

Abstract
The TNF receptor family member BAFFR is essential for providing mature B cells with pro-survival signals and has recently been claimed to transduce these, though not exclusively, via a Syk-dependent signaling hub that feeds into ERK/AKT activation. In this issue of The EMBO Journal, Hobeika et al (2015) describe a synergistic pro-survival scenario involving BAFFR and CD19, which remains functional under Syk null conditions and is able to maintain mature B-cell survival. The authors hence propose a BAFFR-/CD19-driven mechanism to act in parallel with homeostatic NF-κB/AKT activation in non-stimulated B cells.
See also: E Hobeika et al (April 2015)
A large body of literature has convincingly demonstrated that mammalian B-cell development only proceeds normally when an array of cell surface receptors is present and properly functions at certain developmental time windows. One of the most essential components during B-cell development in the bone marrow is the B-cell receptor (BCR), which comes in two forms both programmed to serve discrete development checkpoints: the pre-BCR, in which successfully recombined heavy chains associate with surrogate light chains, to allow for cellular proliferation and further differentiation (positive selection), and the mature BCR, which is rather stringently probed for receptor autoreactivity (negative selection) (Monroe et al, 2003). In both cases, the spleen tyrosine kinase (Syk) associates either chronically or inducibly with the intracellular signaling domains of the BCR as the central player in the corollary signal transduction. After having reached the periphery, the size of the recirculating B-cell pool in the secondary lymphoid organs depends on the interaction of BAFFR (B-cell-activating factor receptor, TNFRSF13C) with its ligand BAFF (BlyS) (Thompson et al, 2001). Binding of BAFF to BAFFR triggers via NIK-/IKK1-dependent signaling the processing of NF-κB2 from the p100 to the p52 form. This, besides the activation of PI3K, alters gene expression and ultimately favors cellular survival. A recent study (Schweighoffer et al, 2013) provided first surprising evidence that Syk, apart from its function in tonic BCR signaling, acts as a BAFFR signaling hub by being activated upon BAFFR stimulation and feeding into the PI3K pathway. Yet, it remained unclear how BAFFR communicates with the BCR signaling subunits (Fig1A).
Figure 1.
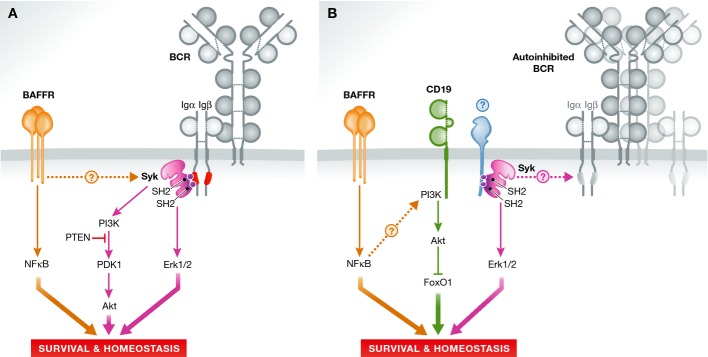
BAFFR provides mature B cells with indispensable survival signals
Inducible deletion of Syk in B cells allows testing its contribution to B-cell receptor (BCR) signaling and to the homeostatic maintenance of B cells. (A) Syk has been suggested to signal Akt and Erk activation downstream of BAFFR. A key step in this route would be BAFFR-induced phosphorylation of Syk and the ITAM motif (red) in the cytoplasmic tail of the BCR-associated Igα signaling chain. (B) An alternative model now describes synergistic pro-survival signaling by BAFFR and CD19 in addition to Syk-mediated survival signals. Here, CD19 signals Akt activation, while BAFFR and Syk signal independently. If Syk signaling under homeostatic conditions involves the BCR or other receptor systems (blue) remains open.
