

Abstract
Background
The AP2/ERF family includes a large number of developmentally and physiologically important transcription factors sharing an AP2 DNA-binding domain. Among them DREB1/CBF and DREB2 factors are known as master regulators respectively of cold and heat/osmotic stress responses.
Experimental Approaches
The manual annotation of AP2/ERF family from Eucalyptus grandis, Malus, Populus and Vitis genomes allowed a complete phylogenetic study for comparing the structure of this family in woody species and the model Arabidopsis thaliana. Expression profiles of the whole groups of EgrDREB1 and EgrDREB2 were investigated through RNAseq database survey and RT-qPCR analyses.
Results
The structure and the size of the AP2/ERF family show a global conservation for the plant species under comparison. In addition to an expansion of the ERF subfamily, the tree genomes mainly differ with respect to the group representation within the subfamilies. With regard to the E. grandis DREB subfamily, an obvious feature is the presence of 17 DREB1/CBF genes, the maximum reported to date for dicotyledons. In contrast, only six DREB2 have been identified, which is similar to the other plants species under study, except for Malus. All the DREB1/CBF and DREB2 genes from E. grandis are expressed in at least one condition and all are heat-responsive. Regulation by cold and drought depends on the genes but is not specific of one group; DREB1/CBF group is more cold-inducible than DREB2 which is mainly drought responsive.
Conclusion
These features suggest that the dramatic expansion of the DREB1/CBF group might be related to the adaptation of this evergreen tree to climate changes when it expanded in Australia.
Introduction
Native to Australia, Eucalyptus tree species are among the fastest growing woody plants in the world and represent about 8% of all planted forest with over 18 million hectares grown in 90 countries [1]. Over the last few decades, Eucalyptus trees became one of the main wood fiber crops and a cost-effective source of lignocellulosic biomass for energy production. Market favorites for pulpwood are E. grandis, E. urophylla and their hybrids, grown in tropical and subtropical regions, and E. globulus, planted in temperate regions. Frost tolerance is limiting their expansion in temperate climates, such as in Southern Europe or Southeastern USA [2]. Being evergreen and evergrowing, the Eucalyptus trees are exposed to cold while growing and all the organs may have to cope with frost.
The 605 Mb E. grandis genome sequence released in January 2011 including about 96% of the expressed gene loci [3] now allows genome-wide investigations. In the present study, this resource was surveyed to manually annotate the AP2/ERF (APETALA2/Ethylene-Responsive element binding Factor) family which is characterized by a highly conserved AP2-DNA binding domain [4, 5]. This transcription factor family is known to play a key role in various developmental and adaptive processes in plants, in particular through biotic and abiotic stress response [6, 7]. Ranging between 132 and 292 genes, this family is one of the largest groups of transcription factors [8, 9] and has been classified into subfamilies [10]. The AP2 subfamily contains proteins exhibiting two AP2/ERF domains and known to be involved in the regulation of developmental processes [11–13]. The RAV (Related to ABI3/VP1) proteins which contain two different DNA-binding domains (AP2/ERF and B3) are regulated by ethylene [14] or brassinosteroids [15] and also contribute to biotic and abiotic stress responses [16]. Exhibiting only one AP2/ERF domain, the ERF and DREB (Dehydration Responsive Element Binding) proteins are key regulators of plant responses to biotic and abiotic stresses [17, 18]. The ERF subfamily includes a large number of proteins binding to the GCC box [19], an ethylene responsive element located in the promoter of pathogenesis-related genes [20]. The DREB factors which recognize the cis-acting sequences CRT (C-Repeat) or DRE (Dehydration Responsive Element) located in the promoters of abiotic stress responsive genes [10]. The DREB subfamily has been divided into six groups (A1-A6) which includes the A1 and A2 groups, commonly known as DREB1/CBF and DREB2 [10].The DREB1/CBF genes are mainly known as cold-responsive whereas most of the DREB2 genes have generally been reported as responsive to water stress or heat-shock. However, there is a growing evidence that the stress regulation of DREB1/CBF and DREB2 genes differs according to the plant species [6, 7],[21], [22, 23]. The demonstration of a functional role of Eucalyptus CBF genes in abiotic stress tolerance was achieved in our hands through overexpression in the hybrid E. urophylla x E. grandis [24]
For the first time, the paper presents a comparison of phylogenetic structure of AP2/ERF families based on manually annotated sequences from four woody plants (Eucalyptus, Populus, Malus and Vitis) and Arabidopsis. Since the number of CBF gene copies was found to be genetically related to stress tolerance [25], the present study may provide new information on the distinct evolution of these species to cope with different environments. The second part of this paper focuses on the DREB1/CBF and DREB2 which are the most described groups with regard to stress response. Interestingly, the comparison of DREB groups from woody species reveals an over-representation of DREB1/CBF members in E. grandis genome, together with a moderate number of DREB2 genes. In addition, gene expression of all the members of DREB1/CBF and DREB2 groups in response to various stresses or in different tissues is presented. The data, which raise the question about specificity of these groups in stress response (cold/heat/drought), are discussed in the context of the adaptive evolution of Eucalyptus.
Materials and Methods
Source of genome databases
The annotated E. grandis genome sequence (V1.1) is available at Phytozome website (http://www.phytozome.net/eucalyptus.php). The genomic sequences of A. thaliana AP2/ERF family were down-loaded from the DATF (Database of Arabidopsis Transcription Factors) database website (http://datf.cbi.pku.edu.cn) [26]. The genomic sequences of the AP2/ERF family genes from Vitis, Populus and Malus were obtained from the Phytozome website and from published data [27], [28], [29], [23]. For each of these woody species, the manual annotation of DREB1/CBF and DREB2 groups was based both on the search of conserved binding domains and the analysis of the complete protein sequence.
Finally, the RNAseq database (http://anjie.bi.up.ac.za/eucGenIE/EucGenIE.html) was used for investigating DREB1 and DREB2 gene expression in different tissues. Raw expression data were obtained from the Eucalyptus genome integrative explorer (http://eucgenie.bi.up.ac.za) [30]. Heat map and hierarchical clustering of genes expression profiles were performed by using expander software (ACGT, http://www.cs.tau.ac.il).
Identification and annotation of E. grandis AP2/ERF proteins
Different approaches were used for the identification and annotation of all E. grandis AP2/ERF family members. First, the PS51032 domain of Prosite and alternatively the Arabidopsis AP2/ERF domains were used as query sequences against the whole translated genome of E. grandis. Because some predicted E. grandis AP2/ERF proteins were not detected in the proteome of Arabidopsis, Vitis, Malus or Populus, a manual annotation of the E. grandis genomic sequences was performed by combining a search of homology (http://blast.ncbi.nlm.nih.gov/), and gene prediction (http://augustus.gobics.de/). In addition, a search for new putative paralogs was performed using SCIPIO, an alignment-based tool [31]. Previously predicted and newly discovered sequences were scanned for the presence of characteristic domains (AP2 and B3) using PROSITE patterns and profiles (http://www.expasy.org/prosite/) or Pfam (http://pfam.sanger.ac.uk/). If a domain (or part of) was missing, a manual annotation was performed to complete or correct the predicted sequence. Finally, the structure and genome localization of each gene was determined by using the SCIPIO program on the resulting exhaustive set of EgrAP2/ERF family members.
Manual annotation and mapping of EgrDREB1/CBF genes
In addition to the automatically predicted sequences, the presence of the CBF signatures (PKK/RPAGRxKFxETRHP and DSAWR) [32] was checked using DiALign on the complete translated E. grandis genome. Following the EUCAGEN consortium recommendation, the EgrCBF sequences were named according to their relative position on the classified scaffolds (1 to 11).Then, the cDNA sequences of four previously described E. gunnii CBFs [33] were compared to the E. grandis genome by using the SPLIGN program proposed on the NCBI website for annotating the genomes of higher eukaryotes [34]. At this stage, the available E. grandis genome may still contain a number of sequencing errors leading to falsely assigned codons [35]. Indeed, some of the predicted sequences appeared to be incorrect since they exhibit wrong positions for the start and/or stop codons or unlikely intronic sequences, compared to typical CBF sequences [36], [37]. To validate the hypothesis of erroneous predictions, new sequencing experiments were performed on DNA and cDNA sequences and analyzed to propose curate sequences. Leaving out the synonym “DREB1” for simplification, the validated EgrCBF proteins were classified according to the E. gunnii orthologs by using the EMBOSS Needle program.
Phylogenetic tree construction
Multiple alignments were prepared using ClustalW [38] using default parameters (gap opening penalty = 10, gap extension penalty = 0.1) and further inspected and visually adjusted with BioEdit [39]. The resulting alignments of complete protein sequences were used in MEGA (version 5) [40] for the construction of phylogenetic trees based on Maximum Likelihood according to Jones-Taylor-Thornton model with uniform rates among sites and complete deletion of gaps/missing data. Bootstrap values were calculated from 100 replicates [41].
Plant material and stress treatments for gene expression analysis
E. grandis plants grown from commercial seeds were obtained from Australia (www.b-and-t-world-seeds.com) and E. gunnii x E. dalrympleana (clone 208) hybrid plants issued from cuttings were provided by FCBA (France). One year-old plants from the two species were cultivated under standard conditions of 25°C day/22°C night, 16 h day-length and 115 μmoles.m2.s-1 light intensity (Lumilux Daylight 58W, Osram, München, Germany). The relative humidity was kept at approximately 80% and plants were watered as needed.
