

Significance
An enigmatic fossil representing the deep-diving, open-ocean whale family Ziphiidae found 740 km inland and at 620 m elevation in West Turkana, Kenya was rediscovered after it went missing for more than 30 years. This stranded whale fossil provides the first constraint on the initiation of east African uplift from near sea level at 17 Ma, limiting the timing and initial elevation of environmental change indicated by geodynamic and climatic modeling, paleosols, isotopes, paleobotany, and the mammalian fossil record. At 17 Ma, elevation was low, rainfall was high, vegetation was forested, and mammalian communities contained immigrants and native African species, including diverse primates. Uplift resulted in increasing aridity and open habitats that drove human evolution.
Keywords: east Africa, Ziphiidae, uplift, drainage, paleoenvironment
Abstract
Timing and magnitude of surface uplift are key to understanding the impact of crustal deformation and topographic growth on atmospheric circulation, environmental conditions, and surface processes. Uplift of the East African Plateau is linked to mantle processes, but paleoaltimetry data are too scarce to constrain plateau evolution and subsequent vertical motions associated with rifting. Here, we assess the paleotopographic implications of a beaked whale fossil (Ziphiidae) from the Turkana region of Kenya found 740 km inland from the present-day coastline of the Indian Ocean at an elevation of 620 m. The specimen is ∼17 My old and represents the oldest derived beaked whale known, consistent with molecular estimates of the emergence of modern strap-toothed whales (Mesoplodon). The whale traveled from the Indian Ocean inland along an eastward-directed drainage system controlled by the Cretaceous Anza Graben and was stranded slightly above sea level. Surface uplift from near sea level coincides with paleoclimatic change from a humid environment to highly variable and much drier conditions, which altered biotic communities and drove evolution in east Africa, including that of primates.
The Turkana ziphiid was found at Loperot in West Turkana, Kenya and described in 1975 by James G. Mead (1), who listed associated fauna, including mollusks, fish, crocodiles, turtles, and mammals, notably primates. Mead (1) detailed the anatomy of the whale fossil, estimated its length in life at some 7 m, and speculated that it was an open-ocean whale that became stranded after swimming up an eastward-flowing river and was then preserved near where it died. After the original publication of this fossil find (1), the specimen went missing until late 2011, when it was rediscovered at Harvard University and returned to the National Museums of Kenya (KNM), where it is curated under the number KNM-LP 52956.
Because the whale was found during the pre-Global Positioning System era, we studied the original 1964 Harvard expedition field notes and catalog to locate the exact site of the ziphiid (2°23′30″ N, 35°52′30″ E by triangulation) (Fig. 1C) in coarse fluvial sandstones and conglomerates of the Lower to Middle Miocene Auwerwer Formation (2–4) ∼7 m below a basalt dated at 17.1 ± 1.0 Ma (4). Although located in an area repeatedly affected by extensional processes since the Cretaceous, the fossil location corresponds to the northern periphery of the Late Cenozoic East African Plateau (EAP) (Fig. 1B). The specimen consists of the rostrum and the ascending processes of the maxillae and premaxillae (length of 82 cm and width of 55 cm) broken from the rest of the skull (1). Five phylogenetically informative characters of KNM-LP 52956 (Fig. 2A and SI Appendix) were scored and entered into a data matrix of 46 characters and 29 fossil and recent taxa (5). A traditional Wagner tree search (one random seed and 10 replications) applied to unweighted and unordered characters yielded 17 most parsimonious trees of 124 steps (Fig. 2B). In all most parsimonious trees, the Turkana ziphiid falls in a derived but unresolved clade with modern Indopacetus, Hyperoodon, and Mesoplodon plus four extinct genera.
Fig. 1.
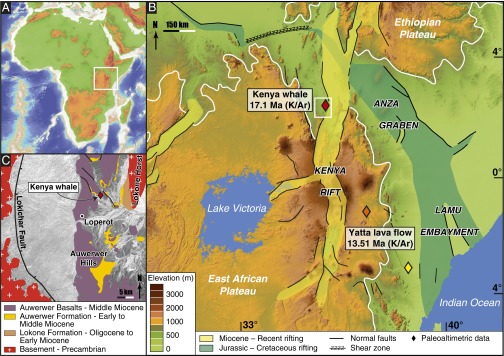
(A) Topography of Africa and bathymetry of the Atlantic and Indian Oceans. The white box indicates the location of the principal map shown in B. (B) Geological setting of the Cenozoic East African Rift System and topography of the present EAP based on a digital elevation model derived from Satellite Radar Topography Mission data. Elevations >1,000 m are enclosed by a white line. Rock ages for the basalt overlaying the beaked whale fossil and the Yatta lava flow phonolites are based on the radioactive decay of potassium (40K) into argon (40Ar). The white box indicates the location of the geological map in C. (C) Geological map of the Oligo-Miocene Lokichar Basin. The red diamond indicates the beaked whale fossil locality (Williams’ Flat) within the Lower to Middle Miocene Auwerwer Formation.
