

Abstract
AIM: To evaluate the expression of different insulin-like growth factor (IGF)-1 mRNA isoforms and IGF-1 receptor (IGF-1R) mRNA in hepatitis C virus (HCV)-infected livers.
METHODS: Thirty-four liver biopsy specimens from chronic hepatitis C (CH-C) patients were obtained before anti-viral therapy. Inflammatory activity (grading) and advancement of fibrosis (staging) were evaluated using a modified point scale of METAVIR. The samples were analyzed using quantitative real-time PCR technique. From fragments of liver biopsies and control liver that were divided and ground in liquid nitrogen, RNA was isolated using RNeasy Fibrous Tissue Mini Kit according to the manufacturer’s instruction. Expression levels of IGF-1 mRNA isoforms (IGF-1A, IGF-1B, IGF-1C, P1, and P2) and IGF-1R mRNA were determined through normalization of copy numbers in samples as related to reference genes: glyceraldehyde-3-phosphate dehydrogenase and hydroxymethylbilane synthase. Results on liver expression of the IGF-1 mRNA isoforms and IGF-1R transcript were compared to histological alterations in liver biopsies and with selected clinical data in the patients. Statistical analysis was performed using Statistica PL v. 9 software.
RESULTS: The study showed differences in quantitative expression of IGF-1 mRNA variants in HCV-infected livers, as compared to the control. Higher relative expression of total IGF-1 mRNA and of IGF-1 mRNAs isoforms (P1, A, and C) in HCV-infected livers as compared to the control were detected. Within both groups, expression of the IGF-1A mRNA isoform significantly prevailed over expressions of B and C isoforms. Expression of P1 mRNA was higher than that of P2 only in CH-C. Very high positive correlations were detected between reciprocal expressions of IGF-1 mRNA isoforms P1 and P2 (r = 0.876). Expression of P1 and P2 mRNA correlated with IGF-1A mRNA (r = 0.891; r = 0.821, respectively), with IGF-1B mRNA (r = 0.854; r = 0.813, respectively), and with IGF-1C mRNA (r = 0.839; r = 0.741, respectively). Expression of IGF-1A mRNA significantly correlated with isoform B and C mRNA (r = 0.956; r = 0.869, respectively), and B with C isoforms (r = 0.868) (P < 0.05 in all cases). Lower expression of IGF-1A and B transcripts was noted in the more advanced liver grading (G2) as compared to G1. Multiple negative correlations were detected between expression of various IGF-1 transcripts and clinical data (e.g., alpha fetoprotein, HCV RNA, steatosis, grading, and staging). Expression of IGF-1R mRNA manifested positive correlation with grading and HCV-RNA.
CONCLUSION: Differences in quantitative expression of IGF-1 mRNA isoforms in HCV-infected livers, as compared to the control, suggest that HCV may induce alteration of IGF-1 splicing profile.
Keywords: Chronic hepatitis C, Insulin-like growth factor-1 receptor, Insulin-like growth factor-1 mRNA isoforms, Quantitative polymerase chain reaction
Core tip: Hepatitis C virus (HCV) may induce alteration of insulin-like growth factor (IGF)-1 splicing profile. A quantitative polymerase chain reaction analysis confirmed higher relative expression of total IGF-1 mRNA and of IGF-1 mRNAs isoforms P1, A, and C in HCV-infected livers as compared to the control. An increase in inflammatory activity (grading) of HCV-infected livers was linked to decreased IGF-1 mRNA expression, an altered profile of mRNA isoforms, and to an increase in IGF-1R mRNA expression. Decreased expression level of IGF-1 mRNA isoforms and an increased liver expression of IGF-1R mRNA were associated with indicators of liver damage (e.g., grading, staging, steatosis, and liver serum enzyme activity), and may be of prognostic significance.
INTRODUCTION
Hepatitis C virus (HCV) is the major causative agent of hepatocellular carcinoma (HCC), mainly through the indirect pathways of chronic inflammation, cell death, and proliferation[1,2]. The function of the insulin-like growth factor (IGF) system in the promotion of cell growth and neoplastic transformation has been previously described[3,4]. The IGF ligands (IGF-1 and -2) bind to the receptors, which include: IGF-1 receptor (IGF-1R), IGF-2R, insulin receptor, and hybrid receptors (IGF-1R/insulin receptor)[3]. The involvement of IGF receptors in the maintenance of the transformed hepatocyte phenotype was also described[5-9]. Most studies documented an increase in IGF-1R expression from preneoplastic lesions to developed HCC[5,9,10]. Other studies found no significant difference in the expression of IGF-1R mRNA between HCC and a control[8]. Similarly, an increase in expression of IGF-1R could not be noted in cultured human hepatocytes[11]. In a few studies on patients with liver cirrhosis and chronic hepatitis C (CH-C), the amounts of IGF-1R transcript were found to increase as compared to the normal liver[12,13]. A more thorough recognition of the role played by IGF system in hepatic carcinogenesis is thought to improve HCC therapy[3,9,14].
The six exons of IGF-1 are alternatively spliced into multiple transcripts, encoding specific circulating and tissue-specific isoforms of the IGF-1 peptide. At the 5’end of the gene, different promoters (P1 and P2), in combination with alternative transcription start sites and differential splicing, generate the mutually-exclusive class 1 and class 2 IGF-1 isoforms[15-17]. At the 3’end of the gene, alternative splicing gives rise to at least three subsets of RNA transcripts, each encoding three distinct C-terminal portions of the unique E-peptide, as well as the 3’-UTR[15,18,19]. Exon 3 encodes parts of the signal peptide and the mature peptide common to all isoforms, while exon 4 encodes the rest of the mature peptide and the proximal part of the E domain. The composition of nucleotides in exons 5 and 6 determines the formation of isoforms A (Ea), B (Eb), and C (Ec) within classes 1 and 2[20]. The biochemical mechanism which controls use of IGF-1 promoters 1 or 2 in alternate splicing remains poorly recognized[16,21-23]. Studies on human liver RNA have demonstrated that IGF-1 transcript undergoes alternate splicing that contains exons 3 and 4, as well as 49 bp of exon 5 and exon 6 (exon 4-5-6)[24]. The role of IGF-1 alternate splicing has been best recognized in skeletal muscular tissue[25-28] and nervous tissue[29,30]. A differential profile of IGF-1 mRNA isoforms was demonstrated in different tumors[31-34]. It remains unknown as to whether HCV and its oncogenic proteins (C, NS3, and NS5A) in vivo may induce alterations in the profile of hepatic IGF-1 gene expression[35].
