylation to activate transcription of several genes in the HOXA cluster. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Lai F, et al. Activating RNAs associate with Mediator to enhance chromatin architecture and transcription. Nature. 2013;494:497–501. doi: 10.1038/nature11884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Lam MT, et al. Rev–Erbs repress macrophage gene expression by inhibiting enhancer-directed transcription. Nature. 2013;498:511–515. doi: 10.1038/nature12209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Orom UA, Shiekhattar R. Long noncoding RNAs usher in a new era in the biology of enhancers. Cell. 2013;154:1190–1193. doi: 10.1016/j.cell.2013.08.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Harigaya Y, et al. Selective elimination of messenger RNA prevents an incidence of untimely meiosis. Nature. 2006;442:45–50. doi: 10.1038/nature04881. This study was the first to reveal that the mRNAs of meiosis-specific genes are targeted for destruction during vegetative growth in S. pombe. The authors defined a cis-RNA motif (the DSR) and a trans-protein factor (Mmi1) that are required for this regulation. [DOI] [PubMed] [Google Scholar]
- 129.Yamashita A, et al. Hexanucleotide motifs mediate recruitment of the RNA elimination machinery to silent meiotic genes. Open Biol. 2012;2:120014. doi: 10.1098/rsob.120014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Tashiro S, Asano T, Kanoh J, Ishikawa F. Transcription-induced chromatin association of RNA surveillance factors mediates facultative heterochromatin formation in fission yeast. Genes Cells. 2013;18:327–339. doi: 10.1111/gtc.12038. [DOI] [PubMed] [Google Scholar]
- 131.Zofall M, et al. RNA elimination machinery targeting meiotic mRNAs promotes facultative heterochromatin formation. Science. 2012;335:96–100. doi: 10.1126/science.1211651. This paper shows that the machinery responsible for clearing meiotic mRNAs during vegetative growth also promotes the formation of heterochromatin islands at the corresponding meiotic gene loci. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Sugiyama T, Sugioka-Sugiyama R. Red1 promotes the elimination of meiosis-specific mRNAs in vegetatively growing fission yeast. EMBO J. 2011;30:1027–1039. doi: 10.1038/emboj.2011.32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Lee NN, et al. Mtr4-like protein coordinates nuclear RNA processing for heterochromatin assembly and for telomere maintenance. Cell. 2013;155:1061–1074. doi: 10.1016/j.cell.2013.10.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Shah S, Wittmann S, Kilchert C, Vasiljeva L. LncRNA recruits RNAi and the exosome to dynamically regulate pho1 expression in response to phosphate levels in fission yeast. Genes Dev. 2014;28:231–244. doi: 10.1101/gad.230177.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Egan ED, Braun CR, Gygi SP, Moazed D. Post-transcriptional regulation of meiotic genes by a nuclear RNA silencing complex. RNA. 2014;20:867–881. doi: 10.1261/rna.044479.114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Yamanaka S, Yamashita A, Harigaya Y, Iwata R, Yamamoto M. Importance of polyadenylation in the selective elimination of meiotic mRNAs in growing S. pombe cells. EMBO J. 2010;29:2173–2181. doi: 10.1038/emboj.2010.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Duret L, Chureau C, Samain S, Weissenbach J, Avner P. The Xist RNA gene evolved in eutherians by pseudogenization of a protein-coding gene. Science. 2006;312:1653–1655. doi: 10.1126/science.1126316. [DOI] [PubMed] [Google Scholar]
- 138.Gapp K, et al. Implication of sperm RNAs in transgenerational inheritance of the effects of early trauma in mice. Nature Neurosci. 2014;17:667–669. doi: 10.1038/nn.3695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Rechavi O, et al. Starvation-induced transgenerational inheritance of small RNAs in C. elegans. Cell. 2014;158:277–287. doi: 10.1016/j.cell.2014.06.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Castel SE, Martienssen RA. RNA interference in the nucleus: roles for small RNAs in transcription, epigenetics and beyond. Nature Rev Genet. 2013;14:100–112. doi: 10.1038/nrg3355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Bernstein E, Caudy AA, Hammond SM, Hannon GJ. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature. 2001;409:363–366. doi: 10.1038/35053110. [DOI] [PubMed] [Google Scholar]
- 142.Elbashir SM, Lendeckel W, Tuschl T. RNA interference is mediated by 21- and 22-nucleotide RNAs. Genes Dev. 2001;15:188–200. doi: 10.1101/gad.862301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Zamore PD, Tuschl T, Sharp PA, Bartel DP. RNAi: double-stranded RNA directs the ATP-dependent cleavage of mRNA at 21 to 23 nucleotide intervals. Cell. 2000;101:25–33. doi: 10.1016/S0092-8674(00)80620-0. [DOI] [PubMed] [Google Scholar]
