

Abstract
While small-molecule kinase inhibitors became the most prominent anticancer drugs, novel combinatorial strategies need to be developed as the fight against cancer is not yet won. We review emerging literature showing that the release of several ectokinases is significantly upregulated in body fluids from cancer patients and that they leave behind their unique signatures on extracellular matrix (ECM) proteins. Our analysis of proteomic data reveals that fibronectin is heavily phosphorylated in cancer tissues particularly within its growth factor binding sites and on domains that regulate fibrillogenesis. We are thus making the case that cancer is not only a disease of cells but also of the ECM. Targeting extracellular kinases or the extracellular signatures they leave behind might thus create novel opportunities in cancer diagnosis as well as new avenues to interfere with cancer progression and malignancy.
Keywords: Cancer marker, drug design, ectokinases, exokinases, extracellular matrix, extracellular phosphorylation, extracellular protein kinase, personalized medicine
Introduction
Since the fight against cancer is far from being won, there is a need to think of new strategies to identify alternative targets for cancer diagnosis and combinatorial therapies. Current challenges include the desire to detect cancer much earlier, to prevent or reduce the emergence of acquired drug resistance 1, and to reduce the often lethal side effects. Even more challenging is the fact that different cancer cells from the same tumor can use different pathways to achieve drug resistance 2. The complexity of pathways that can lead to drug resistance prevents to predict which treatment modality might finally allow the host rather the cancer to survive 3,4. Continued chemotherapy will target only a subset of cancer cells, while the resistant cells continue to grow 2. New strategies are therefore needed to target nonresistant and resistant cancer cells. Protein phosphorylation is the key regulatory posttranslational modification exploited for intracellular signaling 5–7, and kinases require sufficiently high ATP levels to transfer a phosphate group. Today, it is believed that one third of human proteins are phosphorylated 8 and small-molecule kinase inhibitors have thus taken the lead as next generation cancer drugs (Table1) 9. While this is a significant progress, these inhibitors often interfere with other complex intracellular signaling networks thus causing sometimes severe side effects, and need to be combined with other approaches.
Table 1.
Small-molecule kinase inhibitors on the market against kinases
| Name | Trade name | Targeted tyrosine kinase | Disease | Producer |
|---|---|---|---|---|
| Imatinib | Gleevec, Glivec | BCR-Abl | Chronic myelogenous leukemia (CML), gastrointestinal stromal tumors (GISTs), number of other malignancies | Novartis |
| Gefitinib | Iressa | EGFR | Breast, lung, other cancers | AstraZeneca, Teva |
| Erlotinib | Tarceva | EGFR | Nonsmall cell lung cancer (NSCLC), pancreatic cancer, several other types of cancer | Genentech, OSI Pharmaceuticals, Roche |
| Crizotinib | Xalkori | ALK | Nonsmall cell lung cancer (NSCLC) | Pfizer |
| Dasatinib | Sprycel | BCR/Abl and Src family | Chronic myelogenous leukemia (CML), Philadelphia chromosome-positive acute lymphoblastic leukemia (Ph+ ALL) | Bristol-Myers Squibb |
| Lapatinib | Tykerb/Tyverb | HER2 and EGFR | Breast cancer, other solid tumors | GlaxoSmithKline |
| Nilotinib | Tasigna | BCR-ABL, KIT, LCK, EPHA3, EPHA8, DDR1, DDR2, PDGFRB, MAPK11, and ZAK | Chronic myelogenous leukemia | Novartis |
| Pazopanib | Votrient | c-KIT, FGFR, PDGFR, and VEGFR | Renal cell carcinoma, soft tissue sarcoma | GlaxoSmithKline |
| Sunitinib | Sutent | PDGF-Rs, VEGFRs, KIT | Renal cell carcinoma (RCC), gastrointestinal stromal tumor | Pfizer |
| Sorafenib | Nexavar | VEGFR, PDGFR, Raf | Renal cell carcinoma (RCC), unresectable hepatocellular carcinomas (HCC), thyroid cancer | Bayer, Onyx Pharmaceuticals |
| Vandetanib | Caprelsa | VEGFR, EGFR, RET-tyrosine kinase | Tumors of the thyroid gland | AstraZeneca |
| Tofacitinib | Xeljanz, Jakvinus | JAK | Rheumatoid arthritis | Pfizer |
| Ruxolitinib | Jakafi, Jakavi | JAK | Myelofibrosis | Incyte Pharmaceuticals, Novartis |
Current FDA-approved kinase inhibitors on the market in cancer treatment.
Cells secrete a cocktail of enzymes, such as cholinesterases, peptidases, transpeptidases, nucleotidases, phosphodiesterases, ectokinases, and ectophosphatases, which lead to posttranslational modifications of extracellular matrix (ECM) proteins, and the composition of this cocktail depends on cell type, external stimulations, and disease 10. Posttranslational modifications of ECM proteins can affect outside-in cell signaling and consequently cell behavior 11. The massive killing of cancer cells typically increases the local extracellular concentrations of the cytoplasmic content, including ATP, thereby causing additional posttranslational modifications of the ECM. The killing of cancer cells will thus leave behind a “diseased” ECM that can send altered instructive signals to the cells that later invade this cancerous ECM left behind. This has not been considered in the treatment of cancer previously.
Beyond using the concentration of extracellular protein kinases in blood to detect cancer in early stages 12–14, ectokinases and ectophosphatases might serve as new drug targets. Shielded by the plasma membrane, drugs with extracellular targets might cause less side effects as they can less directly interfere with intracellular signaling 15–21. Even though cancer is not only a disease of cells but also leads to posttranslational modifications of the ECM, the intracellular focus has overshadowed potential extracellular opportunities that could be exploited to address some of these challenges. Here, we thus review the indications that cancer is not only a disease of cells but also of the ECM, and how this newly emerging knowledge of extracellular posttranslational modifications can potentially be exploited for cancer diagnosis and treatment.
Extracellular Enzymes and Posttranslational Modifications of ECM Coregulate Cancer Progression
Extracellular strategies are mostly missing although considerable knowledge emerged that the composition and rigidity of the ECM, and consequently ECM cell signaling plays an important role in cancer progression 22,23. The first wave of targeting ECM enzymes was motivated by the finding that cancer tissues show upregulated matrix metalloproteinase (MMP) levels, and it was thought that MMP-induced cleavage of ECM would promote the escape of cancer cells from the site of tumors 24,25 (Fig.1A). Consequently, MMP inhibitors were designed and went into clinical trials, but with devastating negative outcomes 26,27. The main reason for the failure was the lack of appreciation for the complexity of MMP functions and their respective effects on ECM properties and signaling. Only three MMPs had been described at the time when the clinical trials had started, while 23 different MMPs are known today 28. They were found in different cell types with diverse functions including ECM–cell interactions, cell–cell contact, and regulation of soluble factors among many others 27. Broad-spectrum inhibitions of MMPs thus interfere with their diverse regulatory roles and thereby cause major side effects 29.