To quantify CBF and DREB2 transcription in response to stressful conditions, the first experiment was conducted on E. gunnii x E. dalrympleana plants directly transferred from 22°C to 4°C for a 2h-light period followed by a 6h-dark phase, according to previous experiments in our hands [42]. The control plants were kept at 25°C/light for 2h and then at 25°C/dark for 6h. Each sample was composed of all the leaves harvested from two individual plants and pooled for RNA extraction. Transcript quantification was performed on three independent samples (biological replicates).
According to our previous data on Eucalyptus simplified material [43], the second experiment was performed on detached leaves which were submitted to different stress conditions. Thirty leaves (from 10 individual E. grandis plants) were harvested to form three independent samples of ten leaves (biological replicates). The samples were kept for 2h under light followed by a 6h-dark phase, before RNA extraction (on the pool of 10 leaves). For control, heat and cold stress, the petioles of the detached leaves were kept immersed in a liquid medium (¼ MS) in a culture chamber. The stressed samples were submitted either to a cold-shock at 4°C or to a heat-shock at 45°C, while the control samples were kept at 25°C. For drought stress, the detached leaves were left on Whatman paper inside an incubator at 25°C using the same enlightening protocol.
Quantification of gene expression through Real-Time PCR (RT-qPCR)
RNA extraction was performed using a modified protocol of the SV Total RNA Isolation System (Promega, France). The modification consisted in complementing the NTES buffer (0.1 M NaCl; 0.01 M Tris/HCl; pH 7.5; 1 mM EDTA) with 2% polyvinylpyrrolidone (PVP) and 2% 2-mercaptoethanol [44]. Also an additional washing step was included to improve the RNA purity. Using SuperScript II and random primers (Invitrogen, France), cDNA was produced from 3 μg of extracted RNA according to the manufacturer’s instructions. Based on the corrected EgrCBF annotation, oligonucleotide primers were designed to specifically amplify the different members of the family (S1 Table).
Due to the very high similarity between the EgrCBF genes, the design of specific primers for an efficient RT-qPCR amplification of each paralog was challenging, and additional control steps were needed to confirm that the PCR amplification is specific. To check sequence homology, each primer sequence was blasted against both E. grandis and E. gunnii genomes. The use of DNA calculator (http://www.sigma-genosys.com/calc/DNACalc.asp) provided a technical validation for each primer. The final step for controlling the specificity of amplification was the analysis of the PCR products by gel-electrophoresis and dissociation curves.
The RT-qPCR reactions were performed in 10 μL of SYBR Green Master mix (Applied Biosystems), with 100 ng of cDNA and 300 nM of each primer. Three replicates of each cDNA were run in an ABI PRISM 7900HT 295 Sequence Detection System (Applied Biosciences, France) using a program including a first step (50°C for 2 min and 95°C for 10 min) followed by 40 cycles (95°C/15 s and 61°C/1 min). Serine/threonine-protein phosphatase 2A PP2A1 (Eucgr.B03386) and PP2A3 (Eucgr.B03031), IDH (Eucgr.F02901), SAND (Eucgr.B02502) and EF1A (Eucgr.B02473) were used together as reference genes for the normalization of RNA steady-state level, and the relative changes in gene expression were quantified using the 2-ΔΔ Ct method [45].
The relative transcript abundances are presented as the mean expression Log10 values for the experiments based on whole plants. For the experiments on detached leaves, the relative expression values are directly presented as Log10 of induction rate for each stress condition in comparison to the control. In both cases, the experiments were performed on three independent biological replicates (each sample composed of material from individual plants). The mean values were analyzed for significance using the statistical T-test (p value = 0.05).
Results
Annotation and structure of the AP2/ERF transcription factor family from E. grandis in comparison with other plant species
From the 36376 loci encoding putative proteins in the E. grandis genome [3, 46], 209 were identified as AP2/ERF-like proteins based on the presence of an AP2 DNA-binding domain. When several sequences were predicted at the same genomic locus, the selection of a unique sequence was based on the highest similarity with previously described orthologs. Some predicted genes were discarded because they lack a complete AP2 domain or have a very different ORF compared to the AP2/ERF sequences from other plant species. The phylogenetic trees corresponding to each subfamily in A. thaliana and E. grandis are shown in S1, S2, S3 and S4 Figs. The analysis of key residues together with the phylogenetic trees allowed classifying the AP2/ERF members from E. grandis into a soloist and the four typical subfamilies, showing a conserved family structure for the plant species under study.
After the release of the Malus genome sequence, it was reported [3, 23] that the AP2/ERF family contains 259 members grouped according to the Nakano classification [47]. However, based on our manual annotation, four additional CBF sequences were identified and 21 sequences were discarded, (mainly within AP2 subfamily) because they were found as doublets (12 sequences overlapping with other AP2/ERF at the same locus) or exhibit an incomplete AP2 domain (9 sequences). The Malus AP2 subfamily size (51 members) predicted by Girardi and co-workers [23] is significantly smaller following our annotation, since 11 sequences have been discarded and 12 other sequences are proposed as AP2-like genes (because they exhibit high homology with AP2 genes but contain only one AP2 domain). Finally, the 242 AP2/ERF members from Malus were grouped in our hands according to Sakuma classification [10], which allow the structure of the AP2/ERF family to be compared to the four other species under study (Table 1).
Table 1. Comparison of AP2/ERF family composition (number of genes in subfamilies and groups) between Arabidopsis thaliana Vitis vinifera, Populus trichocarpa, Eucalyptus grandis and Malus domestica.
| AP2/ERF family | Groups | A. thaliana | V.vinifera | P. trichocarpa | E. grandis | M. domestica |
|---|---|---|---|---|---|---|
| DREB subfamily | A1 | 6 | 7 | 6 | 17 | 5 + 1p |
| A2 | 8 | 5 | 8 | 6 | 14 | |
| A3 | 1 | 0 | 2 | 1 | 4 | |
| A4 | 16 | 15 | 26 | 16 | 18 | |
| A5 | 16 | 8 | 14 | 17 | 13 | |
| A6 | 10 | 5 | 11 | 7 | 9 | |
| Subtotal | 57 | 40 | 67 | 61 + 3p | 63 +1p | |
| ERF subfamily | B1 | 15 | 11 | 19 | 12 | 19 |
| B2 | 5 | 3 | 6 | 3 | 7 | |
| B3 | 18 | 41 | 35 | 65 | 45 | |
| B4 | 7 | 3 | 7 | 10 | 13 | |
| B5 | 8 | 4 | 8 | 5 | 12 | |
| B6 | 12 | 21 | 26 | 13 | 30 | |
| Subtotal | 65 | 83 | 101 | 103 + 5p | 126 | |
| AP2 subfamily | 18 | 20 | 26 | 21 | 29 + 11 like | |
| RAV subfamily | 6 | 6 | 5 | 6 +2p | 6 | |
| Soloist | 1 | 1 | 1 | 1 | 6 | |
| Total | 147 | 150 | 200 | 202 | 242 |
Pseudogenes are indicated as “p”.
The AP2/ERF family of E. grandis is medium-sized like in Populus, smaller than in Malus but higher than in Vitis and Arabidopsis. These plant species exhibit a similar size for the AP2 and RAV subfamilies and share a highly conserved soloist sequence, which is strongly divergent from the other AP2/ERF proteins and was only duplicated in Malus [23]. ERF, the most represented subfamily in the five species, exhibits an expansion in the tree genomes. With 108 genes, the E. grandis ERF subfamily represents 53.5% of the whole AP2/ERF family, which is in the same range that the other woody species (50.5% for Populus, 52% for Malus and 55.7% for Vitis). Within the ERF subfamily, the distribution of proteins among the B1-B6 groups is similar in Eucalyptus and Vitis but differs from the distribution in the three other species (Fig 1). Of particular note is the B3 group which with 65 members (60% of ERF subfamily) is strongly over-represented in Eucalyptus (Table 1 and Fig 1). In contrast to the ERF expansion, tree DREB subfamilies exhibit a similar size compared to Arabidopsis genome.
Fig 1. Distribution of genes within DREB and ERF subfamilies from Arabidopsis thaliana, Vitis vinifera, Populus trichocarpa Eucalyptus grandis and Malus domestica.

Pie charts depicting as sectors the relative size of each group A1-A6 and B1-B6 for respectively the DREB and ERF subfamilies. Each group is presented as a percentage of the number of corresponding DREB or ERF members, as indicated in bold in Table 1.
According to the literature, the Malus DREB subfamily contains various numbers of DREB1/CBF genes (2–3) and DREB2 genes (13–16) [29], [23]. In our hands, the analysis of conserved domains and complete protein sequences led to identify six CBF and 14 DREB2 factors. For Populus, 17 or 18 DREB2 (A2) genes have been reported [48, 49]. However, the alignment of the DREB2 protein sequences from the species under study showed that 10 of these 18 Populus DREB2 sequences cluster together and are very different from the other A2 factors (shorter sequence and different key residues), especially they do not exhibit the CMIV domain (motif 7) characteristic of the A2 group [50]. Moreover, when we blasted these 10 sequences against the NCBI databases, they were found to be closely related to B6 genes from Arabidopsis and Vitis. Therefore, we considered for further analyses only the eight remaining PtDREB2 (A2) genes for Populus. For Vitis, one VvDREB2 gene was identified in addition to the four members previously reported in the literature [27].
