

Abstract
Human leukocyte antigen (HLA)-G is a non-classical HLA class I molecule with known immune-modulatory functions. Our group identified a subset of human dendritic cells, named DC-10, that induce adaptive interleukin-10-producing T regulatory type 1 (Tr1) cells via the interleukin-10-dependent HLA-G/ILT4 pathway. In this study we aimed at defining the role of HLA-G in DC-10-mediated Tr1 cell differentiation. We analyzed phenotype, functions, and genetic variations in the 3′ untranslated region of the HLA-G locus of in vitro-differentiated DC-10 from 67 healthy donors. We showed that HLA-G expression on DC-10 is donor-dependent. Functional studies demonstrated that DC-10, independently of HLA-G expression, secrete interleukin-10 and negligible levels of interleukin-12. Interestingly, DC-10 with high HLA-G promote allo-specific anergic T cells that contain a significantly higher frequency of Tr1 cells, defined as interleukin-10-producing (P=0.0121) or CD49b+LAG-3+ (P=0.0031) T cells, compared to DC-10 with low HLA-G. We found that the HLA-G expression on DC-10 is genetically imprinted, being associated with specific variations in the 3′ untranslated region of the gene, and it may be finely tuned by microRNA-mediated post-transcriptional regulation. These data highlight the important role of HLA-G in boosting DC-10 tolerogenic activity and confirm that interleukin-10 production by DC-10 is necessary but not sufficient to promote Tr1 cells at high frequency. These new insights into the role of HLA-G in DC-10-mediated induction of Tr1 cells provide additional information for clinical use in Tr1- or DC-10-based cell therapy approaches.
Introduction
Human leukocyte antigen (HLA)-G is a non-classical HLA class I molecule originally identified and characterized in trophoblasts where it confers protection to the semi-allogeneic fetus from the maternal immune system.1,2 More recently, it has become evident that HLA-G plays a role in promoting and maintaining tolerance in autoimmunity and organ transplantation, and immune escape in cancer and infectious diseases. HLA-G is characterized by a relatively low allelic polymorphism and a highly restricted tissue distribution in comparison with classical HLA class I molecules. In healthy conditions, in addition to trophoblasts, HLA-G expression is restricted to amniotic cells, adult thymic medulla, erythroblasts, cornea, pancreatic islets, and endothelial and stem cells.3 By alternative splicing of a single mRNA transcript, seven different HLA-G isoforms, four membrane-bound (HLA-G1, -G2, -G3, and -G4) and three soluble (HLA-G5, -G6, and -G7) can be generated.4,5 Matrix metalloproteinases are responsible for the proteolytic cleavage of HLA-G1 from the membrane, and the release of shed HLA-G1.6 Thus far, the best-characterized isoforms are HLA-G1, the most stable membrane-bound isoform, HLA-G5, and shed HLA-G1.
Several polymorphisms in both the promoter and the 3′ untranslated region (UTR) of the HLA-G gene can influence its expression.7 The best-characterized polymorphism in the 3′UTR is the insertion/deletion (Ins/Del) of 14 base pairs (bp), which has been respectively associated with decreased/increased mRNA stability.8–10 The +3142 G/C single nucleotide polymorphism has also been suggested to control HLA-G expression, since the presence of G may increase the affinity of mRNA for the microRNA (miR)-148a, miR-148b and, miR-152.11,12 Additional less studied single nucleotide polymorphisms are present in the 3′UTR at positions +3001 T/C, +3003 T/C, +3010 G/C, +3027 C/A, +3035 C/T, and +3196 C/G,13 which are potential miRNA-binding sites, and thus may regulate HLA-G expression,14 and +3187 A/G located in the proximity of an AU-rich motif, which affects mRNA stability.15 These polymorphisms cluster in at least 15 different haplotypes identified in Brazilian populations,13,16 and a number of less frequent haplotypes found in other populations.17,18
The functions of HLA-G are oriented towards immune inhibition and induction of tolerance. Through direct binding to the inhibitory receptors Immunoglobulin-Like-Transcript (ILT)2 and ILT4, and Killer cell Immunoglobulin-Like Receptor (KIR)2DL4,19 HLA-G mediates short-term inhibition of natural killer (NK) cells,20 cytotoxic T-lymphocytes,21 B cells,22 and dendritic cells (DC)23,24 and long-term induction of regulatory cells including HLA-G-expressing T regulatory (Treg) cells,23 CD4low and CD8low suppressor T cells,25 interleukin-10 (IL-10)-producing T regulatory type 1 (Tr1) cells,26 and DC-10.27
DC-10 are a subset of human tolerogenic DC that are present in vivo27–29 and can be differentiated in vitro by culturing monocytes in the presence of IL-10. DC-10 secrete IL-10, are CD11c+, express CD14, CD16, HLA-G and ILT4 and, although not activated, display a mature phenotype, being CD86+ and HLA-DR+. The secretion of IL-10 and the expression of membrane-bound HLA-G and ILT4 are critical factors involved in DC-10-mediated induction of Tr1 cells.27 In the present study, we investigated the role of HLA-G in DC-10-mediated Tr1 cell induction and whether polymorphisms present at the 3′UTR of the gene influence the expression of membrane-bound HLA-G on DC-10.
Methods
The methods are described in full in the Online Supplementary Appendix. Peripheral blood was obtained after informed consent in accordance with the Declaration of Helsinki under protocols approved by the ethical committee of the San Raffaele Telethon Institute for Gene Therapy.
Dendritic cell differentiation
CD14+ monocytes were isolated from peripheral blood mononuclear cells by positive selection using CD14 MicroBeads (Miltenyi Biotech, Germany) according to the manufacturer’s instructions. Cells were cultured in RPMI 1640 (Lonza, Italy) supplemented with 10% fetal bovine serum (FBS) (Lonza, Italy) or with 5% human serum (HS) (EuroClone, Italy), 100 U/mL penicillin/streptomycin (Lonza, Italy), 2 mM L-glutamine (Lonza, Italy), (DC medium) at 37°C in the presence of 10 ng/mL recombinant human (rh)IL-4 (R&D Systems, Minneapolis MN, USA) and 100 ng/mL rhGM-CSF (Genzyme, Seattle, WA, USA) with 10 ng/mL of rhIL-10 (BD, Bioscience, CA, USA) for 7 days to differentiate DC-10. Cells cultured with rhIL-4 and rhGM-CSF on day 5 were matured with 1 mg/mL of lipopolysaccharide (Sigma, CA, USA) for 2 more days to generate mature dendritic cells (mDC). At day 7, DC were collected, phenotypically analyzed, and used to stimulate T cells.
Statistical analysis
Sample mean results were compared using the non-parametric Mann-Whitney U test for continuous variables. HLA-G 3′UTR allele and genotype frequencies were obtained by direct counting. Allele and genotype frequencies between populations were compared using the χ2 test. The correlation between membrane-bound HLA-G and ILT4 expression was determined using the Spearman correlation test. DC-10FBS and DC-10HS were compared using a paired t-test. All results are presented as mean values ± standard error of mean (SEM). Differences were regarded as statistically significant at *P<0.05, **P<0.01, and ***P<0.001. The results were analyzed using GraphPad Prism 5.0 (GraphPad Software, Inc. La Jolla, CA, USA).