This study aimed to evaluate the expression of various IGF-1 mRNA isoforms (P1, P2, 1A, 1B, and 1C) and IGF-1R mRNA in chronically HCV-infected livers. Herein we examined if IGF-1 alternative splicing is associated with the degree of liver damage (grading and staging) caused by HCV virus. Results on liver expression of the IGF-1 mRNA isoforms and IGF-1R transcript were compared to histological alterations in liver biopsies and with selected clinical data in the patients. Data concerning changes in IGF-1 alternative splicing in CH-C have not been published up to now. The relationship between liver expression of mRNA IGF-1 isoforms and progression of CH-C to HCC is unclear.
MATERIALS AND METHODS
Patients
The examined group consisted of 34 patients (age: 18-63 years; 18 men and 16 women) with CH-C who were diagnosed and treated in the Department of Infectious Diseases, Poznan University of Medical Sciences in Poznan from 2010 to 2012. Patients were referred to an anti-viral treatment and not previously treated. Infections with other hepatotropic viruses (HBV, HCMV, or EBV) or other reasons of liver damage were excluded (e.g., alcohol abuse, autoimmune hepatitis, NASH, drugs, or history of anti-cancer therapy). Patients with diabetes mellitus, kidney failure or any hormones disturbances were not included in the group. In the study, we used basic clinical data on HCV-infected patients, as well as other results involving biochemical tests on peripheral blood, results of ELISA tests (glucose, insulin), and histopathological examination of liver biopsies. Presence of HCV-specific antibodies was tested using chemiluminescence and ARCHITECT Anti-HCV kits (ABBOTT, Wiesbaden, Germany) in ARCHITECT and 2000 analyzers (ABBOTT). Infection with HCV was confirmed by estimating serum HCV-RNA via the application of GeneProof HEPATITIS C VIRUS HCV tests (GeneProof a.s., Brno, Czech Republic) and manifesting sensitivity of 50 IU/mL. In all patients, HCV genotype was estimated (VERSANT HCV GENOTYPE 2.0 ASSAY, LiPA).
Negative tissue control (n = 7) (patient age: 35-72 years; 4 men, 3 women) involved liver fragments with no morphological traits of organ pathology that were perioperatively sampled from the vicinity of the dissected focal lesion in the liver and a single liver biopsy taken from a patient to diagnose the reasons for elevated aminotransferase activity. Said patient proved to be HCV-negative (absence of specific antibodies and of HCV RNA). The remaining control material also originated from HCV- and HBV-negative patients. Age and sex of the patients were known. The control material was obtained from the Chair and Department of General Surgery, Gastroenterological Oncology and Plastic Surgery, Poznan University of Medical Sciences in Poznan.
Tissue material
Liver biopsy was done in all cases as a routine procedure before antiviral therapy. Basing on USG tests and alpha fetoprotein (AFP) levels, none of the patients had neoplastic growth (HCC). Written informed consent was obtained from every patient before liver biopsy and approval for the study was granted by the institution’s ethical committee (No. 22/09). The excised liver fragment (HCV-infected and control) was divided, cutting off its terminal 0.5 cm fragment, which was immersed in RNA Stabilization Solution (RNAlater®, Applied Biosystems) at -80 °C until use. The remaining part of the fragments obtained from patients were fixed in a buffered 10% solution of formalin and embedded in paraffin. About 5 μm-thick preparations were stained with hematoxylin and eosin and then silver impregnated using standard techniques. Inflammatory activity (grading) and advancement of fibrosis (staging) were evaluated using a modified point scale of METAVIR[36]. This score is composed of a two-letter and two-number scoring system: histological activity (grading: G0 - no activity, G1 - mild activity, G2 - moderate activity, G3 - severe activity) and fibrosis (staging: S0 - no fibrosis, S1 - portal fibrosis without septa, S2 - portal fibrosis with rare septa, S3 - numerous septa without cirrhosis, S4 - cirrhosis). Fatty degeneration of the liver was evaluated using a point scale, in which grade 0 corresponded to an absence of fatty degeneration, while grades 1 and 2 corresponded to < 30% and ≥ 30%-70% hepatocytes showing traits of fatty degeneration, respectively.
Technique of quantitative real-time PCR
RNA was isolated using RNeasy Fibrous Tissue Mini Kit (QIAGEN), according to the manufacturer’s instructions, from fragments of liver biopsies and control liver that were divided and ground in liquid nitrogen. In the course of the procedure, traces of DNA contamination were eliminated using DNases. Total RNA was dissolved in RNase-free water. Quality of RNA preparations was consecutively checked using electrophoresis in an agar-formaldehyde gel. RNA content was quantitated by spectrophotometry. Every RNA sample was subjected to additional digestion with DNase using RNase-Free DNase Set (QIAGEN) in order to avoid contamination with genomic DNA. Subsequently, 1 μg RNA from every sample was subjected to reverse transcription using QuantiTect Reverse Transcription Kit (QIAGEN) and cDNA was obtained. Analysis of gene expression was performed using primers specific for individual mRNA IGF-1 isoforms (as previously described)[34] and selected reference genes: GAPDH (glyceraldehyde-3-phosphate dehydrogenase) and HMBS (hydroxymethylbilane synthase)[37]. The primer sequences for IGF-1R were: forward 5’-GGGAATGGAGTGCTGTATG-3’, reverse 5’-CACAGAAGCTTCGTTGAGAA-3’, GAPDH forward 5’-AAGGTCGGAGTCAACGGATTT-3’, reverse 5’-ACCAGAGTTAAAAGCAGCCCTG-3’, HMBS forward 5’-TGCAACGGCGGAAGAAAA-3’, and reverse 5’-ACGAGGCTTTCAATGTTGCC-3’. The reactions were performed in a final volume of 10 μL. Each sample contained 15 ng/μL cDNA and a mixture of reagents forming the SYBR Green PCR master mix (Applied Biosystems, United Kingdom), with each primer pair at a concentration of 0.25 mmol/L. The reaction was conducted using an automated fluorimeter (Rotor-Gene 6000, Corbett Research). The PCR program was as follows: (1) preliminary denaturation, 95 °C, 10 min; (2) denaturation, 95 °C, 10-15 s; (3) primer annealing, 53-67 °C, 15-35 s; and (4) elongation, 72 °C, 15-40 s. The number of cycles was 40-50. The initial quantity of the product was calculated in relation to the standard curve. Presence of an appropriate product was evaluated by determination of the melting point for a specific PCR product. All samples were amplified in duplicate or triplicate and, in cases in which the results varied by more than 15%, the reactions were repeated. Expression levels of IGF-1 mRNA isoforms (IGF-1A, IGF-1B, IGF-1C, P1, and P2), and IGF-1R mRNA were determined through normalization of copy numbers in samples as related to reference genes (housekeeping genes). In the normalization, reference genes were accepted to include liver-specific GAPDH and HMBS genes, according to literature data[37].