Fig. 2.

(A) Computed tomography rendering of the Turkana ziphiid (KNM-LP 52956) showing 5 of 46 characters from the work by Lambert et al. (5) that could be scored for phylogenetic analysis. 2, Mesorostral groove filled with mesorostral ossification of the vomer with convergent lateral walls of the vomer in the rostrum base area (state 2, derived); 3, mesorostral groove filled with vomer and no mediodorsal contact of the premaxilla (state 0, primitive); 4, prenarial basin absent (state 0, primitive); 27, lacks maxillary alveoli (state 1, derived); 30, rostral prominence lacking (state 0, primitive) (SI Appendix). (B) Beaked whales through time. Ziphiid cladogram calculated using the traditional Wagner tree search phylogenetic analysis program TNT (tree analysis using new technology) (unweighted and unordered; blue dots at nodes indicate clades within Ziphiidae, and numbers indicate Bremer Support values). KNM-LP 52956 falls in a derived clade with modern Indopacetus, Hyperoodon, and Mesoplodon. *Taxa with their earliest records derived from specimens dredged from the sea floor. (Inset) The analysis by Lambert et al. (5) was run with weighted and ordered characters, accounting for topological differences in the resulting trees. Temporal ranges were obtained from the Paleobiology Database (www.paleobiodb.org).
Beaked whales are predicted by molecular clocks to have originated 26.52–35.82 Ma (6). The early record of fossil ziphiids is poor, but at 17.1 ± 1.0 Ma, the Kenyan specimen is currently the most precisely dated ziphiid fossil. Phylogenetic analysis nests the Turkana ziphiid with three modern genera, most notably Mesoplodon, which has species that are estimated to have diverged at 16.6 Ma (6, 7). Thus, the geochronologic constraint provided by the Turkana ziphiid is consistent with molecular predictions.
Ziphiids and Other Cetaceans in Present-Day and Paleorivers
Modern ziphiids are open-ocean, deep-diving specialized suction feeders that prey mainly on squid (8). Phylogenetic analyses incorporating molecular data (9) indicate that they are nested between the freshwater Ganges River dolphin (Platanista) and the Yangtze (Lipotes) plus South American river dolphins (Inia and Pontoporia). The Kenyan whale possesses derived ziphiid characters of the beak, including hyperostosis, reduced gape, and evolutionary loss of teeth (characters not found in any clade of river dolphins). Moreover, the Turkana ziphiid’s total body length falls within the range of modern ziphiids (10) and is up to three times that observed in river dolphins.
A wide variety of present-day marine whales has been observed in rivers, some of which may be associated with seasonal migrations (11, 12). Modern ziphiids occasionally enter rivers, such as occurred in the 2006 stranding of Hyperoodon ampullatus in the Thames River at London. Killer whales (Orcinus orca) have been reported 161 km up the Columbia River at an elevation of 6 m and river depth >10 m (Table 1) (13). In 2007, mother and calf humpback whales (Megaptera novaeangliae) swam 133 km upstream to Sacramento, CA (elevation of 4 m and river depth of ∼10 m) (Table 1) (13). An Antarctic minke whale (Balaenoptera bonaerensis) swam 1,000 km up the Amazon and its tributary the Tapajós, traveling as much as 83 km in 1 day (Table 1) (14). In comparison, South American Sotalia fluvatilis, a river dolphin, has a normal range that reaches an elevation of ∼100 m, where river depths are 10–30 m, 3,600 km up the Amazon drainage from the Atlantic Ocean (Table 1) (15). In each of these cases, the riverine distribution of modern cetaceans is facilitated by sufficient water discharge in low-gradient rivers and the absence of shallow bedrock rapids and waterfalls (15, 16).
Table 1.