Results
In vitro differentiated DC-10 express variable levels of membrane-bound HLA-G
Independently of the donor, DC-10 differentiated in vitro as described in the Methods section were CD11c+CD14+CD1a−CD86+HLA-DR+ (Figure 1A,B). High variability in the expression of membrane-bound HLA-G (ranging from 3.5% to 97.7%) and of ILT4 (ranging from 0.5% to 70.7%; Figure 1A,B) was observed. Notably, ILT4 expression correlated with that of HLA-G (R2=0.46, P<0.0001). mDC differentiated from the same donors of DC-10 were CD11c+CD1a+CD14−CD86bright and did not express HLA-G (<3%) (Figure 1C). Based on the expression of HLA-G, DC-10 were classified as HLA-Ghigh and HLA-Glow when the percentage of HLA-G-expressing cells was higher or lower than 38%, respectively. This threshold value was determined as <95% of the confidence interval of the mean of HLA-G-expressing cells among all DC-10 tested.
Figure 1.
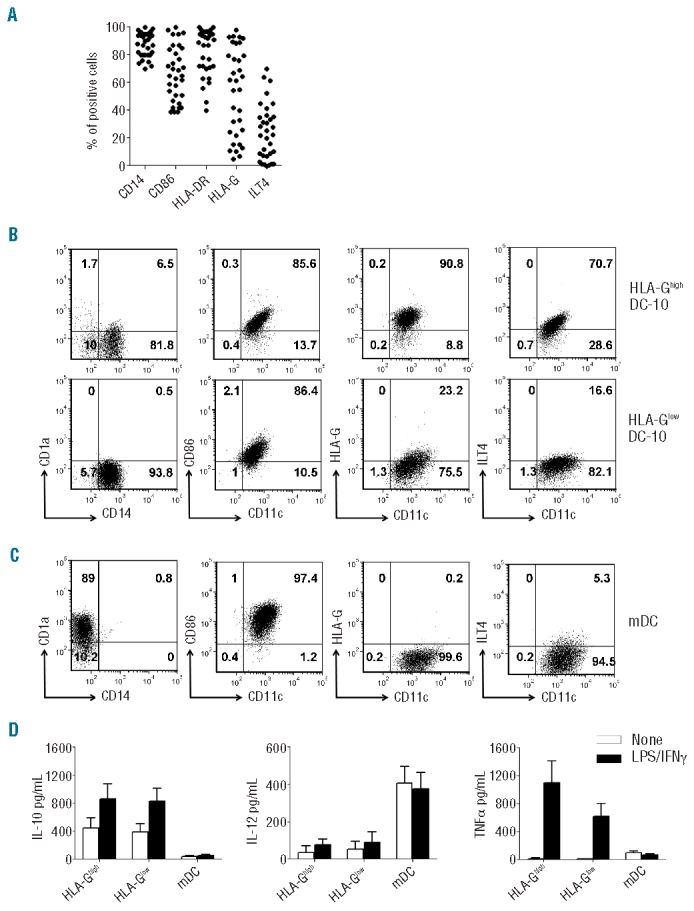
In vitro-differentiated DC-10 express variable levels of membrane-bound HLA-G. Expression levels of CD14, CD1a, CD11c, CD86, HLA-DR, HLA-G, and ILT4 on in vitro differentiated DC-10 and mDC were evaluated by FACS analysis. (A) Percentages of cells positive for the indicated markers in 33 independent donors are shown. (B) One representative donor out of 30 HLA-Ghigh DC-10 donors (upper panels), and one out of 24 HLA-Glow DC-10 donors (lower panels) are presented. Numbers represent percentages of positive cells in each quadrant. (C) One representative donor out of 54 mDC donors is presented. Numbers represent percentages of positive cells in each quadrant. (D) DC-10 were left unstimulated or stimulated with lipopolysaccharide (LPS) and interferon-γ (IFNγ) for 2 days. Concentrations of IL-10, IL-12, and tumor necrosis factor-α (TNFα) in culture supernatants of the indicated cells are shown. Mean±SEM of 11 (HLA-Ghigh DC-10), 15 (HLA-Glow DC-10), and 26 (mDC) independent experiments.
HLA-Ghigh and HLA-Glow DC-10 spontaneously secreted IL-10 at comparable levels, and negligible levels of IL-12 and tumor necrosis factor-α (Figure 1D). Upon stimulation with lipopolysaccharide and interferon-γ, the release of IL-10, but not of IL-12, by DC-10 increased, and no differences were observed between HLA-Ghigh and HLA-Glow DC-10. Activated HLA-Ghigh and HLA-Glow DC-10 secreted comparable amounts of tumor necrosis factor-α (Figure 1D). As expected, mDC differentiated in vitro from the same donors from whom DC-10 were generated, secreted high levels of IL-12, and limited amounts of tumor necrosis factor-α, and of IL-10 at steady state and upon activation (Figure 1D).
To investigate whether the expression of HLA-G by DC-10 was influenced by the culture conditions used for their differentiation (i.e. the presence of FBS), we compared the phenotype and cytokine production profile of DC-10 generated in media supplemented with FBS (DC-10FBS) or HS (DC-10HS). DC-10, independently of the media used, were CD14+CD1a−CD86+HLA−DR+ and expressed HLA-G and ILT4 at variable levels (Figure 2A). DC-10FBS and DC-10HS produced high levels of IL-10 and low amounts of IL-12 spontaneously and tumor necrosis factor-α upon stimulation (Figure 2B). The culture conditions used for DC-10 generation do not affect the phenotype of differentiated DC.
Figure 2.
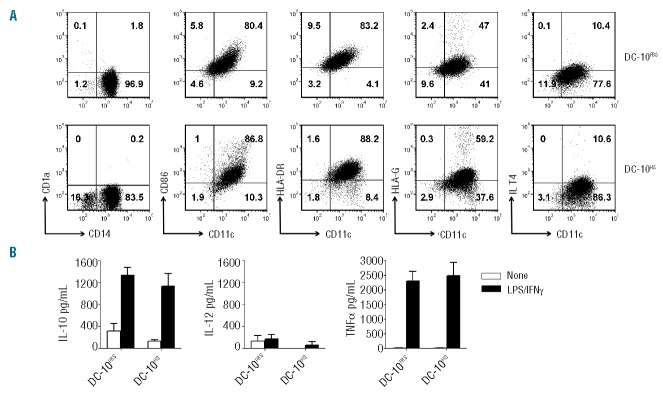
DC-10 differentiated with FBS or HS have comparable phenotypes and cytokine profiles. (A) Expression levels of CD14, CD1a, CD11c, CD86, HLA-DR, HLA-G, and ILT4 on DC-10 differentiated from CD14+ cells isolated from the same donor with FBS (DC-10FBS) or HS (DC-10HS) were evaluated by FACS. One representative donor out of nine tested. Numbers represent percentages of positive cells in each quadrant. (B) DC-10 were left unstimulated or stimulated with lipopolysaccharide (LPS) and interferon-γ (IFNγ) for 2 days. Concentration levels of IL-10, IL-12, and tumor necrosis factor-α (TNFα) in culture supernatants of the indicated cells are shown. Mean±SEM of six independent experiments.