Statistical analysis
At the first stage of statistical analysis, the consistency of all results with the normal distribution of Gauss was verified by using the Shapiro-Wilk test. Parameters of descriptive statistics were subsequently calculated (arithmetic mean, standard deviation, median value, and minimum and maximum value). Data related to quantitative gene expression of IGF-1 mRNA isoforms, IGF-1R mRNA were compared to data for normal liver (negative control) using Mann-Whitney’s test (a non-parametric test for unlinked variables for two groups). In cases of linked variables, the Wilcoxon test was used. For comparing more than two groups, Kruskal-Wallis and multiple comparison Dunn tests were employed. For comparison of the percentage shares of IGF-1 mRNA isoforms, the test of differences between two structural indices was employed. Pearson’s correlation and Spearman’s rank correlation were used to correlate values of variables. Effect of age was also analyzed to determine any correlation between IGF-1 mRNA isoforms, and IGF-1R transcript, and selected clinical data. The results were thought to be statistically significant at P < 0.05. Statistical analysis was performed using Statistica PL v. 9 software (Statsoft, Inc., Tulsa, OK, United States).
RESULTS
Expression of mRNA class 1 (I) and 2 (II) as related to the used promoter P1 or P2
Both among the HCV-infected patients and in the control, transcription of IGF-1 from the first promoter (P1) (class 1) prevailed. In the CH-C group, the shares of transcripts from class 1 (from P1) (76.4%) and class 2 (from P2) (23.6%) were similar to those in the control (62.7% and 37.3%, respectively) (P > 0.05) (Figure 1).
Figure 1.
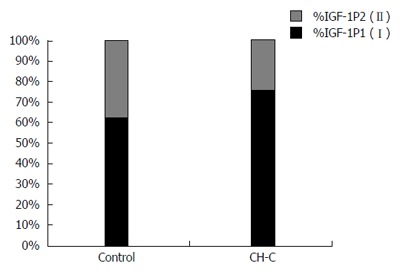
Shares (%) of studied insulin-like growth factor-1 mRNA isoforms of class 1 (I) and class 2 (II) (depending upon employed promoter P1 or P2) as related to the total mRNA for both insulin-like growth factor-1 promoters (100%) in chronic hepatitis C patients and in the control. IGF: Insulin-like growth factor; CH-C: Chronic hepatitis C.
Expression of mRNA isoforms: IGF-1A, IGF-1B, and IGF-1C - shares in percentages
In the CH-C group, expression of the IGF-1A (A) splicing form of mRNA prevailed (92% of all transcripts). Similarly, in the control group, the highest expression was manifested by the A isoform, followed by B and C (Figure 2). Expression of the A isoform was significantly higher in the CH-C group than in the control (92% vs 85%), while expression of isoform B was lower than in the control (5.2% vs 12.9%). No significant differences could be demonstrated between the two compared groups in shares of expressed P1 and P2 transcripts (Figure 3).
Figure 2.
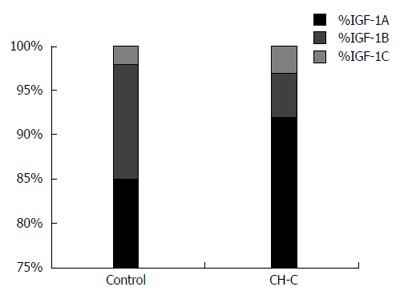
Shares (%) of splicing mRNA isoforms for insulin-like growth factor-1 (1A, 1B, 1C) as related to the total amount of mRNA for all three insulin-like growth factor-1 isoforms (100%) in chronic hepatitis C patients and in the control. IGF: Insulin-like growth factor; CH-C: Chronic hepatitis C.
Figure 3.
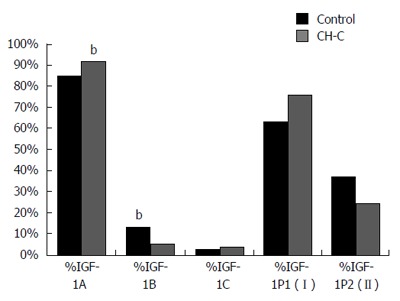
Comparison of shares (%) in expression of insulin-like growth factor-1 mRNA isoforms in patients chronically infected with hepatitis C virus and in the control. bP < 0.01 vs control.
Quantitative analysis of IGF-1 mRNA isoforms in the CH-C and control groups
Detailed analysis of the relative expression manifested by each IGF-1 mRNA isoform, normalized against reference genes (GAPDH and HMBS), demonstrated a significantly higher expression of IGF-1A and IGF-1C mRNA isoforms, of P1 transcripts (class 1), and of total IGF-1 mRNA in the CH-C group as compared to the control. A significantly higher expression of total IGF-1 and all isoforms and classes of IGF-1 mRNA was detected in women as compared to men (Table 1).
Table 1.
Quantitative expression of insulin-like growth factor-1 mRNA isoforms, total mRNA insulin-like growth factor-1, and insulin-like growth factor-1R mRNA (mean ± SD) normalized in relation to housekeeping genes in chronic hepatitis C and in control organ
| mRNA | Control (n = 7) | CH-C (n = 34) | P value |
CH-C |
P value | |
| Women (n = 16) | Men (n = 18) | |||||
| class 1 (P1) | 0.19 ± 0.21 | 0.45 ± 0.45 | P = 0.025 | 0.63 ± 0.59 | 0.28 ± 0.17 | P = 0.003 |
| class 2 (P2) | 0.08 ± 0.07 | 0.15 ± 0.17 | P = 0.164 | 0.20 ± 0.15 | 0.11 ± 0.18 | P = 0.006 |
| IGF-1A | 1.30 ± 1.31 | 5.28 ± 9.94 | P = 0.009 | 7.88 ± 13.83 | 2.97 ± 3.32 | P = 0.027 |
| IGF-1B | 0.18 ± 0.18 | 0.28 ± 0.40 | P = 0.486 | 0.35 ± 0.45 | 0.22 ± 0.35 | P = 0.042 |
| IGF-1C | 0.04 ± 0.05 | 0.21 ± 0.57 | P = 0.031 | 0.35 ± 0.82 | 0.08 ± 0.06 | P = 0.046 |
| Total IGF-1 | 1.52 ± 1.52 | 5.77 ± 10.85 | P = 0.014 | 8.58 ± 15.09 | 3.26 ± 3.70 | P = 0.027 |
| IGF-1R | 0.26 ± 0.18 | 0.14 ± 0.04 | P = 0.016 | 0.14 ± 0.05 | 0.14 ± 0.04 | P = 0.295 |
IGF-1: Insulin-like growth factor 1; CH-C: Chronic hepatitis C.