Comparison of stranding parameters for marine whales in river systems
| Cetacean species | River | River gradient (cm/km) | River depth (m) | Stranding inland (km) | Elevation (m) | Source |
| H. ampullatus | Thames, United Kingdom | 2.9 | 2–11 | 70 | 2 | — |
| M. novaeangliae | Sacramento, United States | 3.0 | ∼10 | 133 | 4 | 13 |
| O. orca | Columbia, United States | 3.7 | >10 | 161 | 6 | 13 |
| Balaenoptera acutorostrata | Tapajós-Amazon, Brazil | 0.1 | 10–30 | 1,000 | 1 | 14 |
| S. fluvatilis | Amazon, Peru | 2.8 | 10–30 | 3,600 | 100 | 15 |
| Turkana ziphiid | Anza River, Kenya | 4? | Unknown | 600–900 | 24–37 | This study |
Elevation of 24–37 m for the Turkana whale is arbitrary and exceeds the elevation of other marine whales stranded in rivers. The river dolphin S. fluvatilis is included for comparison and shows its distance from the coast at 100-m elevation, although its total elevation range extends to 300 m.
Results and Discussion
Rifting, Paleodrainage Evolution, and Surface Uplift of the EAP.
The fluvial sandstones and conglomerates associated with the Turkana ziphiid are within the Oligo-Miocene Lokichar halfgraben (3, 17), which hosts 7-km-thick sedimentary strata related to a fluvial and freshwater lacustrine depositional environment (18). This basin is superimposed on an area of widespread Cretaceous extension associated with the Anza Graben, a northwest to southeast-oriented rift basin, which is also filled with Paleocene to Miocene fluviolacustrine strata that transition upward into marine units toward the Kenya–Somalia coast in the Lamu Embayment (Fig. 1B) (19, 20). The Anza Graben constitutes a failed rift arm, which once linked rifting in the Central African Shear Zone with extensional processes in eastern Kenya that led to the separation of Madagascar from Africa (19).
The size of the ziphiid, the regional context of the fluvial sediments in which it was found, and the fauna associated with it indicate that the path to its stranding site was a sizable low-gradient river. The ziphiid may have mistakenly entered the river while migrating along the east African coast and was subsequently unable to correct its course. Thus, the precisely dated fossil is not only a remarkable zoogeographic record of stranding, but also, it reflects Miocene drainage patterns directed eastward from the developing EAP and provides an important empirical paleoelevation point that helps to temporally limit EAP surface uplift and Tertiary rifting.
In light of the regional paleotectonic conditions that have been inferred from the Lokichar halfgraben sedimentary fill, it is possible that eastward-directed runoff from the continent and fluvial connectivity between the Indian Ocean and the Turkana region during the Early Miocene were guided by the structural grain and sustained thermal subsidence of the Anza Graben (19). The presence of Late Early Miocene freshwater rays (Myliobatiformes) in the Turkana region (21) also attests to the existence of a well-developed fluvial system at that time (22). These Oligo-Miocene paleoenvironmental conditions are compatible with an overall humid climate with precipitation of >1,000 mm/a (millimeters per year), deep freshwater lakes in the rift basins (18), and a vegetative cover with affinities to the Guinea-Congolia/Zambezia phytogeographic zone and ancient lowland forest (23, 24) also present on the Ethiopian Plateau during that interval (25). Similarly, the phylogenetic analysis of Annonaceae suggests the existence of a trans-African forest and a warmer, moister climate before 16.8 Ma (26) during the Miocene climatic optimum (27).
In summary, the Early Miocene paleoenvironment, where the Turkana ziphiid was stranded, was characterized by low-gradient fluvial conditions in a humid, densely vegetated environment. The fluvial system must have had sufficient discharge and connectivity with the open ocean to provide an unrestricted pathway for the upstream migration of the whale.
A large expanse between West Turkana and the Indian Ocean has no record of sediments indicating the former presence of marine conditions. However, between 23 and 17 Ma, when the whale stranded, high sea stands of approximately +10 m (28) flooded the Lamu Embayment (Fig. 3A) as evidenced by Early Miocene shallow coastal marine sediments of the Baratumu Formation recorded in subsurface cores (20). These sediments correspond in age to the Loperot locality and occur some 50 km west of the town of Garissa (20), bringing the sea closer to the West Turkana region. Regardless of its exact route, the distance the whale swam from the sea to its final stranding was considerable. Taking the Anza Rift axis as a reference line for the paleoriver course and discounting meanders, the limits are 900 km if it entered riverine conditions near the current Indian Ocean coast and 600 km if its journey started at the limit of the Lamu Embayment (Fig. 3A and Table 1). Even if a relatively high gradient of 4 cm/km is assumed for the paleoriver used by the Turkana ziphiid, which is greater than most river gradients where cetaceans have been sighted, the whale would have stranded and died at a paleoelevation between 24 and 37 m (Fig. 3A and Table 1). If true, considering the modern elevation of the fossil location at 620 m, the northern periphery of the EAP must have been uplifted by a minimum of 590 m during the last 17 Ma.
