

Abstract
Some of the most devastating agricultural diseases are caused by root-infecting pathogens, yet the majority of studies on these interactions to date have focused on the host responses of aerial tissues rather than those belowground. Fusarium oxysporum is a root-infecting pathogen that causes wilt disease on several plant species including Arabidopsis thaliana. To investigate and compare transcriptional changes triggered by F. oxysporum in different Arabidopsis tissues, we infected soil-grown plants with F. oxysporum and subjected root and leaf tissue harvested at early and late timepoints to RNA-seq analyses. At least half of the genes induced or repressed by F. oxysporum showed tissue-specific regulation. Regulators of auxin and ABA signalling, mannose binding lectins and peroxidases showed strong differential expression in root tissue. We demonstrate that ARF2 and PRX33, two genes regulated in the roots, promote susceptibility to F. oxysporum. In the leaves, defensins and genes associated with the response to auxin, cold and senescence were strongly regulated while jasmonate biosynthesis and signalling genes were induced throughout the plant.
Introduction
Plants are constantly exposed to microbes and pathogens and must coordinate tightly regulated immune responses to ensure their survival and reproductive success. The rhizosphere is a rich source of microorganisms, and some of the most destructive plant diseases are caused by root infecting fungal or oomycete pathogens such as Fusarium, Rhizoctonia, Pythium and Phytophthora species. Although these pathogens infect root tissues, visible disease symptoms often appear in above-ground parts of the plant. Therefore, most studies to date have focused on the response of aerial tissue rather than the roots to such pathogens [1]. Since roots and shoots are normally exposed to different microorganisms, it has been suggested that the two tissues may have evolved defence strategies that differ in amplitude, timing or specificity [2]. However, there is a scarcity of studies comparing the response of roots and shoots to root-infecting fungal pathogens to directly test these hypotheses.
Fusarium oxysporum is a ubiquitous root-infecting fungal pathogen that causes wilt disease on several plant species including Arabidopsis thaliana. F. oxysporum is considered a hemibiotrophic pathogen because it begins its infection cycle as a biotroph but later changes to a necrotroph. In the biotrophic phase, F. oxysporum establishes infection via the roots and travels towards the vasculature. Upon perception of fungal elicitors, plants mount a basal defence response (PTI) characterised by an oxidative burst, cell wall callose deposition and transcriptional changes which is designed to inhibit microbial colonisation. Successful pathogens such as F. oxysporum have evolved mechanisms to overcome this relatively weak defence response and colonise the plant [3]. Once in the vasculature, F. oxysporum travels upwards in the plant and accumulation of fungal mycelia and defence related compounds in the xylem cause vascular wilting. As the infection progresses, F. oxysporum changes to a necrotrophic pathogen, causing foliar necrosis, lesion development and eventual plant death. F. oxysporum is thought to secrete phototoxic compounds which cause root cell collapse and veinal chlorosis in the leaves and is also proposed to reprogramme the host to induce senescence and facilitate disease during the necrotrophic phase of infection [4, 5]. Fungal effectors such as secreted-in-xylem 4 (SIX 4) promote infection most likely by promoting host JA responses [6]. Interestingly, the jasmonate (JA) signalling pathway both negatively and positively contributes to F. oxysporum resistance in A. thaliana. F. oxysporum produces bioactive jasmonates which presumably promote host senescence [7] while the JA receptor mutant coi1 exhibits extreme resistance to F. oxysporum, suggesting that F. oxysporum hijacks the host JA signalling pathway to facilitate disease [8]. Adding to the complexity, JA-dependent defences may also have a relatively small but significant effect on this pathogen [9]. Other phytohormone pathways have also been shown to regulate the host response to F. oxysporum. Ethylene and auxin are both thought to negatively regulate F. oxysporum resistance since the ethylene receptor mutant etr1-1 and auxin signalling and transport mutants show increased resistance to F. oxysporum [10, 11]. The stress hormone ABA also promotes susceptibility to this pathogen [12]. Salicylic acid promotes resistance to F. oxysporum, presumably acting in the biotrophic phase of infection, although this seems to be somewhat isolate-dependent [7, 13–15].
In this study, we comparatively analysed the below- and above-ground defence responses occurring in A. thaliana in response to infection by F. oxysporum. Transcriptional reprogramming by F. oxysporum differed considerably depending on the tissue and timepoint after infection.
Materials and Methods
Plant materials and growth conditions, inoculation, tissue collection and disease assessment
A. thaliana Col-0 seeds were used for RNA-seq analyses. The T-DNA insertion lines arf1-5 (SALK_079046), arf2-6 (CS24600), arf2-8 (CS24602) and double mutant arf2-6/arf1-3 (CS24631) have been previously described [16–18]. T-DNA insertion mutants prx33-1 (SALK_056847C) and prx33-2 (SALK_062314C) were obtained from the Arabidopsis Biological Resource Centre (ABRC).
Inoculations for tissue collection and disease assessment were carried out as follows: seeds were placed on damp soil and imbibed for two days at 4°C to synchronise germination. Plants were then grown under short day conditions (8h photoperiod) at 21°C for four weeks. The F. oxysporum isolate used in this study was strain Fo5176 obtained from Dr Roger Shivas, Queensland Plant Pathology Herbarium, Brisbane, Australia. Inoculations were performed as described previously [12]. Briefly, roots of 4-week-old plants were uprooted and then dipped either in water for the mock inoculation or in a F. oxysporum suspension containing 1 × 106 spores ml-1, replanted and placed under long day growth conditions (16 h photoperiod) at 28°C. At 24 hours and 6 days after inoculation, root and leaf tissue (three independent biological replicates, each containing a pool of ten plants) was harvested and frozen in liquid nitrogen.
Disease was measured by visually assessing symptom development on the leaves at 14 dpi either using a scale of 0–5 with 0 being asymptomatic and 5 being dead as described previously [9] or by percentage of diseased leaves [19]. At least 16 plants were assessed per line.
In-vitro plant growth and F. oxysporum inoculations
In-vitro F. oxysporum inoculation assays were performed as described previously [20]. Sterilised seeds were placed on 1x Murashige and Skoog (MS) agar supplemented with 1% sucrose and incubated at 4°C for two days, then grown under short day conditions (8h photoperiod) at 21°C. The roots of two-week-old seedlings were dipped in either a F. oxysporum suspension containing 1 × 106 spores ml-1 or water under sterile conditions, placed onto sucrose-free 1xMS agar plates and returned to short day growth conditions. At 20 dpi, the number of lateral roots arising from the primary root was counted in a 2cm region starting 0.5cm down from the hypocotyl. Root length was manually measured at 20 dpi. At least 25 plants were assessed per line.
RNA-seq analysis
Total RNA from mock and F. oxysporum-infected tissue was extracted and DNAse treated using the RNeasy mini kit (Qiagen) according to the manufacturer’s instructions. RNA integrity was confirmed using the Agilent 2100 bioanalyser Plant Nano system (Agilent Biotechnologies). Library preparation and sequencing were performed by the Australian Genome Research Facility (AGRF). Messenger RNA was selected using Poly-A tail selection prior to preparation of 50bp single end read libraries. Sequencing was performed on an Illumina HiSeq 2000 system generating approximately from 6.5 to 16 million raw RNA-seq reads per sample.