Analysis of IGF-1 mRNA isoforms in HCV-infected livers as related to activity of inflammation (grading) and of liver fibrosis (staging)
A liver with more intense inflammatory lesions (grading 2, G2) contained a lower expression of all IGF-1 isoforms as compared to their expression in a liver with G1, with the exception of the IGF-1C isoform. No significant differences could be determined between the expression of all IGF-1 mRNA isoforms and the different stages of fibrosis (P > 0.05 in all the cases) (Table 2).
Table 2.
Tissue expression of insulin-like growth factor-1 mRNA isoforms and insulin-like growth factor-1R mRNA (mean ± SD) as related to grading and staging in hepatitis C virus-infected livers
| mRNA | Grading1 1 (n = 15) | Grading 2 (n = 16) | P value | Staging1 1 (n = 21) | Staging 2 (n = 8) | P value |
| class 1 (P1) | 0.63 ± 0.61 | 0.33 ± 0.20 | P = 0.037 | 0.50 ± 0.55 | 0.41 ± 0.23 | P = 0.905 |
| class 2 (P2) | 0.25 ± 0.22 | 0.09 ± 0.06 | P = 0.009 | 0.19 ± 0.20 | 0.11 ± 0.07 | P = 0.549 |
| IGF-1A | 8.67 ± 14.34 | 2.42 ± 1.45 | P = 0.008 | 6.55 ± 12.47 | 3.01 ± 1.77 | P = 0.943 |
| IGF-1B | 0.44 ± 0.55 | 0.15 ± 0.10 | P = 0.012 | 0.34 ± 0.49 | 0.18 ± 0.12 | P = 0.720 |
| IGF-1C | 0.36 ± 0.84 | 0.08 ± 0.06 | P = 0.093 | 0.27 ± 0.72 | 0.09 ± 0.07 | P = 0.582 |
| Total IGF-1 | 9.47 ± 15.66 | 2.65 ± 1.58 | P = 0.006 | 7.16 ± 13.61 | 3.29 ± 1.91 | P = 0.830 |
| IGF-1R | 0.12 ± 0.04 | 0.16 ± 0.03 | P = 0.025 | 0.14 ± 0.04 | 0.16 ± 0.04 | P = 0.198 |
Parameters evaluated in a semi-quantitative scale (see Material and Methods). IGF-1: Insulin-like growth factor 1.
Expression of IGF-1R mRNA
In the CH-C group, expression of IGF-1R mRNA was significantly lower than in the control. No sex-related differences were detected in the expression of the IGF-1R transcript (Table 1). Expression of IGF-1R mRNA was higher in livers with higher grading (G2) as compared to livers with G1 (Table 2).
Expression of various transcripts of IGF-1 and IGF-1R vs the clinical data
Isoforms of IGF-1 mRNA: Expression of total IGF-1 mRNA and of all mRNA isoforms manifested a very strong negative correlation with patient age and BMI value. Expression of all isoforms also demonstrated negative correlations with liver steatosis. Highly negative correlations were detected with activity of ALT, and slightly less pronounced ones (though still negative) with activity of AST. Expression of mRNA isoforms A, B, P1, and P2 showed negative correlations with liver grading. Very poor correlation was detected between liver fibrosis and expression of P2 mRNA. For expression of P2 mRNA, a relatively poor negative correlation was documented, with AFP concentration and HCV viral load. Similarly low were the negative correlations between HCV viral load and expression of IGF-1A mRNA and total IGF-1 mRNA in HCV-infected liver (Table 3).
Table 3.
Values of Spearman’s coefficient for correlation between expression of insulin-like growth factor-1, insulin-like growth factor-1R mRNAs, and clinical data in hepatitis C virus-infected patients
| Class 1 (P1) | Class 2 (P2) | IGF-1A | IGF-1B | IGF-1C | Total IGF-1 | IGF-1R | |
| Age (yr) | -0.5451 | -0.6441 | -0.5821 | -0.6541 | -0.5321 | -0.5961 | 0.3721 |
| BMI | -0.5271 | -0.4521 | -0.4681 | -0.4141 | -0.4891 | -0.4681 | 0.060 |
| Grading | -0.4311 | -0.5691 | -0.4461 | -0.4301 | -0.2851 | -0.4571 | 0.4561 |
| Staging | -0.223 | -0.3461 | -0.182 | -0.187 | -0.160 | -0.190 | 0.264 |
| Steatosis (%) | -0.4151 | -0.5711 | -0.4121 | -0.4081 | -0.4541 | -0.4221 | 0.125 |
| ALT (U/L) | -0.5381 | -0.6161 | -0.5401 | -0.5771 | -0.5331 | -0.5501 | 0.181 |
| AST (U/L) | -0.4341 | -0.5691 | -0.3781 | -0.4001 | -0.3421 | -0.3921 | 0.212 |
| AFP (ng/mL) | -0.291 | -0.3901 | -0.168 | -0.271 | -0.240 | -0.180 | 0.311 |
| HCV RNA (IU/mL) | -0.330 | -0.3881 | -0.3571 | -0.244 | -0.096 | -0.3451 | 0.4031 |
| Total protein (g/dL) | 0.302 | 0.251 | 0.324 | 0.3751 | 0.3661 | 0.319 | 0.141 |
| Albumins (g/dL) | 0.293 | 0.3811 | 0.303 | 0.3491 | 0.222 | 0.295 | 0.123 |
| Gamma globulins (g/dL) | -0.061 | -0.164 | 0.039 | 0.045 | 0.174 | 0.044 | 0.138 |
| Blood platelets (G/L) | 0.273 | 0.3721 | 0.235 | 0.265 | 0.138 | 0.232 | -0.099 |
| Cholesterol (mg/dL) | 0.3861 | 0.286 | 0.4091 | 0.4051 | 0.372 | 0.4041 | 0.048 |
| GGTP (U/L) | -0.384 | -0.6031 | -0.284 | -0.399 | -0.367 | -0.307 | 0.330 |
| Blood glucose (mg/dL) | -0.083 | -0.226 | -0.190 | -0.112 | -0.084 | -0.201 | 0.152 |
| HOMA-IR | -0.067 | -0.157 | -0.237 | -0.140 | -0.184 | -0.234 | -0.019 |
| Insulin (μU/mL) | -0.020 | -0.082 | -0.145 | -0.079 | -0.125 | -0.135 | -0.045 |
The numbers indicate values of r coefficient for which the P < 0.05. IGF-1: Insulin-like growth factor 1; HCV: Hepatitis C virus; AFP: Alpha fetoprotein.
Transcripts of IGF-1R: Expression of IGF-1R mRNA manifested poor positive correlations with age of HCV-infected patients. Moreover, positive correlations were detected between tissue expression of IGF-1R mRNA, grading, and HCV-RNA (Table 3).
