

Abstract
Iatrogenic gestational exposure to diethylstilbestrol (DES) induced alterations of the genital tract and predisposed individuals to develop clear cell carcinoma of the vagina as well as breast cancer later in life. Gestational exposure of rodents to a related compound, the xenoestrogen bisphenol-A (BPA) increases the propensity to develop mammary cancer during adulthood, long after cessation of exposure. Exposure to BPA during gestation induces morphological alterations in both the stroma and the epithelium of the fetal mammary gland at 18 days of age. We postulate that the primary target of BPA is the fetal stroma, the only mammary tissue expressing estrogen receptors during fetal life. BPA would then alter the reciprocal stroma-epithelial interactions that mediate mammogenesis. In addition to this direct effect on the mammary gland, BPA is postulated to affect the hypothalamus and thus in turn affect the regulation of mammotropic hormones at puberty and beyond.
Keywords: fetal mammary gland, mammary gland development, xenoestrogens, endocrine disruptors, environmental exposure, prenatal exposure
1. Introduction
Over the past four decades, strong evidence indicates that fetal exposure to estrogens causes perturbations in mammary gland development leading to mammary cancer manifested during adulthood (1–4). Beside the effects reported from exposure to endogenous and pharmacological estrogens (DES), a new source of concern is fetal exposure to endocrine disrupting chemicals (EDCs), including estrogen mimics (xenoestrogens) (5–7). An EDC is an exogenous chemical, or a mixture of chemicals, that interferes with any aspect of hormone action (8). Among these EDCs, BPA has gained much attention because of its ubiquitous presence in the environment (9–12). The increased incidence of breast, uterine and testicular cancers observed in European populations in the past 50 years has been postulated to be due to EDC exposure during periods of increased vulnerability, such as fetal development and peri-pubertal stages (13–17). This hypothesis was tested by examining the effect of environmentally relevant doses of BPA during fetal life of experimental animals. Perinatal BPA exposure to mice and monkeys resulted in alterations in both the stroma and the epithelium of the developing mammary gland (18–20). Moreover, intraductal hyperplasias, ductal carcinoma in situ (DCIS) and palpable tumors in murine species were documented long after the end of exposure (21–26). What remains to be determined is whether BPA acts exclusively by directly affecting epithelial-stromal interactions in the developing mammary gland or, in addition, by interfering with the hypothalamus-pituitary-ovarian axis (HPOA). In this review, we discuss the effects of fetal exposure to environmentally relevant levels of BPA on fetal mammary gland development and examine the hypothesis that BPA, by virtue of its estrogenicity, increases the risk of developing breast cancer in adulthood by exerting its deleterious effects through the stroma of the developing mammary gland.
2. Fetal exposure to endogenous and pharmacological estrogens
Epidemiological studies suggest that changes in the endocrine-fetal milieu predispose women to diseases that are manifested during adulthood (4;27–29). For instance, non-identical twin birth is considered an indicator of high exposure to estrogen, while pre-eclampsia is regarded as an indicator of low exposure to estrogen. Dizygotic birth correlates with increased risk of breast cancer in the offspring while pre-eclampsia correlates with a lower risk (29–31).
Fetal exposure to pharmacological doses of estrogens increases breast cancer risk at the age of prevalence (28;30;31). Compelling evidence favoring this claim was gathered from the daughters of mothers who were treated with massive doses of DES between 1948 and 1971 in order to prevent spontaneous abortion (32–35). The “DES daughters”, as these women are now commonly referred to, have been diagnosed with a variety of anatomo-physiological abnormalities in the uterus, oviduct and cervix as well as with clear cell adenocarcinoma of the vagina. Recently published data suggest that the DES daughters aged 40 years and older have an increased breast cancer incidence (1;36).
3. Mode of action of xenoestrogens during fetal life
Endogenous estrogens regulate multiple aspects of reproduction and development in males and females. Estrogens bring about their action by binding to nuclear receptors, estrogen receptor alpha (37) and beta (38) as well as the membrane-bound receptor, GPR30 (39). In females, estrogens regulate the morphology and physiology of the reproductive tract at all stages of life, including puberty, pregnancy and menopause (40). In males the generation of null mutants of the estrogen receptor (ER) alpha gene revealed that these mice were infertile. Also in males, it was shown that germ cells do not require ER alpha for development and function, but that somatic cells of the reproductive tract require ER alpha to produce sperm that are capable of fertilization (41). During adulthood, endogenous estrogens mediate biological events in the reproductive tract that, for the most part, are reversible. However, fetal development of the female genital tract is thought not to require estrogens, and excess estrogens are known to produce irreversible alterations, such as those observed in DES daughters.
Xenoestrogens bind to estrogen receptors alpha and beta, both when residing in the nucleus and when attached to the plasma membrane. Additionally, xenoestrogens are known to bind to the membrane receptors GPR30 (42–44) and the orphan receptor ERR gamma (45). Low-dose effects of xenoestrogens may be due to their acting through membrane receptors and to additive action with endogenous estrogens (46).
4. BPA as a xenoestrogens
BPA is commonly used in the manufacturing of plastics and is present in polycarbonate products and epoxy resins. BPA exposure occurs when it leaches into containers holding food, beverages, water and milk, which are later consumed. Additionally, BPA is released from water pipes, dental materials, protective coatings, adhesives, protective window glazing, compact discs, thermal paper and paper coatings (9;47). Pervasive use of BPA increases the risk of exposure to both the developing fetus indirectly through maternal exposure, and the neonate directly through ingestion of infant formula or maternal milk (11;48).
BPA is present in the urine of 95% of a representative sample of a non-institutionalized U.S. population over 6 years of age (49), including pregnant women (50). BPA has also been detected in maternal and fetal serum and placental tissue of newborn humans. The range of BPA concentrations in fetal serum ranged from 0.2 to 9.2 ng/ml, indicating that the developing human fetus and neonate are readily exposed to this chemical (11). To date, BPA is the best-studied xenoestrogen for which effects of exposure have been reported at various time points ranging from fetal to postnatal development.