Figure 1.
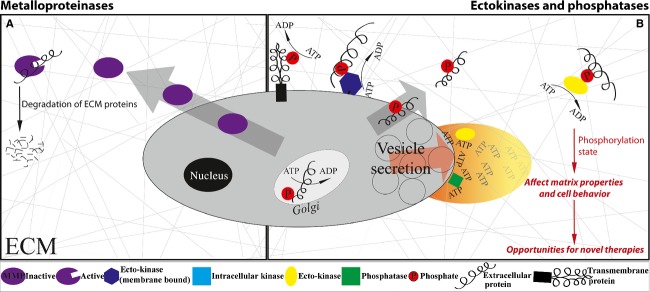
Extracellular enzymes in cancer. (A) To alter the extracellular matrix (ECM) biology of cancer, matrix metalloproteinases (MMPs) served as first extracellular drug targets. The hypothesis was that matrix metalloproteinases (MMP) inhibition can prevent the local degradation of ECM and thus the escape of metastatic stroma cells. (B) Vesicle secretion and cell necrosis transiently releases intracellular content high in kinases and ATP into the surrounding tissue as indicated by the graded plume. For a short time period, the ATP levels are sufficiently high for the kinases to be active in extracellular space leaving behind phosphorylated ECM. Ectokinases and phosphatases thus provide unique opportunities as novel extracellular drug targets.
It is thus timely to consider alternate extracellular strategies, including extracellular enzymes or other means by which to regulate posttranslational modifications (Fig.1B). While the importance of various posttranslational modifications in the ECM are known to regulate cancer progression 23, the significance of ectokinases and ectophosphatases, and the signatures they leave behind, is only now at the verge of being recognized 30. Why should we even consider extracellular phosphorylation since the ATP levels are typically low in extracellular environment? Extracellular ATP can transiently increase to levels that are sufficiently high to activate ectokinases in those tissues that undergo major necrosis and apoptosis, thereby releasing intracellular content 30. Also ATP secretion pathways are significantly upregulated in cancers 31,32 and increased levels of extracellular ATP could recently been measured at tumor sites 33.
Among the reported ectokinases, the most prominent ones are the casein kinase II (CKII) 34, protein kinase A (PKA) 35, protein kinase C (PKC) 36, and the recently reported Fam20C kinase 37. Several ectophosphatases including alkaline phosphatase 38, tartrate-resistant acid phosphatase (TRAP) 39, and the most recently reported PTEN phosphatase 40 have been reported in the ECM. Interestingly, the concentration of the extracellular alkaline phosphatase is already measured routinely as a disease marker in patient's blood samples to detect liver diseases, bone disorders, or cancer and the TRAP is being discussed as a good candidate 39,41.
Taken together, considerable evidence is emerging that posttranslational modifications of ECM coregulate cancer progression, that ectokinases and ectophosphatases are found in body fluids of cancer patients, and that kinases can be transiently active in extracellular space in regions where necrosis or other factors cause the release of ATP. As with any discovery, new ideas and strategies are thus beginning to emerge how to exploit these emerging insights into early cancer detection and therapy.
Striking Signatures of Extracellular Kinase Activity Are Found in Cancer Tissues
Postulating that massive necrosis might temporarily upregulate ectokinase activity in extracellular space, we recently mined published proteomic data and found a significant upregulation of phosphorylated residues in tissue samples from cancer patients 30. This included the phosphorylation of ECM proteins, as well as of cell surface and extracellular domains of transmembrane proteins. Screening more than 60 different extracellular proteins revealed that nearly all can occur in phosphorylated states 30. Most compelling was the finding that the integrin subunits α4 and β1, two key players in cancer progression and signaling, were found in tissue samples to be phosphorylated in their extracellular domains 30,42–44. Since fibronectin 45–47 which is a key component of the ECM is known to be highly upregulated in cancer 48–53, we further analyzed published proteomic data and found that fibronectin is indeed heavily phosphorylated in clinical cancer tissue samples (Fig.2, Table2). Heavily phosphorylated regions in fibronectin include and are associated with growth factor binding sites (FnIII4, FnIII13-14) and with domains that regulate fibronectin fibrillogenesis. This is an important finding since growth factor signaling and ECM fibrillogenesis are essential regulators in cancer malignancy and progression 22. In addition to fibronectin, elevated levels of phosphorylated fibrinogen A are found in the plasma from patients with stage III or IV ovarian cancer compared to healthy controls 54.
Figure 2.

Experimentally verified phosphorylation sites on fibronectin. Schematic representation of plasma fibronectin with modules type I (gray), type II (turquoise), and type III (orange). (A) Locations of various bacterial and cell binding sites on the fibronectin monomer. (B) Experimentally identified phosphorylation sites by mass spectrometry techniques as retrieved from protein data banks Phosida, PhosphoSitePlus, PhosphoNet, HPRD, dbPTM, and UniProt for human Fn (P02751). (C) Locations of protein binding sites on the fibronectin monomer with a special focus on matrix metalloproteinases (MMPs).
Table 2.