The resulting annotation data allow comparing the DREB subfamily (size and representation in %) for the five species under study (Table 1 and Fig 1). The subfamily size in Eucalyptus (64 genes, 31.7%) is similar to Populus (33.5%), but larger than in Malus (26%) and Vitis (26.6%). Within the subfamily, A4 is always the largest group (15 to 26 genes) but the distribution of the groups depends on the plant species (Fig 1). Interestingly, the A1 group (DREB1/CBF) is much more prominent in Eucalyptus (17 genes corresponding to 27% of the DREB subfamily) compared to the other species for which the six or seven genes correspond to 9% (Populus and Malus), 11% (Arabidopsis) and 18% (Vitis). In contrast, the Eucalyptus A2 group (six genes), similar to Vitis group (five genes, 12.5%), is smaller than in Populus and Arabidopsis (eight genes, respectively 12 and 14%) and above all in Malus (14 genes, 22% of DREB subfamily). Taken together this means that the tree species are similar with respect to the total number of DREB genes, but differ in their distribution within the groups especially in the number of genes in the A1 group, which is three times higher in Eucalyptus than in Populus and Malus. These data strongly suggest that the high number of DREB1/CBFs identified in the E. grandis genome is not a common feature in the tree species under study.
Annotation, classification and mapping of EgrCBF genes
The 16 CBF sequences predicted by the Phytozome automatic annotation for E. grandis were then further investigated on the basis of their specific features, such as the presence of “signature sequences” flanking the AP2 domain and the absence of intronic sequence. An additional EgrCBF sequence was identified, when the four EguCBF sequences, previously isolated from E. gunnii, were mapped against the E. grandis genome. According to their relative position on the classified scaffolds (1 to 11), the 17 orthologs of the E. grandis genome were annotated as EgrCBF1 to EgrCBF17. Very interestingly, 14 EgrCBF genes (1–14) are located on the scaffold 1, forming a cluster in a region of about 117 Kb (from 38661043 to 38777958) which does not contain any other gene (Fig 2). Only EgrCBF15, 16 and 17 are located on different loci, EgrCBF15 on scaffold 4 and both EgrCBF16 and 17 on scaffold 5.
Fig 2. Schematic representation of the EgrCBF gene cluster on scaffold 1.
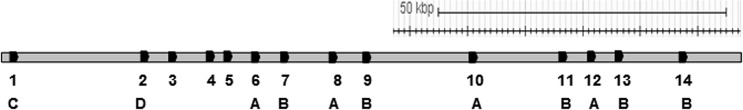
The numbers correspond to EgrCBF1-14. The letters refer to EguCBF1A-, B-, C- or D-like, based on the percentage of identity between CBFs from E. grandis and E. gunnii (except for EgrCBF3, 4 and 5 unlabelled genes) (Table 3).
For the present work, a manual curation was required in particular for three genes (EgrCBF3-4-5) that were incorrectly predicted by the Phytozome automatic annotation. For both EgrCBF4-5 genes, predicted to have an intronic sequence, the translated sequence looked like a typical CBF gene. The confirmation of the absence of intron for the two genes was provided by comparing the sequences amplified from cDNA and genomic DNA. In addition, the proposed EgrCBF4 sequence includes a start-ATG codon at the 768 position (based on position 1 of the currently annotated sequence). The originally predicted EgrCBF3 also lacked a start-ATG codon at the normal position for a typical CBF sequence. Then the comparison of newly sequenced EgrCBF3 gene and EguCBF orthologs confirmed the presence of the four nucleotides at the positions 7–10 which are missing in the Phytozome prediction. The EgrCBF3 sequence could also be isolated from cDNA, showing that EgrCBF3 can be transcribed but, because lacking start-ATG codon, it should not be translated.
The EgrCBF group can be divided into subgroups according to identity (Table 2). The most similar sequences, which share at least 90% of identical residues, correspond to three different subgroups: EgrCBF6-8-10-12, EgrCBF7-9-11-13-14, and EgrCBF16-17. Considering this very high identity level, the corresponding sequences are likely the result from duplication events which occurred more recently than those producing the other paralogs (72 to 83%). When the EgrCBF sequences were compared to the EguCBF1A-B-C-D previously isolated from E. gunnii, 11 sequences were found to exhibit a spectacularly high identity (from 91 to 99%, Table 3). While EgrCBF1 and EgrCBF2 are the closest orthologs of respectively EguCBF1C and D, the two main subgroups (EgrCBF6-8-10-12 and EgrCBF7-9-11-13-14) correspond respectively to EguCBF1A and EguCBF1B, and therefore named CBFA- and B-like.
Table 2. Pairwise sequence comparison (% of identity) between EgrCBF proteins from E. grandis.
| CBF1 | |||||||||||||||||
| CBF 1 | 100 | CBF2 | |||||||||||||||
| CBF 2 | 76 | 100 | CBF3 | ||||||||||||||
| CBF3 | 56 | 69 | 100 | CBF4 | |||||||||||||
| CBF 4 | 67 | 83 | 74 | 100 | CBF5 | ||||||||||||
| CBF 5 | 56 | 68 | 60 | 72 | 100 | CBF6 | |||||||||||
| CBF 6 | 77 | 77 | 60 | 76 | 61 | 100 | CBF7 | ||||||||||
| CBF 7 | 74 | 73 | 56 | 72 | 59 | 80 | 100 | CBF8 | |||||||||
| CBF 8 | 78 | 78 | 61 | 76 | 63 | 97 | 80 | 100 | CBF9 | ||||||||
| CBF 9 | 76 | 75 | 57 | 74 | 59 | 80 | 93 | 81 | 100 | CBF10 | |||||||
| CBF 10 | 74 | 74 | 59 | 74 | 60 | 90 | 78 | 91 | 79 | 100 | CBF11 | ||||||
| CBF 11 | 74 | 74 | 56 | 71 | 57 | 78 | 92 | 79 | 95 | 76 | 100 | CBF12 | |||||
| CBF 12 | 76 | 76 | 62 | 75 | 60 | 91 | 79 | 93 | 80 | 92 | 79 | 100 | CBF 13 | ||||
| CBF 13 | 75 | 74 | 58 | 74 | 59 | 81 | 96 | 82 | 97 | 79 | 95 | 80 | 100 | CBF 14 | |||
| CBF 14 | 76 | 75 | 58 | 74 | 59 | 81 | 95 | 82 | 97 | 79 | 96 | 80 | 98 | 100 | CBF15 | ||
| CBF 15 | 49 | 46 | 41 | 44 | 35 | 48 | 44 | 46 | 46 | 45 | 45 | 47 | 43 | 46 | 100 | CBF16 | |
| CBF 16 | 42 | 38 | 31 | 39 | 30 | 38 | 38 | 40 | 39 | 37 | 40 | 38 | 40 | 39 | 47 | 100 | CBF17 |
| CBF 17 | 42 | 39 | 32 | 39 | 29 | 40 | 38 | 41 | 40 | 38 | 41 | 40 | 40 | 39 | 47 | 92 | 100 |
Bolded numbers correspond to the most identical sequences (>70% identity).
Table 3. Pairwise sequence comparison (% of identity) between EgrCBF and EguCBF proteins respectively from E. grandis and E. gunnii.
| EgrCBF1 | EgrCBF2 | EgrCBF3 | EgrCBF4 | EgrCBF5 | EgrCBF6 | EgrCBF7 | EgrCBF8 | EgrCBF9 | EgrCBF10 | EgrCBF11 | EgrCBF12 | EgrCBF 13 | EgrCBF 14 | EgrCBF15 | EgrCBF16 | EgrCBF17 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EguCBF A | 75 | 75 | 59 | 75 | 60 | 95 | 79 | 95 | 80 | 91 | 78 | 93 | 80 | 81 | 46 | 39 | 41 |
| EguCBF B | 76 | 75 | 57 | 73 | 58 | 81 | 95 | 82 | 98 | 80 | 96 | 81 | 98 | 99 | 46 | 40 | 40 |
| EguCBF C | 95 | 73 | 55 | 66 | 54 | 76 | 72 | 76 | 74 | 73 | 72 | 74 | 74 | 74 | 46 | 40 | 40 |
| EguCBF D | 75 | 97 | 68 | 81 | 67 | 75 | 72 | 76 | 73 | 73 | 72 | 74 | 73 | 74 | 47 | 37 | 40 |
Bolded numbers correspond to the most identical sequences (>90% identity).
Phylogenetic relationships within CBF and DREB2 groups from Eucalyptus, Arabidopsis, Vitis, Malus and Populus
The multiple alignments of CBF protein sequences from Arabidopsis, Vitis, Populus, Malus and Eucalyptus (E. grandis and E. gunnii) were used to generate the phylogenetic tree presented in Fig 3. The CBF paralogs are distributed among either two (Arabidopsis, Eucalyptus and Malus) or three clades (Vitis and Populus). These clades are split into plant specific sub-clades, composed of highly-related paralogs, suggesting that gene duplications seem to have occurred in a plant species specific way. The two clades of the well-characterized Arabidopsis CBF proteins correspond to typical and atypical CBFs (respectively AtCBF1-4 and AtDDF1-2). The small clade (EgrCBF15-17) appears to be phylogenetically related to the AtDDFs. Interestingly, the main clade (EgrCBF1-14) corresponds to the clustered genes on scaffold 1, providing a strong evidence of repeated and recent gene duplications. This large clade might result from successive waves of duplications from one ancestor gene, the most recent leading to the two main sub-clades (CBF-A and-B like genes).