5. Role of the environment during development
The influence of the environment on phenotype determination has been known since the 1880’s from observations in wildlife species. For instance, August Weismann documented that the spring and summer morphs of a butterfly species could be regulated by temperature. However, the gene-centric view of development that dominated 20th century research overlooked these phenomena that revealed phenotypic plasticity (51). The theory proposing that development is the execution of a genetic program gained prevalence from studies in animal models including Drosophila and rodents generated under strictly regulated laboratory conditions. Under these artificial conditions, animals reproduce all year long under light, temperature and strict diet regulations, avoiding external environmental influences. As a consequence, explanations in embryology have been narrowed down to genetics and molecular pathways. However, the phenomenon called polyphenism – one genotype, many phenotypes – contradicts the theory that development is the execution of a genetic program (51). Lately, the role of the environment in phenotype determination is regaining central stage due to its relevance to disease. For example, epidemiological studies showed that malnutrition during fetal development increased the risk to coronary and metabolic diseases during adulthood. Thus the old phenomenon of developmental plasticity is coming back under a medical disguise, namely, the “fetal origins of adult disease” hypothesis (52).
6. Stromal-epithelial interactions in fetal mammary gland development
Besides being influenced by the external environment, embryonic development is mediated by interactions at various levels of biological organization. Epithelial cells interact with neighboring epithelial cells as well as with cells in the surrounding stroma and with the extracellular matrix (ECM). These interactions take place along several spatial and temporal scales that define the shape of the organism (53). The organism imposes constraints at the local and global level via biophysical and biochemical interactions through cell proliferation, cell motility and cell adhesion. Tissue development in appendages like the mammary gland, tooth, feather and hair occur as a result of constant reciprocal interactions between epithelial cells and the surrounding ECM (54;55). Pioneering work by Kratochwil showed that stromal-epithelial interactions are crucial for mammary gland morphogenesis (56). Using an explant model and tissue recombinantion techniques, he separated embryonic day (E) 12 to 16 mammary epithelium from the mesenchyme, and showed that the epithelium develops only after recombination with its mesenchyme. In addition, he showed that recombination with the salivary mesenchyme induces the originally mammary epithelium to display a dichotomous branching pattern typical of the salivary gland. Later, Sakakura et al confirmed these findings in vivo, by recombining E16 mammary epithelium with E14 salivary mesenchyme under the kidney capsule (57). These results clearly pointed out the inductive role of the stroma in mammary and salivary gland morphogenesis.
7. Breast carcinogenesis: a consequence of altered tissue interactions?
Cell-based theories of carcinogenesis, such as the somatic mutation theory (SMT), consider cancer as a disease of cell proliferation and postulate that it occurs as a result of DNA mutations in genes that control cell proliferation (58). Alternatively, the tissue organization field theory (TOFT) postulates that cancer, like morphogenesis is a matter of tissue organization, and it proposes cancer as development gone awry. In fact, pathologists diagnose cancer by looking at altered tissue organization. In the context of TOFT, carcinogens interfere with cell-cell communication and cell-matrix interactions, resulting in disruption at the tissue-level that gives rise to neoplasia (59). From fetal life throughout the life time of the individual, mammary gland morphogenesis involves reciprocal interactions between the stroma and the epithelium (56;57;60;61).
A theory-neutral experimental strategy was used to determine which tissue was the target of a carcinogen. Rats from susceptible strains developed mammary gland tumors when exposed to the chemical carcinogen nitrosomethylurea (NMU). Separately, mammary gland epithelium and mammary gland stroma were exposed to NMU or vehicle and then recombined. The recombination of stroma exposed to NMU with vehicle-exposed epithelial cells resulted in neoplasms (62). The reverse combination did not. This observation showed that the stroma, rather than individual cells in the epithelium, is the target of the carcinogen. These results also pointed to the contextuality of the neoplastic phenotype. In a complementary experimental approach, the epithelial neoplastic phenotype was reversed when epithelial cells isolated from a mammary gland carcinoma were transplanted into a “normal” unexposed stroma (63). From these results, we concluded that tissues are the targets of carcinogens and that TOFT most reliably explains carcinogenesis. Comparable evidence has been gathered for other types of neoplasias such as metastatic breast cancer (64), metastatic melanoma (65) and hepatocellular carcinoma (66) and altogether provide further support for the central role of stromal-epithelial interactions in neoplastic development. Extrapolating from this compelling experimental and clinical evidence, we hypothesized that, BPA exerts its carcinogenic effects by binding to the ERs present in the stroma of the fetal mammary gland.
8. Bisphenol-A: a disruptor of stromal-epithelial interactions?
Fetal exposure to BPA results in morphological alterations in both the stroma and the epithelium of the developing mammary gland (18;19) which can lead to neoplasia later in adulthood (24;26). During fetal development, estrogen receptors α and β are exclusively detected in the stroma of the mammary gland (20;67). The transcriptomal changes induced by BPA and ethinyl estradiol are dissimilar in the stroma from those in the epithelium of fetal exposed mammary glands. This apparently counterintuitive result is easily explained given that different estrogens directly induce slightly different patterns of gene expression in the target ER positive cells (68). The gene expression pattern in the epithelium is more similar between BPA and ethinyl estradiol treatments. This is due to the fact that only a fraction of genes induced in the stroma play a role in stromal-epithelial interactions; and those are the ones that are involved in carrying estrogenic signals to the epithelium. Thus the effects observed in the epithelium must be mediated by the stroma (Figure 1) (20). Altogether this buttresses the hypothesis that BPA exerts its deleterious effects during fetal mammary gland development through the stroma.
Figure 1.

Hierarchical Clustering analyses of the transcriptomal profiles show the effects of perinatal exposure to BPA, ethinyl estradiol (EE2) and vehicle on peri-ductal stroma (left panel) and epithelium (right panel) of WT and null ERα mice. Status of mouse genotype is indicated on the left (Green: WT (+/+), Black: ER knock-out (−/−)) and treatment is indicated on the right of the heat maps. Unique gene clusters observed are shown on the top of the heat maps. Originally published in Wadia et al PLOS ONE 2013 (20).