Overall, DC-10 generated from different donors are mature CD14+ cells, with a high IL-10/IL-12 ratio, and express variable levels of membrane-bound HLA-G and ILT4.
DC-10 with high membrane-bound HLA-G promote T-cell anergy
Naïve CD4+ T cells stimulated with allogeneic HLA-Ghigh and HLA-Glow DC-10, at a 10:1 ratio, proliferated at significantly lower levels compared to T cells primed with mDC generated from the same donor. A reduction in proliferation of 91±7.4% (mean±SEM, n=5) and of 91.3±2.4% (mean±SEM, n=7) induced by HLA-Ghigh and HLA-Glow DC-10, respectively, was observed (Figure 3A and Online Supplementary Figure S1A). Accordingly, the levels of interferon-γ secreted by HLA-Ghigh or HLA-Glow DC-10-stimulated T cells were lower than those produced by mDC-primed T cells (Figure 3A). Similarly, a reduction in prolifetration of 82.5±5.2% (mean±SEM, n=8) and low interferon-γ production was observed when T cells were stimulated with DC-10HS (Figure 3B), confirming that culture conditions do not have an effect on the functional activity of DC-10.
Figure 3.
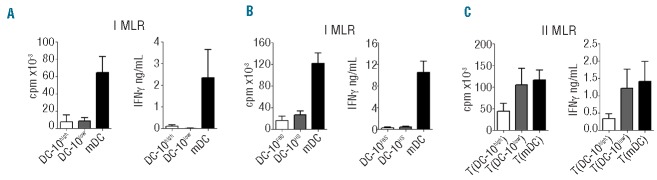
DC-10 with high levels of membrane-bound HLA-G promote T-cell anergy. (A) Naive CD4+ T cells were cultured with allogeneic in vitro-differentiated HLA-Ghigh DC-10, HLA-Glow DC-10, and mDC (ratio 10:1). Proliferative responses were evaluated 4 days after culture by [3H]-thymidine incorporation for an additional 16 h (left panel). Mean±SEM of five DC-10high, seven DC-10low, and 12 mDC independent experiments. Interferon-γ (IFNγ) production was evaluated 4 days after culture (right panel). Mean±SEM of four DC-10high, six DC-10low, and ten mDC independent experiments. (B) Naïve CD4+ T cells were cultured with allogeneic in vitro-differentiated DC-10FBS, and DC-10HS from CD14+ cells isolated from the same donor (ratio 10:1). Proliferative responses were evaluated 4 days after culture by [3H]-thymidine incorporation for an additional 16 h (left panel); IFNγ production was evaluated 4 days after culture (right panel). Mean±SEM of eight independent experiments. (C) Naïve CD4+ T cells were stimulated with allogeneic HLA-Ghigh DC-10 [T(DC-10high)], HLA-Glow DC-10 [T(DC-10low)], or mDC [T(mDC)] for 14 days. After culture, T cells were tested for their ability to proliferate in response to mDC from the same allogeneic donor used in priming. Proliferative responses were evaluated 2 days after culture by [3H]-thymidine incorporation for an additional 16 h (left panel). Mean±SEM of five [T(DC-10high)], seven [T(DC-10low)], and 12 [T(mDC)] independent experiments. IFNg production was evaluated 2 days after culture (right panel). Mean±SEM of five [T(DC-10high)], six [T(DC-10low)], and 11 [T(mDC)] independent experiments.
To investigate the function of T cells primed with HLA-Ghigh and HLA-Glow DC-10, secondary responses were evaluated. After one round of stimulation, T cells primed with HLA-Ghigh DC-10 [T(DC-10high) cells] were hypo-responsive to re-activation with mDC differentiated from the same donor used in primary stimulation, showing a reduction in proliferation of 61.9±18% (mean±SEM, n=5) compared to T cells primed with mDC [T(mDC) cells] (Figure 3C and Online Supplementary Figure S1B). Conversely, T cells obtained with HLA-Glow DC-10 [T(DC-10low) cells] showed high proliferative responses when re-activated with mDC with a limited reduction in proliferation (40.3±13.8%, mean±SEM, n=7; Figure 3C and Online Supplementary Figure S1B). Interferon-γ production by T(DC-10high) cells but not by T(DC-10low) cells re-challenged with mDC was also reduced compared to that secreted by T(mDC) cells (Figure 3C). These data show that high expression of membrane-bound HLA-G on DC-10 is required to promote T-cell anergy in CD4+ T cells.
DC-10 with high membrane-bound HLA-G induce Tr1 cell differentiation
T(DC-10high) cells contained a significantly higher frequency of IL-10-producing cells (on average 10%) compared to both T(DC-10low) cells (on average 3.4%, P=0.0121) and T(mDC) cells (on average 3.5%, P=0.0127) (Figure 4A). Moreover, in T(DC-10high) cells the proportion of interferon-γ-producing cells was on average 58.1%, and the frequency of IL-4-producing cells was very low (2.5±1.2%, mean±SEM, n=3; Figure 4A). Conversely, T(DC-10low) cells contained variable percentages of interferon-γ+ T cells (ranging from 0.3% to 60.8%), and a low frequency of IL-4+ T cells (1.9±0.93%, mean±SEM, n=8) (Figure 4A). The percentages of interferon-γ+ and IL-4+ in T(mDC) cells generated in parallel were comparable to those observed in T(DC-10high) and T(DC-10low) cells (Figure 4A).
Figure 4.