Differential expression analysis was performed using the Tuxedo analysis suite [21]. Briefly, Bowtie2 along with Tophat were used to align generated reads to the TAIR10 A. thaliana reference genome. After expressed transfrags were assembled, Cufflinks was used to quantify gene abundance and transcriptome assemblies were then merged using Cuffmerge. To identify genes differentially expressed by F. oxysporum, Cuffdiff was performed on the following comparisons: F. oxysporum inoculated roots 1 day post inoculation (dpi) versus mock inoculated roots 1dpi (FR1/MR1); F. oxysporum inoculated leaves 1 dpi versus mock inoculated leaves 1dpi (FL1/ML1); FR6/MR6 and FL6/ML6. Statistical analysis was performed within the Cufflinks analysis with false discovery rate and correction for multiple comparisons applied using standard run parameters. Genes considered differentially expressed showed a statistically significant difference in expression values (P<0.05). 0.6–2.2% of reads did not map to the A. thaliana genome. Reads that did not align to annotated transcripts were omitted from the analysis. For reads that mapped to two transcripts, the least significantly aligned transcript(s) were omitted.
To determine the tissue or timepoint specificity of DEGs, venn diagrams and gene lists were obtained using http://genevenn.sourceforge.net/. To determine functionality of DEGs, genone ontology (GO) enrichment analysis was performed using http://bioinfo.cau.edu.cn/agriGO/ [22]. Sequence data are available from NCBI under Sequence Read Archive (SRA) accession SRP052276.
F. oxysporum RT-qPCR
First-strand cDNA was synthesized from 1μg RNA using SuperScript RNA H- Reverse Transcriptase (Invitrogen) and oligo (dT) primer, according to the manufacturer's instructions. RT-qPCR products were amplified in 10μl reactions containing 2μl cDNA, 2x SYBR® Green PCR Master Mix (Applied Biosystems) and 0.3 pmol primers. Plant actin (AT3G18780) was quantified using primers qACT2-F and qACT2-R [23] and F. oxysporum β-tubulin was quantified using primers Foβ-Tub-F and Foβ-Tub-R [9] using the following cycling conditions: 50°C for 2 mins, 95°C for 10 mins, then 40 cycles of: 95°C for 15 sec, 60°C for 40 sec. Quantities were determined against standard curves from cDNA derived from plant or in-vitro grown F. oxysporum using ViiA7 software (version 1.2; Agilent Technologies). Three biological replicates and 3 technical replicates were assessed per sample.
Results and Discussion
To compare the response of A. thaliana roots and leaves to F. oxysporum infection, we performed RNA-seq experiments and identified genes significantly induced or repressed by F. oxysporum in either tissue at 1 or 6 days post inoculation (dpi). We used 1 dpi as an early timepoint when F. oxysporum is presumably in a biotrophic infection phase and 6 dpi when F. oxysporum is undergoing the switch from a biotrophic to necrotrophic lifestyle. At 1 dpi, F. oxysporum—inoculated plants were indistinguishable from mock—inoculated controls and F. oxysporum RNA was detectable in the roots but not in the leaves. By 6 dpi, classic disease symptoms including vein yellowing and chlorosis appeared on the leaves of Col-0 plants and fungal DNA and RNA was detectable in both root and leaf tissue (Fig 1).
Fig 1. Disease symptoms and detection of F. oxysporum RNA in A. thaliana plants after F. oxysporum infection.
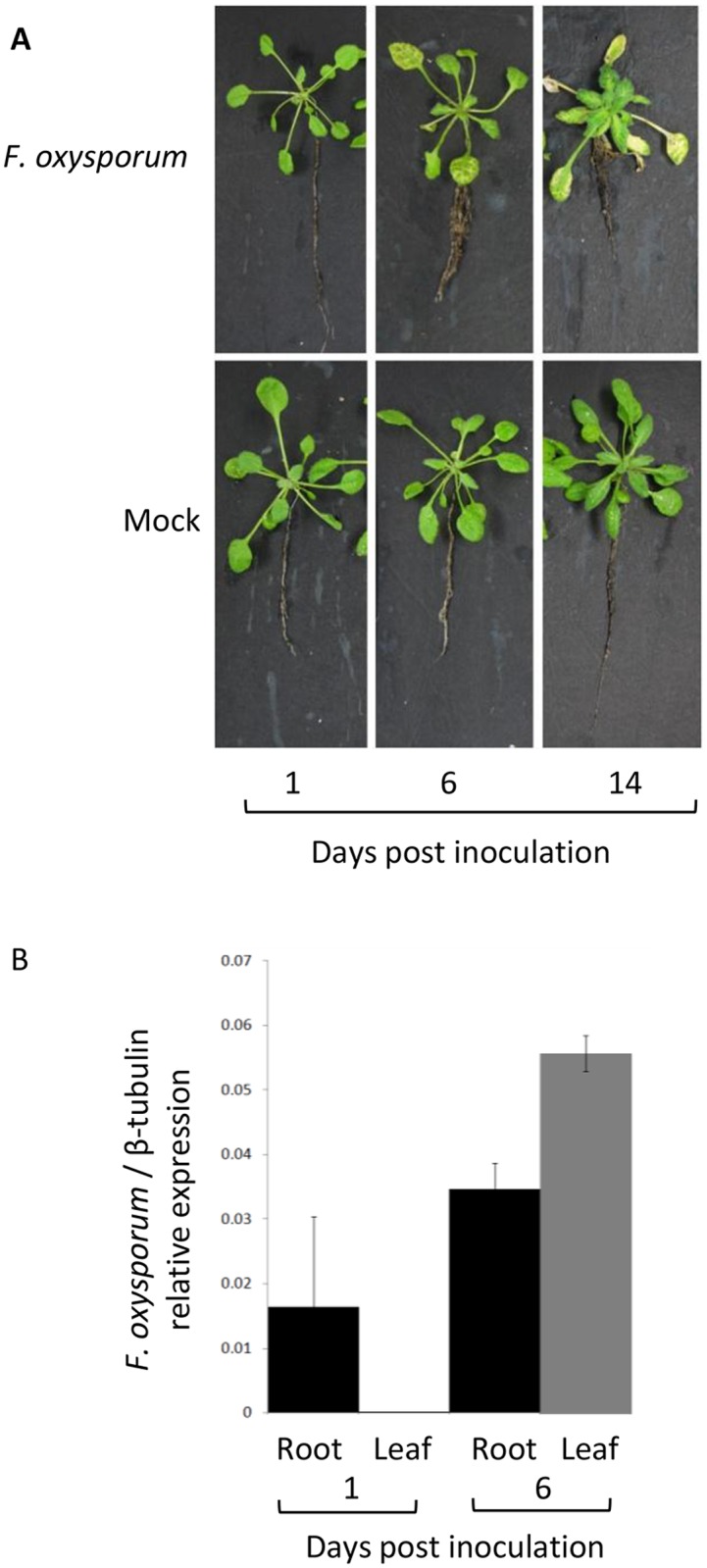
(A) Representative plants were uprooted and photographed at 1, 6 or 14 days after F. oxysporum or mock inoculation. (B) Relative expression of F. oxysporum β-tubulin normalised to A. thaliana actin in root (black) and leaf (grey) tissue. Data show mean relative expression and standard error from 3 biological replicates containing pools of 10 plants.
Tissue and infection stage—specific transcriptional changes are distinct
For differential expression analysis of RNAseq data, we used the cuffdiff analysis tool and found substantial numbers of genes, ranging from 417 to 4089, were regulated by F. oxysporum depending on the timepoint, in roots and leaves. Within each timepoint, similar numbers of genes were repressed as were induced. More genes were regulated in the leaves than in the roots at both timepoints and substantially more genes were regulated at 6 dpi relative to 1 dpi (Fig 2A).