9. Fetal exposure to BPA results in altered mammary gland morphogenesis
Environmentally relevant levels of BPA altered the developing mammary gland in mice exposed in utero from gestational day (gd) 8 to 18. At gd18, the mammary glands were collected and examined to identify alterations in the epithelium and in the stroma (18;20). The stroma of BPA exposed mammary glands displayed alterations in the ECM organization as well as in its cellular components. For instance, collagen deposition was higher near the epithelium while it decreased further away in the loose connective tissue. The gene expression of biomechanical modulators of the ECM, versican and tenascin C, was down-regulated in the periductal stroma of BPA exposed mammary glands (20). Regarding cells in the stroma, BPA increased the number of adipocytes containing lipid droplets and accelerated adipocyte differentiation. BPA also increased apoptosis in the presumptive fat pad as shown by Bax staining (18). These histological changes observed in the presumptive fat pad correlated with up-regulation of adipogenesis related genes (20). The mammary epithelium was overall more developed in BPA-exposed mammary glands compared to controls, as indicated by an increased area subtended by the ducts and ductal extension. BPA delayed ductal lumen formation, indicating that BPA had not produced a mere acceleration in development but its disruption (18). Additionally, fetal mammary glands of in utero BPA-exposed non-human primates showed a higher density of mammary buds and a more advanced epithelial development than the control group (Figure 2) (19).
Figure 2.

Whole mounts of neonatal mammary glands of control and BPA dosed non-human primates. (A) Control; (B) BPA dosed. Scale bar is equivalent to 500 µm. Originally published in Tharp et al., Proc. Natl. Acad. Sci USA 2012 (19).
10. Alterations in the postnatal mammary gland due to in utero exposure to BPA
The mammary glands of mice fetally-exposed to BPA showed increased lateral branching (Figure 3) and increased numbers of epithelial cells expressing PR at 4 months of age (69). At 6 months of age, these mice showed clear changes in their mammary gland histoarchitecture compared to controls. BPA-exposed mammary glands exhibited a more developed epithelial tree and a greater than 300% increase in the relative area of alveolar buds (Figure 4). The overall appearance of mammary tissue of these BPA-exposed virgin mice resembled that of early pregnancy. Additionally, these glands displayed an altered ratio of DNA synthesis between the stroma and the epithelium as detected by BrdU incorporation; DNA synthesis was decreased in the epithelium at 10 days of age and in the stroma at puberty and increased in the stroma at 6 months of age (Figure 5) (70). The mammary gland of male offspring also showed morphological changes due to BPA exposure (Figure 6) (71).
Figure 3.

Increased lateral branching observed in adult mammary glands of mice exposed in utero to BPA. (A) Number of side branches per 500 µm of ductal length in mammary glands of 4 month-old mice treated perinatally with vehicle, 25 ng BPA/kg bw/day and 250 ng BPA/kg bw/day (* p<0.05). Error bars indicate SEM. (B) Whole mounts of adult mammary glands of vehicle (left panel) and 25 ng BPA/kg bw/day (right panel) treated animals. Scale bar represents 1 mm. Originally published in Munoz de Toro et al, Endocrinology (69) Copyright 2005, The Endocrine Society.
Figure 4.

Quantification of ductal and alveolar structures shows an increase in the relative area of (A) ducts, (B) terminal ducts and, (C) alveolar buds in mammary glands of 6 months-old mice exposed in utero to 25 or 250 ng/kg of BPA relative to control. (*p<0.05) relative to control. Originally published in Markey et al. Biol. Reprod. 2001 (70).
Figure 5.

Ratio of DNA synthesis between the stroma and the epithelium. Bar graph represents the ratio of the percentage of epithelial cells that incorporated BrdU to the percentage of stromal cells that incorporated BrdU in the mammary glands of 6 months-old mice exposed in utero to 25 or 250 ng/kg of BPA or control. Originally published in Markey et al. Biol. Reprod. 2001 (70).
Figure 6.

Whole mount mammary glands of male mice at two different ages. These animals were exposed in utero to BPA. The black box indicates samples where the epithelium was small. All samples from the same age were taken at the same magnification. Scale bar represents 1 mm in all panels. Reprinted from Vandenberg et al, Reproductive Toxicology 2013 (71) with permission from Elsevier.
In terms of functionality, the adult mammary gland of BPA-exposed fetuses were more sensitive to estradiol (69) and to progesterone (72). Decreased milk production was observed in lactating F1 females that had been perinatally exposed to BPA. Their milk composition was also altered as it showed a lower content of β-casein than controls (73). We set out to investigate whether BPA could also exert its deleterious effects at the genomic level by inducing epigenetic changes in the developing mammary gland. The DNA methylation profile of mammary glands was analyzed at PND 4, 21 and 50 of WF rats exposed in utero to BPA. BPA-induced alterations in the DNA methylation profile were observed throughout the genome at all three time points (Figure 7). However, genome-wide changes in mRNA expression were only observed at PND 50, i.e., the time at which increased incidence of intraductal hyperplasias and DCIS are observed in this rat strain (74). Therefore, these changes appear to be a consequence of the developmental alterations caused by BPA rather than the cause of neoplastic development later in adulthood. Thus, again, this evidence validates the hypothesis that BPA-induced carcinogenesis is the result of altered stromal-epithelial interactions in the fetal mammary gland.
Figure 7.

BPA-induced changes in DNA methylation observed at PND 4, PND 21 and PND 50 in the mammary glands of perinatally exposed Wistar-Furth rats. The red graphs to the left of each chromosome indicate relative numbers of the transcriptional initiation sites found within a 1 megabase window, which corresponds to the density of promoters. Originally published in Dhimolea et al PLOS ONE 2014 (74).
