

Abstract
The cashew nut releases a substance that is known as cashew nut shell liquid (CNSL). There are both natural (iCNSL) and technical (tCNSL) cashew nut shell liquids. This study used an Artemia salina bioassay to evaluate the toxic effects of iCNSL and tCNSL cashew nut shell liquids. It also evaluated the toxicity, cytotoxicity, and mutagenicity of CNSL and its effects on the damage induced by copper sulfate (CuSO4·5H2O) on the meristems' root of Allium cepa. Effects of the damage induced by CuSO4·5H2O were evaluated before (pre-), during (co-), and after (post-) treatments. The iCNSL contained 94.5% anacardic acid, and the tCNSL contained 91.3% cardanol. The liquids were toxic to A. salina. Toxicity, cytotoxicity, and mutagenicity were observed with iCNSL compared with the negative control. Similarly, iCNSL failed to inhibit the toxicity and cytotoxicity of CuSO4·5H2O. The tCNSL was not toxic, cytotoxic, or mutagenic in any of the concentrations. However, the lowest iCNSL concentrations and all of the tCNSL concentrations had preventive, antimutagenic, and reparative effects on micronuclei and on chromosomal aberrations in the A. cepa. Therefore, protective, modulating, and reparative effects may be observed in the A. cepa, depending on the concentration and type of CNSL used.
1. Introduction
Recent epidemiological studies have shown that medicinal plants may be involved in preventing or delaying the development of various diseases [1, 2]. These plants may act on different targets in signal transduction pathways that may modulate gene expression, cell cycle progression, cellular proliferation, and/or apoptosis [3, 4]. However, adverse effects, such as genotoxicity, mutagenicity, and carcinogenicity [5, 6], can also occur. These effects may be triggered by compounds that interact with deoxyribonucleic acid (DNA), which would cause cellular toxicity and/or genotoxicity [7].
The species Anacardium occidentale (Anacardiaceae) is found in tropical regions worldwide. It is common in Brazil, India, Mozambique, Tanzania, Kenya, Vietnam, Indonesia, and Thailand [8]. Recent studies have been associated with several biological effects of the cashew plant. It can be used as an antioxidant [9, 10] and can be used in dermatitis [11] and also possesses larvicidal [12], antigenotoxic [13], and antimicrobial [14] activities.
The cashew nut releases a liquid that is known as cashew nut shell liquid (CNSL). This liquid is a natural source of phenolic compounds that contribute to its antioxidant [15, 16], antifungal [17], antibacterial [18], larvicidal [19], and nongenotoxic effects in prokaryotic [20, 21] and eukaryotic cells [22]. This liquid is classified into two categories, depending on the extraction method used: natural CNSL (iCNSL) extracted with solvents, and its main components are anacardic acid (62.9%), cardol (23.98%), and cardanol (6.99%) [23] and technical CNSL (tCNSL) wich is prepared by burning the nuts industrially at high temperatures and it contains cardanol (60–65%), cardol (15–20%), polymeric material (10%), and small amounts of metilcardol [24].
The present study aimed to evaluate the toxicity of iCNSL and tCNSL to A. salina and to determine their toxic, cytotoxic, and mutagenic actions and their protective effects against the damage that is induced by CuSO4·5H2O in A. cepa root meristems.
2. Material and Methods
2.1. CNSL Preparation and Doses Choice
Ripe cashew nuts were collected from cashew trees in Teresina in the state of Piauí, Brazil. For iCNSL, extraction the ripe cashew nuts were stored in styrofoam with liquid nitrogen for 5 minutes. Then, they were crushed and subjected to hot extraction Soxhlet extractor with hexane for 16 hours. The iCNSL was then concentrated in a rotary evaporator at 45°C. The tCNSL was provided by a company of the Group Europa–Castanha located in Altos, Teresina, Piauí, in northeastern Brazil. According to the company, the nuts were immersed in a hot bath at 195°C for 3 hours to extract the tCNSL. The tCNSL was then filtered and stored.
The lowest doses used in this study were chosen because recent research on the tCNSL (100–500 µg/mL) has shown antioxidant properties. Thus, doses lower than those tested in the literature have been evaluated to check if they would still be antioxidant and nonmutagenic.
2.2. CNSL Methylation Reaction
Samples of the extracted iCNSL and tCNSL were analysed by gas chromatography coupled to mass spectrometry (GC-MS) in the form of methyl esters. Five milligrams of CNSL was dissolved in 0.5 mL of diethyl ether and transferred to a 5.0 mL flask. A solution of diazomethane in ether (2.0 mL) was then added dropwise at low temperature in an ice bath until outgassing was no longer observed. The flask was sealed with a ground glass stopper and magnetically stirred continuously at room temperature. After 3 hours, the reaction was monitored by thin layer chromatography (TLC). Following ether evaporation, the residue was solubilised in ethyl acetate and analysed by GC-MS [25].
2.3. GC-MS Analysis
Both of the derived CNSL samples were then analysed in a GC-MS system (Shimadzu, GC-17A/MS-QP5050A). The column chromatography DB-5HT (J & W Scientific) was 30 m long and 0.25 mm in diameter, had a film thickness of 0.10 µm, and used helium as a carrier gas (1.0 mL/min). The following parameters were used: interface = 270°C, injector = 250°C, initial temperature = 60°C (2 min), 4.0 mL/min to 180°C (4 min), and 10 mL/min to 260°C (10 min). Identification was achieved by comparing mass spectra (43 to 4500 Daltons, electron impact ionisation, 70 eV) and data from the literature of De Lima et al. [25].
2.4. Artemia salina Test
The method used to assess the toxicity of iCNSL and tCNSL on A. salina was adapted from Meyer et al. [26]. The microcrustaceans were hatched in water as mentioned in Materials and Methods for 48 hours until their larvae were released. Ten A. salina specimens were introduced into each of the three tubes containing a 17.37, 34.75, or 69.50 μg·mL−1 concentration of iCNSL or tCNSL. A nonactive substance (1 : 1 seawater and mineral water) was used as a control. The dead specimens were counted under a stereomicroscope after 24 hours.
2.5. Allium cepa Test
The A. cepa test was adapted from the method reported by Fiskesjö [27]. Each experimental group consisted of iCNSL and tCNSL at concentrations of 17.37, 34.75, or 69.50 μg·mL−1, as well as a solution of 500 µg/mL Tween 20 (solvent), a negative control (dechlorinated water), and a positive control (1.2 μg·mL−1 copper sulphate). Small bulbs of A. cepa (2n = 16) were purchased from supermarkets in Teresina, Piauí.
Following 72 hours of exposure, the roots were measured in centimetres to assess toxicity. The roots were then placed in Carnoy's fixative solution (ethanol/glacial acetic acid 3 : 1 v/v), refrigerated at 4°C for 24 hours, followed by 70% ethanol solution and refrigeration. The roots were subsequently hydrolysed in a hydrochloric acid solution (1 N) and placed in a staining solution (Schiff's dye) for two hours.
The roots were then placed on slides and sectioned in the meristem region. This region of the root was stained with 2% acetic carmine, covered with a cover slip, and then observed under an optical microscope (1000x) to analyse cytotoxicity, mutagenicity, and the effects on the damage induced by copper sulphate. A total of 1,000 cells were analysed on each slide. The following parameters were observed: (a) mitotic index (MI), (b) the frequency of chromosomal aberrations (CA) in anaphase and telophase, and (c) the frequency of micronuclei (MN).
2.6. Effects on Copper Sulphate-Induced Damage
The method used to evaluate antimutagenicity by means of the A. cepa test was adapted from Malini et al. [28]. The present study used CuSO4·5H2O as the genotoxic agent because of its mutagenic potential [29, 30].
Three types of treatments were standardised: (1) pretreatment, in which the bulbs were exposed initially to iCNSL or tCNSL for 48 hours and the roots were then washed in distilled water and placed in 1.2 μg·mL−1 CuSO4·5H2O solutions for 24 hours to germinate; (2) cotreatment, in which the bulbs were placed in tubes that contained iCNSL or tCNSL and a 1.2 μg·mL−1 CuSO4·5H2O solution at a 1 : 1 ratio for 72 hours for germination; and (3) posttreatment, in which the bulbs were first placed in 1.2 μg·mL−1 CuSO4·5H2O solution for 48 hours, and then the roots were rinsed with distilled water and placed in tubes that contained iCNSL or tCNS for 24 hours for germination. The samples were then processed in the similar way as mentioned in the A. cepa test.
2.7. Statistical Analysis
The number of dead A. salina was analysed using the Statistical Package for the Social Sciences (SPSS) software, version 17:0. The IC50 was assessed by probit analysis. The tests were performed in triplicate. The data were analysed using GraphPad Prism software (version 6.03), and the experimental groups were compared with the negative and positive control groups. All results were expressed as mean ± standard deviation (SD). The data were assessed by an analysis of variance (ANOVA) followed by Tukey's test for multiple comparisons for genotoxicity and mutagenicity tests The significance levels were * P < 0.05, ** P < 0.01, and *** P < 0.001.
3. Results
3.1. CNSL Chromatographic Analyses
The main phenolic compounds present in both types of CNSL were identified by GC-MS analysis. The iCNSL contained metilcardol (2.90%), monounsaturated anacardic acid (82.90%), diunsaturated anacardic acid (8.0%), anacardic acid (3.60%), and 2.60% unidentified compounds (Table 1 and Figure 1). The tCNSL exhibited 79.40% monounsaturated cardanol, 8.67% diunsaturated cardanol, 3.23% cardanol, and 8.70% unidentified compounds (Table 1 and Figure 2). These results indicate that the main phenolic compound of iCNSL is monounsaturated anacardic acid and monounsaturated cardanol for the tCNSL.
Table 1.
The investigated components of iCNSL and tCNSL.
| Natural iCNSL* | |||
|---|---|---|---|
| Peak | Retention time (min) | Compounds | Yield |
| 1 | 36.80 | Metilcardol | 2.90 |
| 2 | 37.60 | Monounsaturated anacardic acid | 82.90 |
| 3 | 37.62 | Diunsaturated anacardic acid | 8.00 |
| 4 | 37.70 | Anacardic acid | 3.60 |
| 5 | 38.05 | Unidentified | 2.60 |
|
| |||
| Technical tCNSL | |||
| Peak | Retention time (min) | Compounds | Yield |
|
| |||
| 6 | 18.08 | Monounsaturated cardanol | 79.40 |
| 7 | 18.10 | Diunsaturated cardanol | 8.67 |
| 8 | 18.17 | Cardanol | 3.23 |
| 9 | 20.71 | Unidentified | 8.70 |
*Analysed in the form of methyl esters.