Experimentally verified phosphorylation sites on fibronectin in cancer samples
| Residue (P02751) | Location/binding sites | Reference/databases | Cancer tissues/cells |
|---|---|---|---|
| Y101, Y106, T136 | FnI2, Fn–Fn, Heparin, Tenascin, Fibrin | PhosphositePlus, PhosphoNet | In seven patients samples (Y101): ovarian, liver, lung, esophageal, gastric |
| In one patient sample (Y106): ovarian | |||
| In one patient sample (T136): T-cell leukemia | |||
| Y372 | FnII1, Collagen, Gelatin | PhosphositePlus, PhosphoNet | In three patients samples: ovarian, liver, hepatocellular carcinoma, hepatocyte–liver |
| Y588 | FnI9, Collagen, Gelatin | PhosphositePlus, PhosphoNet | In one patients sample: lung carcinoma |
| Y641 | FnIII1, Fn–Fn | PhosphositePlus, PhosphoNet | In one patients sample: pancreatic carcinoma |
| S904 | Linker FnIII3–FnIII4 | Phosida, PhosphositePlus, PhosphoNet, HPRD,dbPTM 79 | HeLaS3 (cervical cancer) |
| S909 | FnIII4, DNA binding | HPRD, dbPTM 80 | Hela cells |
| Y937, T960, S968, T972 | FnIII4, DNA binding | PhosphositePlus, PhosphoNet | In one patients sample (Y037): gastric |
| In one patients sample (T960): T-cell leukemia | |||
| In one patients sample (S968): T-cell leukemia | |||
| In one patients sample (T972): T-cell leukemia | |||
| Y1042 | FnIII5 | PhosphositePlus, PhosphoNet, dbPTM 81 | Embryonic stem cells |
| Y1206 | FnIII7 | PhosphositePlus, PhosphoNet | In two patients samples: ovarian |
| T1271 | FnIII8, Cell binding region | PhosphositePlus, PhosphoNet | In one patients sample: colorectal |
| T1462 | FnIII10, Cell binding region | PhosphositePlus 82 | 293 (epithelial) |
| T1743, T1762, T1786, S1833, T1840, T1842, T1855, T1860, Y1879, Y1884 | FnIII13, Heparin, Syndecan-4 | PhosphositePlus, PhosphoNet | In one patients sample (T1743): T-cell leukemia |
| In one patients sample (T1762): esophageal | |||
| In one patients sample (T1786): esophageal | |||
| In one patients sample (S1833): liver, cholangiocellular carcinoma | |||
| In two patients samples (1840): cervical | |||
| In one patients sample (T1842): cervical | |||
| In two patients samples (T1855): cervical | |||
| In one patients sample (T1860): cervical | |||
| In one patients sample (Y1879): ovarian | |||
| In two patients samples (Y1884): ovarian, T-cell leukemia | |||
| S2007 | Variable region IIICS, LDV, REDV integrin binding sites | Phosida 83 | Hela cells |
| S2131, S2139 | FnIII15, Cryptic cysteine | 84 | U266 (immortal B lymphocytes derived from multiple myeloma) |
| S2174 | FnIII15, Cryptic cysteine | Phosida, 83,85,86 | Hela cells, HEK, human liver tissue |
| S2182, S2209 | FnIII15, Cryptic cysteine | Phosida 83 | Hela cells |
| S2251 | FnI10, Fibrin binding | 84 | U266 (immortal B lymphocytes derived from multiple myeloma) |
| Y2258 | FnI11 | PhosphositePlus, PhosphoNet | In one patient sample: ovarian |
| S2259, S2285, S2293 | FnI11, Fibrin binding, Protein-disulfide isomerase binding | 84 | U266 (immortal B lymphocytes derived from multiple myeloma) |
| S2294 | FnI12, Fibrin binding, Protein-disulfide isomerase binding | Phosida, 83,85,86 | Hela cells, HEK, human liver tissue |
| S2318 | FnI12, Fibrin binding, Protein-disulfide isomerase binding | dbPTM 84 | U266 (immortal B lymphocytes derived from multiple myeloma) |
| S2328 | FnI12, Fibrin binding, Protein-disulfide isomerase binding | Phosida, 83,85,86 | Hela cells, HEK, Human liver tissue |
| S2341, S2349 | C-terminus, Disulfide bonds for Fn assembly | dbPTM, 84 | Serum |
| Y2350 | C-terminus, Disulfide bonds for Fn assembly | PhosphositePlus, PhosphoNet | In two patients samples: breast, ovarian |
| Y2353 | C-terminus, Disulfide bonds for Fn assembly | PhosphoNet, PhosphositePlus 87 | In 12 patient samples, breast, lung, gastric, liver, hepatocellular carcinoma |
| S2353 | C-terminus, Disulfide bonds for Fn assembly | Phosida, 83,85,86 | Hela cells, HEK, human liver tissue |
| S2354 | C-terminus | PhosphositePlus, PhosphoNet | In one patient sample: ovarian cancer |
| S2376 | C-terminus, Disulfide bonds for Fn assembly | 84 | U266 (immortal B lymphocytes derived from multiple myeloma) |
| S2384 | C-terminus, Disulfide bonds for Fn assembly | Phosida, PhosphoSitePlus, PhosphoNet, dbPTM, UniProt, 83,84,86,88–90 | In 14 patients samples: breast, skin, liver, hepatocellular carcinoma, and surrounding tissue, blood plasma U266 (immortal B lymphocytes derived from multiple myeloma), Hela cells |
| S2419 | C-terminus, Disulfide bonds for Fn assembly | 83,85,86 | Hela cells, HEK, human liver tissue |
| S2432, S2440 | C-terminus, Disulfide bonds for Fn assembly | HPRD 84 | U266 (immortal B lymphocytes derived from multiple myeloma) |
| S2444 | C-terminus, Disulfide bonds for Fn assembly | Phosida 83 | Hela cells |
| S2475 | C-terminus, Disulfide bonds for Fn assembly | HPRD 83,85,86 | Hela cells, U266 (immortal B lymphocytes derived from multiple myeloma) |
Phosphorylated sites by mass spectrometry retrieved from protein databases. Due to lack of track changes and updates of the databases, the reported sites here may differ from the database entries at later points. Table as of Nov. 2014.
Taken together, available data suggest that the upregulated phosphorylation of fibronectin and of some other extracellular proteins is a distinct signature of cancerous ECM. The phosphorylation of the ECM caused by the transient release of ATP by dying cells might thus be physiologically far more important in regulating cancer cell differentiation and tumor progression than previously thought. Indeed, the phosphorylation ratio of peptides increase with tumor size as has been previously shown 13. Any discovery of new signatures how cancer or cancer tissues are different from the norm might offer valuable entrance points for novel diagnostic or therapeutic strategies. Furthermore, extracellular proteins that are highly phosphorylated in some but not in other cancer types might be suitable as novel markers for the early detection of cancers, or perhaps serve as signature of its malignancy.