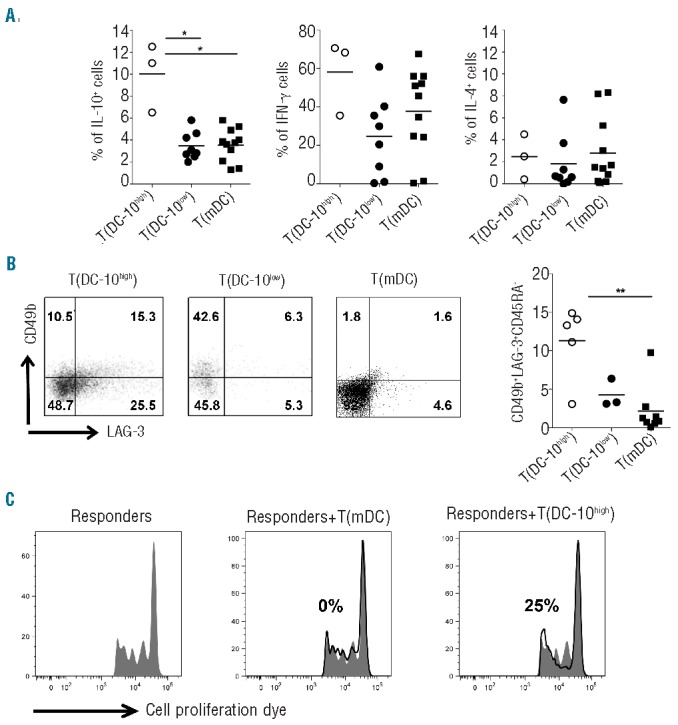
DC-10 with high membrane-bound HLA-G induce Tr1 cell differentiation. Naïve CD4+ T cells were stimulated with allogeneic HLA-Ghigh DC-10 [T(DC-10high)], HLA-Glow DC-10 [T(DC-10low)], or mDC [T(mDC)] for 14 days. (A) After culture, T cells were activated with PMA/IONO (leukocyte activation kit) for 6 h (as described in the Methods section), and cytokine production was determined by intracytoplasmic staining. Percentages of IL-10-, interferon-γ (IFNγ)-, and IL-4-producing cells in T(DC-10high) (n=3), T(DC-10low) (n=8), and T(mDC) (n=11) cells are presented. Each dot represents a single donor and lines indicate mean, *P<0.05. (B) After culture, the frequencies of CD4+ T cells co-expressing CD49b and LAG-3 in T(DC-10high), T(DC-10low), and T(mDC) cells were analyzed by FACS. One representative plot for each cell line (left panels) and percentages of CD49b+LAG-3+CD45RA− in five, three, and eight independent experiments for T(DC-10high), T(DC-10low), and T(mDC) cells, respectively, are shown (right panel). Numbers represent percentages of positive cells in each quadrant. Each dot represents a single donor and lines indicate mean, **P<0.01. (C) T(DC-10high) and T(mDC) cells were tested for their ability to suppress responses of autologous CD4+ T cells activated with mDC (Responders). Percentages of suppression mediated by the indicated T-cell populations are indicated. One representative donor out of three tested is shown.
Tr1 cells can be identified by the co-expression of CD49b and LAG-3.30 Thus, we evaluated the frequency of CD49b+LAG-3+CD45RA− cells in T(DC-10high) and T(DC-10low) cells. Results indicated that T(DC-10high) cells contained a higher frequency of CD49b+LAG-3+CD45RA-cells (on average 11.3%) compared to both T(DC-10low) cells (on average 4.3%) and T(mDC) cells (on average 2.2%, P=0.0031) (Figure 4B). In line with the presence of a high frequency of Tr1 cells, T(DC-10high) cells suppressed the proliferation of autologous CD4+ T cells activated with mDC (40.3±11.1%, mean±SEM, n=3), while T(DC-10low) cells did not (Figure 4C).These findings indicate that high levels of membrane-bound HLA-G improve the ability of DC-10 to promote IL-10-producing Tr1 cells.
HLA-Ghigh and HLA-Glow DC-10 secrete comparable levels of shed HLA-G1
To determine whether the low expression of membrane-bound HLA-G observed on HLA-Glow DC-10 could be ascribed to their ability to shed HLA-G1 from the membrane, we evaluated the amounts of shed HLA-G1 in culture supernatants. We detected shed HLA-G1 in four out of 16 unstimulated DC-10 (1.9±0.4 ng/mL, mean±SEM) and in nine out of 16 stimulated DC-10 (3±0.4 ng/mL, mean±SEM; Figure 5A). Conversely, soluble HLA-G5 was never detected in DC-10 culture supernatants. Interestingly, mDC generated in parallel secreted higher levels of shed HLA-G1 than DC-10 both in steady state and upon activation (P=0.04) (Figure 5A). No differences were observed in the ability of HLA-Ghigh and HLA-Glow DC-10 to release shed HLA-G1 (Figure 5B). Thus, in vitro-differentiated DC-10 secrete small amounts of shed HLA-G1, but not HLA-G5. Moreover, the low expression of membrane-bound HLA-G observed on HLA-Glow DC-10 is not associated with an increased release of shed HLA-G1.
Figure 5.
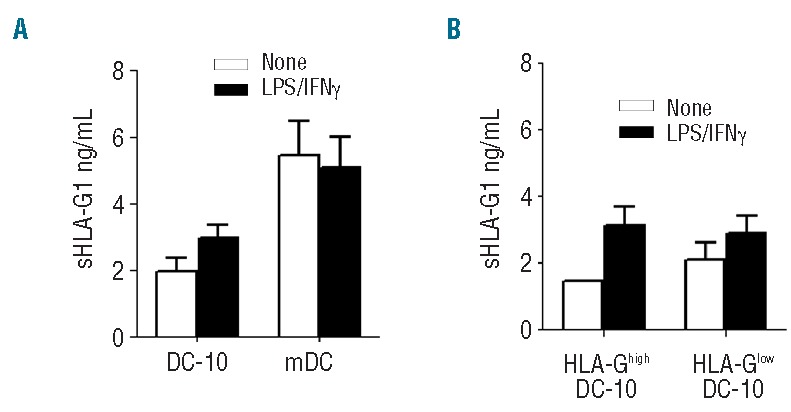
HLA-Ghigh and HLA-Glow DC-10 secrete comparable levels of shed HLA-G1. DC-10 and mDC were left unstimulated or stimulated with lipopolysaccharide (LPS) and interferon-γ (IFNγ) for 2 days. (A) Concentrations of shed HLA-G1 in culture supernatants of DC-10 and mDC are shown. Mean±SEM; n=4 (DC-10) and n=9 (mDC) unstimulated and n=8 (DC-10) and n=12 (mDC) stimulated with LPS and IFNg. (B) Concentration levels of shed HLA-G1 in culture supernatants of HLA-Ghigh and HLA-Glow DC-10 are shown. Mean±SEM; n=1 (HLA-Ghigh DC-10) and n=3 (HLA-Glow DC-10) unstimulated and n=3 (HLA-Ghigh DC-10) and n=6 (HLA-Glow DC-10) stimulated with LPS and IFNg.
Frequencies of 3′UTR HLA-G genotypes in donors with HLA-Ghigh or HLA-Glow DC-10
Polymorphisms at 3′UTR of the HLA-G locus influence its expression,31 thus we investigated whether variability in this region can be associated with membrane-bound HLA-G expression on DC-10. Donors from whom we differentiated DC-10 were typed at the 3′UTR of HLA-G locus and the presence of eight different polymorphic sites was inferred: the 14bp Ins/Del (rs1704), +3003 C/T (rs1707), +3010 C/G (rs1710), +3027 A/C (rs17179101), +3035 C/T (rs17179108), +3142 C/G (rs1063320), +3187 A/G (rs9380142) and +3196 C/G (rs1610696). We observed that the 14bp Ins allele was significantly highly represented both in a single dose (66%) and in double copy (45%) in donors with HLA-Glow DC-10 compared to those with HLA-Ghigh DC-10 (36%, P=0.0009 and 14%, P=0.0065, respectively; Figure 6A). Since 14bp Ins is in strong linkage disequilibrium with G in position +3142,32 we classified donors according to the presence of the 14bp Ins/Del and +3142 C/G polymorphisms as follows: InsG/InsG, DelC/DelC, DelC/InsG, and DelG/X. The relative frequencies of these alleles and genotypes are shown in Figure 6B. As expected, a high frequency of the InsG allele was observed in donors with HLA-Glow DC-10 and the DelC allele was significantly less represented in donors with HLA-Glow compared to those with HLA-Ghigh DC-10 (P=0.0213; Figure 6B).