By applying an elegant, inducible, and B-cell-specific Syk deletion approach (Sykfl/fl;mb1cre-ERT2), Hobeika et al (2015) now reprobe the importance of Syk in BAFFR-mediated survival signaling and arrive at a somewhat different scenario. It is important to mention that constitutive deletion of Syk not only has a dramatic effect on the reactivity of various hematopoietic lineages (Kiefer et al, 1998; Bohmer et al, 2010) but especially on the nascent B-cell pool, essentially abrogating any B-cell development beyond the immature stage (Cheng et al, 1995; Turner et al, 1995). After having validated the functionality of their deletion approach, Hobeika et al (2015) first showed that tamoxifen-mediated deletion of Syk had a heterogenic effect on the survival of peripheral B cells. While splenic marginal zone B cells and peritoneal B1 cells were strongly reduced in numbers, transitional B cells were surprisingly unaffected and mature splenic B cells were only reduced to an amount that allowed further analysis of cellular reactivity. These mature Syk−/− remainder B cells turned out to be unable to mobilize Ca2+ upon BCR or latrunculin stimulation and exhibited inferior total tyrosine phosphorylation upon pervanadate treatment when compared to controls. Interestingly, these cells were unable to activate mTORC1 and exhibited a reduced potential to migrate toward CXCL12, which is in line with the reported function of Syk in B-cell polarization (Pearce et al, 2011). In contrast, Syk deficiency had no effect on TLR4 (LPS), TLR9 (CpG), IL-4, or CD40 stimulation, showing that these receptors function independently.
In the next step, Hobeika et al (2015) addressed the potential of Sykfl/fl;mb1cre-ERT2 B cells to populate Rag2−/−;yc−/− mice, which are void of intrinsic B and T cells under competitive and non-competitive transfer conditions. Even two months post-transfer, a significant portion of Syk-deleted cells persisted in the presence of wild-type competitor B cells, immediately questioning their potential to respond to BAFFR-mediated pro-survival signals. Surprisingly and in noteworthy contrast to Schweighoffer et al (2013), the authors showed that when cultured in vitro, tamoxifen-induced Sykfl/fl B cells still retained responsiveness to BAFF. In the presence of BAFF, tamoxifen-induced Sykfl/fl B cells were only reduced by half in numbers when compared to wild-type control cells and presented with negligible changes in BAFFR expression. To investigate the possibility of a persisting partial dependence on BAFFR signaling in vivo, the authors further treated Sykfl/fl;mb1cre-ERT2 and control mice with an antibody blocking the interaction of BAFF/BAFFR. In accordance with their in vitro findings, blocking BAFFR with neutralizing antibodies under Syk null conditions precipitated a comparable reduction in mature follicular B-cell frequencies in vivo, arguing for an actively employed and functional BAFFR-mediated survival route when Syk is not expressed (Fig1B).
Loss of the BCR mIgM heavy chain leads to the death of B cells, which can be rescued by a constitutively active version of the lipid kinase PI3K (Srinivasan et al, 2009). In a final and laborious approach, Hobeika et al (2015) set out to investigate this previously reported importance of the CD19/PI3K kinase pathway for B-cell homeostasis by intercrossing Sykfl/fl;mb1cre-ERT2 mice with CD19−/− mice. While Sykfl/fl;mb1cre-ERT2 B-cell frequencies resembled those of CD19−/− mice, a combined deficiency yielded severely compromised bone marrow and peripheral B220+CD19+ B-cell numbers. The fact that the homozygous introduction, on top of that, of an inducibly deletable FoxO1 allele (yielding FoxO1fl/fl;Sykfl/fl;mb1cre-ERT2;CD19−/− mice) restored B-cell numbers to levels of Sykfl/fl;mb1cre-ERT2 mice led Hobeika et al (2015) to the conclusion that CD19 indeed boosts B-cell survival via PI3K-mediated degradation of the pro-apoptotic transcription factor FoxO1. In conclusion, the authors therefore suggest synergistic pro-survival functions of BAFFR and CD19 operative in mature B cells (Fig1B).