Figure 1.
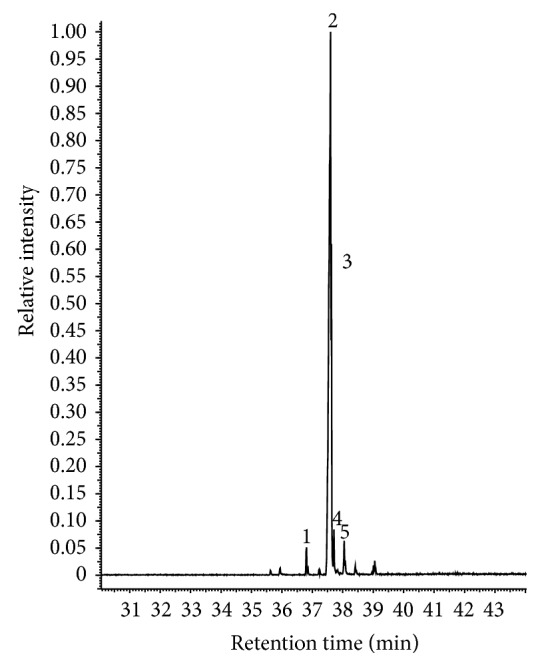
The chromatographic profile of iCNSL obtained by GC-MS analysis. 1: metilcardol, 2: monounsaturated anacardic acid, 3: diunsaturated anacardic acid, 4: anacardic acid, and 5: unidentified.
Figure 2.
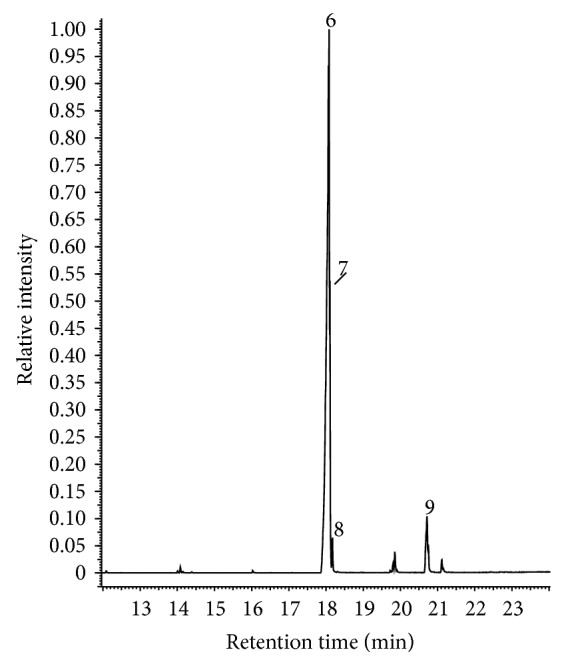
The chromatographic profile of tCNSL obtained by GC-MS analysis. 6: monounsaturated cardanol, 7: diunsaturated cardanol, 8: cardanol, and 9: unidentified.
3.2. Evaluation of iCNSL and tCNSL Toxicity on A. salina
The toxicity of iCNSL and tCNSL on A. salina was evaluated, and the 50% lethal concentration (IC50) values are shown in Figure 3. The IC50 of iCNSL and tCNSL was 36.96 μg·mL−1 and 91.67 μg·mL−1, respectively.
Figure 3.
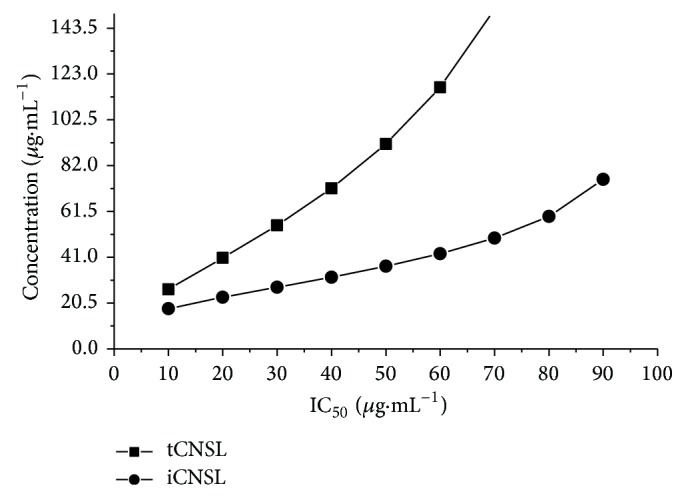
The lethal concentration (LC50) of iCNSL and tCNSL in A. salina toxicity test. Mean of three independent experiments at different iCNSL and tCNSL concentrations.
3.3. Evaluation of iCNSL and tCNSL Toxicity and Cytotoxicity in A. cepa
Analysis of the macroscopic parameter (root growth), which is affected by toxicity in A. cepa root meristems, showed that iCNSL had a significant toxic effect at the highest test concentration (69.50 μg·mL−1) compared with the negative control (P < 0.001). However, tCNSL had no toxic effect (P > 0.05) (Figure 4).
Figure 4.
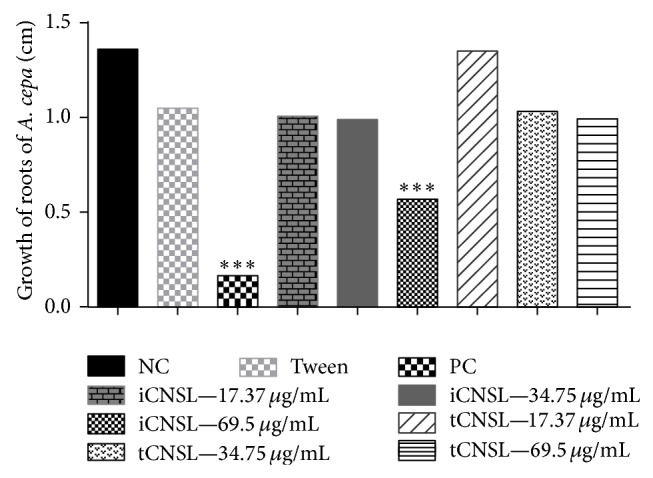
Toxic effects of iCNSL and tCNSL observed by roots growth in A. cepa (cm). Significant compared with the negative control. * P < 0.05; ** P < 0.01 ANOVA. Tukey's test for multiple comparisons between groups.
When A. cepa root meristems were initially exposed to iCNSL or tCNSL in pretreatment and were then treated with a CuSO4·5H2O solution at a 1.2 μg·mL−1 concentration, iCNSL failed to prevent the toxicity of the CuSO4·5H2O and tCNSL (cotreatment) protected against copper sulphate toxicity only at the highest concentration (69.50 μg·mL−1). However, both CNSLs (iCNSL and tCNSL) showed co- or posttreatment effects on CuSO4·5H2O toxicity (Table 2).
Table 2.
Antitoxicity of iCNSL and tCNSL as measured by the effect on A. cepa root growth (cm; mean ± SD, and % growth inhibition) at different concentrations and treatments with copper sulphate (1.2 μg·mL−1).
| Treatments (% inhibition) |
Controls/vehicle | iCNSL | tCNSL | ||||||
|---|---|---|---|---|---|---|---|---|---|
| NC | PC | TWEEN | Concentrations | Concentrations | |||||
| 17.37 μg·mL−1 | 34.75 μg·mL−1 | 69.50 μg·mL−1 | 17.37 μg·mL−1 | 34.75 μg·mL−1 | 69.50 μg·mL−1 | ||||
| Pretreatment (% inhibition) |
NT | NT | NT | 0.51 ± 0.17a∗∗∗,b∗∗
(62.78%) |
0.55 ± 0.18a∗∗∗,b∗∗
(59.86%) |
0.55 ± 0.12a∗∗∗,b∗∗
(59.86%) |
0.45 ± 0.23a∗∗∗.b∗
(67.16%) |
0.71 ± 0.26a∗∗∗,b∗∗∗
(48.18%) |
1.34 ± 0.25b∗∗∗
(2.19%) |
| Cotreatment (% inhibition) |
NT | NT | NT | 0.23 ± 0.09a∗∗∗
(83.09%) |
0.34 ± 0.27a∗∗∗
(75.00%) |
0.38 ± 0.24a∗∗∗
(72.03%) |
0.46 ± 0.09a∗∗∗;b∗
(66.18%) |
0.48 ± 0.20a∗∗∗;b∗
(64.71%) |
0.52 ± 0.32a∗∗∗;b∗∗
(61.23%) |
| Posttreatment (% inhibition) |
NT | NT | NT | 0.32 ± 0.09a∗∗∗
(83.09%) |
0.33 ± 0.11a∗∗∗
(75.00%) |
0.40 ± 0.19a∗∗∗;b∗
(72.00%) |
0.36 ± 0.07a∗∗∗
(66.18%) |
0.38 ± 0.09a∗∗∗
(64.71%) |
0.32 ± 0.05a∗∗∗
(61.77%) |
NC (dechlorinated water). PC (copper sulphate solution). TWEEN (solvent). NT (not tested). 1.000 cells evaluated per bulb, totalling 5,000 cells per group. aSignificant compared with the negative control; bsignificant compared with the positive control. * P < 0.05, ** P < 0.01, and *** P < 0.001 ANOVA. Tukey's test for multiple comparisons between groups.
The mitotic indices (MIs) of A. cepa root meristems that were exposed to different iCNSL and tCNSL concentrations are shown in Figure 5. The iCNSL significantly inhibited (P < 0.05) cell division in A. cepa root meristems at only the highest test concentration (69.50 μg·mL−1). The tCNSL had no cytotoxic effects on MI at any of the test concentrations.
Figure 5.