11. Developmental exposure to BPA increases susceptibility to mammary carcinogenesis
Animal studies showed that BPA exacerbates the effects of carcinogens such as NMU and dimethylbenzanthracene (DMBA) in the mammary gland. Rats that were exposed through lactation to BPA 250 µg/kg/day developed significantly higher numbers of palpable tumors induced by a one-time exposure to DMBA at PND 50 compared to sesame-oil treated animals (23). Interestingly, rats exposed in utero to BPA 250 µg/kg/day showed decreased tumor latency after a one-time DMBA exposure at PND 100 compared to controls (fetally exposed to sesame oil), indicating a shift in the window of sensitivity from PND50 to PND100. Furthermore, the percentage of rats with DMBA-induced tumors and number of tumors per rat were significantly higher in prenatally exposed BPA compared to sesame-oil treated controls (21). In utero exposure of rats to BPA 25 µg/kg/day augmented the responsiveness of the mammary gland to a subcarcinogenic dose of NMU compared to controls. The BPA treated rats showed a significant increase in hyperplastic lesions and further, 13% of the rats developed mammary tumors compared to 0% in controls (22). Altogether, these studies demonstrate that developmental exposure to BPA is able to enhance a carcinogenic insult to the mammary gland. In addition, BPA alone is sufficient to induce mammary gland neoplasia. In utero exposure to BPA induced ductal hyperplasia at all doses (2.5 – 1000 µg/kg/day) and carcinoma in situ at the higher doses (250 and 1000 µg/kg/day) in Wistar-Furth rats (24). Moreover, perinatal exposure to environmentally relevant doses of BPA induced adenocarcinoma of the rat mammary gland at PND90 in Sprague-Dawley rats (26).
12. Is the HPOA involved?
In addition to direct effects on reproductive tissues, perinatal BPA exposure may also exert indirect effects by altering development and function of hypothalamic-pituitary axes, including the HPOA. Studies in BPA-exposed rats and mice have documented alterations in estrous cyclicity (75), ovarian follicular development and in the genital tract (76), and in circulating hormone levels (76;77) all of which could be related to alterations at the hypothalamus and/or pituitary. Changes in the ER levels in the hypothalamus and pituitary have been reported following perinatal exposure to BPA (77–79). Perinatal BPA exposure alters the size and composition of the AVPV (80;81), a brain region essential for cyclic gonadotropin release and the preovulatory LH surge. These results suggest that effects of perinatal BPA exposure at the hypothalamic/pituitary level may contribute to alterations in circulating ovarian hormones and in mammotropic hormones that could in turn influence mammary gland development. Animals exposed to BPA show alveolar development at ages where controls do not (70), suggesting a role for altered prolactin secretion.
13. Conclusions
The Environmental Protection Agency (EPA) defines a carcinogen as a chemical or physical agent capable of causing cancer (82). Under this definition, BPA should now be considered a direct mammary gland carcinogen. In the context of TOFT, a likely explanation for BPA-induced breast carcinogenesis emerges. Namely, by binding to ERs present in the primary mesenchyme, BPA affects collagen deposition and adipocyte differentiation. This modifies the mechanical properties of the ECM resulting in a denser and stiffer tissue, which is consistent with delayed lumen formation. The fact that mammographic density, due to a dense stromal compartment, is a main risk factor for breast cancer buttresses this hypothesis. The affected stroma would induce alterations in epithelial development such as increased branching and ductal growth. Finally, the altered mammary gland displays an abnormal response to natural hormones and is more susceptible to carcinogenic insults, developing intraductal hyperplasias and DCIS later in adulthood (Figure 8).
Figure 8.
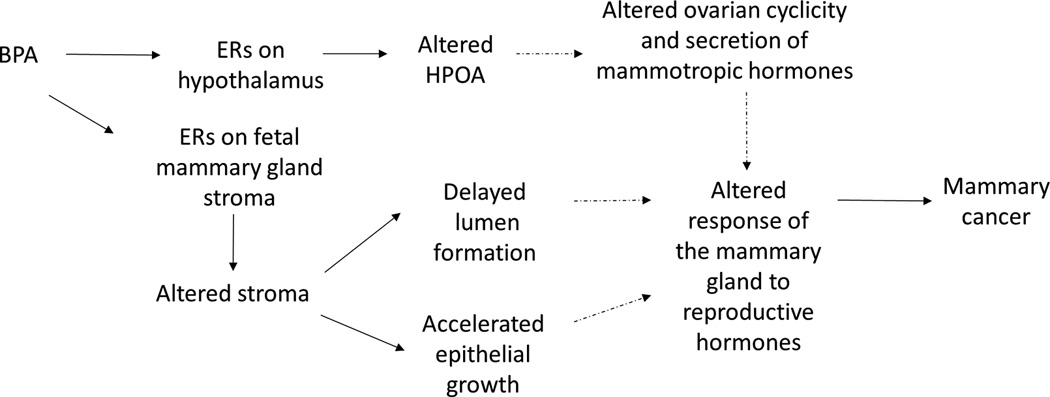
BPA exposure alters mammary gland development and increases the risk of mammary carcinogenesis: BPA binds to the fetal mammary gland mesenchymal ERs, and in turn affects the composition of the ECM increasing tissue rigidity. Increased rigidity delays lumen formation. BPA also induces precocious adipocyte differentiation, which in turn accelerates duct elongation and branching. These changes lead to an increased sensitivity to mammotropic hormones in adulthood. BPA also binds to ER in the hypothalamus, where it alters the control of ovarian cyclicity and the control of the secretion of mammotropic hormones. The solid arrows link effects of in utero exposure to BPA in rodents and non-human primates. Dashed arrows indicate hypothesized links between effects during fetal mammary gland development and mammary carcinogenesis.
Highlights.
The primary target of BPA is the stroma of the fetal mammary gland.
BPA alters reciprocal stroma-epithelium interactions that regulate mammogenesis.
Perinatal BPA exposure sensitizes the mammary gland to additional carcinogen hits.
BPA is a direct mammary gland carcinogen.
Carcinogenesis is development gone awry.
Acknowledgments
We greatly appreciate the editorial contributions by Cheryl Schaeberle and Dr. Beverly Rubin.
Funding statement:
This research was supported by The Avon Foundation grants #02-2009-093, and 02-2011-025 as well as by the National Institute of Environmental Health Sciences, Award Numbers R01ES08314, RC2ES018822. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institute of Environmental Health Sciences or the National Institutes of Health.