New Strategies for Cancer Diagnostics: Quantification of the Concentrations and Activities of Extracellular Protein Kinases
One major challenge is to detect cancer in earlier stages in order to treat patients more successfully. According to recent cancer statistics, the 5-year survival rate dramatically drops if cancer is detected at a late stage 55. Most of the current serum tumor markers are based on the antigen determination method, including CEA, AFP, hCG, PSA, and CA125, but lack tumor specificity and often cannot be used in early cancer screening and diagnosis 56–63. To overcome these shortcomings, novel, cheap, and fast diagnostic tools need to be developed. Measurements of ectokinase and ectophosphatase concentrations and activities in serum might thereby provide new opportunities (Fig.3). Such measurements could be embedded in routinely performed blood tests to screen for cancer long before patients show symptoms. Recent studies with more than 600 patients (374 healthy controls, 229 cancer patients) showed a significant upregulation of ecto-PKA concentrations in serum of cancer patients in contrast to healthy controls 64. While more than 70% of the control patients had undetectable or low ecto-PKA concentrations in serum, more than 85% of the cancer patients had high levels of PKA concentrations, with average activity fivefold higher compared to the healthy controls. In another independent study, sera of 500 patients (295 various cancers, 155 normal controls, 55 without cancer) were analyzed by autoantibody against ecto-PKA. The presented anti-ecto-PKA measurement showed a 90% sensitivity and 80% specificity compared to the conventional methods with 83% sensitivity and 80% specificity 65. Only recently, the quantification of ecto-PKA has been patented as a cancer marker for prostate and breast cancer 66. As suggested by the research group, this approach has the potential to replace the commonly used PSA screening test for prostate cancer and the mammograms screening test for breast cancer, which cost nearly $6 billion annually in the United States alone, with limited reliability of the outcome 67–69. Alternatively or in combination, the ecto-PKC and ecto-CKII are other kinases well suited for phenotyping as they are reported in the ECM and show upregulated levels in secretory vesicles of prostate cancer samples 36,70. Such a path holds considerable promise particularly since a 10-fold increased abundance of ecto-PKC in serum of cancer patients with renal, colon, rectal, adrenal, and lung cancer compared to normal serum has recently been reported 71.
Figure 3.
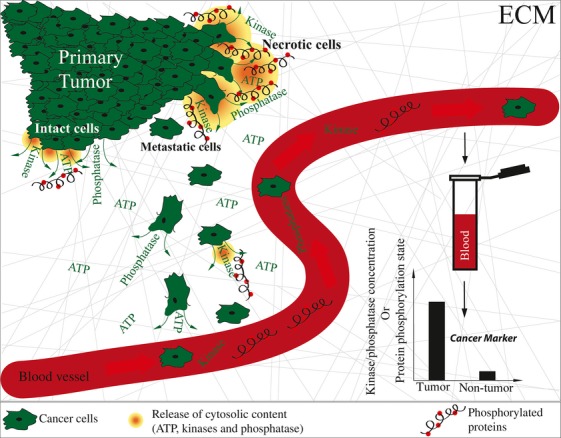
From tumor sites to the blood stream: ectokinases and phosphatases as cancer markers. Enhanced ectokinase and ectophosphatase secretion is seen in intact tumor cells as well as from dying necrotic cells. Blood samples from cancer patients have significantly enhanced ectokinase and ectophosphatase concentrations and activities 64 and might thus serve as novel biomarkers. Several studies report higher selectivity and specificity of markers for early stage cancer detection.
New Strategies for Cancer Treatment: Drug Targeting of Extracellular Protein Kinases and Phosphatases
In the last two decades, intracellular protein kinases have emerged as the most important drug targets in pharmaceutical industry leading to some 20 approved drugs on the market and hundreds more in clinical trials 72. To reduce side effects, a combinatorial approach is needed, one targeting and killing cancer cells while one also tries to prevent or revert the diseased state of ECM. One can further speculate that these drugs might have less side effects as they will not directly interfere with intracellular signaling events 73, but are expected to regulate primarily outside-in cell signaling. Since several important intracellular protein kinases and phosphatase including PKA, PKC, CKII, FAM20C, alkaline phosphatase, and PTEN phosphatase have been found as ectokinases and ectophosphatases, especially in cancer malignancy and progression 30, their potential as novel drug targets has been highlighted 74,75, but not yet systematically exploited. The overexpression of ecto-PKA in secretory vesicles in prostate cancer further points to a putative regulatory role of ectokinases in cancer 70. The expression of the ecto-PKA kinase, as probed in serum of melanoma patients, correlated with the appearance and size of the tumor and tumor removal reduced the levels of ecto-PKA 14. Ecto-PKC is another kinase that has been reported to be present and active in sera of cancer patients with renal, colon, rectal, adrenal, and lung cancer 36,71. Both ecto-PKC and ecto-CKII have been reported to be expressed in secretory vesicles in prostate cancer and they might thus serve as novel targets 70. The role for FAM20C kinase 37, which is present and active in the ECM, is already discussed in the regulation of bone metastasis 76.
Besides protein kinases, protein phosphatases could also serve as potential drug targets. Most recently, monoclonal antibodies were designed to target the extracellular alkaline phosphatase that is expressed on the surface of gastrointestinal cancer cells 77. In addition, the PTEN phosphatase, a tumor suppressor that is known to induce tumor cell death in vitro and in vivo, has been reported to be secreted and subsequently enter other cells where it modifies their signaling and survival 40. Finally, mutations in PTEN and their down regulation are reported to be involved in invasion and metastasis of colorectal carcinomas, indicating PTEN as a novel drug target and a marker for colorectal carcinoma 78. Another advantage is that ectokinases and ectophosphatases could be targeted in cases where other drugs are not efficient anymore due to resistance of the tumor. Consequently, selected extracellular protein kinases and phosphatases might be good candidates for the development of novel drug targets.
Future Perspectives
As extracellular protein phosphorylation is moving into the spotlight of attention, our goal here is to stimulate a thinking process how to best utilize this information for the fight of cancer. An increased understanding of the role of ectokinases and ectophosphatases in the regulation of outside-in signaling pathways in cancer malignancy and progression might result not only in exciting new science but also in the design of new combinatorial drugs that can display their functions in extracellular space 15–21, perhaps complementing conventional therapies, by modulating outside-in cell signaling through the posttranslational modification of extracellular proteins. Starting to apply the knowledge gained in the last 60 years about intracellular protein kinases to the extracellular space offers new opportunities. Ultimately, we need to learn not only how to effectively kill cancer cells but also how to repair diseased cancerous ECM that is left behind and has the potency to send altered instructive signals to newly invading cells.
Acknowledgments
This work was supported by the ERC Advanced Grant (European Research Council, Contract Nr. 233157, V. V.), CCMX Competence Centre for Materials Science and Technology, Grant 0-21108-10, the Swiss Initiative in Systems Biology (SystemsX.ch, “Phosphorylation Modulated Networks of the Cell” [PhosphoNetX]), the Swiss National Science Foundation (SNF, Contract Nr. 3100A0-116236), the Swiss National Science Foundation (Postdoctoral fellowship PBEZP3_145998 to G. Y.), and by the ETH Zurich.