Fig 2. Genes induced or repressed in different tissues at early and late timepoints after F. oxysporum infection.
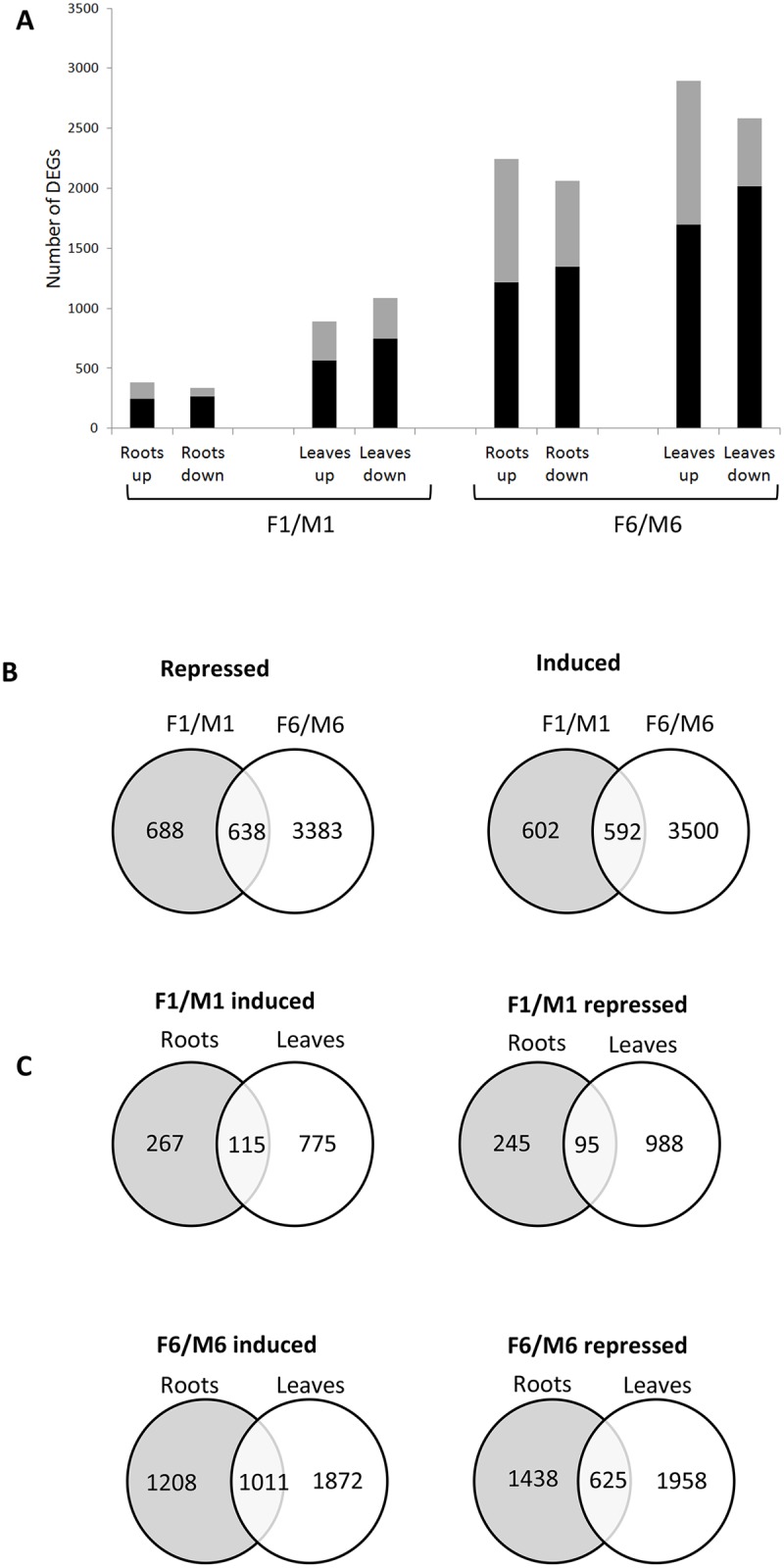
(A) The number of genes induced (up) or repressed (down) by F. oxysporum in roots and leaves at 1 or 6 dpi. Black bars represent genes regulated <2 fold; grey bars represent genes regulated > 2 fold. (B) Overlap in F. oxysporum—responsive genes between two timepoints, 1 (F1/M1) and 6 dpi (F6/M6). (C) Overlap in F. oxysporum—responsive genes between roots and leaves.
Comparison of gene regulation in infection timepoints
We compared the genes regulated at 1 dpi to the genes regulated at 6 dpi. Approximately half of the genes regulated by F. oxysporum at 1 dpi and 14% of the genes regulated at 6 dpi were regulated at both timepoints (Fig 2B). Secondary responses associated with hormone feedback and crosstalk may be occurring to regulate plant growth once infection has been established, accounting for the greater number of DEGs at 6 dpi. In addition, transcriptomic responses to both biotrophic and necrotrophic growth are likely to be occurring during this transitional phase of infection.
Among the genes that were regulated at both timepoints, the fold induction or repression was higher at 6 dpi than at 1 dpi (S1A Fig). The greater amplitude of regulation at the later timepoint in the leaves may be explained by the lack of direct foliar contact with the fungus at 1 dpi compared to 6 dpi. Innate immunity responses generated at 1 dpi may be increased in amplitude to more strongly defend the plant once robust infection has been established at 6 dpi.
Comparison of gene regulation in infected tissues
We next compared DEGs found in roots versus the leaves. More genes were regulated in the leaves than the roots at 1 dpi, even though F. oxysporum RNA was undetectable in the leaves at this early timepoint. In Brassica oleracea, untreated shoots respond to JA within hours of root JA treatment. This systemic response is thought to be due to the reallocation of defence compounds and primary metabolites between roots and shoots [24].
At 1 dpi, 30% of induced root genes and 12% of induced leaf genes were induced in both tissues while 45% of repressed root and 8% of repressed leaf genes were repressed in both tissues (Fig 2C). At 6 dpi, 30–45 and 25–35% of genes regulated in roots and leaves, respectively, were regulated in both tissues.
Following inoculation with the ascomycete pathogen Colletotrichum graminicola, the roots respond more strongly than leaves, while roots and shoots show a quantitatively different response to JA treatment [24, 25]. We asked whether roots responded more strongly to F. oxysporum than the leaves by performing pairwise comparisons on the fold induction or repression of genes that were regulated in both tissues (shared DEGs). Induction of shared DEGs was of a similar order of magnitude in both tissues, while repression of shared DEGs was slightly stronger in the leaves at 1 dpi and slightly stronger in the roots at 6 dpi (S1B Fig). Therefore, in the A. thaliana—F. oxysporum interaction, the amplitude of the transcriptional response is similar between roots and leaves.
To examine the pervasiveness of tissue specificity, we investigated the number of DEGs that showed tissue-specific regulation throughout the experiment. Over 50% of genes that were induced specifically in one tissue at one timepoint were exclusively regulated in that tissue throughout the timecourse (S2A Fig). We next asked whether the genes that show tissue-specific induction at 1 dpi show tissue specific expression under all conditions tested in this experiment. Of the 267 genes induced specifically in the roots at 1 dpi, only 3 were undetectable in leaves in either mock or F. oxysporum inoculated tissue in this experiment, while of the 775 genes induced specifically in the leaves at 1 dpi, 7 were undetectable in roots. These findings demonstrate that tissue-specific F. oxysporum responsive DEGs are not necessarily expressed only in that tissue under normal conditions.