Fig 3. Phylogenetic analyses of CBF proteins (A1 group) from Eucalyptus grandis, Eucalyptus gunnii, Vitis vinifera, Malus domestica, Populus trichocarpa and Arabidopis thaliana by the Maximum Likelihood method using Mega 5.
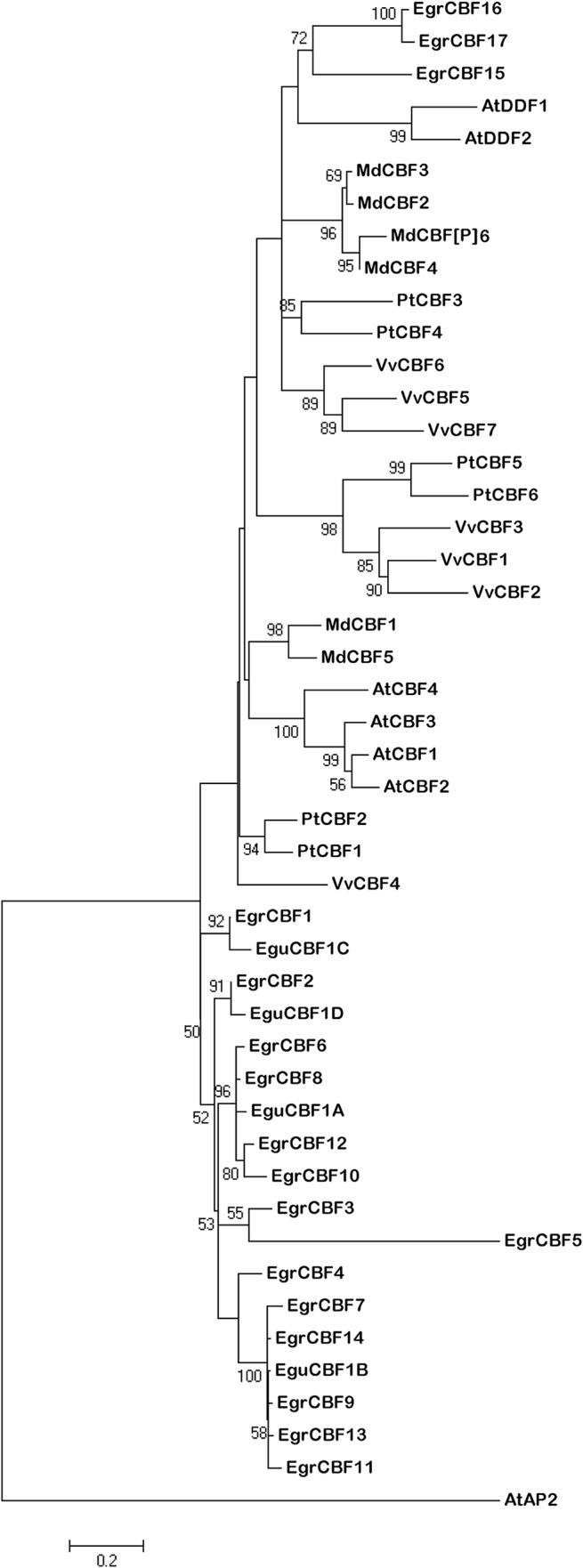
The tree was constructed from the sequence alignement of full-length proteins of the EgrCBF1-17 from E. grandis, Egu CBF1A-B-C-D from E. gunnii, VvCBF1-7 from V. vinifera, PtCBF1-6 from P. trichocarpa, AtCBF1-4 and AtDDF1-2 from A. thaliana and MdCBF1-6 from Malus domestica. AtAP2 (At5G557390.1) is a member of the AP2 subfamily used for rooting the phylogenetic tree. Low Bootstrap values (<50) were removed from the tree.
The same approach with DREB2 protein sequences from Arabidopsis, Vitis, Populus, Malus and Eucalyptus was used for generating the DREB2 phylogenetic tree. As presented on Fig 4, the tree is divided into three main clades, each composed of paralogs from the five species under study, except for one clade with no Malus genes. Similarly to Vitis but in contrast to Populus, Malus and Arabidopsis, the six sequences from Eucalyptus are spread out over the phylogenetic tree, indicating the absence of recent duplication in this DREB group.
Fig 4. Phylogenetic analyses of DREB2 proteins (A2 group) from Eucalyptus grandis, Vitis vinifera, Malus domestica, Populus trichocarpa and Arabidopsis thaliana by the Maximum Likelihood method using Mega 5.
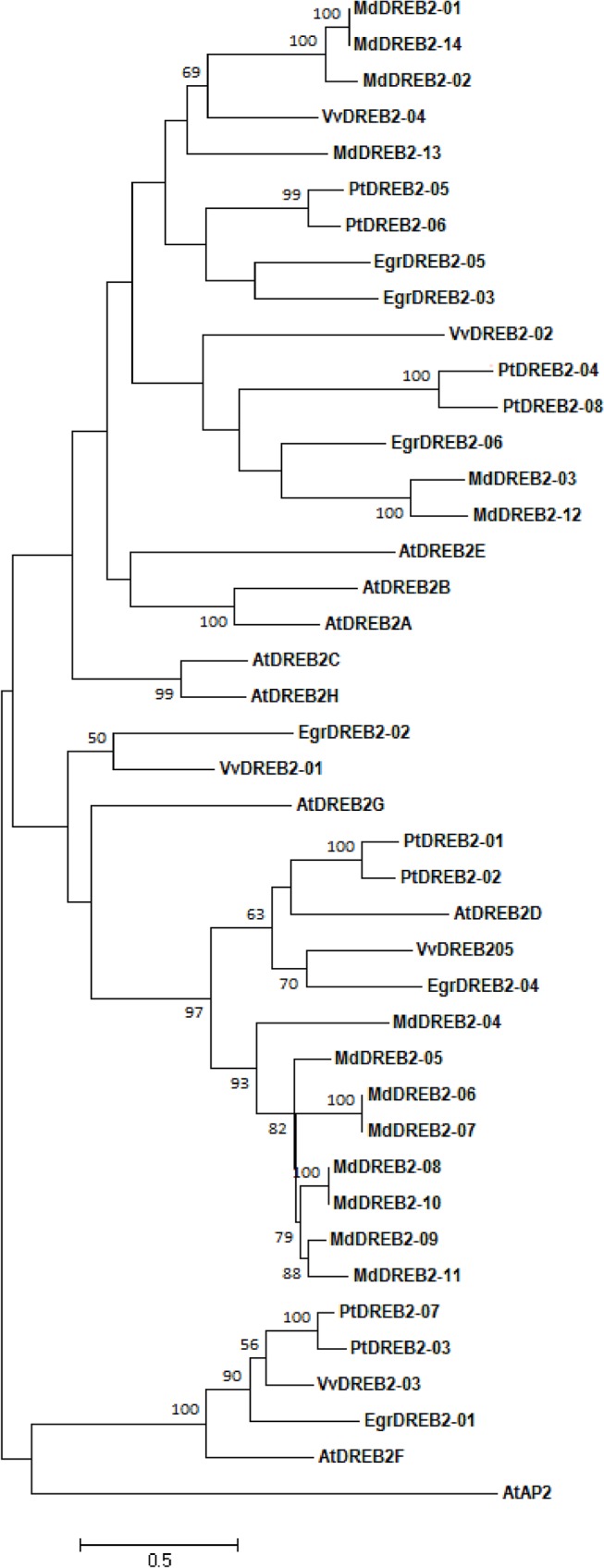
The tree was constructed from the sequence alignment of full-length protein of AtDREB2-(A-H) from A. thaliana, EgrDREB2-(1–6) from E. grandis, VvDREB2-(1–5) from V. vinifera, PtDREB2-(1–8) from P. trichocarpa, MdDREB2-(1–14) from Malus domestica. AtAP2 (At5G557390) is a member of AP2 subfamily used for rooting the phylogenetic tree. Low Bootstrap values (<50) were removed from the tree.
The phylogenetic relationships indicate that the EgrCBF genes result from recent, specific and repeated duplications while the EgrDREB2 members have evolved without specific duplication from sequences prevalent already before the divergence of the species.
Evaluation of the EgrCBF and EgrDREB2 transcription in standard or stressful conditions
The CBF and DREB2 gene expression profiles were first investigated by using RNAseq data from different tissues of E. grandis plants grown in standard conditions. These data only concern five DREB2 in addition to 14 CBF genes automatically annotated. As shown on Fig 5, three groups containing both EgrCBF and EgrDREB2 genes can be distinguished according to their preferential expression in woody tissues (Group I), shoot tips (Group II) or leaf tissues (Group III). Each group includes both CBF and DREB2 genes. Surprisingly, 6 of the 19 CBF/DREB2 genes are preferentially expressed in phloem tissue in addition to DREB2-5 which exhibits the highest expression value (Log10 FKPM) in xylem tissue. In the “leaf group”, the highest expression levels mainly concern mature leaves.
Fig 5. CBF and DREB2 gene expression profiles from RNAseq data corresponding to various tissues from E. grandis.

Raw expression data were obtained from the Eucalyptus genome integrative explorer (http://eucgenie.bi.up.ac.za). Expression analysis is presented for 14 EgrCBF and five EgrDREB2 genes which were automatically annotated, but do not concerns the four additional CBF genes which were manually annotated in our hands. Gene name and accession numbers are mentioned respectively in Alias and ID columns. Heat map and hierarchical clustering of gene expression profiles (right panel) were performed by using expander software (ACGT, http://www.cs.tau.ac.il). For each gene, the highest absolute expression value (Log10 FKPM) is represented in the left panel with the indication of corresponding tissue. YL, young leaves; ML, mature leaves; F, flowers; ST, shoot tips; R, roots P, phloem; X, xylem.