Figure 6.
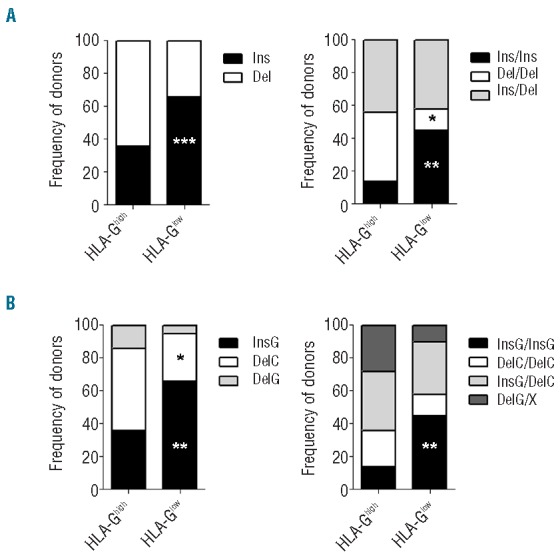
Frequencies of 3′UTR genotypes in donors with HLA-Ghigh and HLA-Glow DC-10. (A) Allele and genotype frequencies for 14 bp Ins/Del polymorphism, and for (B) 14 bp Ins/Del and +3142 G/C polymorphisms in 67 healthy donors (36 HLA-Ghigh DC-10 and 31 HLA-Glow DC-10) are shown. *P<0.05, **P<0.01, ***P<0.001.
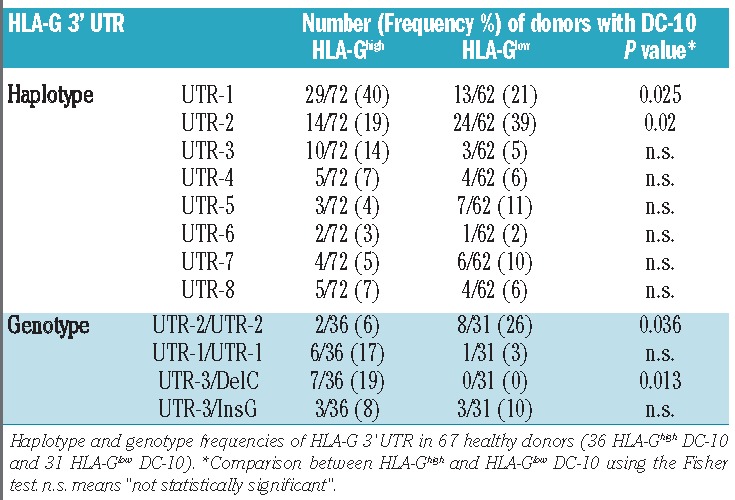
We next defined the frequencies of HLA-G 3′ UTR according to the classification of Castelli et al.,13 and we found that UTR-1 and UTR-2 were overall the most frequent haplotypes in our cohort of donors, UTR-1 being observed in 40% of donors with HLA-Ghigh DC-10, and UTR-2 in 39% of donors with HLA-Glow DC-10 (Table 1). Accordingly, UTR-2/UTR-2 was significantly more highly represented in donors with HLA-Glow DC-10 than in those with HLA-Ghigh DC-10 (P=0.036; Table 1). Moreover, 19% of donors with HLA-Ghigh DC-10 had UTR-3/DelC, whereas none of the donors with HLA-Glow DC-10 had this genotype (P=0.013; Table 1).
Table 1.
Most frequent HLA-G 3′ UTR alleles and genotypes in donors with HLA-Ghigh or HLA-Glow DC-10.
The presence of the 14bp Ins has been associated with low mRNA stability and reduced soluble HLA-G protein production.33 We, therefore, investigated whether the levels of mRNA for HLA-G were variable in DC-10 from different 3′UTR HLA-G typed donors. Overall the amount of HLA-G transcripts was very low, and in seven out of 13 samples resulted below the detection limit. Nevertheless, the highest amounts of HLA-G transcript were from donors with 14bp Ins/Del and 14bp Del/Del genotypes (Online Supplementary Figure S2).
These findings demonstrate that the levels of expression of membrane-bound HLA-G on DC-10 can be influenced by specific polymorphisms at the 3′UTR of the HLA-G locus, and that the UTR-2/UTR-2 genotype is more frequent among donors from whom DC-10 with low HLA-G were generated, while the UTR-3/DelC genotype was more frequent in donors with high HLA-G expression on DC-10.
MicroRNA-152 is highly expressed in in vitro-differentiated DC-10
We hypothesized that miR-152, miR-148a, and miR-148b, which target G at position +3142, may be responsible for the low membrane-bound HLA-G expression observed in DC-10 from donors carrying the UTR-2/UTR-2 genotype. We evaluated the expression of miR-152, miR-2110, and miR-93a predicted to have high binding affinity for HLA-G mRNA14. Results showed that miR-152 was expressed on average 2.1-fold more (n=5) in DC-10 than in CD14+ precursors, whereas miR-2110 and miR-93a were respectively equally or less expressed in DC-10 than in CD14+ cells (Figure 7). These data suggest that the expression of miR-152 could be involved in the post-transcriptional regulation of HLA-G in DC-10, especially in those donors with 3′UTR haplotypes with G in position +3142.
Figure 7.
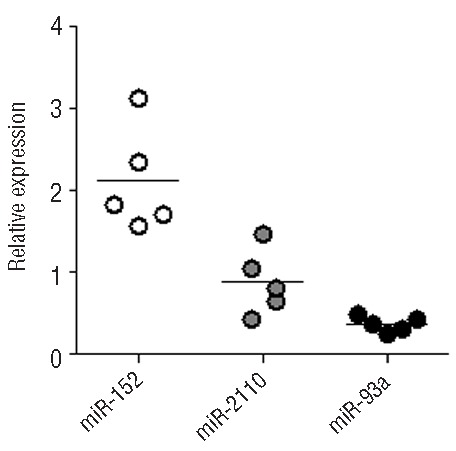
miR-152 is highly expressed by in vitro-differentiated DC-10. miRNA were extracted from in vitro-differentiated DC-10 and the expression of the indicated miRNA was evaluated by reverse transcription polymerase chain reaction. Following normalization to miR-let7a, relative mRNA amounts from DC-10 cells were adjusted to corresponding expression levels of a calibrator (corresponding CD14+ precursors). Numbers represent arbitrary units. Each dot represents a single cell preparation and lines indicate the mean.
Discussion
In the present study, we define the important role of HLA-G in DC-10-mediated induction of Tr1 cells. We show that the expression of membrane-bound HLA-G on DC-10 is donor-dependent and that, in the presence of similar levels of IL-10, a high expression of HLA-G on DC-10 is required to induce, at high frequency, Tr1 cells in vitro. We also established that 3′UTR genetic variations influence the HLA-G expression on DC-10 that might be finely tuned by miRNA-mediated post-transcriptional regulation.