Fig. 3.
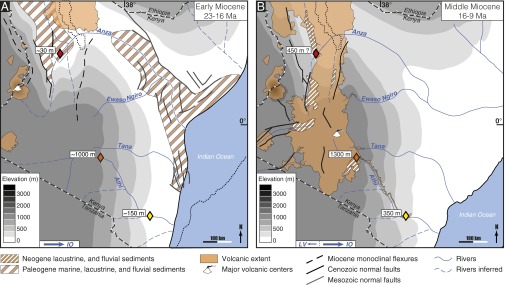
Topographic and drainage evolution in Kenya during the Miocene. Present Indian Ocean coastline, national borders, and Lake Turkana shoreline in the north for orientation. Paleoaltimetric data are shown by diamonds (red, Turkana whale; orange, Yatta lava flow beginning; yellow, Yatta lava flow end). A shows 23–16 Ma: uplift of the EAP to ∼1,000 m in Kenya (33, 34). The plateau area is drained by a major river network, which flows to the east into the Indian Ocean (IO) (29, 30). B shows 16–9 Ma: uplift of the EAP to 1,400 m in Kenya (29). The major river network still flows to the east, but minor rivers also drain to the west through surface downwarps on the EAP interior into the Lake Victoria (LV) basin (29, 30, 46).
The inference of an uplifting EAP after 17 Ma is supported by evidence for the existence of other Middle to Late Miocene eastward-directed paleovalleys that channeled runoff and lava flows away from the EAP. For example, Middle Miocene basaltic and phonolitic lava flows on the eastern margin of the present plateau followed such drainages in the ancestral Ewaso Ngiro, Tana, and Athi Rivers (Fig. 3B) (29, 30). Field evidence for paleoelevation along these eastward drainages was derived from the emplacement of the 13.5-My-old Yatta lava flow (31) along the paleo-Athi River, attesting to phonolitic eruptions before the onset of rifting in the Central EAP at an elevation of 1,400 m (32). This paleotopographic constraint is in good spatiotemporal agreement with modeled elevations of the EAP based on reconstructions of mantle flow (33) and metamorphic density changes (34). The Turkana ziphiid, thus, provides the second line of empirical field evidence for paleoelevation in northern Kenya and the first maximum time constraint for the onset of uplift on the northeastern flank of the EAP. Together with the inferences from the Yatta lava flow, the Turkana ziphiid helps to constrain the onset of uplift of the EAP to between 17 and 13.5 Ma.
Linking Paleoenvironmental Changes to EAP Surface Uplift.
The particular significance of constraining uplift and defining paleoenvironments in this case is the elucidation of evolutionary drivers leading to humans. Terrestrial mammals found with the Turkana ziphiid postdate the Early Miocene faunal interchange with Eurasia, which introduced a number of artiodactyls, rhinos, and carnivores and changed the community composition in which older residents, such as primates and elephants, lived (35, 36). Primates of the Turkana Basin coeval with the Turkana ziphiid include Limnopithecus, Rangwapithecus, Noropithecus (37) and the enigmatic possible hominoids Afropithecus, Turkanapithecus, and Simiolus (Fig. 4) (38).
Fig. 4.
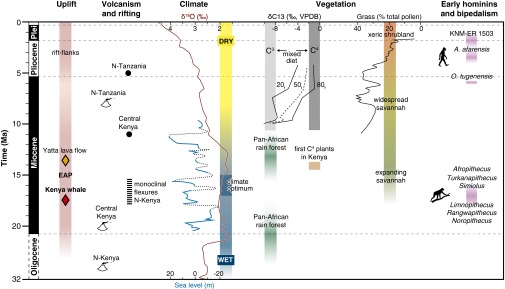
Cenozoic uplift chronology, climatic conditions, paleoenvironment, and early hominin evolution. Paleoaltimetric data (diamonds) of Early to Middle Miocene onset of uplift correlate with major climatic and environmental shifts. Data compilation as follows: onset of volcanism and rifting in Kenya (47); Miocene sea-level fluctuations (48); global deep-sea oxygen isotopes, including the Miocene climate optimum (27); vegetation shifts based on carbon isotopes from herbivore teeth (Xf percentile δ13C values for fossil populations) (45) and pollen data (42) in north Kenya; pan-African rain forest expansion–isolation events (26) and savannah expansion (44); and early primates and bipedalism in northern Kenya (37, 38, 49).