Reciprocal correlations between expressions of IGF-1 mRNA isoforms and IGF-1R mRNA
Very high positive Spearman’s correlations were detected between reciprocal expressions of IGF-1 mRNA isoforms P1 and P2 (r = 0.876), P1 and A (r = 0.891), P1 and B (r = 0.854), P1 and C (r = 0.839); P2 and A (r = 0.821), P2 and B (r = 0.813), P2 and C (r = 0.741), A and B (r = 0.956), A and C (r = 0.869), and B and C (r = 0.868) (P < 0.05 in all cases) in patients with HCV infection. In livers with CH-C, additional significantly weak negative correlations were detected between expression of IGF-1R mRNA and quantities of mRNAs for isoforms of IGF-1A (r = -0.397), IGF-1B mRNAs (r = -0.419), and for total expression of IGF-1 mRNA (r = -0.397) (P < 0.05 in all cases).
In control livers, we confirmed very high positive Spearman’s correlation between reciprocal expression of all IGF-1 mRNA isoforms (data not shown). No significant relationships were detected between expression of IGF-1 mRNA isoforms and IGF-1R mRNA in control livers (data not shown).
DISCUSSION
Studies on expression of various IGF-1 mRNA isoforms have been performed for the first time on livers with CH-C. Our results point to the prevalent expression of mRNA from P1 promoter of IGF-1, both on the control liver and in liver with chronic HCV infection. This confirms the involvement of P1 in the production of 60%-65% and of P2 in the production of approximately 25% of IGF-1 transcripts, which was originally demonstrated in rat liver[17,22]. The percentage shares of the remaining IGF-1 mRNA isoforms (A, B, and C) in the control and HCV-infected liver also demonstrated the prevalence of mRNA isoform A overexpression of mRNAs of the remaining isoforms. In the literature data, there are no references to such results. In studies on human papilloma virus-positive and -negative tissues of uterine cervix carcinoma, a significant prevalence was demonstrated of IGF-1B share over remaining isoforms in tissues with uterine cervix carcinoma as compared to the remaining stages of carcinogenesis, suggesting that this form of the transcript may lead to the formation of the peptide Eb with strongly mitogenic properties[33].
Analysis of the relative values of expression manifested by various IGF-1 mRNA isoforms in the livers of the two examined groups confirmed the quantitative prevalence of isoform A over the remaining variants of IGF-1 transcripts. Prevalence of expression for IGF-1A over IGF-1B was also documented by Ohtsuki et al[38] in such organs as the uterus, ovaries, liver, and kidneys in mice. The authors accentuated organ-specific control of transcription manifested by the gene in the course of development. The prevalence (as high as tenfold) of IGF-1A over IGF-1B mRNA transcripts, both in in vivo conditions (human liver) and in culture (hepatoma cells, macrophage-like cells, and fibroblasts) using RT-PCR technique was also demonstrated by Nagaoka’s team[39]. The dominant expression of IGF-1A transcripts among other transcripts in the liver itself has been described by other authors[16,40].
A detailed quantitative analysis confirmed the higher expression of mRNAs for A and C isoforms (and of total IGF-1 mRNA) in our HCV infected patients as compared to the control. Expression of all studied IGF-1 transcripts has also been significantly higher in women as compared to men with HCV infection. Koczorowska et al[33] demonstrated a quantitatively higher expression of total IGF-1 mRNA in the precancerous stages of the uterine cervix, as well as the activity of both gene promoters at the stage of intraepithelial neoplasia. Brokaw et al[31] demonstrated a significant relationship between higher expression of IGF-1A isoform expression and progression of ovarian carcinoma. Nevertheless, the role of IGF-1 isoform A remains unclear. It has been argued that peptide Ea arising from it may exert mitogenic effects[41] and inhibit growth of neoplastic cells[42].
The quantitative prevalence of expression manifested by IGF-1 isoforms A and C in HCV-infected livers, as compared to the healthy organ, may point to the influence of HCV on alterations in the splicing profile of the gene in humans. Our other observations suggest the influence of mainly non-structural proteins (NS3 and NS5A) on an increase of IGF-1 protein expression in HCV-infected livers[35] and on the augmentation of IGF-1 P1 and P2 mRNA expression mRNA (unpublished data). Another explanation may involve a compensatory increase in the production of IGF-1 mRNA (particularly that of the IGF-1A isoform) under the effect of locally acting growth factors, proinflammatory cytokines, and an increased regeneration of the organ in CH-C. An increased production of two principal IGF-1 transcripts in rabbit skeletal muscle [i.e., muscle L.IGF-1 (resembling isoform A in the liver) and of mRNA mechano-growth factor (MGF, isoform IGF-1B in rabbits; a homologue of IGF-1C in humans)] was detected under muscle stretching and electrical stimulation[43]. Mechanisms of differential IGF-1 mRNA stability were also described[44,45], including the effects of various RNA-binding proteins (e.g., Hu and hnRNP families) on this process[46]. In the patients analyzed in this study, hepatic expression of IGF-1 mRNA isoforms has been significantly lower upon higher activity of inflammation. The relationships have not been as spectacular as those related to fibrosis. We may have examined an insufficient number of patients with more advanced staging (3 patients with staging 3, one with staging 4). Studies have also documented a negative relationship between expression of all IGF-1 mRNA isoforms, steatosis, and ALT activity. The results indicate a coexistence of the more pronounced inflammatory/necrotic lesions in the liver and a lower hepatic production of IGF-1 mRNA isoforms. Since the literature data contains no references to the role of IGF-1 mRNA isoforms in the progression of HCV-related hepatic diseases, the results seem to be pioneering. The other few reports on the subject mainly concern human tumors[31-34]. In the case of hepatoma and HCV-associated HCC, expression of IGF-1 mRNA was studied, but with no references to specific isoforms[8,13,47]. Su et al[47], using Northern blotting, demonstrated numerous IGF-1 transcripts of various sizes in hepatoma cells and in lines of other neoplastic cells (HepG2, Huh-7, PLC/PRF/5, and Hep3B). Each fragment of neoplastic tissue showed a lower expression of IGF-1 mRNA as compared to control tissue. Using the quantitative technique of real-time RT-PCR (similarly to this study), Tovar et al[8] demonstrated a decrease in IGF-1 mRNA expression at the early stages of HCV infection as compared to control, but with no significant differences between preliminary and advanced stages of HCC in patients. Stefano et al[13], using the RT-PCR technique, demonstrated a comparable quantity of IGF-1 mRNA in patients with CH-C and in the control. Another study on cultured rat hepatocytes, also using the RT-PCR technique, demonstrated a 50-fold increase in the expression of IGF-1B mRNA isoform (a homologue of human isoform C) in the cells of obese animals as compared to the hepatocytes of lean individuals[48]. Armakolas et al[32], examining alternative splicing of IGF-1 in prostate carcinoma, demonstrated overexpression of IGF-1C (MGF) in the cells, thereby suggesting the role of the IGF-1 isoform in the stimulation of cell proliferation.