The CBF or DREB2 transcript abundance was quantified on leaves from E. gunnii x E. dalrympleana cold-treated plants in comparison with standard conditions (Fig 6).This expression analysis shows that most of the CBF and DREB2 genes are expressed in standard conditions. The statistical analysis indicates a significant cold induction for most of the CBF genes (CBF1-14) but only two DREB2 genes (DREB2-3 and-5). From the 23 genes under study, CBF14 exhibits the strongest expression under cold and CBF9 shows the highest basal expression. Interestingly, these two genes are close orthologs of respectively EguCBF1B and EguCBF1A, which were chosen for over-expression in E. urophylla x E. grandis transgenic lines [24].
Fig 6. EgrCBF and EgrDREB2 transcript abundance in leaves of E. gunnii x E. dalrympleana plants grown for 8h under 4°C cold (white) or control conditions (black).

Gene expression levels of EgrCBF (C1 to C17) and EgrDREB2 (D2-1 to -6) were quantified on leaves from cold-treated Eucalyptus plants. The experimental values from RT-qPCR (conducted on three biological replicates) were normalized using five reference genes and the mean values (n = 3) were analyzed for significance using the statistical T-test (p value < 0.05). The resulting histograms represent Log10 of relative transcript abundance of the CBF and DREB2 in control or cold conditions. * means a significant 2-fold rate increase in gene expression (cold/control)
In addition, the responses of these genes to different stressful conditions (cold, heat or drought) were evaluated on detached leaves from E. grandis, since the use of this simplified system was validated previously [43]. In this paper, the induction of different CBF genes of cold-stressed pieces of leaves was shown to be representative of CBF expression profiles from Eucalyptus cold-treated plants.
In the present paper, the gene induction rates (stress condition compared to control) are expressed (Fig 7) as the Log10 of the mean values (a 2-fold rate increase in gene expression corresponds to Log10 values > 0.33). In this experiment, all the CBF genes were found to be cold-induced versus 4/6 DREB2 genes. In agreement with the presented data on whole plants, the overall CBF group exhibits a more intense cold response than the DREB2 group. At the opposite, under drought, a higher proportion of DREB2 genes (4/6) are up-regulated compared to the CBF genes (4/17), even though the applied stimulus is overall triggering a much weaker response compared to cold treatment. Finally, all the CBF and DREB2 genes were found to be heat-inducible.
Fig 7. EgrCBF and EgrDREB2 gene induction rates for E. grandis detached leaves incubated for 8 hours under cold (4°C, blue), heat (45°C, red) or drought (yellow), in comparison to control conditions.
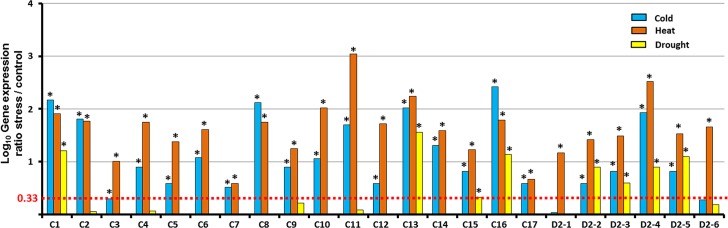
Transcript abundance of EgrCBF (C1 to C17) and EgrDREB2 (D2-1 to -6) was quantified through RT-qPCR on detached leaves from 10 individual E. grandis plants. The experimental values were normalized using two reference genes and the mean values issued from three independent biological replicates (n = 3) were analyzed for significance using the statistical T-test (p value < 0.01). The histograms represent the relative expression values as Log10 of induction rates (stress/control). * means a significant 2-fold rate increase in gene expression (stress/control)
Overall, these analyses confirm that both CBF and DREB2 groups participate in cold, drought and above all heat response in E grandis and E. gunnii x E. dalrympleana. However, CBF group would be mostly involved in cold response and DREB2 mainly in drought response. Moreover, the data evidence that, for these two species of Eucalyptus, regulation is not redundant within the CBF/DREB2 groups since each gene exhibits a distinct pattern of response in different tissues and stressful conditions.
Discussion
A comprehensive search for the Eucalyptus AP2/ERF transcription factors led to the identification of 202 genes which is similar to Populus and ranges in between Vitis (147 genes) and Zea mays (292 members) [51]. A previous comparison of the AP2/ERF gene family size between eudicots, monocots and chlorophyta, have already shown that this size radiates along with land conquest [52]. Actually, most of the observed expansions in AP2/ERF families in plants seem to be associated to recent genome duplications (Eucalyptus, Populus and Malus) or triplications (Brassica rapa) [53], [54], [55], [23]. The structure and phylogeny of the AP2/ERF family are similar for the species under study. The presence of the same subfamilies composed of the same groups suggests that the appearance of the AP2/ERF structure predates the divergence of dicotyledonous species [28, 47]. Compared to Arabidopsis, the tree species under study are characterized by a much larger ERF subfamily beside a similar size for DREB subfamily. Current knowledge states that DREB factors activate genes involved in cold and water deficit response, while ERF factors directly regulate pathogenesis-related (PR) gene expression but some ERF and DREB genes were shown to be activated by both biotic and abiotic stimuli [56]. Actually, most of these ERF in Eucalyptus are B3 factors. More than other ones, these proteins are often associated to the response to both biotic and abiotic stresses, especially through the binding to CE1 element [57],[58], [59].
With regard to the Eucalyptus DREB subfamily, the obvious feature lies in the presence of numerous EgrCBF genes together with a moderate number of EgrDREB2 genes. The phylogenetic analysis suggests that the DREB2 forms existed prior to the plant species divergence, whereas the CBF may have evolved more recently and independently in each plant species. Up to now, the number of CBF genes in dicotyledonous plants is often around six, except for Chrysanthemum [60] and Medicago truncatula [61]. In the first case, the 13 CBF isolated genes are likely related to the hexaploid Chrysanthemum genome; 17 CBF genes have been detected in M. truncatula genome which has experienced high rates of local gene duplications.
The occurrence of clustered CBF genes is a quite common feature for several plant species such as Arabidopsis, wheat and rice [9, 25, 62], but the number of CBF duplication events is much higher in Medicago and Eucalyptus. Duplicated genes can be conserved and associated with sub-functionalization and/or neo-functionalization [63] or become pseudogenes. The intriguing high number of EgrCBF genes may give rise to doubts as to their transcription. Given that all the EgrCBFs are expressed and that only one is predicted not to be translated, it then raises questions about the role of these very similar multiple copies in Eucalyptus. Are they required to ensure an essential function or is there redundancy in their function? Knox and coworkers showed that the presence of two extra-CBF gene copies in a barley genotype contributes to its higher freezing tolerance [25], suggesting that amplification of genomic regions encompassing CBFs would be a mean to increase CBF transcription rate and provide a selective advantage during winter. Also the CBF family expansion in the Pooideae during the Eocene-Oligocene climate cooling was strongly suggested to play a role in the adaptation to a cool climate [64]. Considering its tropical and subtropical origin and ever-growing habit, E. grandis is reported to exhibit a surprisingly high level of frost tolerance as a result of a very rapid hardening capacity [65]. Eucalyptus genus is also known as heat-tolerant [66]. The CBF could, at least in part, participate in the control of the Eucalyptus cold acclimation since over-expression of an E. gunnii CBF ortholog in Eucalyptus was found to increase cold tolerance [24]. Interestingly, the protein corresponding to this transgene is very similar (98–99%) to EgrCBF13 and 14 which were observed as the most expressed in the present paper, such validating their functionality. Therefore, duplication and retention of the CBF genes, most cold- and heat-responsive may contribute to a massive activation of target genes (such as dehydrin) as previously observed during Eucalyptus cold acclimation [67].
On the other hand, the different copies may be not equivalent for regulation or function, at least at the subgroup level. In Arabidopsis, the divergence of the CBF4 gene from the CBF1-3 tandem proved not to be due to selection of entirely new functions, but to diversification of promoter regulatory elements responding to different environmental signals [68, 69]. In E. gunnii, previous studies have shown a complementary regulation of the four CBF genes under various cold conditions [33]. The present study clearly shows that the expression of the 17 EgrCBF genes is differential according to the plant tissue and the applied stress. Not only these genes are not specific to one stress but also they are not equivalent for regulation. Therefore the high number of CBF copies may provide both efficiency and plasticity to the Eucalyptus response to environmental stresses.
The unusual CBF over-representation, in contrast to the moderate number of DREB2 genes in Eucalyptus also raises the question about the adaptation of this species to cold, heat or drought. It was originally thought that CBF genes are the main regulators of cold response when DREB2 group is rather involved in osmotic response, but accumulation of genomic data on different species have shown some conflicts with respect to this classical model [7]. The present study on Eucalyptus species shows that both groups are regulated by both stimuli, but CBF are more cold-sensitive than DREB2 genes which look more involved in drought response. The regulation of CBF genes by various dehydrative stresses was previously shown in E. gunnii [33], Vitis [70], Malus [21], Prunus [22] and Populus [50]. In Arabidopsis, the CBF group includes the strictly cold-induced genes (CBF1-3) together with the drought- and salt-responsive genes (CBF4, DDF1 and DDF2) [71]. Conversely, cold-responsive DREB2 genes have been reported for some grass species [56, 72] Arabidopsis [58], Malus [6] or Populus [50]. Therefore, the numerous CBF and DREB2 genes may together be involved in the control of temperature stress in Eucalyptus. One obvious characteristic of CBF/DREB2 genes with regard to transcriptional regulation in Eucalyptus lies in the strong response to heat which concerns all the 23 members. This feature does not look so common since for example the recent publication [73] on heat transcriptome in poplar states only 19 up-regulated DREB genes versus 23 down-regulated for the six groups (A1-A6 containing 60 genes).