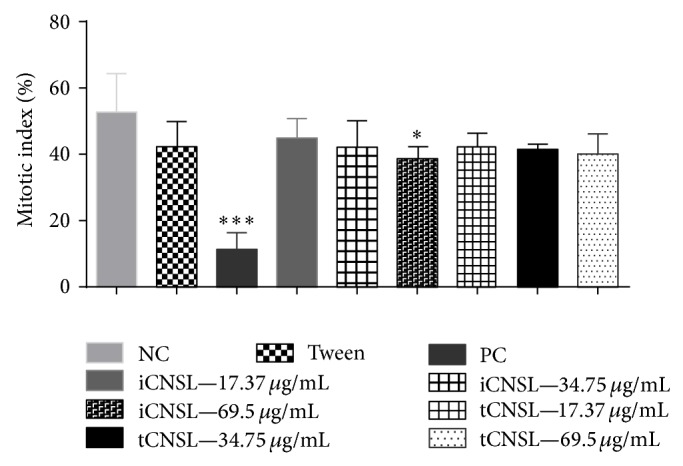
Evaluation of cytotoxic effects of iCNSL and tCNSL observed by inhibition of mitotic index in roots of A. cepa. Significant compared with the negative control. * P < 0.05; ** P < 0.01 ANOVA. Tukey's test for multiple comparisons between groups.
Both iCNSL and tCNSL failed to prevent copper sulphate cytotoxicity in A. cepa root meristems in pretreatment, and neither modulated the cytotoxicity of copper sulphate in cotreatment (Table 3).
Table 3.
Anticytotoxicity of iCNSL and tCNSL as measured by the effect on the cell division rate of A. cepa roots (cm; mean ± standard deviation, and % cell division inhibition) at different concentrations and treatments with copper sulphate (1.2 μg·mL−1).
| Treatments (% inhibition) |
Controls/vehicle | iCNSL | tCNSL | ||||||
|---|---|---|---|---|---|---|---|---|---|
| NC | PC | TWEEN | Concentrations | Concentrations | |||||
| 17.37 µg·mL−1 | 34.75 µg·mL−1 | 69.50 µg·mL−1 | 17.37 µg·mL−1 | 34.75 µg·mL−1 | 69.50 µg·mL−1 | ||||
| Pretreatment (% inhibition) |
NT | NT | NT | 29.28 ± 6.37a∗∗∗;b∗∗
(44.54%) |
23.78 ± 0.68a∗∗∗
(54.96%) |
28.14 ± 1.33a∗∗∗;b∗∗
(46.70%) |
8.92 ± 0.75a∗∗∗
(83.10%) |
9.68 ± 1.46a∗∗∗
(81.66%) |
13.54 ± 3.44a∗∗∗
(74.35%) |
| Cotreatment (% inhibition) |
NT | NT | NT | 24.48 ± 3.89a∗∗∗;b∗
(53.63%) |
23.16 ± 1.91a∗∗∗
(56.13%) |
27.28 ± 2.16a∗∗∗;b∗∗
(48.33%) |
8.38 ± 0.84a∗∗∗
(84.12%) |
9.12 ± 1.46a∗∗∗
(82.72%) |
14.84 ± 2.76a∗∗∗
(71.89%) |
| Posttreatment (% inhibition) |
NT | NT | NT | 25.64 ± 1.86a∗∗∗
(51.43%) |
20.64 ± 2.85a∗∗∗
(60.90%) |
28.52 ± 1.58a∗∗∗;b∗∗∗
(45.98%) |
13.16 ± 1.23a∗∗∗
(75.07%) |
21.72 ± 2.08a∗∗∗
(58.86%) |
32.00 ± 1.30a∗∗∗;b∗∗∗
(39.39%) |
NC (dechlorinated water). PC (copper sulphate solution). TWEEN (solvent). NT (not tested). 1.000 cells evaluated per bulb, totalling 5,000 cells per group. aSignificant compared with the negative control; bsignificant compared with the positive control. * P < 0.05, ** P < 0.01, and *** P < 0.001 ANOVA. Tukey's test for multiple comparisons between groups.
Although iCNSL and tCNSL inhibited (P < 0.001) the cytotoxic activity of copper sulphate at the highest posttreatment test concentration, they also significantly reduced A. cepa root growth (P < 0.001) at this higher concentration (Table 3). These data indicate that neither iCNSL nor tCNSL showed preventive, modulating, and reparative activity against the cytotoxicity induced in A. cepa root meristems by CuSO4·5H2O.
3.4. Evaluation of iCNSL and tCNSL Mutagenicity and Their Effects on Copper Sulphate
Only iCNSL at the highest test concentration (69.50 µg·mL−1) induced MN mutagenicity in A. cepa root meristems (P < 0.05) compared with the negative control. The two lower concentrations of iCNSL and all three concentrations of tCNSL failed to induce MN mutagenicity in A. cepa root meristems (P > 0.05) (Table 4).
Table 4.
Mutagenicity and antimutagenicity of iCNSL and tCNSL as measured by the number of micronuclei (mean ± SD of 1.000 cells per slide, 5 slides per test group) in A. cepa root meristems at different concentrations and treatments with copper sulphate (1.2 µg·mL−1).
| Treatments | Controls/vehicle | iCNSL | tCNSL | ||||||
|---|---|---|---|---|---|---|---|---|---|
| NC | PC | TWEEN | Concentrations | Concentrations | |||||
| 17.37 µg·mL−1 | 34.75 µg·mL−1 | 69.50 µg·mL−1 | 17.37 µg·mL−1 | 34.75 µg·mL−1 | 69.50 µg·mL−1 | ||||
| Without treatment | 1.20 ± 1.09 | 8.40 ± 1.14a∗∗ | 11.80 ± 0.83 | 1.80 ± 0.83 | 1.80 ± 1.48 | 4.00 ± 0.70∗a | 1.00 ± 1.22 | 1.20 ± 0.83 | 2.20 ± 0.83 |
| Pretreatment | NT | NT | NT | 1.40 ± 1.34b∗∗∗ | 1.00 ± 0.90b∗∗∗ | 4.20 ± 1.30a∗;b∗∗∗ | 2.20 ± 2.16b∗∗∗ | 1.80 ± 0.90b∗∗∗ | 2.00 ± 1.41b∗∗∗ |
| Cotreatment | NT | NT | NT | 1.60 ± 0.89b∗ | 1.80 ± 1.48b∗ | 5.40 ± 1.67a∗∗;b∗∗ | 1.00 ± 1.00b∗∗∗ | 1.00 ± 0.70b∗∗∗ | 2.00 ± 1.58b∗∗∗ |
| Posttreatment | NT | NT | NT | 2.2 ± 148b∗∗∗ | 1.00 ± 1.22b∗∗∗ | 4.20 ± 1.48a∗;b∗∗∗ | 1.40 ± 1.14b∗∗∗ | 1.60 ± 1.51b∗∗∗ | 1.80 ± 0.83b∗∗∗ |
NC (dechlorinated water). PC (copper sulphate solution). TWEEN (solvent). NT (not tested). 5.000 cells evaluated per bulb, totalling 5.000 per group. aSignificant compared with the negative control; bsignificant compared with the positive control. * P < 0.05, ** P < 0.01, and *** P < 0.001 ANOVA. Tukey's test for multiple comparisons between groups.
Both iCNSL and tCNSL protect (pretreatment) the DNA of A. cepa root meristems, as observed by the inhibition of MN formation induced by CuSO4·5H2O. However, iCNSL at the highest test concentration did not confer this protection. Similarly, antimutagenic and reparative effects were evidenced by a reduction in MN in co- and posttreatments, except at the highest iCNSL concentration (Table 4). Both iCNSL and tCNSL strongly affected the prevention, modulation, and repair of damage induced by CuSO4·5H2O in A. cepa meristems.
The mutagenicity of the highest iCNSL concentration (69.50 µg·mL−1) was not confirmed when analysing the frequency of chromosomal aberration (CA) because iCNSL at this concentration had already been shown to have mutagenic effects as evidenced by increased MN frequency, and MI inhibition also precluded observation of CA at this concentration. Neither iCNSL nor tCNSL showed mutagenic effects as measured by the frequency of CA compared to the positive control (Figure 6).
Figure 6.
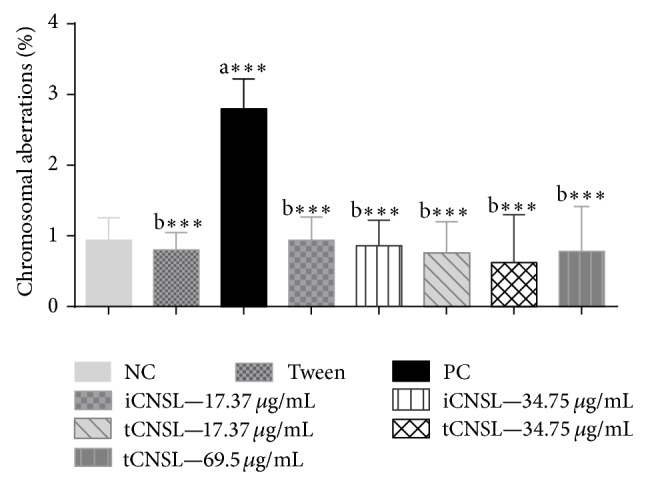
Mutagenic effects of iCNSL and tCNSL by frequency of chromosomal aberrations in A. cepa roots. aSignificant compared with the negative control; bSignificant compared with the positive control. *** P < 0.001 ANOVA. Tukey's test for multiple comparisons between groups.
Chromosome bridges, vagrant and laggard chromosomes, and chromosome fragments are the most notable of the CAs induced by CuSO4·5H2O during anaphase and telophase. The photomicrographic profile of CAs identified in A. cepa root meristems exposed to CuSO4·5H2O is represented in Figure 7. This damage was modulated by iCNSL and tCNSL at the tested concentrations, which suggests that both may have the inhibition mechanisms of aneugenic and/or clastogenic agents.
Figure 7.

The photomicrographic profile of A. cepa root meristems in which pre-, co-, and posttreatment with iCNSL and tCNSL provided protection against the damage caused by exposure to copper sulphate (at a concentration of 1.2 μg/mL). (a) A cell in anaphase with a chromosome bridge and a chromosome fragment, (b) a cell in anaphase with a chromosome bridge, (c) a cell in anaphase with a chromosome bridge and a vagrant chromosome, (d) and (e) cells in anaphase with laggard chromosomes, (f) a cell in anaphase with laggard chromosomes and chromosome fragments, (g) cells in anaphase with laggard and vagrant chromosomes, and (h) and (i) cells in telophase with chromosome fragments.