Conflict of Interest
None declared.
References
- Lippert TH, Ruoff HJ. Volm M. Intrinsic and acquired drug resistance in malignant tumors. The main reason for therapeutic failure. Arzneimittelforschung. 2008;58:261–264. doi: 10.1055/s-0031-1296504. [DOI] [PubMed] [Google Scholar]
- Gottesman MM. Mechanisms of cancer drug resistance. Annu. Rev. Med. 2002;53:615–627. doi: 10.1146/annurev.med.53.082901.103929. [DOI] [PubMed] [Google Scholar]
- Ware KE, Hinz TK, Kleczko E, Singleton KR, Marek LA, Helfrich BA, et al. A mechanism of resistance to gefitinib mediated by cellular reprogramming and the acquisition of an FGF2-FGFR1 autocrine growth loop. Oncogenesis. 2013;2:e39. doi: 10.1038/oncsis.2013.4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santarpia M, De Pas TM, Altavilla G, Spaggiari L. Rosell R. Moving towards molecular-guided treatments: erlotinib and clinical outcomes in non-small-cell lung cancer patients. Future Oncol. 2013;9:327–345. doi: 10.2217/fon.13.6. [DOI] [PubMed] [Google Scholar]
- Hunter T. Why nature chose phosphate to modify proteins. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012;367:2513–2516. doi: 10.1098/rstb.2012.0013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ubersax JA. Ferrell JE., Jr Mechanisms of specificity in protein phosphorylation. Nat. Rev. Mol. Cell Biol. 2007;8:530–541. doi: 10.1038/nrm2203. [DOI] [PubMed] [Google Scholar]
- Manning G, Plowman GD, Hunter T. Sudarsanam S. Evolution of protein kinase signaling from yeast to man. Trends Biochem. Sci. 2002;27:514–520. doi: 10.1016/s0968-0004(02)02179-5. [DOI] [PubMed] [Google Scholar]
- Holt LJ, Tuch BB, Villen J, Johnson AD, Gygi SP. Morgan DO. Global analysis of Cdk1 substrate phosphorylation sites provides insights into evolution. Science. 2009;325:1682–1686. doi: 10.1126/science.1172867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levitzki A. Tyrosine kinase inhibitors: views of selectivity, sensitivity, and clinical performance. Annu. Rev. Pharmacol. Toxicol. 2013;53:161–185. doi: 10.1146/annurev-pharmtox-011112-140341. [DOI] [PubMed] [Google Scholar]
- Goding JW. Ecto-enzymes: physiology meets pathology. J. Leukoc. Biol. 2000;67:285–311. doi: 10.1002/jlb.67.3.285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seger D, Seger R. Shaltiel S. The CK2 phosphorylation of vitronectin. Promotion of cell adhesion via the alpha(v)beta 3-phosphatidylinositol 3-kinase pathway. J. Biol. Chem. 2001;276:16998–17006. doi: 10.1074/jbc.M003766200. [DOI] [PubMed] [Google Scholar]
- Jaros JA, Guest PC, Ramoune H, Rothermundt M, Leweke FM, Martins-de-Souza D, et al. Clinical use of phosphorylated proteins in blood serum analysed by immobilised metal ion affinity chromatography and mass spectrometry. J. Proteomics. 2012;76:36–42. doi: 10.1016/j.jprot.2012.02.015. :Spec No. [DOI] [PubMed] [Google Scholar]
- Kang JH, Asai D, Toita R, Kitazaki H. Katayama Y. Plasma protein kinase C (PKC)alpha as a biomarker for the diagnosis of cancers. Carcinogenesis. 2009;30:1927–1931. doi: 10.1093/carcin/bgp210. [DOI] [PubMed] [Google Scholar]
- Kita T, Goydos J, Reitman E, Ravatn R, Lin Y, Shih WC, et al. Extracellular cAMP-dependent protein kinase (ECPKA) in melanoma. Cancer Lett. 2004;208:187–191. doi: 10.1016/j.canlet.2004.02.018. [DOI] [PubMed] [Google Scholar]
- Dogan SS. Esmaeli B. Ocular side effects associated with imatinib mesylate and perifosine for gastrointestinal stromal tumor. Hematol. Oncol. Clin. North Am. 2009;23:109–114. doi: 10.1016/j.hoc.2008.11.003. ix. [DOI] [PubMed] [Google Scholar]
- Lee MW, Seo CW, Kim SW, Yang HJ, Lee HW, Choi JH, et al. Cutaneous side effects in non-small cell lung cancer patients treated with Iressa (ZD1839), an inhibitor of epidermal growth factor. Acta Derm. Venereol. 2004;84:23–26. doi: 10.1080/00015550310005898. [DOI] [PubMed] [Google Scholar]
- Sipples R. Common side effects of anti-EGFR therapy: acneform rash. Semin. Oncol. Nurs. 2006;22:28–34. doi: 10.1016/j.soncn.2006.01.013. [DOI] [PubMed] [Google Scholar]
- Nelson V, Ziehr J, Agulnik M. Johnson M. Afatinib: emerging next-generation tyrosine kinase inhibitor for NSCLC. Onco. Targets Ther. 2013;6:135–143. doi: 10.2147/OTT.S23165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choueiri TK. Axitinib, a novel anti-angiogenic drug with promising activity in various solid tumors. Curr. Opin. Investig. Drugs. 2008;9:658–671. [PubMed] [Google Scholar]
- Weickhardt AJ, Rothman MS, Salian-Mehta S, Kiseljak-Vassiliades K, Oton AB, Doebele RC, et al. Rapid-onset hypogonadism secondary to crizotinib use in men with metastatic nonsmall cell lung cancer. Cancer. 2012;118:5302–5309. doi: 10.1002/cncr.27450. [DOI] [PubMed] [Google Scholar]
- Hartmann JT, Haap M, Kopp HG. Lipp HP. Tyrosine kinase inhibitors—a review on pharmacology, metabolism and side effects. Curr. Drug Metab. 2009;10:470–481. doi: 10.2174/138920009788897975. [DOI] [PubMed] [Google Scholar]