The presence of a substantial low-gradient river that drained the EAP region first to the north into the Lokichar freshwater basin and then to the southeast toward the Lamu Embayment (Fig. 3) in today’s driest region of northern Kenya, provides compelling evidence for a dramatic environmental change since the Early Miocene. Such a change is compatible with the results of climate models, which link surface uplift of east Africa to modified airflow patterns, aridification, and the establishment of the east African monsoon (39). Uplift of the EAP starting between 17 and 13.5 Ma correlates with a shift toward aridity accompanied by the expansion of C4 plants in east Africa between 14 and 11 Ma (40–42), a change from grassy woodland to wooded grassland documented in paleosols (43), and the expansion of a grass-dominated savannah biome that started in the Middle Miocene and became widespread during the Late Miocene (Fig. 4) (44). Furthermore, recent δ13C isotopic studies on herbivore teeth reveal changes from C3-dominated to mixed C3/C4- or C4-dominated diets from the Late Miocene through the Pliocene in northern Kenya (Fig. 4) (45). Although the age of the Turkana whale predates the aridification that accompanies major vegetation and faunal changes during the Miocene and Pliocene of Africa, including the origin of hominins, its occurrence shows the protracted presence of well-watered, lowland habitats far inland from the coast at an early stage of east African community evolution after major faunal exchange with Eurasia but before the EAP uplift.
Materials and Methods
Rediscovery of the Kenya Beaked Whale (KNM-LP 52956).
The Turkana ziphiid specimen was collected by J. G. Mead, a member of the 1964 Harvard–Kenya Expedition under the leadership of Bryan Patterson. The locality was identified as Williams’ Flat in field notes but referred to by Mead as the Open Pit Turtle Mine, because the specimen was identified in the field as a large turtle (cataloged 14–64K). When Mead was a member of the expedition, he was an undergraduate student at Yale. He then undertook graduate studies at the University of Texas followed by the University of Chicago. He borrowed the specimen for his research and returned it to Harvard in the late 1960s. In 1972, Mead became an assistant curator at the Smithsonian Institution and published his paper on the Turkana ziphiid in 1975 (1). Patterson died in 1979, and the whereabouts of the specimen could no longer be determined. L.L.J. first searched for the specimen in 1980 when he was Head of Paleontology at the KNM. Since then, several searches were made at the Museum of Comparative Zoology, Harvard, the Smithsonian Institution, and the KNM.
In late 2011, a relatively new collections manager at Harvard, Jessica Cundiff, responded to inquiries from L.L.J. with a diligent search. Over time, larger specimen storage at the Museum of Comparative Zoology had apparently changed locations. Cundiff located the Turkana ziphiid in what had formerly been the office of the renowned paleontologist Stephen Jay Gould but was at the time, being used for temporary storage during remodeling. After its rediscovery at Harvard, the late Farish A. Jenkins Jr. immediately approved its transfer to KNM. L.L.J. arranged for its transfer first to Texas for digital scanning, after which he returned it in 2012 to the KNM, where it is now housed and curated under the number KNM-LP 52956.
Scanning and Rendering the Fossil Specimen.
The fossil was surface-scanned with a NextEngine 3D Laser Surface Scanner (Fig. 2A) at the Southern Methodist University Visualization Laboratory. It was scanned at the High-Resolution X-Ray Computed Tomography Facility at the University of Texas in three runs comprising the right posterior, the left posterior, and the beak portions of the preserved skull. The beak portion was scanned as two parallel pieces separated at the point of a histological sample taken by J. G. Mead in 1975 (1). The scans produced three datasets of 211, 208, and 249 slices, respectively, all using the same scanning parameters at an interpixel spacing of 0.292 mm and interslice spacing of 1.45 mm. The datasets and detailed scanning and processing parameters are archived at www.digimorph.org/specimens/Turkana_ziphiid. The slice data were analyzed with Amira 5.3 (FEI Corporation) and ImageJ (www.nih.gov) to determine relationships of bones. Threshold-based isosurface reconstruction was performed in Amira, and the resultant surface models were saved in Wavefront Technologies object format (.obj). These models were then imported into Lightwave 9.6 and aligned to generate figures and virtual reality object (SI Appendix, Movie S1).
Phylogenetic Analysis.