Expression of IGF-1R transcript proved to be lower in our HCV-infected livers than that in the control, and was accompanied by an increase in at least a portion of studied IGF-1 transcripts in the liver. It is known from earlier studies that, even if normal liver represents an organ with the highest production of IGF-1, it contains almost undetectable levels of IGF-1R mRNA[10]. Moreover, marked expression of the receptor used to be noted mainly in Kupffer, vascular endothelial, and stellate cells, but not in hepatocytes[49,50]. In this study, expression of IGF-1R mRNA in the CH-C group has manifested positive correlation with grading. Therefore, it seems that local production of IGF-1R mRNA does not increase until HCV-associated hepatic lesions become pronounced, which was noted to be accompanied by a reduced production of total IGF-1 mRNA (and of certain isoforms). A lowered expression of IGF-1, coexisting with an increased production of IGF-1R in uterine carcinoma, was also described[33]. In studies on more advanced stages of liver carcinogenesis, an increase in IGF-1R expression was already detected in preneoplastic focal lesions in the liver, in HCC itself, and in cell lines of human hepatoma[5,10]. No such increase was detected in human hepatocyte cultures[11]. Studies by Price et al[6] on the rat model demonstrated a higher expression of IGF-1R mRNA in the control liver as compared to HCC, and it was only the cooperation of two proteins [IGF-1 and hepatocyte growth factor-scatter factor (HGF-SF)] that stimulated mitogenesis of hepatocytes in the animals. In patients with liver cirrhosis and CH-C, increased amounts of IGF-1R transcript were detected as compared to normal liver[12,13]. Tovar et al[8] failed to detect a significant difference in the expression of IGF-1R mRNA in hepatocellular carcinoma as compared to a control in human HCC.
Differences in quantitative expression of IGF-1 mRNA isoforms in HCV-infected livers, as compared to the control, suggest that HCV may induce alteration of the IGF-1 splicing profile. An increase in the grading of HCV-infected livers was linked to decreased IGF-1 mRNA expression, an altered profile of mRNA isoforms, and to an increase in IGF-1R mRNA expression. The demonstration of increased tissue expression of IGF-1R mRNA and the decreased expression level of IGF-1 mRNA isoforms, accentuated in line with increasing liver damage, may be of a prognostic significance.
COMMENTS
Background
Insulin-like growth factor 1 (IGF-1) represents a well-recognized pro-proliferative factor.
Research frontiers
No studies are available on the role played by local expression involving various IGF-1 mRNA isoforms in chronic hepatitis C in vivo.
Innovations and breakthroughs
A quantitative polymerase chain reaction analysis used in the study confirmed the higher expression of total IGF-1 mRNA and of IGF-1 mRNAs isoforms A and C in hepatitis C virus-infected livers as compared to the control.
Applications
The demonstration of decreased expression levels of IGF-1 mRNA isoforms and an increased tissue expression of IGF-1R mRNA, associated with indicators of liver damage (e.g., grading, staging, steatosis, and liver serum enzyme activity), may be of prognostic significance.
Peer-review
This paper is intriguing but presents important clinical limitations, mostly relating to the low number of patients with chronic hepatitis C investigated, to the low prevalence of elderly patients with severe forms of the disease, and to the insufficient number of controls.
Footnotes
Supported by Minister of Education and Science, Warsaw, Poland, No. NN401009437.
Ethics approval: Committee on Bioethics, Poznan University of Medical Sciences, 61-701 Poznan, Poland (No. 22/09).
Institutional animal care and use committee: Not applicable.
Conflict-of-interest: All authors declare that they have no relevant or material financial interests that relate to the research described in this paper.
Data sharing: No additional data available.
Open-Access: This article is an open-access article which was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Peer-review started: August 27, 2014
First decision: September 15, 2014
Article in press: January 8, 2015
P- Reviewer: Liu YX, Sagnelli E S- Editor: Ma YJ L- Editor: Rutherford A E- Editor: Wang CH
References
- 1.de Oliveria Andrade LJ, D’Oliveira A, Melo RC, De Souza EC, Costa Silva CA, Paraná R. Association between hepatitis C and hepatocellular carcinoma. J Glob Infect Dis. 2009;1:33–37. doi: 10.4103/0974-777X.52979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Yang JD, Roberts LR. Hepatocellular carcinoma: A global view. Nat Rev Gastroenterol Hepatol. 2010;7:448–458. doi: 10.1038/nrgastro.2010.100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Wu J, Zhu AX. Targeting insulin-like growth factor axis in hepatocellular carcinoma. J Hematol Oncol. 2011;4:30. doi: 10.1186/1756-8722-4-30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kasprzak A, Adamek A. The insulin-like growth factor (IGF) signaling axis and hepatitis C virus-associated carcinogenesis (review) Int J Oncol. 2012;41:1919–1931. doi: 10.3892/ijo.2012.1666. [DOI] [PubMed] [Google Scholar]
- 5.Tsai TF, Yauk YK, Chou CK, Ting LP, Chang C, Hu CP, Han SH, Su TS. Evidence of autocrine regulation in human hepatoma cell lines. Biochem Biophys Res Commun. 1988;153:39–45. doi: 10.1016/s0006-291x(88)81186-0. [DOI] [PubMed] [Google Scholar]
- 6.Price JA, Kovach SJ, Johnson T, Koniaris LG, Cahill PA, Sitzmann JV, McKillop IH. Insulin-like growth factor I is a comitogen for hepatocyte growth factor in a rat model of hepatocellular carcinoma. Hepatology. 2002;36:1089–1097. doi: 10.1053/jhep.2002.36158. [DOI] [PubMed] [Google Scholar]