- Lu P, Weaver VM. Werb Z. The extracellular matrix: a dynamic niche in cancer progression. J. Cell Biol. 2012;196:395–406. doi: 10.1083/jcb.201102147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leeming DJ, Bay-Jensen AC, Vassiliadis E, Larsen MR, Henriksen K. Karsdal MA. Post-translational modifications of the extracellular matrix are key events in cancer progression: opportunities for biochemical marker development. Biomarkers. 2011;16:193–205. doi: 10.3109/1354750X.2011.557440. [DOI] [PubMed] [Google Scholar]
- Egeblad M. Werb Z. New functions for the matrix metalloproteinases in cancer progression. Nat. Rev. Cancer. 2002;2:161–174. doi: 10.1038/nrc745. [DOI] [PubMed] [Google Scholar]
- Zucker S. Vacirca J. Role of matrix metalloproteinases (MMPs) in colorectal cancer. Cancer Metastasis Rev. 2004;23:101–117. doi: 10.1023/a:1025867130437. [DOI] [PubMed] [Google Scholar]
- Overall CM. Lopez-Otin C. Strategies for MMP inhibition in cancer: innovations for the post-trial era. Nat. Rev. Cancer. 2002;2:657–672. doi: 10.1038/nrc884. [DOI] [PubMed] [Google Scholar]
- Coussens LM, Fingleton B. Matrisian LM. Matrix metalloproteinase inhibitors and cancer: trials and tribulations. Science. 2002;295:2387–2392. doi: 10.1126/science.1067100. [DOI] [PubMed] [Google Scholar]
- Kessenbrock K, Plaks V. Werb Z. Matrix metalloproteinases: regulators of the tumor microenvironment. Cell. 2010;141:52–67. doi: 10.1016/j.cell.2010.03.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sang QX, Jin Y, Newcomer RG, Monroe SC, Fang X, Hurst DR, et al. Matrix metalloproteinase inhibitors as prospective agents for the prevention and treatment of cardiovascular and neoplastic diseases. Curr. Top. Med. Chem. 2006;6:289–316. doi: 10.2174/156802606776287045. [DOI] [PubMed] [Google Scholar]
- Yalak G. Vogel V. Extracellular phosphorylation and phosphorylated proteins: not just curiosities but physiologically important. Sci. Signal. 2012;5:re7. doi: 10.1126/scisignal.2003273. [DOI] [PubMed] [Google Scholar]
- Stagg J. Smyth MJ. Extracellular adenosine triphosphate and adenosine in cancer. Oncogene. 2010;29:5346–5358. doi: 10.1038/onc.2010.292. [DOI] [PubMed] [Google Scholar]
- Adinolfi E, Amoroso F. Giuliani AL. P2X7 receptor function in bone-related cancer. J. Osteoporos. 2012;2012:637863. doi: 10.1155/2012/637863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pellegatti P, Raffaghello L, Bianchi G, Piccardi F, Pistoia V. Di Virgilio F. Increased level of extracellular ATP at tumor sites: in vivo imaging with plasma membrane luciferase. PLoS ONE. 2008;3:e2599. doi: 10.1371/journal.pone.0002599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodriguez FA, Contreras C, Bolanos-Garcia V. Allende JE. Protein kinase CK2 as an ectokinase: the role of the regulatory CK2beta subunit. Proc. Natl. Acad. Sci. USA. 2008;105:5693–5698. doi: 10.1073/pnas.0802065105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cvijic ME, Kita T, Shih W, DiPaola RS. Chin KV. Extracellular catalytic subunit activity of the cAMP-dependent protein kinase in prostate cancer. Clin. Cancer Res. 2000;6:2309–2317. [PubMed] [Google Scholar]
- Babinska A, Hogan MV, Sobocki T, Sobocka MB, Ehrlich YH. Kornecki E. Identification of ecto-PKC on surface of human platelets: role in maintenance of latent fibrinogen receptors. Am. J. Physiol. Heart Circ. Physiol. 2000;278:H2008–H2019. doi: 10.1152/ajpheart.2000.278.6.H2008. [DOI] [PubMed] [Google Scholar]
- Tagliabracci VS, Engel JL, Wen J, Wiley SE, Worby CA, Kinch LN, et al. Secreted kinase phosphorylates extracellular proteins that regulate biomineralization. Science. 2012;336:1150–1153. doi: 10.1126/science.1217817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheibe RJ, Kuehl H, Krautwald S, Meissner JD. Mueller WH. Ecto-alkaline phosphatase activity identified at physiological pH range on intact P19 and HL-60 cells is induced by retinoic acid. J. Cell. Biochem. 2000;76:420–436. doi: 10.1002/(sici)1097-4644(20000301)76:3<420::aid-jcb10>3.0.co;2-f. [DOI] [PubMed] [Google Scholar]
- Yamada S, Tsuruya K, Yoshida H, Taniguchi M, Haruyama N, Tanaka S, et al. The clinical utility of serum tartrate-resistant acid phosphatase 5b in the assessment of bone resorption in patients on peritoneal dialysis. Clin. Endocrinol. 2013;78:844–851. doi: 10.1111/cen.12070. [DOI] [PubMed] [Google Scholar]
- Hopkins BD, Fine B, Steinbach N, Dendy M, Rapp Z, Shaw J, et al. A secreted PTEN phosphatase that enters cells to alter signaling and survival. Science. 2013;341:399–402. doi: 10.1126/science.1234907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiwanitkit V. High serum alkaline phosphatase levels, a study in 181 Thai adult hospitalized patients. BMC Fam. Pract. 2001;2:2. doi: 10.1186/1471-2296-2-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desgrosellier JS. Cheresh DA. Integrins in cancer: biological implications and therapeutic opportunities. Nat. Rev. Cancer. 2010;10:9–22. doi: 10.1038/nrc2748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yao ES, Zhang H, Chen YY, Lee B, Chew K, Moore D, et al. Increased beta1 integrin is associated with decreased survival in invasive breast cancer. Cancer Res. 2007;67:659–664. doi: 10.1158/0008-5472.CAN-06-2768. [DOI] [PubMed] [Google Scholar]