In addition to a TNT traditional Wagner tree search, a second analysis using the Branch and Bound algorithm in PAUP confirmed the topology obtained from TNT. Bremer support was calculated using TNT with suboptimal trees retained. In the calculated Majority Rule tree, the Turkana ziphiid fell with >75% support in an unresolved Hyperoodontinae along with Indopacetus, Mesoplodon, Africanacetus, and Ihlengesi, with which the Turkana ziphiid shares the same scores for all five characters that could be observed. Lambert et al. (5) ran their analysis with weighted and ordered characters. Our analyses were run unweighted and unordered, which accounts for the topological differences in the resulting trees (compare with Fig. 2B, Inset) (notably, that Ziphiidae is paraphyletic when characters are unweighted and unordered). The positions of Tasmacetus and Nazcacetus also differ markedly, with these taxa forming a clade sister to Imocetus, Tusciziphius, and Globicetus. Jackknifing the phylogeny reveals very poor support for all interclade relationships. Temporal ranges in Fig. 2 were obtained from the Paleobiology Database (www.paleobiodb.org).
Supplementary Material
Acknowledgments
The rediscovery and repatriation of KNM-LP 52956 was an extended effort joined by E. Mbua and the curatorial staff of the National Museums of Kenya, L. J. Flynn, J. G. Mead, M. Leakey, R. Leakey, A. K. Behrensmeyer, J. Cundiff, and the late Farish A. Jenkins Jr. B. Jacobs, A. Grossman, C. Liutkus-Pierce, C. MacLeod, and C. Jenkins provided valuable help and comments. We thank C. Potter and J. Ososky for guiding us through the Smithsonian collections and R. MacPhee (American Museum of Natural History), E. Fordyce (University of Otago), and O. Mateus (Museu da Lourinhã) for access to specimens. We also thank J. G. Mead for encouragement, providing field data, and cooperating at every turn. We thank two anonymous reviewers for their comments. Funding was provided by the Institute for the Study of Earth and Man at Southern Methodist University (SMU) and SMU Engaged Learning (to A.L.), and the Research Training Group GRK1364 of the German Research Foundation (DFG).
Footnotes
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Data deposition: The 3D digital model of KNM-LP 52956 reported in this paper has been archived at www.digimorph.org/specimens/Turkana_ziphiid.
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1421502112/-/DCSupplemental.
References
- 1.Mead JG. A fossil beaked whale (Cetacea: Ziphiidae) from the Miocene of Kenya. J Paleontol. 1975;45(4):745–751. [Google Scholar]
- 2.Boschetto HB, Brown FH, McDougall I. Stratigraphy of the Lothidok Range, northern Kenya, and K/Ar ages of its Miocene primates. J Hum Evol. 1992;22(1):47–71. [Google Scholar]
- 3.Tiercelin J-J, et al. Hydrocarbon potential of the Meso-Cenozoic Turkana Depression, northern Kenya. I. Reservoirs: Depositional environments, diagenetic characteristics, and source rock–reservoir relationships. Mar Pet Geol. 2004;21(1):41–62. [Google Scholar]
- 4.Ducrocq S, et al. New Oligocene vertebrate localities from northern Kenya (Turkana Basin) J Vertebr Paleontol. 2010;30(1):293–299. [Google Scholar]
- 5.Lambert O, de Muizon C, Bianucci G. The most basal beaked whale Ninoziphius platyrostris Muizon, 1983: Clues on the evolutionary history of the family Ziphiidae (Cetacea: Odontoceti) Zool J Linn Soc. 2013;167(4):569–598. [Google Scholar]
- 6.McGowen MR, Spaulding M, Gatesy J. Divergence date estimation and a comprehensive molecular tree of extant cetaceans. Mol Phylogenet Evol. 2009;53(3):891–906. doi: 10.1016/j.ympev.2009.08.018. [DOI] [PubMed] [Google Scholar]
- 7.Dalebout ML, Steel D, Baker CS. Phylogeny of the beaked whale genus Mesoplodon (Ziphiidae: Cetacea) revealed by nuclear introns: Implications for the evolution of male tusks. Syst Biol. 2008;57(6):857–875. doi: 10.1080/10635150802559257. [DOI] [PubMed] [Google Scholar]