Abbreviations
- EDC
Endocrine Disrupting Chemicals
- BPA
bisphenol A
- DES
diethylstilbisterol
- HPOA
hypothalamus-pituitary-ovarian axis
- DCIS
ductal carcinoma in situ
- GPR30
G-protein coupled receptor 30
- ERR
Estrogen receptor related
- ECM
extracellular matrix
- NMU
nitrosomethyl urea
- TOFT
Tissue Organization Field Theory
- PND
postnatal day
- GD
gestational day
- AVPV
anteroventral periventricular nucleus
- EPA
Environmental Protection Agency
- DMBA
dimethylbenzanthracene
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Contributor Information
Tessie Paulose, Email: Tessie.Paulose@tufts.edu.
Lucia Speroni, Email: Lucia.Speroni@tufts.edu.
Carlos Sonnenschein, Email: Carlos.Sonnenschein@tufts.edu.
Reference List
- 1.Palmer JR, Wise LA, Hatch EE, Troisi R, Titus-Ernstoff L, Strohsnitter W, et al. Prenatal diethylstilbestrol exposure and risk of breast cancer. Cancer Epidem Biomar. 2006;15:1509–1514. doi: 10.1158/1055-9965.EPI-06-0109. [DOI] [PubMed] [Google Scholar]
- 2.de Assis S, Warri A, Cruz MI, Laja O, Tian Y, Zhang B, et al. High-fat or ethinyl-oestradiol intake during pregnancy increases mammary cancer risk in several generations of offspring. Nat Commun. 2012;3:1053. doi: 10.1038/ncomms2058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.NCI (National Cancer Institute) Diethylstilbestrol (DES) and Cancer. 2011 Available from: URL: http://www.cancer.gov/cancertopics/factsheet/Risk/DES.
- 4.Trichopoulos D. Is breast cancer initiated in utero? Epidemiology. 1990;1:95–96. [PubMed] [Google Scholar]
- 5.Fenton SE, Hamm JT, Birnbaum L, Youngblood GL. Persistent abnormalities in the rat mammary gland following gestational and lactational exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) Toxicol Sci. 2002;67:63–74. doi: 10.1093/toxsci/67.1.63. [DOI] [PubMed] [Google Scholar]
- 6.Tomooka Y, Bern HA. Growth of mouse mammary glands after neonatal sex hormone treatment. J Nat Cancer Inst. 1982;69:1347–1352. [PubMed] [Google Scholar]
- 7.Vandenberg LN, Maffini MV, Sonnenschein C, Rubin BS, Soto AM. Bisphenol-A and the great divide: A review of controversies in the field of endocrine disruption. Endocr Rev. 2009;30:75–95. doi: 10.1210/er.2008-0021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zoeller RT, Brown TR, Doan L, Gore AC, Skakkebaek N, Soto AM, et al. Endocrine-dsrupting chemicals and public health protection: A statement of principles from The Endocrine Society. Endocrinology. 2012;153:4097–4110. doi: 10.1210/en.2012-1422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Burridge E. Chemical profile: bisphenol A. ICIS Chemical Business. 2008;274:48. [Google Scholar]
- 10.Calafat AM, Ye X, Wong LY, Reidy JA, Needham LL. Exposure of the U.S. population to bisphenol A and 4-tertiary-Octylphenol: 2003–2004. Environ Health Perspect. 2008;116:39–44. doi: 10.1289/ehp.10753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Schonfelder G, Wittfoht W, Hopp H, Talsness CE, Paul M, Chahoud I. Parent bisphenol A accumulation in the human maternal-fetal-placental unit. Environ Health Perspect. 2002;110:A703–A707. doi: 10.1289/ehp.110-1241091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sun Y, Irie M, Kishikawa N, Wada M, Kuroda N, Nakashima K. Determination of bisphenol A in human breast milk by HPLC with column-switching and fluorescence detection. Biomed Chromatogr. 2004;18:501–507. doi: 10.1002/bmc.345. [DOI] [PubMed] [Google Scholar]
- 13.Sharpe RM, Skakkebaek NE. Are oestrogens involved in falling sperm count and disorders of the male reproductive tract? Lancet. 1993;341:1392–1395. doi: 10.1016/0140-6736(93)90953-e. [DOI] [PubMed] [Google Scholar]
- 14.Skakkebaek NE, Meyts ER, Jorgensen N, Carlsen E, Petersen PM, Giwercman A, et al. Germ cell cancer and disorders of spermatogenesis: an environmental connection? APMIS. 1998;106:3–12. doi: 10.1111/j.1699-0463.1998.tb01314.x. [DOI] [PubMed] [Google Scholar]
- 15.Hoyer AP, Grandjean P, Jorgensen T, Brock JW, Hartvig HB. Organochloride exposure and risk of breast cancer. Lancet. 1998;352:1816–1820. doi: 10.1016/S0140-6736(98)04504-8. [DOI] [PubMed] [Google Scholar]
- 16.Cohn BA, Wolff MS, Cirillo PM, Sholtz RI. DDT and breast cancer in young women: New data on the significance of age at exposure. Environ Health Perspect. 2007;115:1406–1414. doi: 10.1289/ehp.10260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Skakkebaek NE, Rajpert-DeMeyts E, Main KM. Testicular dysgenesis syndrome: an increasingly common developmental disorder with environmental aspects. Hum Reprod. 2001;16:972–978. doi: 10.1093/humrep/16.5.972. [DOI] [PubMed] [Google Scholar]