- Nikolopoulos SN, Blaikie P, Yoshioka T, Guo W. Giancotti FG. Integrin beta4 signaling promotes tumor angiogenesis. Cancer Cell. 2004;6:471–483. doi: 10.1016/j.ccr.2004.09.029. [DOI] [PubMed] [Google Scholar]
- Pankov R. Yamada K. Fibronectin at a glance. J. Cell Sci. 2002;115:3861–3863. doi: 10.1242/jcs.00059. [DOI] [PubMed] [Google Scholar]
- Hynes RO. The extracellular matrix: not just pretty fibrils. Science. 2009;326:1216–1219. doi: 10.1126/science.1176009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vogel V. Mechanotransduction involving multimodular proteins: converting force into biochemical signals. Annu. Rev. Biophys. Biomol. Struct. 2006;35:459–488. doi: 10.1146/annurev.biophys.35.040405.102013. [DOI] [PubMed] [Google Scholar]
- Bissell DM. Chronic liver injury, TGF-beta, and cancer. Exp. Mol. Med. 2001;33:179–190. doi: 10.1038/emm.2001.31. [DOI] [PubMed] [Google Scholar]
- Helleman J, Jansen MP, Ruigrok-Ritstier K, van Staveren IL, Look MP, van Meijer- Gelder ME, et al. Association of an extracellular matrix gene cluster with breast cancer prognosis and endocrine therapy response. Clin. Cancer Res. 2008;14:5555–5564. doi: 10.1158/1078-0432.CCR-08-0555. [DOI] [PubMed] [Google Scholar]
- Hynes RO. Cell-matrix adhesion in vascular development. J. Thromb. Haemost. 2007;5:32–40. doi: 10.1111/j.1538-7836.2007.02569.x. [DOI] [PubMed] [Google Scholar]
- Zeng ZZ, Jia Y, Hahn NJ, Markwart SM, Rockwood KF. Livant DL. Role of focal adhesion kinase and phosphatidylinositol 3′-kinase in integrin fibronectin receptor-mediated, matrix metalloproteinase-1-dependent invasion by metastatic prostate cancer cells. Cancer Res. 2006;66:8091–8099. doi: 10.1158/0008-5472.CAN-05-4400. [DOI] [PubMed] [Google Scholar]
- Ricciardelli C. Rodgers RJ. Extracellular matrix of ovarian tumors. Semin. Reprod. Med. 2006;24:270–282. doi: 10.1055/s-2006-948556. [DOI] [PubMed] [Google Scholar]
- Fornaro M, Plescia J, Chheang S, Tallini G, Zhu YM, King M, et al. Fibronectin protects prostate cancer cells from tumor necrosis factor-alpha-induced apoptosis via the AKT/survivin pathway. J. Biol. Chem. 2003;278:50402–50411. doi: 10.1074/jbc.M307627200. [DOI] [PubMed] [Google Scholar]
- Ogata Y, Heppelmann CJ, Charlesworth MC, Madden BJ, Miller MN, Kalli KR, et al. Elevated levels of phosphorylated fibrinogen-alpha-isoforms and differential expression of other post-translationally modified proteins in the plasma of ovarian cancer patients. J. Proteome Res. 2006;5:3318–3325. doi: 10.1021/pr060344+. [DOI] [PubMed] [Google Scholar]
- Siegel R, Naishadham D. Jemal A. Cancer statistics, 2013. CA Cancer J. Clin. 2013;63:11–30. doi: 10.3322/caac.21166. [DOI] [PubMed] [Google Scholar]
- Molina R, Barak V, van Dalen A, Duffy MJ, Einarsson R, Gion M, et al. Tumor markers in breast cancer- European Group on Tumor Markers recommendations. Tumour Biol. 2005;26:281–293. doi: 10.1159/000089260. [DOI] [PubMed] [Google Scholar]
- Leonard GD, Low JA, Berman AW. Swain SM. CA 125 elevation in breast cancer: a case report and review of the literature. Breast J. 2004;10:146–149. doi: 10.1111/j.1075-122x.2004.21374.x. [DOI] [PubMed] [Google Scholar]
- Tricoli JV, Schoenfeldt M. Conley BA. Detection of prostate cancer and predicting progression: current and future diagnostic markers. Clin. Cancer Res. 2004;10:3943–3953. doi: 10.1158/1078-0432.CCR-03-0200. [DOI] [PubMed] [Google Scholar]
- Bremnes RM, Sirera R. Camps C. Circulating tumour-derived DNA and RNA markers in blood: a tool for early detection, diagnostics, and follow-up? Lung Cancer. 2005;49:1–12. doi: 10.1016/j.lungcan.2004.12.008. [DOI] [PubMed] [Google Scholar]
- Qin LX. Tang ZY. Recent progress in predictive biomarkers for metastatic recurrence of human hepatocellular carcinoma: a review of the literature. J. Cancer Res. Clin. Oncol. 2004;130:497–513. doi: 10.1007/s00432-004-0572-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tarro G, Perna A. Esposito C. Early diagnosis of lung cancer by detection of tumor liberated protein. J. Cell. Physiol. 2005;203:1–5. doi: 10.1002/jcp.20195. [DOI] [PubMed] [Google Scholar]
- Crawford NP, Colliver DW. Galandiuk S. Tumor markers and colorectal cancer: utility in management. J. Surg. Oncol. 2003;84:239–248. doi: 10.1002/jso.10325. [DOI] [PubMed] [Google Scholar]
- Gretzer MB. Partin AW. PSA markers in prostate cancer detection. Urol. Clin. North Am. 2003;30:677–686. doi: 10.1016/s0094-0143(03)00057-0. [DOI] [PubMed] [Google Scholar]
- Wang H, Li M, Lin W, Wang W, Zhang Z, Rayburn ER, et al. Extracellular activity of cyclic AMP-dependent protein kinase as a biomarker for human cancer detection: distribution characteristics in a normal population and cancer patients. Cancer Epidemiol. Biomarkers Prev. 2007;16:789–795. doi: 10.1158/1055-9965.EPI-06-0367. [DOI] [PubMed] [Google Scholar]
- Nesterova MV, Johnson N, Cheadle C, Bates SE, Mani S, Stratakis CA, et al. Autoantibody cancer biomarker: extracellular protein kinase A. Cancer Res. 2006;66:8971–8974. doi: 10.1158/0008-5472.CAN-06-1049. [DOI] [PubMed] [Google Scholar]
- Puskas R. Held D. 2013. Measurement of PKA for cancer detection. Patent. US_8455200_B2.