From our results, one may therefore hypothesize that, in Eucalyptus, the large CBF group would complete the limited number of DREB2 genes, for regulating multiple abiotic stress responses. This does not seem so surprising because expansion of Eucalyptus into Australia occurred during the late Miocene when the climate became colder and drier [74]. This means that this tree was submitted to a double selection pressure. As regards the cold response, the higher number of CBF genes in Eucalyptus compared to other trees highlights the very distinct functional strategies to face winter mainly based on frost tolerance for Eucalyptus versus frost avoidance for Populus or Malus. In addition, all the CBF and DREB2 genes exhibit a significant induction by heat stress. It is now commonly accepted that Eucalyptus became the dominant sclerophyll forest in Australia, about 140 000 years ago, when Aboriginal people arrived because of “fire dick” agriculture [75]. It is proposed that Eucalyptus plants were merely pre-adapted for fire survival and expansion [66]. The presence of a large number of heat-responsive CBFs might be a piece of this pre-adaptation by controlling proteins involved in heat protection, like HSP. In conclusion, CBF and DREB2 may be key players of cross-talks between cold, heat and drought responses which appear to be strong in Eucalyptus both for the regulatory pathways and the resulting adaptive mechanisms. The unusual expansion of the EgrCBF group, together with a differential gene regulation could be a way of counterbalancing the permanent exposure to a changing environment and may contribute to a wider response to a range of environmental changes including at least heat shock.
In conclusion, this investigation of the AP2/ERF family in Eucalyptus with regard to other woody species reveals some common features, such as family size and global structure. As more and more reported for plants, the stress-related groups CBF and DREB2 prove differentially but not specifically regulated by various abiotic stresses, CBF members participating mainly in the cold response and DREB2 genes in the drought response. More surprising with regard to other species is the strong response to heat stress for all the CBF and DREB2 members.
Above all, Eucalyptus genome is marked out by a dramatic over-representation of CBF genes, which does not appear like a woody plant feature. This result gives evidence of intense and recent duplication events and suggests their importance in the adaptation of this ever-growing tree to cold, drought and above all heat. Overall, this result strongly supports the hypothesis of Myburg and coworkers [3] that the intense genome-wide duplications in Eucalyptus (the highest reported to date) may be important for adaptive evolution of this tree in dynamically changing environments.
Finally, these data make now possible to envisage a functional characterization including spatiotemporal expression studies for identifying the most relevant candidate genes (CBF and DREB2) towards improving stress tolerance (cold, drought or combined abiotic stresses).
Supporting Information
The tree was generated using Mega 5 program by the Maximum Likelihood method. Bootstrap values are indicated at each branch. For the E. grandis DREB subfamily, the relation-ship between Phytozome gene IDs and the names used in this paper is indicated in S2 Table.
(TIF)
The tree was generated using Mega 5 program by the Maximum Likelihood method. Bootstrap values are indicated at each branch. For the E. grandis ERF subfamily, the relation-ship between Phytozome gene IDs and the names used in this paper is indicated in S2 Table.
(TIF)
The tree was generated using Mega 5 program by the Maximum Likelihood method. Bootstrap values are indicated at each branch. For the E. grandis RAV subfamily, the relation-ship between Phytozome gene IDs and the names used in this paper is indicated in S2 Table.
(TIF)
The tree was generated using Mega 5 program by Maximum Likelihood method. Bootstrap values are indicated at each branch. For the E. grandis AP2 subfamily, subfamily, the relation-ship between Phytozome gene IDs and the names used in this paper is indicated in S2 Table.
(TIF)
(DOC)
“Eucagen name” associates the subfamily name with the relative position on the scaffolds (1 to 11). “Generic name” given to EgrCBF(1–17) and EgrDREB2-(1–6) from literature uses and genome location.
(DOC)
Acknowledgments
We acknowledge A. Myburg for providing us with RNAseq data on different tissues from E. grandis. We are also very grateful to Dr A. Nassuth from Guelph University (Canada) for critical reviewing the manuscript and English editing.
Data Availability
All relevant data are within the paper and its supporting information files, accession numbers and genome location on genomic sequence (Phytozome database).
Funding Statement
This work was funded by Fibre Excellence (31, St Gaudens, France) associated with the Midi Pyrénées Regional Council (France) and also supported by the French Ministry for Scientific Research (University Toulouse III (UPS), CNRS and Labex Tulip (ANR-10-LABX-41); the Vietnamese Government provided the PBC grant. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. FAO. State of the World's Forests 2007 Rome: Electronic Publishing Policy and Support Branch, Communication Division, FAO; 2007. [Google Scholar]
- 2. Hinchee M, Zhang C, Chang S, Cunningham M, Hammond W, Nehra N. Biotech Eucalyptus can sustainably address society's need for wood: the example of freeze tolerant Eucalyptus in the southeastern U.S. BMC Proceedings. 2011;5(Suppl 7):I24 PubMed 10.1186/1753-6561-5-S7-I24 [DOI] [Google Scholar]
- 3. Myburg AA, Grattapaglia D, Tuskan GA, Hellsten U, Hayes RD, Grimwood J, et al. The genome of Eucalyptus grandis. Nature. 2014;510(7505):356–62. Epub 2014/06/12. 10.1038/nature13308 PubMed . [DOI] [PubMed] [Google Scholar]
- 4. Okamuro JK, Caster B, Villarroel R, Van Montagu M, Jofuku KD. The AP2 domain of APETALA2 defines a large new family of DNA binding proteins in Arabidopsis. Proc Nat Ac Sci USA. 1997;94(13):7076–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Feng JX, Liu D, Pan Y, Gong W, Ma LG, Luo JC, et al. An annotation update via cDNA sequence analysis and comprehensive profiling of developmental, hormonal or environmental responsiveness of the Arabidopsis AP2/EREBP transcription factor gene family. Plant Mol Biol. 2005;59(6):853–68. 10.1007/s11103-005-1511-0 PubMed . [DOI] [PubMed] [Google Scholar]
- 6. Matsukura S, Mizoi J, Yoshida T, Todaka D, Ito Y, Maruyama K, et al. Comprehensive analysis of rice DREB2-type genes that encode transcription factors involved in the expression of abiotic stress-responsive genes. Mol Genet Genomics. 2010;283(2):185–96. 10.1007/s00438-009-0506-y PubMed . [DOI] [PubMed] [Google Scholar]
- 7. Lata C, Prasad M. Role of DREBs in regulation of abiotic stress responses in plants. J Exp Bot. 2011;62(14):4731–48. err210 [pii] 10.1093/jxb/err210 PubMed . [DOI] [PubMed] [Google Scholar]
- 8. Zhou M-L, Yi-Xiong T, Yan-min W. Genome-wide analysis of APE/ERF transcription factor family in Zea maize Current Bioinformatics. 2013;7(3). doi: 10.2174/157489312802 460776 [Google Scholar]
- 9. Sharoni AM, Nuruzzaman M, Satoh K, Shimizu T, Kondoh H, Sasaya T, et al. Gene structures, classification and expression models of the AP2/EREBP transcription factor family in rice. Plant Cell Physiol. 2011;52(2):344–60. pcq196 [pii] 10.1093/pcp/pcq196 PubMed . [DOI] [PubMed] [Google Scholar]
- 10. Sakuma Y, Liu Q, Dubouzet J, Abe H, Shinozaki K, Yamaguchi-Shinozaki K. DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration- and cold-inducible gene expression. Biochem Biophys Res Commun. 2002;290(3):998–1009. [DOI] [PubMed] [Google Scholar]
- 11. Elliott RC, Betzner AS, Huttner E, Oakes MP, Tucker WQ, Gerentes D, et al. AINTEGUMENTA, an APETALA2-like gene of Arabidopsis with pleiotropic roles in ovule development and floral organ growth. Plant Cell. 1996;8(2):155–68. PubMed [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Chuck G, Meeley RB, Hake S. The control of maize spikelet meristem fate by the APETALA2-like gene indeterminate spikelet1. Genes Dev. 1998;12(8):1145–54. PubMed [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Boutilier K, Offringa R, Sharma VK, Kieft H, Ouellet T, Zhang L, et al. Ectopic expression of BABY BOOM triggers a conversion from vegetative to embryonic growth. Plant Cell. 2002;14(8):1737–49. PubMed [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Alonso JM, Stepanova AN, Solano R, Wisman E, Ferrari S, Ausubel FM, et al. Five components of the ethylene-response pathway identified in a screen for weak ethylene-insensitive mutants in Arabidopsis. Proc Natl Acad Sci U S A. 2003;100(5):2992–7. 0438070100 [pii] 10.1073/pnas.0438070100 PubMed [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Hu YX, Wang YX, Liu XF, Li JY. Arabidopsis RAV1 is down-regulated by brassinosteroid and may act as a negative regulator during plant development. Cell Res. 2004;14(1):8–15. 10.1038/sj.cr.7290197 PubMed . [DOI] [PubMed] [Google Scholar]