- 18.Vandenberg LN, Maffini MV, Wadia PR, Sonnenschein C, Rubin BS, Soto AM. Exposure to environmentally relevant doses of the xenoestrogen bisphenol-A alters development of the fetal mouse mammary gland. Endocrinology. 2007;148:116–127. doi: 10.1210/en.2006-0561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Tharp AP, Maffini MV, Hunt PA, Vandevoort CA, Sonnenschein C, Soto AM. Bisphenol A alters the development of the rhesus monkey mammary gland. Proc Natl Acad Sci U S A. 2012 May 7;109:8190–8195. doi: 10.1073/pnas.1120488109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wadia PR, Cabaton NJ, Borrero MD, Rubin BS, Sonnenschein C, Shioda T, et al. Low-dose BPA exposure alters the mesenchymal and epithelial transcriptomes of the mouse fetal mammary gland. PLoS One. 2013;8:e63902. doi: 10.1371/journal.pone.0063902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Betancourt AM, Eltoum IA, Desmond RA, Russo J, Lamartiniere CA. In utero exposure to Bisphenol A shifts the window of susceptibility for mammary carcinogenesis in the rat. Environ Health Perspect. 2010 Jul 30;118:1614–1619. doi: 10.1289/ehp.1002148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Durando M, Kass L, Piva J, Sonnenschein C, Soto AM, Luque EH, et al. Prenatal bisphenol A exposure induces preneoplastic lesions in the mammary gland in Wistar rats. Environ Health Perspect. 2007;115:80–86. doi: 10.1289/ehp.9282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Jenkins S, Raghuraman N, Eltoum I, Carpenter M, Russo J, Lamartiniere CA. Oral exposure to Bisphenol A increases dimethylbenzanthracene-induced mammary cancer in rats. Environ Health Perspect. 2009;117:910–915. doi: 10.1289/ehp.11751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Murray TJ, Maffini MV, Ucci AA, Sonnenschein C, Soto AM. Induction of mammary gland ductal hyperplasias and carcinoma in situ following fetal Bisphenol A exposure. Reproductive Toxicology. 2007;23:383–390. doi: 10.1016/j.reprotox.2006.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Weber Lozada K, Keri RA. Bisphenol A increases mammary cancer risk in two distinct mouse models of breast cancer. Biol Reprod. 2011;85:490–497. doi: 10.1095/biolreprod.110.090431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Acevedo N, Davis B, Schaeberle CM, Sonnenschein C, Soto AM. Perinatally administered Bisphenol A as a potential mammary gland carcinogen in rats. Environ Health Perspect. 2013;121:1040–1046. doi: 10.1289/ehp.1306734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Markey CM, Rubin BS, Soto AM, Sonnenschein C. Endocrine disruptors from Wingspread to environmental developmental biology. J Steroid Biochem Molec Biol. 2003;83:235–244. doi: 10.1016/s0960-0760(02)00272-8. [DOI] [PubMed] [Google Scholar]
- 28.Ekbom A, Trichopoulos D, Adami HO, Hsieh CC, Lan SJ. Evidence of prenatal influences on breast cancer risk. Lancet. 1992;340:1015–1018. doi: 10.1016/0140-6736(92)93019-j. [DOI] [PubMed] [Google Scholar]
- 29.Braun MM, Ahlbom A, Floderus B, Brinton LA, Hoover RN. Effect of twinship on incidence of cancer of the testis, breast, and other sites (Sweden) CCC. 1995;6:519–524. doi: 10.1007/BF00054160. [DOI] [PubMed] [Google Scholar]
- 30.Potischman N, Troisi R. In-utero and early life exposures in relation to risk of breast cancer. CCC. 1999;10:561–573. doi: 10.1023/a:1008955110868. [DOI] [PubMed] [Google Scholar]
- 31.Tamimi R, Lagiou P, Vatten LJ, Mucci L, Trichopoulos D, Hellerstein S, et al. Pregnancy hormones, pre-eclampsia, and implications for breast cancer risk in the offspring. Cancer Epidemiol Biomarkers Prev. 2003;12:647–650. [PubMed] [Google Scholar]
- 32.Herbst AL, Ulfelder H, Poskanzer DC. Adenocarcinoma of the vagina: association of maternal stilbestrol therapy with tumor appearance in young women. New Engl J Med. 1971;284:878–881. doi: 10.1056/NEJM197104222841604. [DOI] [PubMed] [Google Scholar]
- 33.Herbst AL, Bern HA. Developmental Effects of Diethylstilbestrol (DES) in Pregnancy. New York: Thieme-Stratton; 1988. [Google Scholar]
- 34.Herbst AL, Anderson D. Clear cell adenocarcinoma of the vagina and cervix secondary to intrauterine exposure to diethylstilbestrol. Seminars in Surgical Oncology. 1990;6:343–346. doi: 10.1002/ssu.2980060609. [DOI] [PubMed] [Google Scholar]
- 35.Mittendorf R. Teratogen update: carcinogenesis and teratogenesis associated with exposure to diethylstilbestrol (DES) in utero. Teratology. 1995;51:435–445. doi: 10.1002/tera.1420510609. [DOI] [PubMed] [Google Scholar]
- 36.Hoover RN, Hyer M, Pfeiffer RM, Adam E, Bond B, Cheville AL, et al. Adverse health outcomes in women exposed in utero to diethylstilbestrol. N Engl J Med. 2011;365:1304–1314. doi: 10.1056/NEJMoa1013961. [DOI] [PubMed] [Google Scholar]
- 37.Green S, Walter P, Kumar V, Krust A, Bornert JM, Argos P, et al. Human oestrogen receptor cDNA: sequence, expression and homology to v-erb-A. Nature. 1986;320:134–139. doi: 10.1038/320134a0. [DOI] [PubMed] [Google Scholar]
- 38.Kuiper GG, Enmark E, Pelto-Huikko M, Nilsson S, Gustafsson JA. Cloning of a novel receptor expressed in rat prostate and ovary. Proc Nat Acad Sci USA. 1996;93:5925–5930. doi: 10.1073/pnas.93.12.5925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Carmeci C, Thompson DA, Ring HZ, Francke U, Weigel RJ. Identification of a gene (GPR30) with homology to the G-protein-coupled receptor superfamily associated with estrogen receptor expression in breast cancer. Genomics. 1997;45:607–617. doi: 10.1006/geno.1997.4972. [DOI] [PubMed] [Google Scholar]
- 40.Couse JF, Korach KS. Estrogen receptor null mice: what have we learned and where will they lead us? Endocr Rev. 1999;20:358–417. doi: 10.1210/edrv.20.3.0370. [DOI] [PubMed] [Google Scholar]
- 41.Couse JF, Mahato D, Eddy EM, Korach KS. Molecular mechanism of estrogen action in the male: insights from the estrogen receptor null mice. Reprod Fertil Dev. 2001;13:211–219. doi: 10.1071/rd00128. [DOI] [PubMed] [Google Scholar]