Comparison with published transcriptomic studies in the F. oxysporum—A. thaliana interaction
We found that F. oxysporum inoculation regulated transcription of a similar subset of novel genes in soil-grown plants as in agar-grown seedlings studied previously by [20] (Table A in S1 File). By separating root and leaf tissue prior to sequencing, we were able to show that several of the novel DEGs identified by [20] were regulated in specific tissues. In addition, less than half of the genes found to be regulated by F. oxysporum in the roots by [26] were similarly regulated in our study (Table B in S1 File). The reasons for this discrepancy are unclear, but could be due to differences in the sampling time points (2 dpi in [20] versus 1 or 6 dpi in this study) or the different technologies (microarray in [26] vs RNA-seq in this study) used in the transcriptome analyses. A recent study has demonstrated that RNA-seq has greater sensitivity and accuracy than microarray analyses [27]. Weakly expressed DEGs may have been detected using RNA-seq in our study but been below the sensitivity threshold using a microarray.
Key functional processes showing tissue specificity
Gene ontology (GO) term singular enrichment analysis was used to identify differences in functionality between genes regulated in roots and shoots (Table C in S1 File). In general, the most significantly overrepresented functional categories were defense-related. Fig 3 summarises key processes that are transcriptionally responsive to F. oxysporum in A. thaliana. The observed induction of JA-associated genes and auxin biosynthesis genes and repression of photosynthetic genes in the leaves is consistent with trends found in previous transcriptomic studies [11, 20]. F. oxysporum-regulation of mannose binding lectins and cold-responsive genes has not been described in detail elsewhere.
Fig 3. Key processes that undergo transcriptional reprogramming in response to F. oxysporum infection.

Arrows indicate trend of transcriptional regulation in each of the functional categories, which were chosen based on the strength of the transcriptional response strength and the number of genes regulated. An example of a gene in each functional category is given. Upon perception of F. oxysporum in the roots, the basal defence response is elicited. F. oxysporum effectors, toxins and hormone mimics such as bioactive JAs trigger transcriptional changes and production of host-derived signalling compounds. Systemic movement of both fungal and host derived signalling molecules elicit transcriptional changes in the leaves ahead of the fungus. Transcriptional changes in both tissues are a consequence of active defence responses (i.e. production of antifungal compounds, defence signalling) F. oxysporum- driven manipulation (i.e. increased senescence in the leaves to facilitate necrotrophic infection), and the response of the plant to stress incurred by infection (i.e. changes in photosynthesis and flowering time minimise the fitness cost to the plant).
Plant-wide processes responsive to F. oxysporum infection
Genes repressed in either tissue at 6 dpi showed an enrichment of genes associated with general biosynthesis and metabolomic processes which could be a consequence of the plant reallocating resources from development to defense. Given the established roles of JA and auxin in the F. oxysporum—A. thaliana interaction [8, 11], it could be expected that biosynthesis, signalling and transport genes in these hormone pathways may be regulated. Indeed, the majority of JA and auxin biosynthesis genes were induced in both tissues at 6 dpi. Several of the most strongly regulated auxin biosynthesis genes such as CYP83B1 and CYB71A13 catalyse the production of indole glucosinolates, which act as antimicrobial metabolites. JA signalling associated genes were also strongly induced in both tissues, with all JAZ genes induced at 6 dpi (Tables 1 and 2). The majority of JA and auxin biosynthesis genes were induced in both tissues. JA signalling associated genes were also strongly induced in both tissues, with all JAZ genes induced at 6 dpi (Tables 1 and 2).
Table 1. JA signalling and biosynthetic genes are induced in both tissues.
| TAIR | Description | F1/M1 | F6/M6 | ||
|---|---|---|---|---|---|
| Roots | Shoots | Roots | Shoots | ||
| JA biosynthesis | |||||
| AT3G25760 | ALLENE OXIDE CYCLASE 1 (AOC1) | - | - | - | 8.4 |
| AT3G25770 | ALLENE OXIDE CYCLASE 2 (AOC2) | - | - | - | 5.3 |
| AT1G19640 | JASMONIC ACID CARBOXYL METHYLTRANSFERASE (JMT) | - | - | - | 14.3 |
| AT1G20510 | OPC-8:0 CoA ligase1 (OPCL1) | - | - | 3.4 | 5.4 |
| AT1G74100 | SULFOTRANSFERASE 16 (SOT16) | - | - | 1.5 | 5.1 |
| AT2G06050 | OXOPHYTODIENOATE-REDUCTASE 3 (OPR3) | - | - | 3.8 | 9.8 |
| AT4G16760 | ACYL-COA OXIDASE 1 (ACX1) | - | - | 2.3 | 2.0 |
| AT3G48520 | CYTOCHROME P450 (CYP94B3) | - | 1.9 | 1.8 | 2.3 |
| AT2G46370 | JAR1 | - | - | 4.3 | 1.4 |
| JAZ | |||||
| AT1G19180 | JAZ1 | 1.6 | - | 6.2 | 5.6 |
| AT1G74950 | JAZ2 | - | - | 4.9 | 6.4 |
| AT3G17860 | JAZ3 | - | - | 3.6 | 1.8 |
| AT1G48500 | JAZ4 | - | - | 2.1 | 2.2 |
| AT1G17380 | JAZ5 | - | 1.9 | 9.3 | 8.9 |
| AT1G72450 | JAZ6 | 1.7 | - | 4.3 | 4.4 |
| AT2G34600 | JAZ7 | - | - | 3.7 | 7.3 |
| AT1G30135 | JAZ8 | - | - | - | 9.0 |
| AT1G70700 | JAZ9 | - | - | 5.7 | 5.6 |
| AT5G13220 | JAZ10 | 2.1 | 3.2 | 72.5 | 15.2 |
| JA-inducible defensins / antimicrobial proteins | |||||
| AT5G24770 | VEGETATIVE STORAGE PROTEIN 2 (VSP2) | 2.0 | - | 2.0 | 7.8 |
| AT5G24780 | VEGETATIVE STORAGE PROTEIN 1 (VSP1) | 2.1 | 3.1 | 2.0 | 12.2 |
| AT1G72260 | THIONIN 2.1 (THI2.1) | - | - | - | 14.0 |
| AT5G06870 | POLYGALACTURONASE INHIBITING PROTEIN 2 (PGIP2) | - | - | 16.0 | 4.0 |
| AT1G61120 | TERPENE SYNTHASE 04 (TPS04) | - | 3.4 | - | 13.3 |
| AT3G12500 | PATHOGENESIS-RELATED 3 (PR3) | 1.9 | - | 2.7 | 2.2 |
| AT5G44420 | PDF1.2 | - | 2.8 | - | 65.1 |
| AT2G26010 | PDF1.3 | - | - | - | 159.9 |
| AT5G44430 | PDF1.2C | - | - | - | 186.0 |
| AT2G26020 | PDF1.2b | - | - | - | 37.0 |
| AT5G63660 | PDF2.5 | - | - | 2.3 | - |
| AT1G19610 | PDF1.4 | - | 0.4 | 0.7 | 0.6 |
| AT1G13609 | DEFL family | - | 0.5 | - | - |
| AT2G16367 | DEFL family | - | - | - | 6.2 |
| AT1G64195 | DEFL family | - | 0.6 | - | 5.2 |
| AT2G43530 | DEFL family | - | - | 12.3 | 4.7 |
| AT1G13607 | DEFL family | - | 0.5 | - | 3.1 |
| AT1G76954 | DEFL family | - | - | - | 2.8 |
| AT3G59930 | DEFL family | - | - | 2.4 | 2.4 |
| AT3G05727 | DEFL family | - | 0.5 | - | 2.0 |
| AT4G22230 | DEFL family | - | - | 2.5 | - |
| AT4G22235 | DEFL family | - | - | 4.4 | - |
| AT4G22212 | DEFL family | - | - | 6.5 | - |
| AT4G22214 | DEFL family | 2.3 | - | 38.7 | - |
| AT2G43510 | TRYPSIN INHIBITOR PROTEIN 1 (TI1) | 2.2 | - | 2.6 | 1.4 |
JA biosynthetic, signalling or responsive genes regulated >2 fold. Number indicates fold induction or repression by F. oxysporum relative to mock treatment.