Similar to other populations of tolerogenic DC previously described, G-CSF-induced DC34,35 and dermal CD141+ cells,36 we confirmed that DC-10 differentiated in vitro are CD14+. Importantly, we define that DC-10 expressed variable levels of HLA-G depending on the donor, and accordingly, DC-10 can be classified into HLA-Ghigh or HLA-Glow. We found that the expression of HLA-G on DC-10 correlated with that of ILT4, but not with the expression of ILT2 or of ILT3, which were highly expressed on DC-10 (data not shown). These findings are similar to those showing that in transfected antigen-presenting cell lines, the ectopic expression of HLA-G promotes the up-regulation of ILT2, ILT3, and ILT4.37
By proteolytic shedding, the HLA-G1 transmembrane can be released as shed HLA-G1, which retains all the functions of the membrane counterpart.6,38 We postulated that low levels of HLA-G on DC-10 may be associated with an increased production of shed HLA-G1. However, comparably low amounts of shed HLA-G1 were generated from both HLA-Glow and HLA-Ghigh DC-10. The production of shed HLA-G1 is regulated by the activity of matrix metalloproteinases,38 and specifically by matrix metalloproteinase-2.39 Data from the gene expression profile of in vitro-differentiated DC-10 indicated that matrix metalloproteinases are present in DC-10 (Comi et al., unpublished data). Thus, it may be hypothesized that in HLA-Glow DC-10 the great majority of HLA-G1 molecules are rapidly cleaved and released as shed HLA-G1, whereas the higher amounts of HLA-G1 synthesized in HLA-Ghigh DC-10 saturate the activity of matrix metalloproteinases and, as a result, more HLA-G1 is expressed as the transmembrane isoform. Future investigation will elucidate this possibility.
Constitutive high HLA-G expression confers to antigen-presenting cells the ability to inhibit primary T-cell responses.23 We herein show that DC-10 displayed low stimulatory activity regardless of the levels of HLA-G expression. This result is not surprising, since DC-10, independently of HLA-G expression, secrete high levels of IL-10, which directly inhibit T-cell responses in vitro.40–43 We have demonstrated that IL-10, either exogenously added or derived by DC-10, inhibited the proliferation of allogeneic naïve T cells in vitro.44,45
DC-10 promote Tr1 cell differentiation via the IL-10-induced HLA-G/ILT4 pathway.27 Although IL-10, ILT4, and HLA-G are important for DC-10 tolerogenic activity, their relative contributions in inducing Tr1 cells were not investigated. The identification of DC-10 that spontaneously expressed high or low HLA-G finally allowed us to demonstrate that the high levels of HLA-G on DC-10 are required for efficient induction of anergic T cells, and consequently Tr1 cells. Indeed, we proved that HLA-Ghigh DC-10 promoted Tr1 cells, identified as IL-10-producing or as CD49b+LAG-3+CD45RA− 30 T cells, at higher frequency than HLA-Glow DC-10. In addition, our data sustain the key role of IL-10 in DC-10-induced T-cell anergy (and Tr1 cells),27,43–45 since, in a limited number of donors, T cells primed with HLA-Glow DC-10 became hypo-responsive to secondary stimulation. Data presented in this study demonstrated that, although IL-10-derived by DC-10 is necessary to promote T-cell anergy, it is not sufficient to promote the differentiation of Tr1 cells at high frequency. The low HLA-G (and ILT4) expression on DC-10 hampers the amplification of the DC-10-mediated tolerogenic loop, consisting in inhibiting T-cell activation via ILT2 and promoting HLA-G and ILT4 expression on neighboring DC-10, and the consequent induction of IL-10, which sustains Tr1 cell generation. Since HLA-G expression on DC-10 is genetically determined, future studies aimed at up-regulating HLA-G in donors prone to generate HLA-Glow DC-10 could definitively prove our hypothesis.
Studies investigating 3′UTR HLA-G polymorphisms and HLA-G expression have primarily focused on the association between the 14bp Ins/Del polymorphism and soluble HLA-G isoforms.33,46 Only recently have these analyses been broadened to other polymorphisms in the 3′UTR region of HLA-G and to membrane-bound HLA-G.47,48 In the present study, we screened the association of multiple variations in the 3′UTR HLA-G, considering alleles, genotypes, haplotypes, and diplotypes, with membrane-bound HLA-G expression on DC-10. We found that the majority of donors with HLA-Glow DC-10 carried haplotypes, such as UTR-2, UTR-5, UTR-7, containing 14bp Ins and G in position +3142, both variations previously associated with low mRNA stability.10,14 These results are in line with a recent work showing that UTR-2 and UTR-5 are associated with low expression of soluble HLA-G.48
The presence of G at position +3142 increases the affinity of miR-152, miR-148a, and miR-148b for mRNA, promoting the repression of HLA-G expression.14 Moreover, the over-expression of miR-148a or miR-152 in JEG-3 or LCL721.221 cell lines promotes post-transcriptional down-regulation of membrane-bound HLA-G.49,50 Interestingly, analysis of the miRNome profile of DC-10 and its comparison with that of mDC identified ten differentially expressed miRNA, including miR-148a (Comi et al., unpublished data). Moreover, miR-152 was highly expressed in DC-10 as compared to CD14+ precursors. Thus, it can be postulated that post-transcriptional regulation of mRNA encoding for HLA-G may occur in DC-10 generated from donors carrying a double dose of 3′UTR haplotypes with +3142 G. The higher frequency of UTR-2, UTR-5, and UTR-7 in donors with HLA-Glow DC-10 supports this hypothesis. Although several questions regarding the mechanisms associated with polymorphic sites at 3′UTR of HLA-G need to be addressed, the present study evidences a link between the presence of 14bp Ins and the low expression of HLA-G, and indicates that this effect can be finely tuned by additional variations present at the 3′UTR, such as the +3142 C/G, possibly via miRNA-mediated post-transcriptional regulation.
In conclusion, this study provides new insights into the role of HLA-G in DC-10-mediated Tr1 cell induction: (i) high expression of HLA-G is required for efficient induction of Tr1 cells via DC-10; (ii) the contribution of soluble HLA-G isoforms in DC-10-mediated induction of Tr1 cells is negligible; and (iii) 3′UTR HLA-G haplotypes and genotypes can be associated with the expression of membrane-bound HLA-G on DC-10 and, consequently, with their ability to prime naïve T cells to become Tr1 cells. To our knowledge, this is the first comprehensive study in which 3′UTR polymorphisms have been associated with membrane-bound HLA-G expression and with tolerogenic properties of a specific cell subset. DC-10 are currently used to generate allospecific Tr1 cells for adoptive Treg-based cell therapy, but they represent an interesting therapeutic tool to induce or re-establish immunological tolerance in different clinical settings including allogeneic transplantation or autoimmune diseases. Results on the influence of 3′UTR genotypes on HLA-G expression on DC-10 and their tolerogenic activity provide new important tools for donor selection in several clinical settings such as hematopoietic stem cell and solid organ transplantation, and will also be relevant for understanding the immunological mechanisms underlying autoimmune diseases and cancer.