- 42.Watson CS, Pappas TC, Gametchu B. The other estrogen receptor in the plasma membrane: implications for the actions of environmental estrogens. Environ Health Perspect. 1995;103(Suppl):41–50. doi: 10.1289/ehp.95103s741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Nadal A, Ropero AB, Laribi O, Maillet M, Fuentes E, Soria B. Nongenomic actions of estrogens and xenoestrogens by binding at a plasma membrane receptor unrelated to estrogen recpetor α and estrogen receptor β. Proc Nat Acad Sci USA. 2000 Oct 10;97:11603–11608. doi: 10.1073/pnas.97.21.11603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Watson CS, Bulayeva NN, Wozniak AL, Finnerty CC. Signaling from the membrane via membrane estrogen receptor-alpha: estrogens, xenoestrogens, and phytoestrogens. Steroids. 2005;70:364–371. doi: 10.1016/j.steroids.2005.03.002. [DOI] [PubMed] [Google Scholar]
- 45.Heard DJ, Norby PL, Holloway J, Vissing H. Human ERRgamma, a third member of the estrogen receptor-related receptor (ERR) subfamily of orphan nuclear receptors: tissue-specific isoforms are expressed during development and in the adult. Mol Endocrinol. 2000;14:382–392. doi: 10.1210/mend.14.3.0431. [DOI] [PubMed] [Google Scholar]
- 46.Sheehan DM, Willingham E, Gaylor D, Bergeron JM, Crews D. No threshold dose for estradiol-induced sex reversal of turtle embryos: How little is too much? Environ Health Perspect. 1999;107:155–159. doi: 10.1289/ehp.99107155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Vandenberg LN, Hauser R, Marcus M, Olea N, Welshons WV. Human exposure to bisphenol A (BPA) Reproductive Toxicology. 2007;24:139–177. doi: 10.1016/j.reprotox.2007.07.010. [DOI] [PubMed] [Google Scholar]
- 48.Ye X, Kuklenyik Z, Needham J, Calafat AM. Measuring environmental phenols and chlorinated organic chemicals in breast milk using automated on-line column switching-high performance liquid chromatography-isotope dilution tandem mass spectrometry. Journal of Chromatography B. 2006;831:110–115. doi: 10.1016/j.jchromb.2005.11.050. [DOI] [PubMed] [Google Scholar]
- 49.Calafat AM, Kuklenyik Z, Reidy JA, Caudill SP, Ekong J, Needham JL. Urinary concentrations of Bisphenol A and 4-Nonylphenol in a human reference population. Environ Health Perspect. 2005;113:391–395. doi: 10.1289/ehp.7534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Woodruff TJ, Zota AR, Schwartz JM. Environmental chemicals in pregnant women in the United States: NHANES 2003–2004. Environ Health Perspect. 2011;119:878–885. doi: 10.1289/ehp.1002727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Gilbert SF. Mechanisms for the environmental regulation of gene expression: ecological aspects of animal development. J Biosci. 2005;30:65–74. doi: 10.1007/BF02705151. [DOI] [PubMed] [Google Scholar]
- 52.Barker DJP, Eriksson JG, Osmond C. Fetal origins of adult disease: strength of effects and biological basis. Int J Epidemiol. 2002;31:1235–1239. doi: 10.1093/ije/31.6.1235. [DOI] [PubMed] [Google Scholar]
- 53.de la Rosa LN. Becoming organisms: the organisation of development and the development of organisation. Hist Phil Life Sci. 2010;32:289–315. [PubMed] [Google Scholar]
- 54.Widelitz RB, Veltmaat JM, Mayer JA, Foley J, Chuong CM. Mammary glands and feathers: comparing two skin appendages which help define novel classes during vertebrate evolution. Semin Cell Dev Biol. 2007 Apr;18:255–266. doi: 10.1016/j.semcdb.2007.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Mikkola ML, Millar SE. The mammary bud as a skin appendage: unique and shared aspects of development. J Mammary Gland Biol Neoplasia. 2006;11:187–203. doi: 10.1007/s10911-006-9029-x. [DOI] [PubMed] [Google Scholar]
- 56.Kratochwil K. Organ specificity in mesenchymal induction demonstrated in the embryonic development of the mammary gland of the mouse. Dev Biol. 1969;20:46–71. doi: 10.1016/0012-1606(69)90004-9. [DOI] [PubMed] [Google Scholar]
- 57.Sakakura T, Nishizuka Y, Dawe CJ. Mesenchyme-dependent morphogenesis and epithelium-specific cytodifferentiation in mouse mammary gland. Science. 1976;194:1439–1441. doi: 10.1126/science.827022. [DOI] [PubMed] [Google Scholar]
- 58.Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011 Mar 4;144:646–674. doi: 10.1016/j.cell.2011.02.013. [DOI] [PubMed] [Google Scholar]
- 59.Soto AM, Sonnenschein C. The tissue organization field theory of cancer: A testable replacement for the somatic mutation theory. BioEssays. 2011 May;33:332–340. doi: 10.1002/bies.201100025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Veltmaat JM, Mailleux AA, Thiery JP, Bellusci S. Mouse embryonic mammogenesis as a model for the molecular regulation of pattern formation. Differentiation. 2003;71:1–17. doi: 10.1046/j.1432-0436.2003.700601.x. [DOI] [PubMed] [Google Scholar]
- 61.Cowin P, Wysolmerski J. Molecular mechanisms guiding embryonic mammary gland development. Cold Spring Harb Perspect Biol. 2010 Jun 1;2:a003251. doi: 10.1101/cshperspect.a003251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Maffini MV, Soto AM, Calabro JM, Ucci AA, Sonnenschein C. The stroma as a crucial target in rat mammary gland carcinogenesis. J Cell Sci. 2004;117:1495–1502. doi: 10.1242/jcs.01000. [DOI] [PubMed] [Google Scholar]