Table 2. Regulation of auxin biosynthetic, signalling and transport genes.
| TAIR10 | Name | F1/M1 | F6/M6 | ||
|---|---|---|---|---|---|
| Roots | Shoots | Roots | Shoots | ||
| Auxin biosynthesis | |||||
| AT4G13770 | CYP83A1 | - | - | 2.1 | 1.5 |
| AT4G31500 | CYP83B1 / ALTERED TRYPTOPHAN REGULATION 4 (ATR4) | - | - | - | 5.1 |
| AT2G30770 | CYP71A13 | 2.4 | - | 12.3 | 2.0 |
| AT1G24100 | UGT74B1 | - | - | 1.4 | 3.5 |
| AT1G04610 | YUCCA 3 (YUC3) | - | - | 3.0 | - |
| AT4G28720 | YUCCA 8 (YUC8) | - | - | 2.4 | 0.3 |
| AT2G20610 | SUPERROOT 1 (SUR1) | 1.4 | - | - | 2.4 |
| AT5G05730 | ANTHRANILATE SYNTHASE ALPHA SUBUNIT 1 (ASA1) | - | - | 3.9 | 4.5 |
| AT1G70560 | TRYPTOPHAN AMINOTRANSFERASE OF ARABIDOPSIS 1 (TAA1) | - | - | 2.3 | - |
| AT3G44300 | NITRILASE 2 (NIT2) | - | 2.6 | 2.5 | - |
| AT5G05590 | PHOSPHORIBOSYLANTHRANILATE ISOMERASE 2 (PAI2) | - | - | - | 2.2 |
| AT1G44350 | IAA-LEUCINE RESISTANT (ILR)-LIKE GENE 6 (ILL6) | - | - | 9.3 | 5.0 |
| AT1G51780 | IAA-LEUCINE RESISTANT (ILR)-LIKE GENE 5 (ILL5) | - | - | - | 30.9 |
| AT1G51760 | IAA-ALANINE RESISTANT 3 (IAR3) / JASMONIC ACID RESPONSIVE 3 (JAR3) | - | - | 3.1 | 5.9 |
| AT1G12200 | FLAVIN MONOOXYGENASE (FMO) | 1.3 | 0.1 | - | 1.4 |
| GRETCHEN HAGEN 3 family | |||||
| AT1G28130 | GH3.17 | - | - | 0.7 | 0.5 |
| AT5G13370 | GH3 family | 2.1 | 1.5 | 2.8 | |
| AT4G27260 | WES1 /GH3.5 | - | 0.7 | 0.6 | 0.3 |
| Auxin transport | |||||
| AT1G78100 | AUXIN UP-REGULATED F-BOX PROTEIN 1 (AUF1) | - | - | 0.4 | 0.6 |
| AT1G22220 | AUXIN UP-REGULATED F-BOX PROTEIN 2 (AUF2) | - | - | - | 2.3 |
| AT4G39403 | POLARIS (PLS) | - | - | - | 0.0 |
| AT2G38120 | AUXIN RESISTANT 1 (AUX1) | - | - | 0.5 | 0.5 |
| AT2G21050 | LIKE AUXIN RESISTANT 2 (LAX2) | 1.4 | - | 0.8 | 0.4 |
| AT5G13930 | TRANSPARENT TESTA 4 / CHALCONE SYNTHASE | - | 2.8 | - | 1.5 |
| AT3G05630 | PHOSPHOLIPASE D P2 | - | - | - | 2.2 |
| AT5G57090 | PIN 2 | - | - | 2.6 | - |
| AT1G70940 | PIN 3 | - | - | - | 0.5 |
| AT1G23080 | PIN 7 | - | - | 0.6 | 0.5 |
| AT1G76520 | PIN-LIKES 3 | - | - | 2.1 | 1.6 |
| AT2G17500 | PIN-LIKES 5 | - | - | - | 4.3 |
| AT2G14820 | NAKED PINS IN YUC MUTANTS 2 (NPY2) | - | - | 2.3 | - |
| AT2G23050 | NPY4 | - | - | 2.1 | - |
| AT5G67440 | NPY3 | - | - | 0.4 | - |
| AT4G37590 | NPY5 | - | - | 0.3 | - |
| Auxin/Indole-3-acetic acid (Aux/IAA) family | |||||
| AT5G43700 | IAA4 | - | 0.7 | 0.7 | 0.4 |
| AT5G65670 | IAA9 | - | - | 0.3 | - |
| AT4G14550 | IAA14 | - | - | 2.5 | - |
| AT1G19850 | IAA24 | - | - | 0.3 | - |
| AT3G16500 | IAA26 | - | - | 0.4 | - |
| AT5G25890 | IAA28 | 1.4 | 1.7 | 0.1 | - |
| AT4G32280 | IAA29 | - | 0.5 | - | 0.2 |
| AT3G62100 | IAA30 | - | - | 0.7 | 0.3 |
| Auxin response factors (ARFs) | |||||
| AT5G62000 | ARF2 | - | - | 0.4 | - |
| AT5G37020 | ARF8 | - | 1.5 | 0.4 | - |
| AT2G28350 | ARF10 | - | - | 0.5 | - |
| AT3G61830 | ARF18 | - | - | 0.5 | 0.8 |
| AT1G19220 | ARF19 | - | - | 0.4 | 0.8 |
| AT4G17788 | miRNA 160 targetting ARF10, ARF16, ARF17 | 24.2 | - | 0.8 | 55.0 |
| AT1G31173 | miRNA 167D targetting ARF6 and ARF8 | - | - | - | 0.6 |
Auxin signalling or auxin-responsive genes that are regulated >2 fold. Number indicates fold induction or repression by F. oxysporum relative to mock treatment.
Plant processes responsive to F. oxysporum infection in roots
The most strongly regulated genes in the roots are listed in Tables D and E in S1 File. Genes with known defensive roles were strongly regulated in the roots. For example, ER-LOCALIZED DnaJ-LIKE PROTEIN 3b (AT3G62600), a component of the ER quality control machinery involved in basal immunity [28] was strongly repressed in the roots at 1 dpi. TUBBY LIKE PROTEIN 5 (AT1G43640) promotes root colonisation by the mutualistic fungus Piriformospora indica [29] and was induced 73 fold in the roots at 6 dpi. Anthocyanins are suggested to play an important role in response to abiotic or biotic stress [30]. Genes involved in anthocyanin accumulation were some of the most strongly regulated genes in the roots at 1 dpi. Interestingly, of these FLS4 [31] were repressed while TRANSPARENT TESTA19 (TT19; [32]) and TT3 [33] were both induced.