- 8.Geisler JH, McGowen MR, Yang G, Gatesy J. A supermatrix analysis of genomic, morphological, and paleontological data from crown Cetacea. BMC Evol Biol. 2011;11(2011):112. doi: 10.1186/1471-2148-11-112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Madsen PT, Aguilar de Soto N, Tyack PL, Johnson M. Beaked whales. Curr Biol. 2014;24(16):R728–R730. doi: 10.1016/j.cub.2014.06.041. [DOI] [PubMed] [Google Scholar]
- 10.MacLeod CD. How big is a beaked whale? A review of body length and sexual size dimorphism in the family Ziphiidae. J Cetacean Res Manage. 2006;7(3):310–318. [Google Scholar]
- 11.Mitchell E, Kozicki VM. Autumn stranding of a Northern Bottlenose Whale (Hyperoodon ampullatus) in the Bay of Fundy, Nova Scotia. J Fish Res Board Can. 1975;32(7):1019–1040. [Google Scholar]
- 12.MacLeod CD, Pierce GJ, Santos MB. Geographic and temporal variations in strandings of beaked whales (Ziphiidae) on the coasts of the United Kingdom and the Republic of Ireland 1800-2002. J Cetacean Res Manag. 2004;6(1):79–86. [Google Scholar]
- 13.Gulland FMD, et al. Health assessment, antibiotic treatment, and behavioural responses to herding efforts of a cow-calf pair of humback whales (Megaptera novaeangliae) in the Sacramento River Delta, California. Aquat Mamm. 2008;34(2):182–192. [Google Scholar]
- 14.Balensiefer DC, Marcondes MCC, Pretto DJ, Cypriano-Souza AL, Luna FO. Antarctic minke whale (Balaenoptera bonaerensis, Burmeister, 1867) in the Tapajós River, Amazon Basin, Brazil. Aquat Mamm. 2014;40(2):201–206. [Google Scholar]
- 15.Gomez-Salazar C, et al. Update on the freshwater distribution of Sotalia in Colombia, Ecuador, Peru, Venezuela and Suriname. LAJAM. 2010;8(1-2):171–178. [Google Scholar]
- 16.Hrbek T, et al. A new species of river dolphin from Brazil or: How little do we know our biodiversity. PLoS ONE. 2014;9(1):e83623. doi: 10.1371/journal.pone.0083623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Morley CK, et al. Tectonic evolution of the northern Kenya Rift. J Geol Soc London. 1992;149(3):333–348. [Google Scholar]
- 18.Talbot MR, et al. Hydrocarbon potential of the Meso-Cenozoic Turkana Depression, northern Kenya. II. Source rocks: Quality, maturation, depositional environments and structural control. Mar Pet Geol. 2004;21(1):63–78. [Google Scholar]
- 19.Bosworth W, Morley CK. Structural and stratigraphic evolution of the Anza rift, Kenya. Tectonophysics. 1994;236(1-4):93–115. [Google Scholar]
- 20.Nyagah K. Stratigraphy, depositional history and environments of deposition of Cretaceous through Tertiary strata in the Lamu Basin, southeast Kenya and implications for reservoirs for hydrocarbon exploration. Sediment Geol. 1995;96(1-2):43–71. [Google Scholar]
- 21.Leakey RE, Leakey MGA. New Miocene hominoid from Kenya. Nature. 1986;324(6093):143–146. [Google Scholar]
- 22.Feibel CS. Freshwater stingrays from the Plio-Pleistocene of the Turkana Basin, Kenya and Ethiopia. Lethaia. 1993;26(4):359–366. [Google Scholar]
- 23.Vincens A, Tiercelin J-J, Buchet G. New Oligocene-early Miocene microflora from the southwestern Turkana Basin: Paleoenvironmental implications on the northern Kenya Rift. Paleogeogr Paleocl. 2006;239(3-4):470–486. [Google Scholar]
- 24.Linder HP. The evolution of African plant diversity. Front Ecol Evol. 2014;2(38):1–14. [Google Scholar]
- 25.Pan AD, Jacobs BF, Currano ED. Dioscoreaceae fossils from the late Oligocene and early Miocene of Ethiopia. Bot J Linn Soc. 2014;175(1):17–28. [Google Scholar]
- 26.Couvreur TLP, Chatrou LW, Sosef MSM, Richardson JE. Molecular phylogenetics reveal multiple tertiary vicariance origins of the African rain forest trees. BMC Biol. 2008;6:54–63. doi: 10.1186/1741-7007-6-54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zachos J, Pagani M, Sloan L, Thomas E, Billups K. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science. 2001;292(5517):686–693. doi: 10.1126/science.1059412. [DOI] [PubMed] [Google Scholar]
- 28.Miller KG, Mountain GS, Wright JD, Browning JV. A 180-million-year record of sea level and ice volume variations from continental margin and deep-sea isotopic records. Oceanography (Wash DC) 2011;24(2):40–53. [Google Scholar]