Moreover, both types of CNSL showed protective, antimutagenic, and DNA-repair effects in regard to the damage induced by copper sulphate (Table 5).
Table 5.
Antimutagenicity of iCNSL and tCNSL as measured by the frequency of chromosomal aberrations (mean ± SD of 1.000 cells per slide, 5 slides per test group) in A. cepa root meristems at different concentrations (pre-, co-, and posttreatment) with copper sulphate (1.2 µg·mL−1).
| Groups with concentration | Chromosome bridges | Chromosomal aberration/5000 cells | Total frequency (%) | |||
|---|---|---|---|---|---|---|
| Vagrant chromosomes | Laggard chromosomes | Fragments | Total frequency | |||
| iCNSL pretreatment | ||||||
| 17,37 µg·mL−1 | 0.40 ± 0.89b∗ | 1.60 ± 0.50b∗∗∗ | 4.00 ± 4.84 | 2.00 ± 1.22 | 8.00 ± 3.46b∗∗∗ | 0.80 ± 0.34b∗∗∗ |
| 34,75 µg·mL−1 | 0.60 ± 0.89b∗ | 1.20 ± 1.09b∗∗∗ | 2.60 ± 2.96b∗ | 2.60 ± 1.94 | 7.00 ± 4.06b∗∗∗ | 0.70 ± 0.40b∗∗∗ |
| tCNSL pretreatment | ||||||
| 17,37 µg·mL−1 | 1.20 ± 1.00 | 1.00 ± 0.00b∗∗∗ | 8.00 ± 4.41 | 1.20 ± 0.83b∗ | 11.40 ± 7.33b∗∗∗ | 1.14 ± 0.73b∗∗∗ |
| 34,75 µg·mL−1 | 0.75 ± 0.50 | 1.20 ± 0.44b∗∗∗ | 8.20 ± 3.11 | 2.40 ± 1.51 | 12.80 ± 3.76b∗∗∗ | 1.28 ± 0.37b∗∗∗ |
| 69,5 µg·mL−1 | 0.80 ± 0.40 | 1.60 ± 0.44b∗∗∗ | 8.00 ± 4.00 | 2.60 ± 1.81 | 13.00 ± 5.61b∗∗∗ | 1.30 ± 0.56b∗∗∗ |
| iCNSL cotreatment | ||||||
| 17,37 µg·mL−1 | 0.40 ± 0.89b∗ | 1.00 ± 0.00b∗∗∗ | 3.40 ± 1.81b∗ | 1.60 ± 1.14 | 6.40 ± 2.51b∗∗∗ | 0.64 ± 0.25b∗∗∗ |
| 34,75 µg·mL−1 | 0.20 ± 0.44b∗ | 1.20 ± 0.44b∗∗∗ | 5.40 ± 2.60 | 1.00 ± 0.70b∗∗ | 7.80 ± 2.77b∗∗∗ | 0.78 ± 0.27b∗∗∗ |
| tCNSL cotreatment | ||||||
| 17,37 µg·mL−1 | 0.60 ± 0.54b∗ | 1.60 ± 1.14b∗∗∗ | 5.20 ± 1.78 | 2.00 ± 1.00 | 9.40 ± 2.51b∗∗∗ | 0.94 ± 0.25b∗∗∗ |
| 34,75 µg·mL−1 | 0.60 ± 0.54b∗ | 1.20 ± 1.64b∗∗∗ | 4.60 ± 2.19 | 2.40 ± 1.14 | 8.80 ± 0.30b∗∗∗ | 8.80 ± 3.03b∗∗∗ |
| 69,50 µg·mL−1 | 1.60 ± 1.94 | 3.40 ± 1.67b∗∗∗ | 8.80 ± 5.35 | 3.40 ± 2.79 | 17.20 ± 5.76b∗∗∗ | 1.72 ± 0.57b∗∗∗ |
| iCNSL posttreatment | ||||||
| 17,37 µg·mL−1 | 0.20 ± 0.44b∗∗ | 1.20 ± 0.83b∗∗∗ | 1.60 ± 1.57b∗∗ | 1.60 ± 1.14 | 4.60 ± 1.81b∗∗∗ | 0.46 ± 0.18b∗∗∗ |
| 34,75 µg·mL−1 | 0.20 ± 0,44b∗ | 2.00 ± 1.00b∗∗∗ | 4.00 ± 4.35 | 2.40 ± 0.89 | 8.60 ± 4.27b∗∗∗ | 0.86 ± 0.42b∗∗∗ |
| tCNSL posttreatment | ||||||
| 17,37 µg·mL−1 | 1.20 ± 0.44 | 1.00 ± 1.22b∗∗∗ | 6.00 ± 4.84 | 1.00 ± 1.22b∗ | 9.20 ± 7.46b∗∗∗ | 0.92 ± 0.74b∗∗∗ |
| 34,75 µg·mL−1 | 0.80 ± 0.44 | 2.60 ± 1.51b∗∗∗ | 7.40 ± 2.52 | 2.60 ± 2.60 | 13.00 ± 6.51b∗∗∗ | 1.30 ± 0.65b∗∗∗ |
| 69,50 µg·mL−1 | 1.20 ± 0.44 | 3.80 ± 1.78b∗∗ | 7.00 ± 4.12 | 2.80 ± 0.83 | 14.80 ± 4.08b∗∗ | 1.48 ± 0.40b∗∗ |
NC (dechlorinated water). PC (copper sulphate solution). TWEEN (solvent). NT (not tested). 5.000 cells evaluated per bulb, totalling 5,000 per group. aSignificant compared with the negative control; bsignificant compared with the positive control. * P < 0.05, ** P < 0.01, and *** P < 0.001 ANOVA. Tukey's test for multiple comparisons between groups.
4. Discussion
Genomic instability is a common cause of cancer. Cancer cells are more susceptible than normal cells to DNA-damaging agents. This increased susceptibility provides a path for therapeutic intervention. Phytochemicals may affect the genome and trigger damage to DNA and repair mechanisms [31]. However, many of such dietary substances have been associated with a decreased risk of cancer, for example, breast cancer [32, 33].
Differences between the two types of CNSL were identified by the GC-MS analysis. The iCNSL contained a mixture of anacardic acids that constituted 94.5% of its composition. Conversely, tCNSL exhibited a mixture of cardanols that constituted more than 90% of its composition (Table 1). These percentages corroborate the studies by Philip et al. [34], who reported that anacardic acid may exceed 80% of iCNSL composition. This percentage may be as high as 90%, according to Das and Ganesh [35]. Previous studies had found that cardanol is the main component of tCNSL [24, 36]. The difference in chemical composition between the two types of CNSL might result from the preparation process used to obtain tCNSL, which involves heating the cashew nuts to 180–200°C. This heat causes anacardic acid to undergo decarboxylation and conversion into cardanol, leading to the higher cardanol concentration in tCNSL [37].
The chemical compositions of both iCNSL and tCNSL recorded in the present study differ from the chemical composition reported in the literature [15, 23, 24]. The origin of the cashew nuts, the weather conditions, and, particularly, the extraction process used may account for the differences between this and other studies with regard to the ratio of phenolic compounds found in iCNSL [35], and the operating and heating conditions may affect the ratios recorded in tCNSL [38].
The A. salina bioassay is considered to be useful for preliminary assessments of general toxicity, and it correlates well with cytotoxic activity against some types of solid tumours in humans [26]. The IC50 of iCNSL in A. salina was lower than that of tCNSL (Figure 3). Plant extracts with LC50 values under 1,000 μg·mL−1 are considered to be active and to have toxic activity [39, 40]. Therefore, although iCNSL and tCNSL had different LC50 values, both are considered to have shown toxic effects in the acute toxicity test with A. salina. Guerra [41] suggests that the Artemia bioassay could be used for the toxicity evaluations of compounds that are rich in phenols.
The most significant toxicity results found in this study (Figure 3) were obtained when A. salina was treated with iCNSL. This finding may be related to the presence of anacardic acids in iCNSL because anacardic acids have been reported to be cytotoxic [22]. Other studies have reported that anacardic acid derivatives, such as isonicotinoyl hydrazone, show in vitro activity against Mycobacterium smegmatis [42].
Muroi et al. [43] showed that different types of unsaturations present in the chain of anacardic acids are related to increased antibacterial activity against Staphylococcus aureus. The synergistic effects decrease with an increasing number of double bonds in the chain. A possible explanation for this effect is that the introduction of unsaturation or branching into the hydrophobic groups increases the surfactant water solubility and, therefore, increases the activity [44]. The high toxicity of iCNSL at the highest test concentration in this bioassay was most likely due to the presence of anacardic acid and to its additive and/or synergistic effects.
Cardanol may account for the toxic activity of tCNSL in the A. salina bioassay (Figure 3). Studies of the wastewater from cashew-processing factories show that cardanol, which is the main component of tCNSL, is toxic to A. salina [45]. The iCNSL contains primarily anacardic acids, and tCNSL contains primarily cardanols. This difference confounds conclusions regarding whether anacardic acids or cardanol was the active agent responsible for the effects observed in this study.
Toxicity was also characterised in this study by a reduction in the growth of A. cepa root meristems. The highest concentration of iCNSL (69.50 μg·mL−1) caused significant root growth inhibition (P < 0.001) compared with the negative control. However, toxic activity was not observed for any of the three concentrations of tCNSL (Figure 3).
The iCNSL concentration of 69.50 μg·mL−1 was more than double the IC50 that was observed in A. salina (36.96 μg·mL−1), but the two lower concentrations were not toxic to A. cepa root meristems (Figure 3). Another study, in which three species of molluscs of the same genus (Biomphalaria straminea, B. tenagophila, and B. glabrata) were treated with a 20 ppm hexane extract, showed mortality rates that ranged from 97.1% to 100% after 24 hours of exposure [46].
The A. cepa test system is a key in vivo model for the evaluation of root growth after direct treatment with a substance of interest and for the prediction of DNA damage. The test is considered to be an effective preselection tool for toxicity and genotoxicity studies [47] because the results can be extrapolated to other animals and plants [47].
The A. cepa test also provides other macroscopic parameters that indicate the toxicity of chemicals and environmental pollutants. These toxicity parameters include very large roots, which indicate cellular proliferation; colour changes; and the presence of tumours [48]. This test reveals toxic [26] and cytotoxic effects [49].