- 144.Parrish S, Fleenor J, Xu S, Mello C, Fire A. Functional anatomy of a dsRNA trigger: differential requirement for the two trigger strands in RNA interference. Mol Cell. 2000;6:1077–1087. doi: 10.1016/s1097-2765(00)00106-4. [DOI] [PubMed] [Google Scholar]
- 145.Song JJ, et al. The crystal structure of the Argonaute2 PAZ domain reveals an RNA binding motif in RNAi effector complexes. Nature Struct Biol. 2003;10:1026–1032. doi: 10.1038/nsb1016. [DOI] [PubMed] [Google Scholar]
- 146.Wang Y, et al. Nucleation, propagation and cleavage of target RNAs in Ago silencing complexes. Nature. 2009;461:754–761. doi: 10.1038/nature08434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Ma JB, Ye K, Patel DJ. Structural basis for overhang-specific small interfering RNA recognition by the PAZ domain. Nature. 2004;429:318–322. doi: 10.1038/nature02519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Ma JB, et al. Structural basis for 5′-end-specific recognition of guide RNA by the A. fulgidus Piwi protein. Nature. 2005;434:666–670. doi: 10.1038/nature03514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Song JJ, Smith SK, Hannon GJ, Joshua-Tor L. Crystal structure of Argonaute and its implications for RISC slicer activity. Science. 2004;305:1434–1437. doi: 10.1126/science.1102514. [DOI] [PubMed] [Google Scholar]
- 150.Matranga C, Tomari Y, Shin C, Bartel DP, Zamore PD. Passenger-strand cleavage facilitates assembly of siRNA into Ago2-containing RNAi enzyme complexes. Cell. 2005;123:607–620. doi: 10.1016/j.cell.2005.08.044. [DOI] [PubMed] [Google Scholar]
- 151.Rand TA, Petersen S, Du F, Wang X. Argonaute2 cleaves the anti-guide strand of siRNA during RISC activation. Cell. 2005;123:621–629. doi: 10.1016/j.cell.2005.10.020. [DOI] [PubMed] [Google Scholar]
- 152.Miyoshi K, Tsukumo H, Nagami T, Siomi H, Siomi MC. Slicer function of Drosophila Argonautes and its involvement in RISC formation. Genes Dev. 2005;19:2837–2848. doi: 10.1101/gad.1370605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Buker SM, et al. Two different Argonaute complexes are required for siRNA generation and heterochromatin assembly in fission yeast. Nature Struct Mol Biol. 2007;14:200–207. doi: 10.1038/nsmb1211. [DOI] [PubMed] [Google Scholar]
- 154.Baulcombe D. RNA silencing in plants. Nature. 2004;431:356–363. doi: 10.1038/nature02874. [DOI] [PubMed] [Google Scholar]
- 155.Dalmay T, Hamilton A, Rudd S, Angell S, Baulcombe DC. An RNA-dependent RNA polymerase gene in Arabidopsis is required for posttranscriptional gene silencing mediated by a transgene but not by a virus. Cell. 2000;101:543–553. doi: 10.1016/s0092-8674(00)80864-8. [DOI] [PubMed] [Google Scholar]
- 156.Sijen T, et al. On the role of RNA amplification in dsRNA-triggered gene silencing. Cell. 2001;107:465–476. doi: 10.1016/s0092-8674(01)00576-1. [DOI] [PubMed] [Google Scholar]
- 157.Stuwe E, Toth KF, Aravin AA. Small but sturdy: small RNAs in cellular memory and epigenetics. Genes Dev. 2014;28:423–431. doi: 10.1101/gad.236414.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Khurana JS, Theurkauf W. piRNAs, transposon silencing, and Drosophila germline development. J Cell Biol. 2010;191:905–913. doi: 10.1083/jcb.201006034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Senti KA, Brennecke J. The piRNA pathway: a fly’s perspective on the guardian of the genome. Trends Genet. 2010;26:499–509. doi: 10.1016/j.tig.2010.08.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Ishizu H, Nagao A, Siomi H. Gatekeepers for Piwi–piRNA complexes to enter the nucleus. Curr Opin Genet Dev. 2011;21:484–490. doi: 10.1016/j.gde.2011.05.001. [DOI] [PubMed] [Google Scholar]
- 161.Brennecke J, et al. Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell. 2007;128:1089–1103. doi: 10.1016/j.cell.2007.01.043. [DOI] [PubMed] [Google Scholar]
- 162.Gunawardane LS, et al. A slicer-mediated mechanism for repeat-associated siRNA 5′ end formation in Drosophila. Science. 2007;315:1587–1590. doi: 10.1126/science.1140494. References 161 and 162 reported the discovery of piRNA amplification by the ping-pong mechanism. [DOI] [PubMed] [Google Scholar]
- 163.Halic M, Moazed D. Dicer-independent primal RNAs trigger RNAi and heterochromatin formation. Cell. 2010;140:504–516. doi: 10.1016/j.cell.2010.01.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Yu R, Jih G, Iglesias N, Moazed D. Determinants of heterochromatic siRNA biogenesis and function. Mol Cell. 2014;53:262–276. doi: 10.1016/j.molcel.2013.11.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Kawakami K, Hayashi A, Nakayama J, Murakami Y. A novel RNAi protein, Dsh1, assembles RNAi machinery on chromatin to amplify heterochromatic siRNA. Genes Dev. 2012;26:1811–1824. doi: 10.1101/gad.190272.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Marasovic M, Zocco M, Halic M. Argonaute and Triman generate dicer-independent priRNAs and mature siRNAs to initiate heterochromatin formation. Mol Cell. 2013;52:173–183. doi: 10.1016/j.molcel.2013.08.046. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.