Hormone signalling
JA and ethylene-associated genes were strongly regulated in the roots. ETO1-like, a paralog of ETO1 which represses ethylene biosynthesis was the most strongly repressed gene at 1 dpi. The second most highly-induced gene in infected roots at 1 dpi is NATA1 that encodes an ornithine N-delta-acetyltransferase involved in the production of JA-induced defensive metabolite [34] while at 6 dpi, the most responsive gene (induced 485 fold) was TAT3, a tyrosine aminotransferase responsive to JA. Several uncharacterised DEFL genes such as AT4G22214 (37 fold induction at 6 dpi) showed strong induction specifically in the roots (Table 1).
ABA is a major player in abiotic stress regulation but also contributes to defence [35]. It has been proposed that ABA may act as a root-to-shoot systemic resistance signal in some plant-microbe interactions [25]. Several ABA-associated genes were strongly regulated by F. oxysporum in the roots. At 6 dpi, the ABA receptor RCAR3 that was recently implicated in the immune response to bacteria [36] was repressed >10 fold; SNF1-related kinase2.2 (AT3G50500) which regulates the drought stress response [37] was repressed >40 fold and the ABA transporter PLEIOTROPIC DRUG RESISTANCE 12 (AT1G15520), which regulates resistance to the pathogen bacteria Ralstonia solanacearum [38] was induced 15 fold. At 1 dpi, AT5G35210, which encodes a PHD transcription factor required for ABI4 activation [39], was repressed >7 fold in the roots. Drought stress caused by the invasion of xylem vessels by F. oxysporum may contribute to the strong regulation of ABA-associated transcripts upon infection.
Mannose-binding lectins
Members of the Mannose-binding lectin (MBL) superfamily protein were among the most strongly induced genes in the roots (Table 3). AT1G52100 and AT5G35940 were induced >50 fold at 6 dpi, while AT5G28520 was induced >8 fold at 1 dpi. Although the specific function of MBLs in plant defence is currently unknown, it is thought that MBLs recognise potential invaders by binding to specific carbohydrates on foreign microorganisms [40]. Recently, a MBL from pepper was shown to mediate basal immunity and SA-mediated defences [38], while a MBL from strawberry promotes resistance to C. acutatum [41]. Ginkbilobin-2, a secreted MBL from Ginkgo biloba, inhibits the growth of F. oxysporum [42] and several other MBLs exhibit antifungal activity [43–46].
Table 3. Mannose-binding lectin superfamily (MBL) genes regulated >2 fold by F. oxysporum.
| TAIR10 | F1/M1 | F6/M6 | ||
|---|---|---|---|---|
| Roots | Leaves | Roots | Leaves | |
| AT1G78820 | - | - | 0.4 | 0.6 |
| AT3G51710 | - | - | 0.7 | 0.5 |
| AT5G49870 | - | - | 2.0 | - |
| AT5G38550 | - | - | 2.2 | - |
| AT5G38540 | - | - | 2.5 | - |
| AT2G25980 | - | - | 2.6 | - |
| AT1G52050 | 0.6 | - | 2.7 | - |
| AT1G52000 | - | - | 2.8 | 5.6 |
| AT1G52060 | 0.6 | - | 3.1 | - |
| AT1G52130 | - | - | 4.0 | - |
| AT5G28520 | 8.8 | - | 4.5 | - |
| AT1G52070 | 0.6 | - | 5.5 | - |
| AT5G35940 | - | - | 56.4 | - |
| AT1G52100 | - | - | 56.5 | 1.8 |
| AT1G52000 | - | - | 2.8 | 5.6 |
| AT3G16450 | 1.5 | - | 1.8 | 2.4 |
Oxidative burst
One of the earliest responses of roots to F. oxysporum infection is the oxidative burst [47]. 'Response to oxidative stress’ was a highly significant functional GO category of induced genes in the roots at 6 dpi (Table C in S1 File). Reactive oxygen species (ROS) act as signalling molecules to activate plant defences and are mainly generated by NADPH oxidases and apoplastic peroxidases [48]. Previous studies show that the leaf NADPHs RBOHD and RBOHF, that were induced in both tissues in this study (induced 1.7–4.4 fold at 6 dpi) have opposite effects on F. oxysporum disease development [20].
Numerous peroxidases were strongly induced by F. oxysporum in the roots (Fig 4A). PRX53 is JA-responsive and promotes resistance to the cyst nematode H. schachtii [49] while PRX33 is required for ROS formation in response to F. oxysporum filtrate and is a major contributor of MAMP-triggered ROS production [50, 51]. We inoculated prx33-1 and prx33-2, which contain T-DNA insertions in the first intron of PRX33 with F. oxysporum and found that they were more resistant to F. oxysporum than WT plants suggesting that PRX33 promotes susceptibility to F. oxysporum (Fig 4B and 4C). Interestingly, A. thaliana antisense or T-DNA insertion lines with reduced levels of PRX33 and PRX34 are more susceptible to various bacterial and fungal pathogens [50, 52] and show defective SA signaling and impaired PTI responses [53]. Numerous studies associate ROS production with defence, however recent data suggest that ROS may also promote disease development of necrotrophic pathogens. For example, Medicago truncatula plants with reduced ROS-producing capabilities in the roots show enhanced resistance to the root rot pathogen Aphanomyces euteiches [54]; transgenic potato plants that produce increased levels of pathogen-inducible ROS are more susceptible to Alternaria solani [55] while RBOHB-generated ROS promotes Botrytis cinerea lesion development in Nicotiana benthamiana [56]. In addition to basal defence signaling and modulation of defence responses such as the hypersensitive response and cell wall cross linking, ROS affect diverse physiological processes including regulation of lateral root emergence [57] and leaf senescence [58]. Indeed, prx33/prx34 knockdown plants show delayed senescence [59]. A possible explanation for PRX33-mediated susceptibility to F. oxysporum is that that PRX33- generated ROS in the roots acts as a systemic signal to accelerate senescence in the leaves, promoting the transition from a biotrophic to necrotrophic lifestyle of F. oxysporum.
Fig 4. PRX33 is induced by F. oxysporum and promotes susceptibility to F. oxysporum.

(A) The fold induction or repression of peroxidases in response to F. oxysporum. (B) Representative F. oxysporum- inoculated WT and prx33 mutant plants at 14 days post inoculation. (C) Mean disease score and standard error from 16 plants. Asterisk indicates significant difference relative to WT (P<0.01).
Photosynthesis
Interestingly, although photosynthetic genes are generally expressed at low levels in the roots of healthy plants, ‘photosynthesis’ was the most highly enriched GO category in genes induced specifically in the roots at 1 dpi (Table C in S1 File). Eighteen genes associated with photosynthesis were induced 2–3 fold in the roots at 1 dpi (Table F in S1 File). Intriguingly, 16 of these genes were repressed in the leaves at 6 dpi. Similarly to the transcriptional patterns of photosynthesis genes in our study, cotton plants exposed to drought stress show an increased transcription of photosynthesis related genes in the roots and a repression of such genes in the leaves [60] suggesting cross talk between the drought and F. oxysporum responses.