The effects of extracts against damage caused by toxic agents have been analysed in recent studies assessing root size via the A. cepa test [28, 50]. Certain metals, including copper, may inhibit root growth, most likely by inhibiting cell division, and may also cause toxicity and cytotoxicity [51].
In the present study, only the highest test concentration of tCNSL showed preventive effects when exposed to the CuSO4·5H2O solution in A. cepa meristems. However, neither iCNSL nor tCNSL prevented the toxicity induced by CuSO4·H2O in co- or posttreatment applications (Table 3).
Root growth is regulated by the combination of cell division activity in mitotically active meristems and cell elongation in the regions that are proximal to root apices [52]. Only the highest concentration of iCNSL had significant antiproliferative activity (P < 0.05) compared with the negative control (Figure 5). This finding suggests that this concentration caused disturbance in meristem proliferation in A. cepa.
Macroscopic parameters are associated with toxicity and may likewise be associated with a reduction in the MI, which would affect DNA replication and protein synthesis [53]. No preventive, modulating, and reparative activities of CNSL against the cytotoxicity induced by copper sulphate were observed in this study (Table 4).
Oliveira et al. [23] also found that anacardic acid is a larger component of iCNSL than of tCNSL (Table 1) and that anacardic acid may have had prooxidant effects in A. cepa meristems. Recent studies indicate that antiproliferative effects on mammalian cell cultures are associated with oxidative stress [54] because the production of reactive oxygen species (ROS) impacts root growth and may inhibit growth and cell division [55].
The tCNSL showed no cytotoxic effect (P > 0.05) at any concentration. A positive correlation was found between inhibited root growth and reduced MI at the highest iCNSL concentration (Figures 4 and 5).
The MI is calculated by dividing the number of dividing cells by the total number of cells observed and is expressed as a percentage [56]. A reduction in the MI can be interpreted as cell death [57]. The present study showed a mitodepressive effect of iCNSL at the highest test concentration (69.50 μg·mL−1) on A. cepa cell division. The mitodepressive effect may have resulted from abnormal cellular conditions caused by the treatment. The reduction in the MI may have been related to early prophase arrest [58], inhibition of DNA synthesis, or cell cycle arrest at the G2 phase, which would prevent cells from entering mitosis [59]. The reduced MI also inhibits microtubule formation and nucleoprotein synthesis and reduces the ATP levels that provide energy for spindle elongation, microtubule dynamics, and chromosome movement [60].
Kubo et al. [61] also showed that anacardic acid and cardol may have a moderate cytotoxic effect. The inhibition of prooxidant enzymes may account for this effect. The volume of the hydrophobic side chain and its ability to act as a surfactant would explain its cytotoxic effect. Cardol has also been shown to be cytotoxic at a dose of 0.01 mM in HeLa cells [62]. The results of these two studies might confirm the cytotoxic action of the highest concentration of iCNSL which contains anacardic acid and cardol [18].
Acevedo et al. [22] showed that the anacardic acid present in Amphipterygium adstringens has cytotoxic effects in the peripheral lymphocytes of mice treated with doses of less than 10 mg/kg. The cytotoxic effects were evidenced by decreases in polychromatic and normochromatic erythrocytes. Anacardic acid from A. adstringens is also cytotoxic against Gram-positive bacteria in dental abscesses, has molluscicidal activity [63], inhibits apoptosis in chick embryonic neuronal cells [64], and inhibits breast cancer (MCF-7 and MDA-MB 231) cervical cancer cell lines and other types of tissues, including lung, liver, bladder, and melanoma [65].
Recent studies have also shown that a combination of anacardic acid and lunasin, which is another natural plant extract, may exhibit anticarcinogenic properties. These compounds act on the regulation of the expression of several genes involved in the cell cycle, apoptosis, and signal transduction [66]. Both compounds have a strong inhibitory effect on a number of cancer cell lines [67–69]. For example, [70] reported inhibition of the growth of HepG2 and U266 tumour cells treated with 60 µM of anacardic acid for 24, 48, and 72 hours.
The frequencies of CA and MN are commonly used to detect genotoxicity [59, 71]. This study investigated genotoxicity based on the frequency of MN and CAs, such as chromosome bridges, vagrant and laggard chromosomes, and chromosome fragments (Figure 7).
Iarmarcovai et al. [72] characterise micronuclei as small, spherical bodies that consist of genetic material that is not incorporated into the main nucleus during the final stages of mitosis. MN may result from the failure of acentric chromosome fragments to incorporate into the cell nucleus and clastogenicity (DNA breaks) or from whole chromosomes of aneugenic origin (disturbance in the mitotic spindle). The iCNSL at a concentration of 69.50 µg·mL−1 is thought to have induced genotoxic effects by means of clastogenic mechanisms (Table 5) because the MN that were generated at this concentration are considered small. Small MN are indicative of clastogenic action [73] resulting from genotoxic stress [74, 75].
The mutagenic response that occurred at the highest iCNSL concentration might have resulted from chromosomal instability, phenotypes, and cellular changes caused by genetic defects and/or exogenous exposure [76]. However, previous studies found that iCNSL did not have a mutagenic effect in the Ames test [22] or the MN test in mice bone marrow [77].
We, therefore, hypothesise that the MN formed at the highest iCNSL concentration resulted from breaks that occurred during cell division, possibly due to unrepaired or incorrectly repaired damage or to poor chromosome separation as result of mitotic malfunction. These events may have resulted from oxidative stress [78] and therefore from an integrated response to instability of the genetic material [79] that reflected various chromosomal changes [71].
However, iCNSL and tCNSL had no significant genotoxic effects on the frequency of CAs compared with the negative control (Figure 7). The iCNSL at the highest test concentration showed no preventive, antimutagenic, and reparative responses against CuSO4·5H2O. The iCNSL at the two lower concentrations and tCNSL did show preventive, antimutagenic, and reparative activities, as indicated by the reduced frequency of MN (Table 5). These results are consistent with the decrease in the frequency of CAs that resulted from the inhibition of damage induced by CuSO4·5H2O (Table 5).
Several experimental models have shown that synthetic or natural resorcinolic lipids do not cause DNA damage at low concentrations, which suggests that they have anticancer activity [80]. The results of these studies are consistent with the results of the present study, in which no genotoxicity was found at the lowest concentrations of iCNSL and tCNSL.
The present study documented the genotoxic effects of CuSO4·5H2O, which are explained by the ability of it causing DNA damage [29]. The chemical components of iCNSL and tCNSL may have protected, modulated, and repaired the oxidative effects of CuSO4·5H2O in A. cepa meristems. Cardol and cardanol were found to exhibit in vitro antioxidant effects in studies of the chemical characteristics of CNSL. These compounds have these effects because they scavenge free radicals, including the hydroxyl radical [23].
Other studies have also reported that the genotoxicity of chemical agents may be repaired by phenolic compounds with antioxidant and radical-scavenging activities [81]. Chromosomal aberrations consist of changes in chromosome structure that result in breaks or exchange of chromosomal material. These types of damage are usually lethal to cells, but some are viable and may have somatic or hereditary genetic effects [82].
Chromosomal fragments in cells indicate chromosomal breaks and may be related to anaphase bridges [83], disturbances in microtubule assembly, and cell death [84]. The results of the present study (Table 5) show that iCNSL and tCNSL failed to induce the fragment type of CA and therefore did not cause anaphase bridges when compared to the control group.
Antimutagenic compounds are able to induce some metabolic enzymes that may act as enzymatic inhibitors of mutagenic agents or inhibitors of promutagens in pretreatment experiments [85, 86]. The preventive, antimutagenic, and reparative effects of iCNSL and tCNSL observed in A. cepa (Tables 4 and 5) (except at the highest iCNSL concentration) were corroborated with the study of [15], who reported that tCNSL effect protected against oxidative stress (at a concentration of 100–500 μg·mL−1) in S. cerevisiae that were defective in antioxidant enzymes. The protection against damage caused by H2O2 occurs via bioantimutagenic mechanisms, but it occurs by means of dysmutagenesis in concurrent treatment. Bioantimutagenic agents act on the physiological mechanisms of DNA protection and repair and reverse the mutagenic effects and prevent their persistence [85, 87]. Thus, CNSL most likely acted as a bioantimutagenic and dysmutagenic agent and showed a stronger antimutagenic effect.
Components of phenolic lipids, including anacardic acid and alkylresorcinol, have antigenotoxic activity in vitro because of the ability of lipids to interact with biological membranes [88]. This is confirmed by the presence of hydrophilic and hydrophobic regions in their structures, which give lipids an amphipathic character that is responsible for their affinity for biological membranes. This character allows the phenolic lipids to be incorporated easily into cell membranes [89].
Cardanol also has antioxidant effects [23], and phenolic compounds with this ability can suppress genotoxicity [81]. Deszcz and Kozubek [90] noted that alkylresorcinols may be characterised as antioxidants when they are at very low concentrations, and they protect free fatty acids and phospholipids against peroxidation induced by the iron and autooxidation of biological membranes. These activities may constitute the main factor accounting for the antimutagenic activity exhibited by these compounds.
The present study confirms the strong antimutagenic, preventive, and restorative effects of CNSL. De Lima et al. [25] observed that iCNSL (at a concentration of 200 μg·mL−1) had an antioxidant effect in S. cerevisiae. Andrade et al. [15] observed that 100 μg·mL−1 of tCNSL might reduce free radical levels by 88.9% in the DPPH test and that it scavenges hydroxyl radicals by means of xanthine oxidase, resulting in antioxidant activity with an IC50 = 702 µg/mL.
Melo Cavalcante et al. [91] also confirmed that A. occidentale pulp has antioxidant effects against H2O2 at pre-, co-, and posttreatment in Salmonella typhimurium, as assessed by the Ames test. The authors attributed these effects to the pulp's chemical components, which include anacardic acid. These components may also protect S. typhimurium (TA102) against the damage induced by aflatoxin B1 via several mechanisms [92]. Cashew juice and cajuina (processed juice) reduce damage to the peripheral blood cells of mice. The juice caused a 60.82% reduction in damage and the cajuina caused an 82.19% reduction in damage, compared with cyclophosphamide. Further, the juice and the cajuina reduced the number of CAs in the bone marrow of mice by 53% and 65%, respectively. These effects may be related to the antioxidant activities of their components [13]. The results reported by de Carvalho Melo-Cavalcante et al. [13] confirm the (concentration-dependent) antimutagenic and antigenotoxic effects of iCNSL and tCNSL observed in the present study.