- 7.Zhang YC, Wang XP, Zhang LY, Song AL, Kou ZM, Li XS. Effect of blocking IGF-I receptor on growth of human hepatocellular carcinoma cells. World J Gastroenterol. 2006;12:3977–3982. doi: 10.3748/wjg.v12.i25.3977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Tovar V, Alsinet C, Villanueva A, Hoshida Y, Chiang DY, Solé M, Thung S, Moyano S, Toffanin S, Mínguez B, et al. IGF activation in a molecular subclass of hepatocellular carcinoma and pre-clinical efficacy of IGF-1R blockage. J Hepatol. 2010;52:550–559. doi: 10.1016/j.jhep.2010.01.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Yan XD, Yao M, Wang L, Zhang HJ, Yan MJ, Gu X, Shi Y, Chen J, Dong ZZ, Yao DF. Overexpression of insulin-like growth factor-I receptor as a pertinent biomarker for hepatocytes malignant transformation. World J Gastroenterol. 2013;19:6084–6092. doi: 10.3748/wjg.v19.i36.6084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Scharf JG, Dombrowski F, Ramadori G. The IGF axis and hepatocarcinogenesis. Mol Pathol. 2001;54:138–144. doi: 10.1136/mp.54.3.138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Scharf JG, Schmidt-Sandte W, Pahernik SA, Ramadori G, Braulke T, Hartmann H. Characterization of the insulin-like growth factor axis in a human hepatoma cell line (PLC) Carcinogenesis. 1998;19:2121–2128. doi: 10.1093/carcin/19.12.2121. [DOI] [PubMed] [Google Scholar]
- 12.Morali G, Shitrit AB, Eran M, Freier S, Reinus C, Braverman D. Hepatic production of insulin-like growth factors in normal and diseased liver. Hepatogastroenterology. 2005;52:1511–1515. [PubMed] [Google Scholar]
- 13.Stefano JT, Correa-Giannella ML, Ribeiro CM, Alves VA, Massarollo PC, Machado MC, Giannella-Neto D. Increased hepatic expression of insulin-like growth factor-I receptor in chronic hepatitis C. World J Gastroenterol. 2006;12:3821–3828. doi: 10.3748/wjg.v12.i24.3821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Maki RG. Small is beautiful: insulin-like growth factors and their role in growth, development, and cancer. J Clin Oncol. 2010;28:4985–4995. doi: 10.1200/JCO.2009.27.5040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Bell GI, Stempien MM, Fong NM, Rall LB. Sequences of liver cDNAs encoding two different mouse insulin-like growth factor I precursors. Nucleic Acids Res. 1986;14:7873–7882. doi: 10.1093/nar/14.20.7873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Adamo ML. Regulation of insulin-like growth factor I gene expression. Implications for normal and pathological growth. Diabetes Rev. 1995;3:2–27. [Google Scholar]
- 17.Adamo ML, Ben-Hur H, LeRoith D, Roberts CT. Transcription initiation in the two leader exons of the rat IGF-I gene occurs from disperse versus localized sites. Biochem Biophys Res Commun. 1991;176:887–893. doi: 10.1016/s0006-291x(05)80269-4. [DOI] [PubMed] [Google Scholar]
- 18.Simmons JG, Van Wyk JJ, Hoyt EC, Lund PK. Multiple transcription start sites in the rat insulin-like growth factor-I gene give rise to IGF-I mRNAs that encode different IGF-I precursors and are processed differently in vitro. Growth Factors. 1993;9:205–221. doi: 10.3109/08977199309010833. [DOI] [PubMed] [Google Scholar]
- 19.Rotwein P. Two insulin-like growth factor I messenger RNAs are expressed in human liver. Proc Natl Acad Sci USA. 1986;83:77–81. doi: 10.1073/pnas.83.1.77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Temmerman L, Slonimsky E, Rosenthal N. Class 2 IGF-1 isoforms are dispensable for viability, growth and maintenance of IGF-1 serum levels. Growth Horm IGF Res. 2010;20:255–263. doi: 10.1016/j.ghir.2010.03.002. [DOI] [PubMed] [Google Scholar]
- 21.Pell JM, Saunders JC, Gilmour RS. Differential regulation of transcription initiation from insulin-like growth factor-I (IGF-I) leader exons and of tissue IGF-I expression in response to changed growth hormone and nutritional status in sheep. Endocrinology. 1993;132:1797–1807. doi: 10.1210/endo.132.4.8462477. [DOI] [PubMed] [Google Scholar]
- 22.Mittanck DW, Kim SW, Rotwein P. Essential promoter elements are located within the 5’ untranslated region of human insulin-like growth factor-I exon I. Mol Cell Endocrinol. 1997;126:153–163. doi: 10.1016/s0303-7207(96)03979-2. [DOI] [PubMed] [Google Scholar]
- 23.Wang L, Wang X, Adamo ML. Two putative GATA motifs in the proximal exon 1 promoter of the rat insulin-like growth factor I gene regulate basal promoter activity. Endocrinology. 2000;141:1118–1126. doi: 10.1210/endo.141.3.7344. [DOI] [PubMed] [Google Scholar]
- 24.Chew SL, Lavender P, Clark AJ, Ross RJ. An alternatively spliced human insulin-like growth factor-I transcript with hepatic tissue expression that diverts away from the mitogenic IBE1 peptide. Endocrinology. 1995;136:1939–1944. doi: 10.1210/endo.136.5.7720641. [DOI] [PubMed] [Google Scholar]
- 25.Yang S, Alnaqeeb M, Simpson H, Goldspink G. Cloning and characterization of an IGF-1 isoform expressed in skeletal muscle subjected to stretch. J Muscle Res Cell Motil. 1996;17:487–495. doi: 10.1007/BF00123364. [DOI] [PubMed] [Google Scholar]
- 26.Mills P, Lafrenière JF, Benabdallah BF, El Fahime el M, Tremblay JP. A new pro-migratory activity on human myogenic precursor cells for a synthetic peptide within the E domain of the mechano growth factor. Exp Cell Res. 2007;313:527–537. doi: 10.1016/j.yexcr.2006.10.032. [DOI] [PubMed] [Google Scholar]
- 27.Barton ER, DeMeo J, Lei H. The insulin-like growth factor (IGF)-I E-peptides are required for isoform-specific gene expression and muscle hypertrophy after local IGF-I production. J Appl Physiol (1985) 2010;108:1069–1076. doi: 10.1152/japplphysiol.01308.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Matheny RW, Nindl BC, Adamo ML. Minireview: Mechano-growth factor: a putative product of IGF-I gene expression involved in tissue repair and regeneration. Endocrinology. 2010;151:865–875. doi: 10.1210/en.2009-1217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Dluzniewska J, Sarnowska A, Beresewicz M, Johnson I, Srai SK, Ramesh B, Goldspink G, Górecki DC, Zabłocka B. A strong neuroprotective effect of the autonomous C-terminal peptide of IGF-1 Ec (MGF) in brain ischemia. FASEB J. 2005;19:1896–1898. doi: 10.1096/fj.05-3786fje. [DOI] [PubMed] [Google Scholar]