Acknowledgments
We thank Prof. E.A. Donadi and Dr. R. Sales de Albuquerque, from the Division of Clinical Immunology, Department of Medicine, School of Medicine of Ribeirão Preto, University of São Paulo, São Paulo, Brazil, for technical support with the molecular biology.
Footnotes
The online version of this article has a Supplementary Appendix.
Funding
This work was supported by Telethon Italy grant number TGT11E02, and by the Italian Ministry of Health, RF-2011-02346774, and a European grant for European cooperation in science and technology (Action to Focus and Accelerate Cell-based Tolerance-inducing Therapies).
Dr. Giada Amodio conducted this study as partial fulfillment of her PhD in Molecular Medicine, Program in Basic and Applied Immunology, San Raffaele University, Milan, Italy.
Authorship and Disclosures
Information on authorship, contributions, and financial & other disclosures was provided by the authors and is available with the online version of this article at www.haematologica.org.
References
- 1.Kovats S, Main EK, Librach C, Stubblebine M, Fisher SJ, DeMars R. A class I antigen, HLA-G, expressed in human trophoblasts. Science. 1990;248(4952):220–223. [DOI] [PubMed] [Google Scholar]
- 2.Rouas-Freiss N, Goncalves RM, Menier C, Dausset J, Carosella ED. Direct evidence to support the role of HLA-G in protecting the fetus from maternal uterine natural killer cytolysis. Proc Natl Acad Sci USA. 1997;94(21):11520–11525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Curigliano G, Criscitiello C, Gelao L, Goldhirsch A. Molecular pathways: human leukocyte antigen G (HLA-G). Clin Cancer Res. 2013;19(20):5564–5571. [DOI] [PubMed] [Google Scholar]
- 4.Fujii T, Ishitani A, Geraghty DE. A soluble form of the HLA-G antigen is encoded by a messenger ribonucleic acid containing intron 4. J Immunol. 1994;153(12):5516–5524. [PubMed] [Google Scholar]
- 5.Ishitani A, Geraghty DE. Alternative splicing of HLA-G transcripts yields proteins with primary structures resembling both class I and class II antigens. Proc Natl Acad Sci USA. 1992;89(9):3947–3951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Park GM, Lee S, Park B, et al. Soluble HLA-G generated by proteolytic shedding inhibits NK-mediated cell lysis. Biochem Biophys Res Commun. 2004;313(3):606–611. [DOI] [PubMed] [Google Scholar]
- 7.Carosella ED. The tolerogenic molecule HLA-G. Immunol Lett. 2011;138(1):22–24. [DOI] [PubMed] [Google Scholar]
- 8.Rousseau P, Le Discorde M, Mouillot G, Marcou C, Carosella ED, Moreau P. The 14 bp deletion-insertion polymorphism in the 3′ UT region of the HLA-G gene influences HLA-G mRNA stability. Hum Immunol. 2003;64(11):1005–1010. [DOI] [PubMed] [Google Scholar]
- 9.Rebmann V, van der Ven K, Passler M, Pfeiffer K, Krebs D, Grosse-Wilde H. Association of soluble HLA-G plasma levels with HLA-G alleles. Tissue Antigens. 2001;57(1):15–21. [DOI] [PubMed] [Google Scholar]
- 10.Hviid TV, Hylenius S, Rorbye C, Nielsen LG. HLA-G allelic variants are associated with differences in the HLA-G mRNA isoform profile and HLA-G mRNA levels. Immunogenetics. 2003;55(2):63–79. [DOI] [PubMed] [Google Scholar]
- 11.Tan Z, Randall G, Fan J, et al. Allele-specific targeting of microRNAs to HLA-G and risk of asthma. Am J Hum Genet. 2007;81(4):829–834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Veit TD, Chies JA. Tolerance versus immune response – microRNAs as important elements in the regulation of the HLA-G gene expression. Transpl Immunol. 2009;20(4):229–231. [DOI] [PubMed] [Google Scholar]
- 13.Castelli EC, Mendes-Junior CT, Deghaide NH, et al. The genetic structure of 3′ untranslated region of the HLA-G gene: polymorphisms and haplotypes. Genes Immun. 2010;11(2):134–141. [DOI] [PubMed] [Google Scholar]
- 14.Castelli EC, Moreau P, Oya e Chiromatzo A, et al. In silico analysis of microRNAS targeting the HLA-G 3′ untranslated region alleles and haplotypes. Hum Immunol. 2009;70(12):1020–1025. [DOI] [PubMed] [Google Scholar]
- 15.Yie SM, Li LH, Xiao R, Librach CL. A single base-pair mutation in the 3′-untranslated region of HLA-G mRNA is associated with pre-eclampsia. Mol Hum Reprod. 2008;14(11):649–653. [DOI] [PubMed] [Google Scholar]
- 16.Lucena-Silva N, Monteiro AR, de Albuquerque RS, et al. Haplotype frequencies based on eight polymorphic sites at the 3′ untranslated region of the HLA-G gene in individuals from two different geographical regions of Brazil. Tissue Antigens. 2012;79(4):272–278. [DOI] [PubMed] [Google Scholar]
- 17.Larsen MH, Hylenius S, Andersen AM, Hviid TV. The 3′-untranslated region of the HLA-G gene in relation to pre-eclampsia: revisited. Tissue Antigens. 2010;75(3):253–261. [DOI] [PubMed] [Google Scholar]
- 18.Alvarez M, Piedade J, Balseiro S, Ribas G, Regateiro F. HLA-G 3′-UTR SNP and 14-bp deletion polymorphisms in Portuguese and Guinea-Bissau populations. Int J Immunogenet. 2009;36(6):361–366. [DOI] [PubMed] [Google Scholar]
- 19.Shiroishi M, Kuroki K, Rasubala L, et al. Structural basis for recognition of the non-classical MHC molecule HLA-G by the leukocyte Ig-like receptor B2 (LILRB2/LIR2/ILT4/CD85d). Proc Natl Acad Sci USA. 2006;103(44):16412–16417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Riteau B, Menier C, Khalil-Daher I, et al. HLA-G1 co-expression boosts the HLA class I-mediated NK lysis inhibition. Int Immunol. 2001;13(2):193–201. [DOI] [PubMed] [Google Scholar]
- 21.Le Gal FA, Riteau B, Sedlik C, et al. HLA-G-mediated inhibition of antigen-specific cytotoxic T lymphocytes. Int Immunol. 1999;11(8):1351–1356. [DOI] [PubMed] [Google Scholar]
- 22.Naji A, Menier C, Morandi F, et al. Binding of HLA-G to ITIM-bearing Ig-like transcript 2 receptor suppresses B cell responses. J Immunol. 2014;192(4):1536–1546. [DOI] [PubMed] [Google Scholar]
- 23.LeMaoult J, Krawice-Radanne I, Dausset J, Carosella ED. HLA-G1-expressing antigen-presenting cells induce immunosuppressive CD4+ T cells. Proc Natl Acad Sci USA. 2004;101(18):7064–7069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Horuzsko A, Lenfant F, Munn DH, Mellor AL. Maturation of antigen-presenting cells is compromised in HLA-G transgenic mice. Int Immunol. 2001;13(3):385–394. [DOI] [PubMed] [Google Scholar]