- 29.Wichura H, Bousquet R, Oberhänsli R, Strecker MR, Trauth M. Evidence for middle Miocene uplift of the East African Plateau. Geology. 2010;38(6):543–546. [Google Scholar]
- 30.Veldkamp A, Schoorl JM, Wijbrans JR, Claessens L. Mount Kenya volcanic activity and the Late Cenozoic landscape reorganisation in the upper Tana fluvial system. Geomorphology (Amst) 2012;145-146:19–31. [Google Scholar]
- 31.Veldkamp A, et al. Late Cenozoic fluvial dynamics of the River Tana, Kenya, an uplift dominated record. Quat Sci Rev. 2007;26(22-24):2897–2912. [Google Scholar]
- 32.Wichura H, Bousquet R, Oberhänsli R. Emplacement of the mid-Miocene Yatta lava flow, Kenya: Implications for modelling long channelled lava flows. J Volcanol Geotherm Res. 2010;198(3-4):325–338. [Google Scholar]
- 33.Moucha R, Forte AM. Changes in African topography driven by mantle convection. Nat Geosci. 2011;4(10):707–712. [Google Scholar]
- 34.Duesterhoeft E, Bousquet R, Wichura H, Oberhänsli R. Anorogenic plateau formation: The importance of density changes in the lithosphere. J Geophys Res. 2012;117(B7):B07204. [Google Scholar]
- 35.Kappelman J, et al. Oligocene mammals from Ethiopia and faunal exchange between Afro-Arabia and Eurasia. Nature. 2003;426(6966):549–552. doi: 10.1038/nature02102. [DOI] [PubMed] [Google Scholar]
- 36.Rasmussen DT, Gutierrez M. Mammalian fauna from the late Oligocene of northwestern Kenya. Palaeontogr Abt A. 2009;288:1–52. [Google Scholar]
- 37.Grossman A, Liutkus-Pierce C, Kyongo B, M’Kirera F. New fauna from Loperot contributes to the understanding of Early Miocene catarrhine communities. Int J Primatol. 2014;2014(35):1253–1274. [Google Scholar]
- 38.Leakey M, Grossman A, Gutiérrez M, Fleagle JG. Faunal change in the Turkana Basin during the late Oligocene and Miocene. Evol Anthropol. 2011;20(6):238–253. doi: 10.1002/evan.20338. [DOI] [PubMed] [Google Scholar]
- 39.Sepulchre P, et al. Tectonic uplift and Eastern Africa aridification. Science. 2006;313(5792):1419–1423. doi: 10.1126/science.1129158. [DOI] [PubMed] [Google Scholar]
- 40.Morgan ME, Kingston JD, Marino BD. Carbon isotopic evidence for the emergence of C4 plants in the Neogene from Pakistan and Kenya. Nature. 1994;367(6459):162–165. [Google Scholar]
- 41.Kingston JD, Hill A, Marino BD. Isotopic evidence for neogene hominid paleoenvironments in the Kenya rift valley. Science. 1994;264(5161):955–959. doi: 10.1126/science.264.5161.955. [DOI] [PubMed] [Google Scholar]
- 42.Feakins SJ, et al. Northeast African vegetation change over 12 m.y. Geology. 2013;41(3):295–298. [Google Scholar]
- 43.Retallack GJ, Dugas DP, Bestland EA. Fossil soils and grasses of a middle miocene East african grassland. Science. 1990;247(4948):1325–1328. doi: 10.1126/science.247.4948.1325. [DOI] [PubMed] [Google Scholar]
- 44.Jacobs BF. Palaeobotanical studies from tropical Africa: Relevance to the evolution of forest, woodland and savannah biomes. Philos Trans R Soc Lond B Biol Sci. 2004;359(1450):1573–1583. doi: 10.1098/rstb.2004.1533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Uno KT, et al. Late Miocene to Pliocene carbon isotope record of differential diet change among East African herbivores. Proc Natl Acad Sci USA. 2011;108(16):6509–6514. doi: 10.1073/pnas.1018435108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.King BC. Structural evolution of the Gregory rift valley. In: Bishop WW, editor. Geological Background to Fossil Man. Geology Society London; London: 1978. p. 2254. [Google Scholar]
- 47.Baker BH, Wohlenberg J. Structure and evolution of the Kenya rift valley. Nature. 1971;229(5286):538–542. doi: 10.1038/229538a0. [DOI] [PubMed] [Google Scholar]
- 48.Komintz MA, et al. Late Cretaceous to Miocene sea-level estimates from New Jersey and Delaware coastal plain coreholes: An error analysis. Basin Res. 2008;20(2):211–226. [Google Scholar]
- 49.Richmond BG, Jungers WL. Orrorin tugenensis femoral morphology and the evolution of hominin bipedalism. Science. 2008;319(5870):1662–1665. doi: 10.1126/science.1154197. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.