5. Conclusions
In summary, this study showed that anacardic acids are the primary components of iCNSL and cardanol of tCNSL. Both iCNSL and tCNSL showed protective (pretreatment), modulating (cotreatment), and reparative (posttreatment) in vivo effects against the damage induced by copper sulphate in A. cepa meristems at the lowest concentrations evaluated. Therefore, CNSL, which is a natural and renewable product extracted from the cashew nut shell, can be the basis for further studies to determine the mechanisms activated by its components and the mechanisms by which their synergism produces beneficial effects. These studies might be precursors for the production of biotechnological products.
Acknowledgments
The authors thank the National Council of Technological and Scientific Development (CNPq/Brazil) and the Research Laboratory in Genetic Toxicological, UFPI, Brazil.
Conflict of Interests
The authors declare no conflict of interests.
References
- 1.Dahech I., Farah W., Trigui M., et al. Antioxidant and antimicrobial activities of Lycium shawii fruits extract. International Journal of Biological Macromolecules. 2013;60:328–333. doi: 10.1016/j.ijbiomac.2013.05.020. [DOI] [PubMed] [Google Scholar]
- 2.Hasnat M. A., Pervin M., Lim B. O. Acetylcholinesterase inhibition and in vitro and in vivo antioxidant activities of Ganoderma lucidum grown on germinated brown rice. Molecules. 2013;18(6):6663–6678. doi: 10.3390/molecules18066663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Büchner F. L., Bueno-de-Mesquita H. B., Linseisen J., et al. Fruits and vegetables consumption and the risk of histological subtypes of lung cancer in the European prospective investigation into cancer and nutrition (EPIC) Cancer Causes and Control. 2010;21(3):357–371. doi: 10.1007/s10552-009-9468-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Soerjomataram I., Oomen D., Lemmens V., et al. Increased consumption of fruit and vegetables and future cancer incidence in selected European countries. European Journal of Cancer. 2010;46(14):2563–2580. doi: 10.1016/j.ejca.2010.07.026. [DOI] [PubMed] [Google Scholar]
- 5.Akaneme F. I., Amaefule C. C. Evaluation of the cytotoxicity and genotoxicity of aqueous leaf extracts of Azadirachta indica A. Juss using the Allium test . Journal of Medicinal Plants Research. 2012;6(23):3898–3907. [Google Scholar]
- 6.Munari C. C., de Oliveira P. F., de Souza Lima I. M., et al. Evaluation of cytotoxic, genotoxic and antigenotoxic potential of Solanum lycocarpum fruits glicoalkaloid extract in V79 cells. Food and Chemical Toxicology. 2012;50(10):3696–3701. doi: 10.1016/j.fct.2012.07.028. [DOI] [PubMed] [Google Scholar]
- 7.Rietjens I. M. C. M., Martena M. J., Boersma M. G., Spiegelenberg W., Alink G. M. Molecular mechanisms of toxicity of important food-borne phytotoxins. Molecular Nutrition & Food Research. 2005;49(2):131–158. doi: 10.1002/mnfr.200400078. [DOI] [PubMed] [Google Scholar]
- 8.Mazzetto S. E., Lomonaco D., Mele G. Cashew nut oil: opportunities and challenges in the context of sustainable industrial development. Quimica Nova. 2009;32(3):732–741. doi: 10.1590/s0100-40422009000300017. [DOI] [Google Scholar]
- 9.Moo-Huchin V. M., Moo-Huchin M. I., Estrada-Leon R. J., et al. Antioxidant compounds, antioxidant activity and phenolic content in peel from three tropical fruits from Yucatan, Mexico. Food Chemistry. 2015;166:17–22. doi: 10.1016/j.foodchem.2014.05.127. [DOI] [PubMed] [Google Scholar]
- 10.Zielinski A. A. F., Ávila S., Ito V., Nogueira A., Wosiacki G., Haminiuk C. W. I. The association between chromaticity, phenolics, carotenoids, and in vitro antioxidant activity of frozen fruit pulp in Brazil: an application of chemometrics. Journal of Food Science. 2014;79(4):C510–C516. doi: 10.1111/1750-3841.12389. [DOI] [PubMed] [Google Scholar]
- 11.Bae-Harboe Y.-S. C., Masterpol K. S. Botanical briefs: cashew apple (Anacardium occidentale) Cutis. 2013;92(4):174–176. [PubMed] [Google Scholar]
- 12.Guissoni A. C. P., Silva I. G., Geris R., Cunha L. C., Silva H. H. G. Larvicidal activity of Anacardium occidentale as an alternative to control Aedes aegypti and its toxicity in Rattus norvegicus . Revista Brasileira de Plantas Medicinais. 2013;15(3):363–367. doi: 10.1590/s1516-05722013000300008. [DOI] [Google Scholar]
- 13.de Carvalho Melo-Cavalcante A. A., de Moura Dantas S. M. M., de Sousa Leite A., et al. In vivo antigenotoxic and anticlastogenic effects of fresh and processed cashew (Anacardium occidentale) apple juices. Journal of Medicinal Food. 2011;14(7-8):792–798. doi: 10.1089/jmf.2010.0153. [DOI] [PubMed] [Google Scholar]
- 14.Ayepola O., Ishola R. Evaluation of antimicrobial activity of Anacardium occidentale (Linn.) Advances in Medical and Dental Sciences. 2009;3(1):1–3. [Google Scholar]
- 15.Andrade T. D. A. D., Araújo B. Q., Citó A. M. D. L., et al. Antioxidant properties and chemical composition of technical Cashew Nut Shell Liquid (tCNSL) Food Chemistry. 2011;126(3):1044–1048. doi: 10.1016/j.foodchem.2010.11.122. [DOI] [Google Scholar]
- 16.Michodjehoun-Mestres L., Souquet J.-M., Fulcrand H., Bouchut C., Reynes M., Brillouet J.-M. Monomeric phenols of cashew apple (Anacardium occidentale L.) Food Chemistry. 2009;112(4):851–857. doi: 10.1016/j.foodchem.2008.06.056. [DOI] [Google Scholar]
- 17.Kannan V. R., Sumathi C., Balasubramanian V., Ramesh N. Elementary chemical profiling and antifungal properties of cashew (Anacardium occidentale L.) Nuts. Botany Research International. 2009;2(4):253–257. [Google Scholar]
- 18.Parasa L. S., Tumati S. R., Kumar C., Chicurupati S., Rao G. In vitro antimicrobial activity of cashew (Anacardium occidentale, L.) nuts shell liquid against methicillin resistant Staphylococcus aureus (MRSA) clinical isolates. International Journal of Pharmacy Science. 2011;3:436–440. [Google Scholar]
- 19.Mukhopadhyay A. K., Hati A. K., Tamizharasu W., Sathya Babu P. Larvicidal properties of cashew nut shell liquid (Anacardium occidentale L) on immature stages of two mosquito species. Journal of Vector Borne Diseases. 2010;47(4):257–260. [PubMed] [Google Scholar]
- 20.George J., Kuttan R. Mutagenic, carcinogenic and cocarcinogenic activity of cashewnut shell liquid. Cancer Letters. 1997;112(1):11–16. doi: 10.1016/s0304-3835(96)04540-5. [DOI] [PubMed] [Google Scholar]
- 21.Polasa K., Rukmini C. Mutagenicity tests of cashewnut shell liquid, rice-bran oil and other vegetable oils using the Salmonella typhimurium/microsome system. Food and Chemical Toxicology. 1987;25(10):763–766. doi: 10.1016/0278-6915(87)90231-6. [DOI] [PubMed] [Google Scholar]
- 22.Acevedo H. R., Rojas M. D., Arceo S. D. B., et al. Effect of 6-nonadecyl salicylic acid and its methyl ester on the induction of micronuclei in polychromatic erythrocytes in mouse peripheral blood. Mutation Research—Genetic Toxicology and Environmental Mutagenesis. 2006;609(1):43–46. doi: 10.1016/j.mrgentox.2006.06.002. [DOI] [PubMed] [Google Scholar]
- 23.Oliveira M. S. C., Morais S. M. D., Magalhães D. V., et al. Antioxidant, larvicidal and antiacetylcholinesterase activities of cashew nut shell liquid constituents. Acta Tropica. 2011;117(3):165–170. doi: 10.1016/j.actatropica.2010.08.003. [DOI] [PubMed] [Google Scholar]
- 24.Kumar P. P., Paramashivappa R., Vithayathil P. J., Rao P. V. S., Rao A. S. Process for isolation of cardanol from technical cashew (Anacardium occidentale l.) Nut shell liquid. Journal of Agricultural and Food Chemistry. 2002;50(16):4705–4708. doi: 10.1021/jf020224w. [DOI] [PubMed] [Google Scholar]
- 25.De Lima S. G., Feitosa C. M., Citó A. M., et al. Effects of immature cashew nut-shell liquid (Anacardium occidentale) against oxidative damage in Saccharomyces cerevisiae and inhibition of acetylcholinesterase activity. Genetics and Molecular Research. 2008;7(3):806–818. doi: 10.4238/vol7-3gmr473. [DOI] [PubMed] [Google Scholar]
- 26.Meyer B. N., Ferrigni N. R., Putnam J. E. Brine shrimp: a convenient general bioassay for active plant constituents. Planta Medica. 1982;45(1):31–34. doi: 10.1055/s-2007-971236. [DOI] [PubMed] [Google Scholar]
- 27.Fiskesjö G. The Allium test as a standard in environmental monitoring. Hereditas. 1985;102(1):99–112. doi: 10.1111/j.1601-5223.1985.tb00471.x. [DOI] [PubMed] [Google Scholar]
- 28.Malini M., Marin-Morales M. A., Mantovani M. S., et al. Determination of the antimutagenicity of an aqueous extract of Rhizophora mangle L. (Rhizophoraceae), using in vivo and in vitro test systems. Genetics and Molecular Biology. 2010;33(1):176–181. doi: 10.1590/s1415-47572009005000106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Banu B. S., Ishaq M., Danadevi K., Padmavathi P., Ahuja Y. R. DNA damage in leukocytes of mice treated with copper sulfate. Food and Chemical Toxicology. 2004;42(12):1931–1936. doi: 10.1016/j.fct.2004.07.007. [DOI] [PubMed] [Google Scholar]