- Smith DS, Humphrey PA. Catalona WJ. The early detection of prostate carcinoma with prostate specific antigen: the Washington University experience. Cancer. 1997;80:1852–1856. [PubMed] [Google Scholar]
- Elmore JG, Armstrong K, Lehman CD. Fletcher SW. Screening for breast cancer. JAMA. 2005;293:1245–1256. doi: 10.1001/jama.293.10.1245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harris L, Fritsche H, Mennel R, Norton L, Ravdin P, Taube S, et al. American Society of Clinical Oncology 2007 update of recommendations for the use of tumor markers in breast cancer. J. Clin. Oncol. 2007;25:5287–5312. doi: 10.1200/JCO.2007.14.2364. [DOI] [PubMed] [Google Scholar]
- Babiker AA, Ronquist G, Nilsson B. Ekdahl KN. Overexpression of ecto-protein kinases in prostasomes of metastatic cell origin. Prostate. 2006;66:675–686. doi: 10.1002/pros.20268. [DOI] [PubMed] [Google Scholar]
- Cho YS, Park YG, Lee YN, Kim MK, Bates S, Tan L, et al. Extracellular protein kinase A as a cancer biomarker: its expression by tumor cells and reversal by a myristate-lacking Ca and RIIb subunit overexpression. Proc. Natl Acad. Sci. USA. 2000;18:835–840. doi: 10.1073/pnas.97.2.835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen P. Alessi DR. Kinase drug discovery–what's next in the field? ACS Chem. Biol. 2013;8:96–104. doi: 10.1021/cb300610s. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ehrlich YH. Kornecki E. Ecto-protein kinases as mediators for the action of secreted ATP in the brain. Prog. Brain Res. 1999;120:411–426. doi: 10.1016/s0079-6123(08)63573-6. [DOI] [PubMed] [Google Scholar]
- Redegeld FA, Caldwell CC. Sitkovsky MV. Ecto-protein kinases: ecto-domain phosphorylation as a novel target for pharmacological manipulation? Trends Pharmacol. Sci. 1999;20:453–459. doi: 10.1016/s0165-6147(99)01399-1. [DOI] [PubMed] [Google Scholar]
- Wright X. Wright JF. Biological insights into therapeutic protein modifications throughout trafficking and their biopharmaceutical applications. Int. J. Cell Biol. 2013;2013:273086. doi: 10.1155/2013/273086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo X. Wang XF. New secrets behind bone metastasis. Cell Res. 2012;22:1309–1311. doi: 10.1038/cr.2012.99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li M, Gao J, Feng R, Wang Y, Chen X, Sun J, et al. Generation of monoclonal antibody MS17-57 targeting secreted alkaline phosphatase ectopically expressed on the surface of gastrointestinal cancer cells. PLoS One. 2013;8:e77398. doi: 10.1371/journal.pone.0077398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li XH, Zheng HC, Takahashi H, Masuda S, Yang XH. Takano Y. PTEN expression and mutation in colorectal carcinomas. Oncol. Rep. 2009;22:757–764. doi: 10.3892/or_00000497. [DOI] [PubMed] [Google Scholar]
- Daub H, Olsen JV, Bairlein M, Gnad F, Oppermann FS, Korner R, et al. Kinase-selective enrichment enables quantitative phosphoproteomics of the kinome across the cell cycle. Mol. Cell. 2008;31:438–448. doi: 10.1016/j.molcel.2008.07.007. [DOI] [PubMed] [Google Scholar]
- Sugiyama N, Masuda T, Shinoda K, Nakamura A, Tomita M. Ishihama Y. Phosphopeptide enrichment by aliphatic hydroxy acid-modified metal oxide chromatography for nano-LC-MS/MS in proteomics applications. Mol. Cell Proteomics. 2007;6:1103–1109. doi: 10.1074/mcp.T600060-MCP200. [DOI] [PubMed] [Google Scholar]
- Rigbolt KT, Prokhorova TA, Akimov V, Henningsen J, Johansen PT, Kratchmarova I, et al. System-wide temporal characterization of the proteome and phosphoproteome of human embryonic stem cell differentiation. Sci. Signal. 2011;4:rs3. doi: 10.1126/scisignal.2001570. [DOI] [PubMed] [Google Scholar]
- Goswami T. Bryan B. Methods for the isolation of phosphoproteins and phosphopeptides for mass spectrometry analysis: toward increased functional phosphoproteomics. Sample Preparation in Biological Mass Spectrometry. 2011:627–655. 2011 Date: 20 May 2011. [Google Scholar]
- Olsen JV, Blagoev B, Gnad F, Macek B, Kumar C, Mortensen P, et al. Global, in vivo, and site-specific phosphorylation dynamics in signaling networks. Cell. 2006;127:635–648. doi: 10.1016/j.cell.2006.09.026. [DOI] [PubMed] [Google Scholar]
- Zhou W, Ross MM, Tessitore A, Ornstein D, Vanmeter A, Liotta LA, et al. An initial characterization of the serum phosphoproteome. J. Proteome Res. 2009;8:5523–5531. doi: 10.1021/pr900603n. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruse CI, McClatchy DB, Lu B, Cociorva D, Motoyama A, Park SK, et al. Motif-specific sampling of phosphoproteomes. J. Proteome Res. 2008;7:2140–2150. doi: 10.1021/pr800147u. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han G, Ye M, Liu H, Song C, Sun D, Wu Y, et al. Phosphoproteome analysis of human liver tissue by long-gradient nanoflow LC coupled with multiple stage MS analysis. Electrophoresis. 2010;31:1080–1089. doi: 10.1002/elps.200900493. [DOI] [PubMed] [Google Scholar]
- Rikova K, Guo A, Zeng Q, Possemato A, Yu J, Haack H, et al. Global survey of phosphotyrosine signaling identifies oncogenic kinases in lung cancer. Cell. 2007;131:1190–1203. doi: 10.1016/j.cell.2007.11.025. [DOI] [PubMed] [Google Scholar]
- Lee HJ, Na K, Kwon MS, Kim H, Kim KS. Paik YK. Quantitative analysis of phosphopeptides in search of the disease biomarker from the hepatocellular carcinoma specimen. Proteomics. 2009;9:3395–3408. doi: 10.1002/pmic.200800943. [DOI] [PubMed] [Google Scholar]
- Bennetzen MV, Larsen DH, Bunkenborg J, Bartek J, Lukas J. Andersen JS. Site-specific phosphorylation dynamics of the nuclear proteome during the DNA damage response. Mol. Cell Proteomics. 2010;9:1314–1323. doi: 10.1074/mcp.M900616-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han G, Ye M, Zhou H, Jiang X, Feng S, Jiang X, et al. Large-scale phosphoproteome analysis of human liver tissue by enrichment and fractionation of phosphopeptides with strong anion exchange chromatography. Proteomics. 2008;8:1346–1361. doi: 10.1002/pmic.200700884. [DOI] [PubMed] [Google Scholar]