Host processes responsive to F. oxysporum infection in leaves
The most strongly regulated genes in the leaves are listed in Tables G and H in S1 File. The ethylene response factor RAP2.6 (AT1G43160, induced 11 fold in leaves at 1 dpi and in both tissues at 6 dpi) contributes to resistance against the beet cyst nematode Heterodera schachtii [61] while the glutaredoxin ROXY-2 (AT5G14070; repressed 7 fold in leaves at 6 dpi) promotes susceptibility to Botrytis cinerea [62].
Defense / antimicrobial genes
JA-associated defence genes, such as PDF1.2C and PDF1.3 (induced >150 fold 6 dpi) showed strong leaf-specific regulation at 6 dpi (Table 1). PR1, a classic marker of SA-mediated defences was induced during initial infection (4 fold up 1 dpi leaves) but was then downregulated (5 fold down 6 dpi leaves). Such a transcription pattern is typical of a hemibiotroph, whereupon SA defences elicited during the biotrophic phase are antagonised by JA signalling during the necrotrophic phase. In general, however, relatively few SA signalling genes were regulated by F. oxysporum in our experiment. One exception was SABATH methyltransferase AT3G11480 (induced 18 fold at 6 dpi in leaves), which is required for systemic acquired resistance and basal immune responses [63].
Photosynthesis
Consistent with published data from seedlings [20], photosynthesis was the most overrepresented functional GO category in repressed genes in leaves at 6 dpi (Table C in S1 File), which may be a consequence of pathogen-induced chlorosis and senescence. Sixty-one genes associated with photosynthesis were repressed at 6 dpi, however only 10 of these genes were repressed >2 fold (Table F in S1 File).
Circadian rhythm, cold response, and flowering time
Two of the most strongly regulated leaf genes modulate the circadian clock, cold response and flowering time. FKF1, a flavin-binding kelch repeat F-box protein and PSEUDO-RESPONSE REGULATOR 5 were repressed >8 fold in leaves at 1 dpi [64]. Genes encoding several other cold response regulators including CBFs (CRT/DRE binding factor)/DREB (DRE-binding factor such as CBF1, CBF2, COR72, COR413 were also strongly repressed in the leaves. Genes encoding ICE1 and ICE2 and the JAZ proteins, which mediate cross talk between cold and JA signalling [65] and SOC1 and FLC, which mediate cross talk between cold and flowering pathways [66] were all regulated by F. oxysporum in this study. Altering the transition to flowering in response to biotic stress is hypothesised to be a mechanism by the plant to maintain a balance between defence and development. Pathogen infection often results in accelerated flowering time to ensure reproductive success of the plant during stress [67].
Senescence
Delayed senescence has been associated with increased F. oxysporum resistance. For example, COI1, which promotes JA-triggered leaf senescence [68] is required for susceptibility to F. oxysporum [8] while cpr5/hys1 mutants that show accelerated senescence are more susceptible to F. oxysporum [5]. Whereas leaf tissue becomes necrotic and senescent in late stages of infection, roots do not show obvious signs of necrosis after infection (Fig 1A), prompting us to explore the hypothesis that senescence-associated transcriptional changes occur predominantly in the foliar tissue. At 1 dpi, plant senescence markers SAG29 and SAG12 were strongly induced in the leaves (>11 fold), indicating that senescence-associated responses are an important part of the infection and/or lesion development process. By 6 dpi, however, both genes were repressed >2 fold. One hypothesis to explain the marked change in SAG gene expression throughout the infection timecourse is that upon establishment of infection, F. oxysporum manipulates the host to promote leaf senescence to allow the advancing fungus to enter the necrotrophic infection phase and fully colonize the host. Once foliar infection has been established, the host might inhibit the senescence response in a counter attack to inhibit disease symptoms and fungal accumulation. SAG29 encodes a member of the SWEET sucrose efflux transporter family proteins [69] that are associated with plant defense (reviewed by [70, 71]) while SAG12 encodes a cysteine protease implicated in a range of senescence and cell death responses [72, 73]. Therefore the change in transcription of these genes throughout the timecourse could alternatively represent an early immune response against F. oxysporum that is inhibited by the fungus as infection progresses.
Chlorophyllase (CHL1, AT1G19670), the first enzyme involved in chlorophyll degradation, was induced 13 fold in leaves at 6 dpi. CHL1 promotes the production of ROS and resistance to the necrotrophic fungal pathogen Alternaria brassicicola while it negatively regulates resistance to the necrotrophic bacterial pathogen Erwinia carotovora [74].
Tissue-specific regulation of auxin signalling and transport
Exogenous treatment with auxin does not alter the response of A. thaliana plants to F. oxysporum, however several mutants compromised in auxin signalling and transport show increased resistance to F. oxysporum, suggesting that auxin promotes susceptibility to F. oxysporum [11, 75]. In agar-grown seedlings, a marked inhibition of root elongation and increased lateral root proliferation is evident in F. oxysporum–inoculated seedlings relative to mock inoculated seedlings (Fig 5A). Similarly, roots of soil-grown F. oxysporum-inoculated plants are shorter and appear bushier relative to mock-inoculated plants by 6 dpi (Fig 1A, Fig 5B). Such phenotypes are reminiscent of plants treated with auxin. Given that F. oxysporum enters the plant through lateral root (LR) initials [11, 76], an increased proliferation of LR might aid F. oxysporum infection. Several genes differentially expressed >2 fold in the roots, including CONSTANS-LIKE 3 (COL3), ARF19, IAA14 [77], IAA28 [78] and the chitinase-like protein CTL1 [79] regulate lateral root growth or formation. CALLOSE SYNTHASE 3 (CS3; AT5G13000) inhibits callose accumulation in emerging lateral roots [80]. Strong repression of (>8 fold) of CS3 in the roots at 6 dpi may be a mechanism to increase physical barriers against F. oxysporum entry.
Fig 5. Auxin-related phenotypes and role of ARF2 in the A. thaliana—F. oxysporum interaction.

(A) F. oxysporum inoculation triggers root growth inhibition and lateral root (LR) proliferation in agar-grown Col-0 seedlings. (i) Two week old seedlings were inoculated with water (mock) or F. oxysporum and photographed at 9 days post inoculation. (ii) Mean primary root (PR) length or (iii) mean number of LRs per cm PR in mock or F. oxysporum—inoculated agar-grown seedlings measured at 9 dpi. (B) F. oxysporum inoculation triggers root growth inhibition in soil-grown Col-0 plants. Mean PR length in mock (grey bars) or F. oxysporum (black bars)–inoculated soil-grown plants at 1, 6 and 14 dpi. Data shown are mean and standard error from >13 plants. Asterisk indicates significant difference between mock and F. oxysporum treatment (P<0.05). (C) Representative F. oxysporum- inoculated WT (Col-0) and mutant plants at 14 days post inoculation (dpi). (D) Mean percentage of diseased leaves per plant and standard error from at least 30 plants per line. Asterisks indicate significant difference relative to WT (*P<0.05; **P<0.01).
We identified genes involved in auxin signalling or transport that were induced or repressed >2 fold (Table 2). Loss of function mutants of three auxin transport regulators that are F. oxysporum-responsive: AUX1 (both tissues), PIN2 (roots) and TRANSPARENT TESTA 4 (leaves), have previously been shown to contribute to the host response to F. oxysporum [11].