- 30.Yildiz M., Cğerci I. H., Konuk M., Fatih Fidan A., Terzi H. Determination of genotoxic effects of copper sulphate and cobalt chloride in Allium cepa root cells by chromosome aberration and comet assays. Chemosphere. 2009;75(7):934–938. doi: 10.1016/j.chemosphere.2009.01.023. [DOI] [PubMed] [Google Scholar]
- 31.Rajendran P., Ho E., Williams D. E., Dashwood R. H. Dietary phytochemicals, HDAC inhibition, and DNA damage/repair defects in cancer cells. Clinical Epigenetics. 2011;3, article 4 doi: 10.1186/1868-7083-3-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kaefer C. M., Milner J. A. The role of herbs and spices in cancer prevention. The Journal of Nutritional Biochemistry. 2008;19(6):347–361. doi: 10.1016/j.jnutbio.2007.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ramos S. Cancer chemoprevention and chemotherapy: dietary polyphenols and signalling pathways. Molecular Nutrition & Food Research. 2008;52(5):507–526. doi: 10.1002/mnfr.200700326. [DOI] [PubMed] [Google Scholar]
- 34.Philip J. Y. N., Buchweishaija J., Mkayula L. L., Ye L. Preparation of molecularly imprinted polymers using anacardic acid monomers derived from cashew nut shell liquid. Journal of Agricultural and Food Chemistry. 2007;55(22):8870–8876. doi: 10.1021/jf0718289. [DOI] [PubMed] [Google Scholar]
- 35.Das P., Ganesh A. Bio-oil from pyrolysis of cashew nut shell—a near fuel. Biomass and Bioenergy. 2003;25(1):113–117. doi: 10.1016/s0961-9534(02)00182-4. [DOI] [Google Scholar]
- 36.Ikeda R., Tanaka H., Uyama H., Kobayashi S. Synthesis and curing behaviors of a crosslinkable polymer from cashew nut shell liquid. Polymer. 2002;43(12):3475–3481. doi: 10.1016/S0032-3861(02)00062-9. [DOI] [Google Scholar]
- 37.Attanasi O. A., Berretta S., Fiani C., Filippone P., Mele G., Saladino R. Synthesis and reactions of nitro derivatives of hydrogenated cardanol. Tetrahedron. 2006;62(25):6113–6120. doi: 10.1016/j.tet.2006.03.105. [DOI] [Google Scholar]
- 38.Lubi M. C., Thachil E. T. Cashew nut shell liquid (CNSL)—a versatile monomer for polymer synthesis. Designed Monomers and Polymers. 2000;3(2):123–153. doi: 10.1163/156855500300142834. [DOI] [Google Scholar]
- 39.dos Santos H. M., Jr., Oliveira D. F., De Carvalho D. A., et al. Evaluation of native and exotic Brazilian plants for anticancer activity. Journal of Natural Medicines. 2010;64(2):231–238. doi: 10.1007/s11418-010-0390-0. [DOI] [PubMed] [Google Scholar]
- 40.Arcanjo D. D. R., Albuquerque A. C. M., Melo-Neto B., Santana L. C. L. R., Medeiros M. G. F., Citó A. M. G. L. Bioactivity evaluation against Artemia salina Leach of medicinal plants used in Brazilian Northeastern folk medicine. Brazilian Journal of Biology. 2012;72(3):505–509. doi: 10.1590/s1519-69842012000300013. [DOI] [PubMed] [Google Scholar]
- 41.Guerra R. Ecotoxicological and chemical evaluation of phenolic compounds in industrial effluents. Chemosphere. 2001;44(8):1737–1747. doi: 10.1016/s0045-6535(00)00562-2. [DOI] [PubMed] [Google Scholar]
- 42.Swamy B. N., Suma T. K., Rao G. V., Reddy G. C. Synthesis of isonicotinoylhydrazones from anacardic acid and their in vitro activity against Mycobacterium smegmatis . European Journal of Medicinal Chemistry. 2007;42(3):420–424. doi: 10.1016/j.ejmech.2006.09.009. [DOI] [PubMed] [Google Scholar]
- 43.Muroi H., Nihei K.-I., Tsujimoto K., Kubo I. Synergistic effects of anacardic acids and methicillin against methicillin resistant Staphylococcus aureus . Bioorganic and Medicinal Chemistry. 2004;12(3):583–587. doi: 10.1016/j.bmc.2003.10.046. [DOI] [PubMed] [Google Scholar]
- 44.Rosen M. J. Surfactants and Interfacial Phenomena. Wiley; 1989. [Google Scholar]
- 45.Pimentel M. R., De Lima D. P., Martins L. R., Beatriz A., Santaella S. T., Lotufo L. V. C. Ecotoxicological analysis of cashew nut industry effluents, specifically two of its major phenolic components, cardol and cardanol. Pan-American Journal of Aquatic Sciences. 2009;4(3):363–368. [Google Scholar]
- 46.de Souza C. P., Mendes N. M., Jannotti-Passos L. K., Pereira J. P. The use of cashew nut shell of caju (Anacardium occidentale) as alternative molluscicide. Revista do Instituto de Medicina Tropical de São Paulo. 1992;34(5):459–466. [PubMed] [Google Scholar]
- 47.Bagatini M. D., Vasconcelos T. G., Laughinghouse H. D., IV, Martins A. F., Tedesco S. B. Biomonitoring hospital effluents by the Allium cepa L. test. Bulletin of Environmental Contamination and Toxicology. 2009;82(5):590–592. doi: 10.1007/s00128-009-9666-z. [DOI] [PubMed] [Google Scholar]
- 48.Herrero O., Pérez Martín J. M., Fernández Freire P., Carvajal López L., Peropadre A., Hazen M. J. Toxicological evaluation of three contaminants of emerging concern by use of the Allium cepa test. Mutation Research/Genetic Toxicology and Environmental Mutagenesis. 2012;743(1-2):20–24. doi: 10.1016/j.mrgentox.2011.12.028. [DOI] [PubMed] [Google Scholar]
- 49.Rossato L. V., Tedesco S. B., Laughinghouse H. D., IV, Farias J. G., Nicoloso F. T. Alterations in the mitotic index of Allium cepa induced by infusions of Pluchea sagittalis submitted to three different cultivation systems. Anais da Academia Brasileira de Ciências. 2010;82(4):857–860. doi: 10.1590/s0001-37652010000400007. [DOI] [PubMed] [Google Scholar]
- 50.Silva D. S. B. S., Barboza B., Garcia A. C. F. S., et al. Investigation of protective effects of Erythrina velutina extract against MMS induced damages in the root meristem cells of Allium cepa . Brazilian Journal of Pharmacognosy. 2013;23(2):273–278. doi: 10.1590/s0102-695x2013005000006. [DOI] [Google Scholar]
- 51.Dimitrova I., Ivanova E. Effect of heavy metal soil pollution on some morphological and cytogenetical characteristics of flax (Linum usitatissum L.) Journal of Balkan Ecology. 2003;4:212–218. [Google Scholar]
- 52.Shishkova S., Rost T. L., Dubrovsky J. G. Determinate root growth and meristem maintenance in angiosperms. Annals of Botany. 2008;101(3):319–340. doi: 10.1093/aob/mcm251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Tkalec M., Malarić K., Pavlica M., Pevalek-Kozlina B., Vidaković-Cifrek Ž. Effects of radiofrequency electromagnetic fields on seed germination and root meristematic cells of Allium cepa L. Mutation Research: Genetic Toxicology and Environmental Mutagenesis. 2009;672(2):76–81. doi: 10.1016/j.mrgentox.2008.09.022. [DOI] [PubMed] [Google Scholar]
- 54.Martín J. M. P., Peropadre A., Herrero Ó., Freire P. F., Labrador V., Hazen M. J. Oxidative DNA damage contributes to the toxic activity of propylparaben in mammalian cells. Mutation Research—Genetic Toxicology and Environmental Mutagenesis. 2010;702(1):86–91. doi: 10.1016/j.mrgentox.2010.07.012. [DOI] [PubMed] [Google Scholar]
- 55.Jones D. L., Blancaflor E. B., Kochian L. V., Gilroy S. Spatial coordination of aluminium uptake, production of reactive oxygen species, callose production and wall rigidification in maize roots. Plant, Cell & Environment. 2006;29(7):1309–1318. doi: 10.1111/j.1365-3040.2006.01509.x. [DOI] [PubMed] [Google Scholar]
- 56.Kumari M., Mukherjee A., Chandrasekaran N. Genotoxicity of silver nanoparticles in Allium cepa . Science of the Total Environment. 2009;407(19):5243–5246. doi: 10.1016/j.scitotenv.2009.06.024. [DOI] [PubMed] [Google Scholar]
- 57.Ping K. Y., Darah I., Yusuf U. K., Yeng C., Sasidharan S. Genotoxicity of Euphorbia hirta: an Allium cepa assay. Molecules. 2012;17(7):7782–7791. doi: 10.3390/molecules17077782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Briand C. H., Kapoor B. M. The cytogenetic effects of sodium salicylate on the root meristem cells of Alium sativum L. Cytologia. 1989;54(2):203–209. doi: 10.1508/cytologia.54.203. [DOI] [Google Scholar]
- 59.Türkoğlu Ş. Evaluation of genotoxic effects of sodium propionate, calcium propionate and potassium propionate on the root meristem cells of Allium cepa . Food and Chemical Toxicology. 2008;46(6):2035–2041. doi: 10.1016/j.fct.2008.01.043. [DOI] [PubMed] [Google Scholar]
- 60.Majewska A., Wolska E., Śliwińska E., et al. Antimitotic effect, G2/M accumulation, chromosomal and ultrastructure changes in meristematic cells of Allium cepa L. root tips treated with the extract from Rhodiola rosea roots. Caryologia. 2003;56(3):337–351. doi: 10.1080/00087114.2003.10589343. [DOI] [Google Scholar]