- 30.Quesada A, Micevych P, Handforth A. C-terminal mechano growth factor protects dopamine neurons: a novel peptide that induces heme oxygenase-1. Exp Neurol. 2009;220:255–266. doi: 10.1016/j.expneurol.2009.08.029. [DOI] [PubMed] [Google Scholar]
- 31.Brokaw J, Katsaros D, Wiley A, Lu L, Su D, Sochirca O, de la Longrais IA, Mayne S, Risch H, Yu H. IGF-I in epithelial ovarian cancer and its role in disease progression. Growth Factors. 2007;25:346–354. doi: 10.1080/08977190701838402. [DOI] [PubMed] [Google Scholar]
- 32.Armakolas A, Philippou A, Panteleakou Z, Nezos A, Sourla A, Petraki C, Koutsilieris M. Preferential expression of IGF-1Ec (MGF) transcript in cancerous tissues of human prostate: evidence for a novel and autonomous growth factor activity of MGF E peptide in human prostate cancer cells. Prostate. 2010;70:1233–1242. doi: 10.1002/pros.21158. [DOI] [PubMed] [Google Scholar]
- 33.Koczorowska MM, Kwasniewska A, Gozdzicka-Jozefiak A. IGF1 mRNA isoform expression in the cervix of HPV-positive women with pre-cancerous and cancer lesions. Exp Ther Med. 2011;2:149–156. doi: 10.3892/etm.2010.174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kasprzak A, Szaflarski W, Szmeja J, Andrzejewska M, Przybyszewska W, Kaczmarek E, Koczorowska M, Kościński T, Zabel M, Drews M. Differential expression of IGF-1 mRNA isoforms in colorectal carcinoma and normal colon tissue. Int J Oncol. 2013;42:305–316. doi: 10.3892/ijo.2012.1706. [DOI] [PubMed] [Google Scholar]
- 35.Kasprzak A, Adamek A, Przybyszewska W, Szaflarski W, Sterzyńska K, Seraszek A, Mozer-Lisewska I, Kaczmarek E, Biczysko W. Expression of IGF-I and viral proteins (C, NS3, NS5A) in livers of patients with chronic HCV infection. Adv Clin Exp Med. 2011;20:263–273. [Google Scholar]
- 36.Bedossa P, Poynard T. An algorithm for the grading of activity in chronic hepatitis C. The METAVIR Cooperative Study Group. Hepatology. 1996;24:289–293. doi: 10.1002/hep.510240201. [DOI] [PubMed] [Google Scholar]
- 37.Cicinnati VR, Shen Q, Sotiropoulos GC, Radtke A, Gerken G, Beckebaum S. Validation of putative reference genes for gene expression studies in human hepatocellular carcinoma using real-time quantitative RT-PCR. BMC Cancer. 2008;8:350. doi: 10.1186/1471-2407-8-350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ohtsuki T, Otsuki M, Murakami Y, Maekawa T, Yamamoto T, Akasaka K, Takeuchi S, Takahashi S. Organ-specific and age-dependent expression of insulin-like growth factor-I (IGF-I) mRNA variants: IGF-IA and IB mRNAs in the mouse. Zoolog Sci. 2005;22:1011–1021. doi: 10.2108/zsj.22.1011. [DOI] [PubMed] [Google Scholar]
- 39.Nagaoka I, Someya A, Iwabuchi K, Yamashita T. Expression of insulin-like growth factor-IA and factor-IB mRNA in human liver, hepatoma cells, macrophage-like cells and fibroblasts. FEBS Lett. 1991;280:79–83. doi: 10.1016/0014-5793(91)80208-k. [DOI] [PubMed] [Google Scholar]
- 40.Barton ER. The ABCs of IGF-I isoforms: impact on muscle hypertrophy and implications for repair. Appl Physiol Nutr Metab. 2006;31:791–797. doi: 10.1139/h06-054. [DOI] [PubMed] [Google Scholar]
- 41.Tian XC, Chen MJ, Pantschenko AG, Yang TJ, Chen TT. Recombinant E-peptides of pro-IGF-I have mitogenic activity. Endocrinology. 1999;140:3387–3390. doi: 10.1210/endo.140.7.7044. [DOI] [PubMed] [Google Scholar]
- 42.Chen MJ, Chiou PP, Lin P, Lin CM, Siri S, Peck K, Chen TT. Suppression of growth and cancer-induced angiogenesis of aggressive human breast cancer cells (MDA-MB-231) on the chorioallantoic membrane of developing chicken embryos by E-peptide of pro-IGF-I. J Cell Biochem. 2007;101:1316–1327. doi: 10.1002/jcb.21254. [DOI] [PubMed] [Google Scholar]
- 43.McKoy G, Ashley W, Mander J, Yang SY, Williams N, Russell B, Goldspink G. Expression of insulin growth factor-1 splice variants and structural genes in rabbit skeletal muscle induced by stretch and stimulation. J Physiol. 1999;516(Pt 2):583–592. doi: 10.1111/j.1469-7793.1999.0583v.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Hepler JE, Van Wyk JJ, Lund PK. Different half-lives of insulin-like growth factor I mRNAs that differ in length of 3‘ untranslated sequence. Endocrinology. 1990;127:1550–1552. doi: 10.1210/endo-127-3-1550. [DOI] [PubMed] [Google Scholar]
- 45.Oberbauer AM. The Regulation of IGF-1 Gene Transcription and Splicing during Development and Aging. Front Endocrinol (Lausanne) 2013;4:39. doi: 10.3389/fendo.2013.00039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Lee EK, Gorospe M. Minireview: posttranscriptional regulation of the insulin and insulin-like growth factor systems. Endocrinology. 2010;151:1403–1408. doi: 10.1210/en.2009-1123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Su TS, Liu WY, Han SH, Jansen M, Yang-Fen TL, P’eng FK, Chou CK. Transcripts of the insulin-like growth factors I and II in human hepatoma. Cancer Res. 1989;49:1773–1777. [PubMed] [Google Scholar]
- 48.Tenoutasse S, Van Vliet G, Ledru E, Deal C. IGF-I transcript levels in whole-liver tissue, in freshly isolated hepatocytes, and in cultured hepatocytes from lean and obese Zucker rats. Horm Res. 2003;59:135–141. doi: 10.1159/000069066. [DOI] [PubMed] [Google Scholar]
- 49.Alexia C, Fallot G, Lasfer M, Schweizer-Groyer G, Groyer A. An evaluation of the role of insulin-like growth factors (IGF) and of type-I IGF receptor signalling in hepatocarcinogenesis and in the resistance of hepatocarcinoma cells against drug-induced apoptosis. Biochem Pharmacol. 2004;68:1003–1015. doi: 10.1016/j.bcp.2004.05.029. [DOI] [PubMed] [Google Scholar]
- 50.Jiang Y, Wang L, Gong W, Wei D, Le X, Yao J, Ajani J, Abbruzzese JL, Huang S, Xie K. A high expression level of insulin-like growth factor I receptor is associated with increased expression of transcription factor Sp1 and regional lymph node metastasis of human gastric cancer. Clin Exp Metastasis. 2004;21:755–764. doi: 10.1007/s10585-005-1198-2. [DOI] [PubMed] [Google Scholar]