- 16. Sohn KH, Lee SC, Jung HW, Hong JK, Hwang BK. Expression and functional roles of the pepper pathogen-induced transcription factor RAV1 in bacterial disease resistance, and drought and salt stress tolerance. Plant Mol Biol. 2006;61(6):897–915. PubMed . [DOI] [PubMed] [Google Scholar]
- 17. Agarwal P, Jha B. Transcription factors in plants and ABA dependent and independent abiotic stress signalling. Biol Plant. 54: Springer; 2010. p. 201–12. [Google Scholar]
- 18.Mizoi J, Shinozaki K, Yamaguchi-Shinozaki K. AP2/ERF family transcription factors in plant abiotic stress responses. Biochim Biophys Acta. 2011. S1874-9399(11)00142-8 [pii] 10.1016/j.bbagrm.2011.08.004 PubMed . [DOI] [PubMed]
- 19. Fujimoto SY, Ohta M, Usui A, Shinshi H, Ohme-Takagi M. Arabidopsis ethylene-responsive element binding factors act as transcriptional activators or repressors of GCC box-mediated gene expression. Plant Cell. 2000;12(3):393–404. PubMed [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Ohme-Takagi M, Shinshi H. Ethylene-inducible DNA binding proteins that interact with an ethylene-responsive element. Plant Cell. 1995;7(2):173–82. 7/2/173 [pii] 10.1105/tpc.7.2.173 PubMed [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Zhao T, Liang D, Wang P, Liu J, Ma F. Genome-wide analysis and expression profiling of the DREB transcription factor gene family in Malus under abiotic stress. Mol Genet Genomics. 2012;287(5):423–36. 10.1007/s00438-012-0687-7 PubMed . [DOI] [PubMed] [Google Scholar]
- 22. Artlip TS, Wisniewski ME, Bassett CL, Norelli JL. CBF gene expression in peach leaf and bark tissues is gated by a circadian clock. Tree Physiol. 2013;33(8):866–77. Epub 2013/08/21. 10.1093/treephys/tpt056 PubMed . [DOI] [PubMed] [Google Scholar]
- 23. Girardi CL, Rombaldi CV, Dal Cero J, Nobile PM, Laurens F, Bouzayen M, et al. Genome-wide analysis of the AP2/ERF superfamily in apple and transcriptional evidence of ERF involvement in scab pathogenesis. Scientia Horticulturae. 2013;151(0):112–21. 10.1016/j.scienta.2012.12.017. [DOI] [Google Scholar]
- 24. Navarro M, Ayax C, Martinez Y, Laur J, El Kayal W, Marque C, et al. Two EguCBF1 genes overexpressed in Eucalyptus display a different impact on stress tolerance and plant development. Plant Biotechnol J. 2011;9(1):50–63. Epub 2010/05/25. 10.1111/j.1467-7652.2010.00530.x PubMed . [DOI] [PubMed] [Google Scholar]
- 25. Knox AK, Dhillon T, Cheng H, Tondelli A, Pecchioni N, Stockinger EJ. CBF gene copy number variation at Frost Resistance-2 is associated with levels of freezing tolerance in temperate-climate cereals. Theor Appl Genet. 2010;121(1):21–35. 10.1007/s00122-010-1288-7 PubMed . [DOI] [PubMed] [Google Scholar]
- 26. Guo A, He K, Liu D, Bai S, Gu X, Wei L, et al. DATF: a database of Arabidopsis transcription factors. Bioinformatics. 2005;21(10):2568–9. bti334 [pii] 10.1093/bioinformatics/bti334 PubMed . [DOI] [PubMed] [Google Scholar]
- 27. Zhuang J, Peng RH, Cheng ZM, Zhang J, Cai B, Zhang Z, et al. Genome-wide analysis of the putative AP2/ERF family genes in Vitis vinifera. Scientia Horticulturae. 2009;123(1):73–81. 10.1016/j.scienta.2009.08.002 PubMed WOS:000271159000013. [DOI] [Google Scholar]
- 28. Licausi F, Giorgi FM, Zenoni S, Osti F, Pezzotti M, Perata P. Genomic and transcriptomic analysis of the AP2/ERF superfamily in Vitis vinifera. BMC Genomics. 2010;11:719 1471-2164-11-719 [pii] 10.1186/1471-2164-11-719 PubMed [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Zhuang J, Chen JM, Yao QH, Xiong F, Sun CC, Zhou XR, et al. Discovery and expression profile analysis of AP2/ERF family genes from Triticum aestivum. Mol Biol Rep. 2011;38(2):745–53. 10.1007/s11033-010-0162-7 PubMed . [DOI] [PubMed] [Google Scholar]
- 30.Hefer C, Mizrachi E, Joubert F, Myburg A, editors. The Eucalyptus genome integrative explorer (EucGenIE): a resource for Eucalyptus genomics and transcriptomics. IUFRO Tree Biotechnology Conference; 2011 2011; Arraial d’Ajuda, Bahia, Brazil: BMC Proceedings.
- 31. Odronitz F, Pillmann H, Keller O, Waack S, Kollmar M. WebScipio: an online tool for the determination of gene structures using protein sequences. BMC Genomics. 2008;9:422 1471-2164-9-422 [pii] 10.1186/1471-2164-9-422 PubMed [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Jaglo KR, Kleff S, Amundsen KL, Zhang X, Haake V, Zhang JZ, et al. Components of the Arabidopsis C-repeat/dehydration-responsive element binding factor cold-response pathway are conserved in Brassica napus and other plant species. Plant Physiology. 2001;127(3):910–7. [PMC free article] [PubMed] [Google Scholar]
- 33. Navarro M, Marque G, Ayax C, Keller G, Borges JP, Marque C, et al. Complementary regulation of four Eucalyptus CBF genes under various cold conditions. J Exp Bot. 2009;60(9):2713–24. erp129 [pii] 10.1093/jxb/erp129 PubMed [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Kapustin Y, Souvorov A, Tatusova T, Lipman D. Splign: algorithms for computing spliced alignments with identification of paralogs. Biol Direct. 2008;3:20 1745-6150-3-20 [pii] 10.1186/1745-6150-3-20 PubMed [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Grattapaglia D, Vaillancourt RE, Shepherd M, Thumma BR, Foley W, Külheim C, et al. Progress in Myrtaceae genetics and genomics: Eucalyptus as the pivotal genus. Tree Genetics & Genomes. 2012;8:463–508. [Google Scholar]
- 36. Galiba G, Vagujfalvi A, Li CX, Soltesz A, Dubcovsky J. Regulatory genes involved in the determination of frost tolerance in temperate cereals. Plant Sci. 2009;176(1):12–9. 10.1016/j.plantsci.2008.09.016 PubMed WOS:000261551900002. [DOI] [Google Scholar]
- 37. Medina J, Catalá R, Salinas J. The CBFs: three arabidopsis transcription factors to cold acclimate. Plant Sci. 2011;180(1):3–11. S0168-9452(10)00187-1 [pii] 10.1016/j.plantsci.2010.06.019 PubMed . [DOI] [PubMed] [Google Scholar]
- 38. Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, et al. Clustal W and Clustal X version 2.0. Bioinformatics. 2007;23(21):2947–8. btm404 [pii] 10.1093/bioinformatics/btm404 PubMed . [DOI] [PubMed] [Google Scholar]
- 39. Hall T. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl Acids Symp Ser. 1999;41:95–8. [Google Scholar]
- 40. Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol. 2011;28(10):2731–9. msr121 [pii] 10.1093/molbev/msr121 PubMed [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Felsenstein J. Evolutionary trees from DNA sequences: a maximum likelihood approach. J Mol Evol. 1981;17(6):368–76. PubMed . [DOI] [PubMed] [Google Scholar]
- 42. El Kayal W, Navarro M, Marque G, Keller G, Marque C, Teulieres C. Expression profile of CBF-like transcriptional factor genes from Eucalyptus in response to cold. J Exp Bot. 2006;57(10):2455–69. 10.1093/jxb/erl019 PubMed WOS:000239389700030. [DOI] [PubMed] [Google Scholar]
- 43. Navarro M, Marque G, Ayax C, Keller G, Borges JP, Marque C, et al. Complementary regulation of four Eucalyptus CBF genes under various cold conditions. J Exp Bot. 2009;60(9). 10.1093/jxb/erp129 PubMed WOS:000266957900025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Jones J, Dunsmuir P, Bedbrook J. High level expression of introduced chimeric genes in regenerated transformed plants. The EMBO Journal. 1985;4:2411–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCT method. Methods. 2001;25(4):402–8. [DOI] [PubMed] [Google Scholar]
- 46. Myburg AA, Griffin AR, Sederoff RR, Whetten RW. Comparative genetic linkage maps of Eucalyptus grandis, Eucalyptus globulus and their F1 hybrid based on a double pseudo-backcross mapping approach. Theoretical and Applied Genetics. 2003;107(6):1028–42. PubMed . [DOI] [PubMed] [Google Scholar]
- 47. Nakano T, Suzuki K, Fujimura T, Shinshi H. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol. 2006;140(2):411–32. pp.105.073783 [pii] 10.1104/pp.105.073783 PubMed [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Zhuang J, Cai B, Peng RH, Zhu B, Jin XF, Xue Y, et al. Genome-wide analysis of the AP2/ERF gene family in Populus trichocarpa. Biochem Biophys Res Commun. 2008;371(3):468–74. S0006-291X(08)00781-X [pii] 10.1016/j.bbrc.2008.04.087 PubMed . [DOI] [PubMed] [Google Scholar]