- 63.Maffini MV, Calabro JM, Soto AM, Sonnenschein C. Stromal regulation of neoplastic development: Age-dependent normalization of neoplastic mammary cells by mammary stroma. Am J Pathol. 2005;167:1405–1410. doi: 10.1016/S0002-9440(10)61227-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Bussard KM, Smith GH. Human breast cancer cells are redirected to mammary epithelial cells upon interaction with the regenerating mammary gland microenvironment in-vivo. PLoS ONE. 2012;7:e49221. doi: 10.1371/journal.pone.0049221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Hendrix MJ, Seftor EA, Seftor RE, Kasemeier-Kulesa J, Kulesa PM, Postovit LM. Reprogramming metastatic tumour cells with embryonic microenvironments. Nat Rev Cancer. 2007;7:246–255. doi: 10.1038/nrc2108. [DOI] [PubMed] [Google Scholar]
- 66.Coleman WB, Wennerberg AE, Smith GJ, Grisham JW. Regulation of the differentiation of diploid and some aneuploid rat liver epithelial (stemlike) cells by the hepatic microenvironment. Am J Pathol. 1993 May;142:1373–1382. [PMC free article] [PubMed] [Google Scholar]
- 67.Lemmen JG, Broekhof JLM, Kuiper GGJM, Gustafsson JA, Van Der Saag PT, van der Burg B. Expression of estrogen receptor alpha and beta during mouse embryogenesis. Mech Dev. 1999;81:163–167. doi: 10.1016/s0925-4773(98)00223-8. [DOI] [PubMed] [Google Scholar]
- 68.Shioda T, Chesnes J, Coser KR, Zou L, Hur J, Dean KL, et al. Importance of dosage standardization for interpreting transcriptomal signature profiles: Evidence from studies of xenoestrogens. Proc Nat Acad Sci USA. 2006;103:12033–12038. doi: 10.1073/pnas.0605341103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Munoz de Toro MM, Markey CM, Wadia PR, Luque EH, Rubin BS, Sonnenschein C, et al. Perinatal exposure to Bisphenol A alters peripubertal mammary gland development in mice. Endocrinology. 2005;146:4138–4147. doi: 10.1210/en.2005-0340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Markey CM, Luque EH, Munoz de Toro MM, Sonnenschein C, Soto AM. In utero exposure to bisphenol A alters the development and tissue organization of the mouse mammary gland. Biol Reprod. 2001;65:1215–1223. doi: 10.1093/biolreprod/65.4.1215. [DOI] [PubMed] [Google Scholar]
- 71.Vandenberg LN, Schaeberle CM, Rubin BS, Sonnenschein C, Soto AM. The male mammary gland: a target for the xenoestrogen bisphenol A. Reproductive Toxicology. 2013;37:15–23. doi: 10.1016/j.reprotox.2013.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Ayyanan A, Laribi O, Schuepbach-Mallepell S, Schrick C, Gutierrez M, Tanos T, et al. Perinatal exposure to bisphenol a increases adult mammary gland progesterone response and cell number. Mol Endocrinol. 2011 Nov;25:1915–1923. doi: 10.1210/me.2011-1129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Kass L, Altamirano GA, Bosquiazzo VL, Luque EH, Munoz de Toro M. Perinatal exposure to xenoestrogens impairs mammary gland differentiation and modifies milk composition in Wistar rats. Reproductive Toxicology. 2012;33:390–400. doi: 10.1016/j.reprotox.2012.02.002. [DOI] [PubMed] [Google Scholar]
- 74.Dhimolea E, Wadia PR, Murray TJ, Settles ML, Treitman JD, Sonnenschein C, et al. Prenatal exposure to BPA alters the epigenome of the rat mammary gland and increases the propensity to neoplastic development. PLoS ONE. 2014 doi: 10.1371/journal.pone.0099800. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Rubin BS, Murray MK, Damassa DA, King JC, Soto AM. Perinatal exposure to low doses of bisphenol-A affects body weight, patterns of estrous cyclicity and plasma LH levels. Environ Health Perspect. 2001;109:675–680. doi: 10.1289/ehp.01109675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Markey CM, Coombs MA, Sonnenschein C, Soto AM. Mammalian development in a changing environment: exposure to endocrine disruptors reveals the developmental plasticity of steroid-hormone target organs. Evolution and Development. 2003;5:1–9. doi: 10.1046/j.1525-142x.2003.03011.x. [DOI] [PubMed] [Google Scholar]
- 77.Khurana S, Ranmal S, Ben-Jonathan N. Exposure of newborn male and female rats to environmental estrogens: delayed and sustained hyperprolatinemia and alterations in estrogen receptor expression. Endocrinology. 2000;141:4512–4517. doi: 10.1210/endo.141.12.7823. [DOI] [PubMed] [Google Scholar]
- 78.Monje L, Varayoud J, Munoz-de-Toro M, Luque EH, Ramos JG. Exposure of neonatal female rats to bisphenol A disrupts hypothalamic LHRH pre-mRNA processing and estrogen receptor alpha expression in nuclei controlling estrous cyclicity. Reprod Toxicol. 2010 Oct 15; doi: 10.1016/j.reprotox.2010.08.004. [DOI] [PubMed] [Google Scholar]
- 79.Ramos JG, Varayoud J, Kass L, Rodriguez H, Costabel L, Munoz de Toro MM, et al. Bisphenol a induces both transient and permanent histofunctional alterations of the hypothalamic-pituitary-gonadal axis in prenatally exposed male rats. Endocrinology. 2003;144:3206–3215. doi: 10.1210/en.2002-0198. [DOI] [PubMed] [Google Scholar]
- 80.Rubin BS, Lenkowski JR, Schaeberle CM, Vandenberg LN, Ronsheim PM, Soto AM. Evidence of altered brain sexual differentiation in mice exposed perinatally to low environmentally relevant levels of bisphenol A. Endocrinology. 2006;147:3681–3691. doi: 10.1210/en.2006-0189. [DOI] [PubMed] [Google Scholar]
- 81.Patisaul HB, Fortino AE, Polston EK. Neonatal genistein or bisphenol-A exposure alters sexual differentiation of the AVPV. Neurotoxicol Teratol. 2006 Jan;28:111–118. doi: 10.1016/j.ntt.2005.11.004. [DOI] [PubMed] [Google Scholar]
- 82.Technology Transfer Network Air Toxics; Glossary of Key Terms. 2005 Available from: URL: http://www.epa.gov/ttn/atw/natamain/gloss1.html.