- 25.Naji A, Le Rond S, Durrbach A, et al. CD3+CD4low and CD3+CD8low are induced by HLA-G: novel human peripheral blood suppressor T-cell subsets involved in transplant acceptance. Blood. 2007;110(12):3936–3948. [DOI] [PubMed] [Google Scholar]
- 26.Gregori S, Magnani CF, Roncarolo MG. Role of human leukocyte antigen-G in the induction of adaptive type 1 regulatory T cells. Hum Immunol. 2009;70(12):966–969. [DOI] [PubMed] [Google Scholar]
- 27.Gregori S, Tomasoni D, Pacciani V, et al. Differentiation of type 1 T regulatory cells (Tr1) by tolerogenic DC-10 requires the IL-10-dependent ILT4/HLA-G pathway. Blood. 2010;116(6):935–944. [DOI] [PubMed] [Google Scholar]
- 28.Amodio G, Mugione A, Sanchez AM, et al. HLA-G expressing DC-10 and CD4(+) T cells accumulate in human decidua during pregnancy. Hum Immunol. 2013;74(4):406–411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Locafaro G, Amodio G, Tomasoni D, Tresoldi C, Ciceri F, Gregori S. HLA-G expression on blasts and tolerogenic cells in patients affected by acute myeloid leukemia. J Immunol Res. 2014;2014:636292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Gagliani N, Magnani CF, Huber S, et al. Coexpression of CD49b and LAG-3 identifies human and mouse T regulatory type 1 cells. Nat Med. 2013;19(6):739–746. [DOI] [PubMed] [Google Scholar]
- 31.Donadi EA, Castelli EC, Arnaiz-Villena A, Roger M, Rey D, Moreau P. Implications of the polymorphism of HLA-G on its function, regulation, evolution and disease association. Cell Mol Life Sci. 2011;68(3):369–395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Castelli EC, Mendes-Junior CT, Veiga-Castelli LC, Roger M, Moreau P, Donadi EA. A comprehensive study of polymorphic sites along the HLA-G gene: implication for gene regulation and evolution. Mol Biol Evol. 2011;28(11):3069–3086. [DOI] [PubMed] [Google Scholar]
- 33.Hviid TV, Rizzo R, Christiansen OB, Melchiorri L, Lindhard A, Baricordi OR. HLA-G and IL-10 in serum in relation to HLA-G genotype and polymorphisms. Immunogenetics. 2004;56(3):135–141. [DOI] [PubMed] [Google Scholar]
- 34.Rutella S, Bonanno G, Pierelli L, et al. Granulocyte colony-stimulating factor promotes the generation of regulatory DC through induction of IL-10 and IFN-alpha. Eur J Immunol. 2004;34(5):1291–1302. [DOI] [PubMed] [Google Scholar]
- 35.Rossetti M, Gregori S, Roncarolo MG. Granulocyte-colony stimulating factor drives the in vitro differentiation of human dendritic cells that induce anergy in naive T cells. Eur J Immunol. 2010;40(11):3097–3106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Chu CC, Ali N, Karagiannis P, et al. Resident CD141 (BDCA3)+ dendritic cells in human skin produce IL-10 and induce regulatory T cells that suppress skin inflammation. J Exp Med. 2012;209(5):935–945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.LeMaoult J, Zafaranloo K, Le Danff C, Carosella ED. HLA-G up-regulates ILT2, ILT3, ILT4, and KIR2DL4 in antigen presenting cells, NK cells, and T cells. FASEB J. 2005;19(6):662–664. [DOI] [PubMed] [Google Scholar]
- 38.Dong Y, Lieskovska J, Kedrin D, Porcelli S, Mandelboim O, Bushkin Y. Soluble non-classical HLA generated by the metalloproteinase pathway. Hum Immunol. 2003;64(8):802–810. [DOI] [PubMed] [Google Scholar]
- 39.Rizzo R, Trentini A, Bortolotti D, et al. Matrix metalloproteinase-2 (MMP-2) generates soluble HLA-G1 by cell surface proteolytic shedding. Mol Cell Biochem. 2013;381(1–2):243–255. [DOI] [PubMed] [Google Scholar]
- 40.Taga K, Mostowski H, Tosato G. Human interleukin-10 can directly inhibit T-cell growth. Blood. 1993;81(11):2964–2971. [PubMed] [Google Scholar]
- 41.de Waal Malefyt R, Yssel H, de Vries JE. Direct effects of IL-10 on subsets of human CD4+ T cell clones and resting T cells. Specific inhibition of IL-2 production and proliferation. J Immunol. 1993;150(11):4754–4765. [PubMed] [Google Scholar]
- 42.Groux H, Bigler M, de Vries JE, Roncarolo MG. Interleukin-10 induces a long-term antigen-specific anergic state in human CD4+ T cells. J Exp Med. 1996;184(1):19–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Groux H, O’Garra A, Bigler M, et al. A CD4+ T-cell subset inhibits antigen-specific T-cell responses and prevents colitis. Nature. 1997;389(6652):737–742. [DOI] [PubMed] [Google Scholar]
- 44.Bacchetta R, Gregori S, Serafini G, et al. Molecular and functional characterization of allogantigen-specific anergic T cells suitable for cell therapy. Haematologica. 2010;95(12):2134–2143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Gregori S, Roncarolo MG, Bacchetta R. Methods for in vitro generation of human type 1 regulatory T cells. Methods Mol Biol. 2011;677:31–46. [DOI] [PubMed] [Google Scholar]
- 46.Rizzo R, Hviid TV, Stignani M, et al. The HLA-G genotype is associated with IL-10 levels in activated PBMCs. Immunogenetics. 2005;57(3–4):172–181. [DOI] [PubMed] [Google Scholar]
- 47.Rizzo R, Audrito V, Vacca P, et al. HLA-G is a component of the CLL escape repertoire to generate immune suppression: impact of HLA-G 14 bp (rs66554220) polymorphism. Haematologica. 2014;99(5):888–896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Martelli-Palomino G, Pancotto JA, Muniz YC, et al. Polymorphic sites at the 3′ untranslated region of the HLA-G gene are associated with differential HLA-G soluble levels in the Brazilian and French population. PloS One. 2013;8(10):e71742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Zhu XM, Han T, Wang XH, et al. Overexpression of miR-152 leads to reduced expression of human leukocyte antigen-G and increased natural killer cell mediated cytolysis in JEG-3 cells. Am J Obstet Gynecol. 2010;202(6):592.e1–7. [DOI] [PubMed] [Google Scholar]
- 50.Manaster I, Goldman-Wohl D, Greenfield C, et al. MiRNA-mediated control of HLA-G expression and function. PloS One. 2012;7(3):e33395. [DOI] [PMC free article] [PubMed] [Google Scholar]