- 61.Kubo I., Nitoda T., Tocoli F. E., Green I. R. Multifunctional cytotoxic agents from Anacardium occidentale . Phytotherapy Research. 2011;25(1):38–45. doi: 10.1002/ptr.3109. [DOI] [PubMed] [Google Scholar]
- 62.Hemshekhar M., Sebastin Santhosh M., Kemparaju K., Girish K. S. Emerging roles of anacardic acid and its derivatives: a pharmacological overview. Basic and Clinical Pharmacology and Toxicology. 2012;110(2):122–132. doi: 10.1111/j.1742-7843.2011.00833.x. [DOI] [PubMed] [Google Scholar]
- 63.Kubo I., Komatsu S., Ochi M. Molluscicides from the cashew Anacardium occidentale and their large-scale isolation. Journal of Agricultural and Food Chemistry. 1986;34(6):970–973. doi: 10.1021/jf00072a010. [DOI] [Google Scholar]
- 64.Ahlemeyer B., Selke D., Schaper C., Klumpp S., Krieglstein J. Ginkgolic acids induce neuronal death and activate protein phosphatase type-2C. European Journal of Pharmacology. 2001;430(1):1–7. doi: 10.1016/S0014-2999(01)01237-7. [DOI] [PubMed] [Google Scholar]
- 65.Sung B., Pandey M. K., Ann K. S., et al. Anacardic acid (6-nonadecyl salicylic acid), an inhibitor of histone acetyltransferase, suppresses expression of nuclear factor-κB-regulated gene products involved in cell survival, proliferation, invasion, and inflammation through inhibition of the inhibitory subunit of nuclear factor-KBa kinase, leading to potentiation of apoptosis. Blood. 2008;111(10):4880–4891. doi: 10.1182/blood-2007-10-117994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Hsieh C. C., Hernández-Ledesma B., de Lumen B. O. Lunasin-aspirin combination against NIH/3T3 cells transformation induced by chemical carcinogens. Plant Foods for Human Nutrition. 2011;66(2):107–113. doi: 10.1007/s11130-011-0229-1. [DOI] [PubMed] [Google Scholar]
- 67.Seong Y.-A., Shin P.-G., Kim G.-D. Anacardic acid induces mitochondrial-mediated apoptosis in the A549 human lung adenocarcinoma cells. International Journal of Oncology. 2013;42(3):1045–1051. doi: 10.3892/ijo.2013.1763. [DOI] [PubMed] [Google Scholar]
- 68.Tan J., Chen B., He L., et al. Anacardic acid (6-pentadecylsalicylic acid) induces apoptosis of prostate cancer cells through inhibition of androgen receptor and activation of p53 signaling. Chinese Journal of Cancer Research. 2012;24(4):275–283. doi: 10.1007/s11670-012-0264-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Wu Y., He L., Zhang L., et al. Anacardic acid (6-pentadecylsalicylic acid) inhibits tumor angiogenesis by targeting Src/FAK/Rho GTpases signaling pathway. Journal of Pharmacology and Experimental Therapeutics. 2011;339(2):403–411. doi: 10.1124/jpet.111.181891. [DOI] [PubMed] [Google Scholar]
- 70.Huang H., Hua X., Liu N., et al. Anacardic acid induces cell apoptosis associated with induction of ATF4-dependent endoplasmic reticulum stress. Toxicology Letters. 2014;228(3):170–178. doi: 10.1016/j.toxlet.2014.05.012. [DOI] [PubMed] [Google Scholar]
- 71.Majer B. J., Grummt T., Uhl M., Knasmüller S. Use of plant bioassays for the detection of genotoxins in the aquatic environment. Acta Hydrochimica et Hydrobiologica. 2005;33(1):45–55. doi: 10.1002/aheh.200300557. [DOI] [Google Scholar]
- 72.Iarmarcovai G., Bonassi S., Botta A., Baan R. A., Orsière T. Genetic polymorphisms and micronucleus formation: a review of the literature. Mutation Research—Reviews in Mutation Research. 2008;658(3):215–233. doi: 10.1016/j.mrrev.2007.10.001. [DOI] [PubMed] [Google Scholar]
- 73.Leme D. M., Marin-Morales M. A. Allium cepa test in environmental monitoring: a review on its application. Mutation Research/Reviews in Mutation Research. 2009;682(1):71–81. doi: 10.1016/j.mrrev.2009.06.002. [DOI] [PubMed] [Google Scholar]
- 74.Decordier I., Papine A., Plas G., et al. Automated image analysis of cytokinesis-blocked micronuclei: an adapted protocol and a validated scoring procedure for biomonitoring. Mutagenesis. 2009;24(1):85–93. doi: 10.1093/mutage/gen057. [DOI] [PubMed] [Google Scholar]
- 75.Heddle J. A., Fenech M., Hayashi M., MacGregor J. T. Reflections on the development of micronucleus assays. Mutagenesis. 2011;26(1):3–10. doi: 10.1093/mutage/geq085. [DOI] [PubMed] [Google Scholar]
- 76.Fernandes T. C. C., Mazzeo D. E. C., Marin-Morales M. A. Mechanism of micronuclei formation in polyploidizated cells of Allium cepa exposed to trifluralin herbicide. Pesticide Biochemistry and Physiology. 2007;88(3):252–259. doi: 10.1016/j.pestbp.2006.12.003. [DOI] [Google Scholar]
- 77.Carvalho A. L. N., Annoni R., Silva P. R. P., et al. Acute, subacute toxicity and mutagenic effects of anacardic acids from cashew (Anacardium occidentale Linn.) in mice. Journal of Ethnopharmacology. 2011;135(3):730–736. doi: 10.1016/j.jep.2011.04.002. [DOI] [PubMed] [Google Scholar]
- 78.Fenech M. Cytokinesis-block micronucleus assay evolves into a “cytome” assay of chromosomal instability, mitotic dysfunction and cell death. Mutation Research—Fundamental and Molecular Mechanisms of Mutagenesis. 2006;600(1-2):58–66. doi: 10.1016/j.mrfmmm.2006.05.028. [DOI] [PubMed] [Google Scholar]
- 79.Migliore L., Coppedè F., Fenech M., Thomas P. Association of micronucleus frequency with neurodegenerative diseases. Mutagenesis. 2011;26(1):85–92. doi: 10.1093/mutage/geq067. [DOI] [PubMed] [Google Scholar]
- 80.Buonanno F., Quassinti L., Bramucci M., et al. The protozoan toxin climacostol inhibits growth and induces apoptosis of human tumor cell lines. Chemico-Biological Interactions. 2008;176(2-3):151–164. doi: 10.1016/j.cbi.2008.07.007. [DOI] [PubMed] [Google Scholar]
- 81.Romero-Jiménez M., Campos-Sánchez J., Analla M., Muñoz-Serrano A., Alonso-Moraga Á. Genotoxicity and anti-genotoxicity of some traditional medicinal herbs. Mutation Research: Genetic Toxicology and Environmental Mutagenesis. 2005;585(1-2):147–155. doi: 10.1016/j.mrgentox.2005.05.004. [DOI] [PubMed] [Google Scholar]
- 82.Swierenga S. H. H., Heddle J. A., Sigal E. A., et al. Recommended protocols based on a survey of current practice in genotoxicity testing laboratories, IV. Chromosome aberration and sister-chromatid exchange in Chinese hamster ovary, V79 Chinese hamster lung and human lymphocyte cultures. Mutation Research—Fundamental and Molecular Mechanisms of Mutagenesis. 1991;246(2):301–322. doi: 10.1016/0027-5107(91)90050-x. [DOI] [PubMed] [Google Scholar]
- 83.Singh R. J. Plant Cytogenetics. Boca Raton, Fla, USA: CRC Press; 2003. [Google Scholar]
- 84.Çelik T. A., Aslantürk Ö. S. Evaluation of cytotoxicity and genotoxicity of Inula viscosa leaf extracts with Allium test. Journal of Biomedicine and Biotechnology. 2010;2010:8. doi: 10.1155/2010/189252.189252 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Kada T., Inoue T., Namiki N. Environmental desmutagens and antimutagens. In: Klekowski E. J., editor. Environmental Mutagenesis and Plant Biology. New York, NY, USA: Praeger; 1982. pp. 137–151. [Google Scholar]
- 86.Kuroda Y., Jain A. K., Tezuka H., Kada T. Antimutagenicity in cultured mammalian cells. Mutation Research—Fundamental and Molecular Mechanisms of Mutagenesis. 1992;267(2):201–209. doi: 10.1016/0027-5107(92)90064-9. [DOI] [PubMed] [Google Scholar]
- 87.de Flora S. Mechanisms of inhibitors of mutagenesis and carcinogenesis. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis. 1998;402(1-2):151–158. doi: 10.1016/s0027-5107(97)00292-3. [DOI] [PubMed] [Google Scholar]
- 88.Parikka K., Rowland I. R., Welch R. W., Wähälä K. In vitro antioxidant activity and antigenotoxicity of 5-n-alkylresorcinols. Journal of Agricultural and Food Chemistry. 2006;54(5):1646–1650. doi: 10.1021/jf052486e. [DOI] [PubMed] [Google Scholar]
- 89.Kozubek A., Tyman J. H. P. Resorcinolic lipids, the natural non-isoprenoid phenolic amphiphiles and their biological activity. Chemical Reviews. 1999;99(1):1–26. doi: 10.1021/cr970464o. [DOI] [PubMed] [Google Scholar]
- 90.Deszcz L., Kozubek A. Higher cardol homologs (5-alkylresorcinols) in rye seedlings. Biochimica et Biophysica Acta. 2000;1483(2):241–250. doi: 10.1016/s1388-1981(99)00187-0. [DOI] [PubMed] [Google Scholar]
- 91.Melo Cavalcante A. A., Rubensam G., Picada J. N., Gomes da Silva E., Fonseca Moreira J. C., Henriques J. A. P. Mutagenicity, antioxidant potential, and antimutagenic activity against hydrogen peroxide of cashew (Anacardium occidentale) apple juice and cajuina. Environmental and Molecular Mutagenesis. 2003;41(5):360–369. doi: 10.1002/em.10158. [DOI] [PubMed] [Google Scholar]
- 92.Cavalcante A. A. M., Rübensam G., Erdtmann B., Brendel M., Henriques J. A. P. Cashew (Anacardium occidentale) apple juice lowers mutagenicity of aflatoxin B1 in S. typhimurium TA102. Genetics and Molecular Biology. 2005;28(2):328–333. doi: 10.1590/s1415-47572005000200025. [DOI] [Google Scholar]