- 49. Xu W, Li F, Ling L, Liu A. Genome-wide survey and expression profiles of the AP2/ERF family in castor bean (Ricinus communis L.). BMC Genomics. 2013;14(1):785 Epub 2013/11/15. 10.1186/1471-2164-14-785 PubMed . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Chen Y, Yang J, Wang Z, Zhang H, Mao X, Li C. Gene Structures, Classification, and Expression Models of the DREB Transcription Factor Subfamily in Populus trichocarpa. ScientificWorldJournal. 2013;2013:954640 Epub 2013/12/11. 10.1155/2013/954640 PubMed . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Zhou M-L, Tang Y-X, Wu Y-m. Genome-Wide Analysis of AP2/ERF Transcription Factor Family in Zea Mays. Current Bioinformatics. 2012;7(3):324–32. [Google Scholar]
- 52. Dietz KJ, Vogel MO, Viehhauser A. AP2/EREBP transcription factors are part of gene regulatory networks and integrate metabolic, hormonal and environmental signals in stress acclimation and retrograde signalling. Protoplasma. 2010;245(1–4):3–14. 10.1007/s00709-010-0142-8. PubMed 10.1007/s00709-010-0196-7 [DOI] [PubMed] [Google Scholar]
- 53. Wang H, Moore MJ, Soltis PS, Bell CD, Brockington SF, Alexandre R, et al. Rosid radiation and the rapid rise of angiosperm-dominated forests. Proc Natl Acad Sci U S A. 2009;106(10):3853–8. Epub 2009/02/19. 10.1073/pnas.0813376106 PubMed [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Myburg A, Grattapaglia D, Tuskan G, Jenkins J, Schmutz J, Mizrachi E, et al. The Eucalyptus grandis Genome Project: Genome and transcriptome resources for comparative analysis of woody plant biology. BMC Proceedings. 2011;5(Suppl 7):I20 PubMed 10.1186/1753-6561-5-S7-I20 [DOI] [Google Scholar]
- 55. Song J, Liu J, Weng M, Huang Y, Luo L, Cao P, et al. Cloning of galactinol synthase gene from Ammopiptanthus mongolicus and its expression in transgenic Photinia serrulata plants. Gene. 2013;513(1):118–27. 10.1016/j.gene.2012.10.058 PubMed . [DOI] [PubMed] [Google Scholar]
- 56. Xu ZS, Chen M, Li LC, Ma YZ. Functions of the ERF transcription factor family in plants. Botany-Botanique. 2008;86(9):969–77. 10.1139/b08-041 PubMed WOS:000259575700001. [DOI] [Google Scholar]
- 57. Lee SJ, Cho DI, Kang JY, Kim SY. An ARIA-interacting AP2 domain protein is a novel component of ABA signaling. Mol Cells. 2009;27(4):409–16. 10.1007/s10059-009-0058-3 PubMed . [DOI] [PubMed] [Google Scholar]
- 58. Lee SJ, Kang JY, Park HJ, Kim MD, Bae MS, Choi HI, et al. DREB2C interacts with ABF2, a bZIP protein regulating abscisic acid-responsive gene expression, and its overexpression affects abscisic acid sensitivity. Plant Physiol. 2010;153(2):716–27. 10.1104/pp.110.154617 PubMed [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Zhang W, Zou A, Miao J, Yin Y, Tian R, Pang Y, et al. LeERF-1, a novel AP2/ERF family gene within the B3 subcluster, is down-regulated by light signals in Lithospermum erythrorhizon. Plant Biol (Stuttg). 2011;13(2):343–8. Epub 2011/02/12. 10.1111/j.1438-8677.2010.00375.x PubMed . [DOI] [PubMed] [Google Scholar]
- 60. Tong Z, Hong B, Yang Y, Li Q, Ma N, Ma C, et al. Overexpression of two chrysanthemum DgDREB1 group genes causing delayed flowering or dwarfism in Arabidopsis. Plant Mol Biol. 2009;71(1–2):115–29. 10.1007/s11103-009-9513-y PubMed . [DOI] [PubMed] [Google Scholar]
- 61. Tayeh N, Bahrman N, Sellier H, Bluteau A, Blassiau C, Fourment J, et al. A tandem array of CBF/DREB1 genes is located in a major freezing tolerance QTL region on Medicago truncatula chromosome 6. BMC Genomics. 2013;14:814 Epub 2013/11/23. 10.1186/1471-2164-14-814 PubMed . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Lin H, Zhu W, Silva JC, Gu X, Buell CR. Intron gain and loss in segmentally duplicated genes in rice. Genome Biol. 2006;7(5):R41 gb-2006-7-5-r41 [pii] 10.1186/gb-2006-7-5-r41 PubMed [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Duarte JM, Cui LY, Wall PK, Zhang Q, Zhang XH, Leebens-Mack J, et al. Expression pattern shifts following duplication indicative of subfunctionalization and neofunctionalization in regulatory genes of Arabidopsis. Molecular Biology and Evolution. 2006;23(2):469–78. 10.1093/molbev/msj051 PubMed WOS:000234718800023. [DOI] [PubMed] [Google Scholar]
- 64. Sandve SR, Fjellheim S. Did gene family expansions during the Eocene-Oligocene boundary climate cooling play a role in Pooideae adaptation to cool climates? Mol Ecol. 2010;19(10):2075–88. MEC4629 [pii] 10.1111/j.1365-294X.2010.04629.x PubMed . [DOI] [PubMed] [Google Scholar]
- 65. Florence RG. Ecology and silviculture of eucalypt forests: Csiro; 1996. [Google Scholar]
- 66. Bowman DMJS. Australian rainforests: islands of green in a land of fire Cambridge University Press; 2000. [Google Scholar]
- 67.Keller G, Cao P, San Clemente H, El Kayal W, Marque C, Teulières C. Transcript profiling combined with functional annotation of 2,662 ESTs provides a molecular picture of Eucalyptus gunnii cold acclimation. Trees—Structure and Function. 2013:1–23. 10.1007/s00468-013-0918-5 [DOI]
- 68. Haake V, Cook D, Riechmann JL, Pineda O, Thomashow MF, Zhang JZ. Transcription factor CBF4 is a regulator of drought adaptation in Arabidopsis. Plant Physiology. 2002;130(2):639–48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Lawton-Rauh A. Evolutionary dynamics of duplicated genes in plants. Mol Phylogen Evol. 2003;29(3):396–409. 10.1016/j.ympev.2003.07.004 PubMed WOS:000186738000003. [DOI] [PubMed] [Google Scholar]
- 70. Xiao H, Tattersall EA, Siddiqua MK, Cramer GR, Nassuth A. CBF4 is a unique member of the CBF transcription factor family of Vitis vinifera and Vitis riparia. Plant Cell and Environment. 2008;31(1):1–10. PubMed . [DOI] [PubMed] [Google Scholar]
- 71. Haake V, Cook D, Riechmann JL, Pineda O, Thomashow MF, Zhang JZ. Transcription factor CBF4 is a regulator of drought adaptation in Arabidopsis. Plant Physiol. 2002;130(2):639–48. 10.1104/pp.006478 PubMed [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Chen J, Xia X, Yin W. Expression profiling and functional characterization of a DREB2-type gene from Populus euphratica. Biochem Biophys Res Commun. 2009;378(3):483–7. 10.1016/j.bbrc.2008.11.071 [DOI] [PubMed] [Google Scholar]
- 73. Song Y, Chen Q, Ci D, Shao X, Zhang D. Effects of high temperature on photosynthesis and related gene expression in poplar. BMC Plant Biol. 2014;14:111 Epub 2014/04/30. 10.1186/1471-2229-14-111 PubMed [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Martin HA. Cenozoic climatic change and the development of the arid vegetation in Australia. Journal of Arid Environments. 2006;66(3):533–63. 10.1016/j.jaridenv.2006.01.009. [DOI] [Google Scholar]
- 75. Moss PT, Kershaw PA. The last glacial cycle from the humid tropics of northeastern Australia: comparison of a terrestrial and a marine record. Palaeogeogr, Palaeoclimatol, Palaeoecol. 2000;155(1–2):155–76. 10.1016/s0031-0182(99)00099-1 [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The tree was generated using Mega 5 program by the Maximum Likelihood method. Bootstrap values are indicated at each branch. For the E. grandis DREB subfamily, the relation-ship between Phytozome gene IDs and the names used in this paper is indicated in S2 Table.
(TIF)
The tree was generated using Mega 5 program by the Maximum Likelihood method. Bootstrap values are indicated at each branch. For the E. grandis ERF subfamily, the relation-ship between Phytozome gene IDs and the names used in this paper is indicated in S2 Table.
(TIF)
The tree was generated using Mega 5 program by the Maximum Likelihood method. Bootstrap values are indicated at each branch. For the E. grandis RAV subfamily, the relation-ship between Phytozome gene IDs and the names used in this paper is indicated in S2 Table.
(TIF)
The tree was generated using Mega 5 program by Maximum Likelihood method. Bootstrap values are indicated at each branch. For the E. grandis AP2 subfamily, subfamily, the relation-ship between Phytozome gene IDs and the names used in this paper is indicated in S2 Table.
(TIF)
(DOC)
“Eucagen name” associates the subfamily name with the relative position on the scaffolds (1 to 11). “Generic name” given to EgrCBF(1–17) and EgrDREB2-(1–6) from literature uses and genome location.
(DOC)
Data Availability Statement
All relevant data are within the paper and its supporting information files, accession numbers and genome location on genomic sequence (Phytozome database).