The study by Hobeika et al (2015) significantly deepens our insight into the mechanisms ensuring appropriate populations of peripheral B cells. While it provides strong genetic evidence for PI3K-mediated survival signaling from CD19, the origin of the Syk-mediated survival signal in resting B cells remains to be clarified. The long-standing model of stimulation-induced recruitment and activation of Syk to the BCR has recently been supported in a number of elegant studies investigating the nano-composition of the B-cell membrane (reviewed in Maity et al, 2014). Consequently, in resting peripheral B cells, the BCR would be unlikely to signal survival via Syk. Possible scenarios explaining the pro-survival function of Syk might involve recurrent, highly transient episodes of BCR-mediated activation or other receptor systems that might engage Syk by cooption of ITAM-bearing receptors.
References
- Bohmer R, Neuhaus B, Buhren S, Zhang D, Stehling M, Bock B, Kiefer F. Regulation of developmental lymphangiogenesis by Syk(+) leukocytes. Dev Cell. 2010;18:437–449. doi: 10.1016/j.devcel.2010.01.009. [DOI] [PubMed] [Google Scholar]
- Cheng AM, Rowley B, Pao W, Hayday A, Bolen JB, Pawson T. Syk tyrosine kinase required for mouse viability and B-cell development. Nature. 1995;378:303–306. doi: 10.1038/378303a0. [DOI] [PubMed] [Google Scholar]
- Hobeika E, Levit-Zerdou E, Anastasopoulou V, Pohlmeyer R, Altmeier S, Alsadeq A, Dobenecker M-W, Pelanda R, Reth M. CD19 and BAFF-R can signal to promote B cell survival in the absence of Syk. EMBO J. 2015;34:925–939. doi: 10.15252/embj.201489732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiefer F, Brumell J, Al-Alawi N, Latour S, Cheng A, Veillette A, Grinstein S, Pawson T. The Syk protein tyrosine kinase is essential for Fcgamma receptor signaling in macrophages and neutrophils. Mol Cell Biol. 1998;18:4209–4220. doi: 10.1128/mcb.18.7.4209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maity PC, Yang J, Klaesener K, Reth M. The nanoscale organization of the B lymphocyte membrane. Biochim Biophys Acta. 2014;1853:830–840. doi: 10.1016/j.bbamcr.2014.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monroe JG, Bannish G, Fuentes-Panana EM, King LB, Sandel PC, Chung J, Sater R. Positive and negative selection during B lymphocyte development. Immunol Res. 2003;27:427–442. doi: 10.1385/IR:27:2-3:427. [DOI] [PubMed] [Google Scholar]
- Pearce G, Audzevich T, Jessberger R. SYK regulates B-cell migration by phosphorylation of the F-actin interacting protein SWAP-70. Blood. 2011;117:1574–1584. doi: 10.1182/blood-2010-07-295659. [DOI] [PubMed] [Google Scholar]
- Schweighoffer E, Vanes L, Nys J, Cantrell D, McCleary S, Smithers N, Tybulewicz VL. The BAFF receptor transduces survival signals by co-opting the B cell receptor signaling pathway. Immunity. 2013;38:475–488. doi: 10.1016/j.immuni.2012.11.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Srinivasan L, Sasaki Y, Calado DP, Zhang B, Paik JH, DePinho RA, Kutok JL, Kearney JF, Otipoby KL, Rajewsky K. PI3 kinase signals BCR-dependent mature B cell survival. Cell. 2009;139:573–586. doi: 10.1016/j.cell.2009.08.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson JS, Bixler SA, Qian F, Vora K, Scott ML, Cachero TG, Hession C, Schneider P, Sizing ID, Mullen C, Strauch K, Zafari M, Benjamin CD, Tschopp J, Browning JL, Ambrose C. BAFF-R, a newly identified TNF receptor that specifically interacts with BAFF. Science. 2001;293:2108–2111. doi: 10.1126/science.1061965. [DOI] [PubMed] [Google Scholar]
- Turner M, Mee PJ, Costello PS, Williams O, Price AA, Duddy LP, Furlong MT, Geahlen RL, Tybulewicz VL. Perinatal lethality and blocked B-cell development in mice lacking the tyrosine kinase Syk. Nature. 1995;378:298–302. doi: 10.1038/378298a0. [DOI] [PubMed] [Google Scholar]