Auxin-associated F. oxysporum-responsive genes in leaves and roots
POLARIS, which was the most strongly repressed gene in the leaves at 6 dpi (Table H in S1 File) encodes a peptide that is auxin-inducible and promotes ethylene-mediated auxin flux and biosynthesis [81]. SMALL AUXIN UPREGULATED RNAs (SAURs) are members of a large gene family containing early-responsive auxin genes of largely unknown function [82]. Thirty SAURs or SAUR-like genes were regulated >2 fold by F. oxysporum and the majority of them were repressed in the leaves at one or both timepoints (Table I in S1 File).
Four members of the five NAKED PINS IN YUC genes, which have been implicated in lateral organogenesis and the root gravitrophic response [83] were regulated specifically in roots at 6 dpi (Table 2).
AUXIN RESPONSE FACTORs (ARFs) are transcription factors that regulate auxin-responsive gene expression and are repressed by AUX/IAA proteins [84]. The majority of the ARFs and AUX/IAA transcripts that were differentially expressed >2 fold in this study were repressed by F. oxysporum. Moreover, several of these genes were repressed specifically in the roots (Table 2). Transcripts corresponding to ARF1 and the closely related protein ARF2 were repressed (1.8 and 2.5 fold, respectively) specifically in the roots at 6 dpi. To determine whether ARF1 or ARF2 play functional roles in F. oxysporum—A. thaliana interaction, we inoculated the previously described arf2, arf2/arf1 and arf1 mutants [16–18] with F. oxysporum. arf1 showed a slight but significant increase in resistance relative to WT plants while independent arf2 and arf1/arf2 mutants were significantly more resistant than both arf1 and WT plants (Fig 5C and 5D). These data suggest that ARF2 and ARF1 promote susceptibility to F. oxysporum.
While F. oxysporum-triggered transcriptional repression of ARF1 and 2 occurs in the roots, it is unclear whether arf1 and arf2-mediated resistance occurs via the roots. ARF2 has been implicated in the control of lateral root growth [85], which might perturb F. oxysporum entry. In addition to promoting lateral organ formation and inhibiting root elongation, auxin also promotes leaf senescence [86]. arf2 plants exhibit delayed leaf senescence and reduced transcription of senescence-inducing genes including SAG12. arf1 loss-of-function mutants show a normal senescence phenotype, but act additively with arf2 to delay senescence in the arf1 arf2 double mutant [16, 87]. Delayed leaf senescence in the arf2 mutants might delay the switch to necrotrophic growth of F. oxysporum, resulting in reduced disease symptoms.
ARF1 and ARF2 have also been shown to promote susceptibility to the biotrophic oomycete Hyaloperonospora arabidopsidis [88] and necrotrophic fungus Sclerotinia sclerotiorum [89], respectively. ARF1 negatively regulates the accumulation of glucosinolates [88] while ARF2 seems to mediate ABA signalling [89]. Altered ABA, SA or JA defence signalling due to perturbed crosstalk with auxin signalling in these mutants may also contribute to the enhanced F. oxysporum resistance phenotype.
Conclusion
Roots and leaves play different biological roles but are dependent on each other for growth and responses to stress. In this article, we presented a comprehensive analysis of the transcriptional response of A. thaliana above-ground and below-ground tissue to F. oxysporum infection. As examples of the functional significance of our transcriptome analyses, we demonstrated that ARF2 and PRX33, two genes differentially expressed in the roots, both promote susceptibility to F. oxysporum. Interestingly, both genes also promote leaf senescence, prompting us to suggest that ARF2 or PRX33 might propagate root-to-shoot signals that accelerate the transition from biotrophic to necrotrophic growth and thus symptom development of F. oxysporum. Genetic modification of PRX33 or ARF2 in crops may increase resistance to F. oxysporum in the field. However, potential benefits of such a strategy need to be considered alongside possible pleiotrophic effects such as altered flowering time, which could lead to yield penalties. Our findings demonstrate the existence of infection stage and tissue-specific transcriptional responses and enhance our understanding of the interaction between plants and hemibiotrophic root-infecting fungal pathogens. Future studies will focus on the functional dissection of genes that are differentially regulated by F. oxysporum in the roots and have not previously been shown to play a role in defence.
Supporting Information
The mean fold induction or repression of genes that were regulated in (A) both timepoints (middle circles, Fig 2B) or in (B) both tissues (each middle circle, Fig 2D) was compared using pairwise comparisons. Data shown are mean fold change and standard error observations. Asterisk indicates significant difference (P<0.05) in a paired T-test.
(TIF)
Proportion of genes showing tissue-specific induction at one timepoint (indicated above Venn diagram) that show tissue-specific regulation throughout the time-course. The number of DEGs that are only ever regulated by F. oxysporum in the designated tissue are shown in parenthesis.
(TIF)
Table A, Novel F. oxysporum-regulated genes identified in 2 week old seedlings [16] and their regulation in this study. Table B, Genes regulated by F. oxysporum in the roots of A. thaliana soil-grown plants at 2 dpi [17] and their regulation in this study. Table C, Results of gene ontology (GO) singular enrichment analysis (SEA comparing the top five most highly overrepresented functional gene ontology categories between tissues for each timepoint. Table D, The ten most strongly induced or repressed genes in the roots at 1 dpi. Highlighted values indicate a fold change >2. Table E, The ten most strongly induced or repressed genes in the roots at 6 dpi. Highlighted values indicate a fold change >2. Table F, Photosynthesis associated genes differentially regulated by F. oxysporum. Highlighted values indicate a fold change >2. Table G, The ten most strongly induced or repressed genes in the leaves at 1dpi. Highlighted values indicate a fold change >2. Table H, The ten most strongly induced or repressed genes in the leaves at 6dpi. Highlighted values indicate a fold change >2. Table I, SMALL AUXIN UPREGULATED (SAUR) and SAUR-like genes regulated > 2 fold by F. oxysporum. Highlighted values indicate a fold change >2.
(XLSX)
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was partially funded by the CSIRO OCE (Office of the Chief Executive) postdoctoral fellowship to RL. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. De Coninck B, Timmermans P, Vos C, Cammue BPA, Kazan K. What lies beneath: belowground defense strategies in plants, Trends in Plant Science. 2014;20: 91–101. 10.1016/j.tplants.2014.09.007 [DOI] [PubMed] [Google Scholar]
- 2. Balmer D, Mauch-Mani B. More beneath the surface? Root versus shoot antifungal plant defenses. Front Plant Sci. 2013;4: 256 10.3389/fpls.2013.00256 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Jones JD, Dangl JL. The Plant immune system. Nature. 2006;16: 323–9. [DOI] [PubMed] [Google Scholar]
- 4. Dong X, Xiong Y, Ling N, Shen Q, Guo S. Fusaric acid accelerates the senescence of leaf in banana when infected by Fusarium. World J Microbiol Biotechnol. 2014;30:1399–408. 10.1007/s11274-013-1564-1 [DOI] [PubMed] [Google Scholar]
- 5. Schenk PM, Kazan K, Rusu AG, Manners JM, Maclean DJ. The SEN1 gene of Arabidopsis is regulated by signals that link plant defence responses and senescence. Plant Physiol Biochem. 2005;43: 997–1005. [DOI] [PubMed] [Google Scholar]
- 6. Thatcher LF, Gardiner DM, Kazan K, Manners JM. A highly conserved effector in Fusarium oxysporum is required for full virulence on Arabidopsis. Mol Plant Microbe Interact. 2012;25: 180–90. 10.1094/MPMI-08-11-0212 [DOI] [PubMed] [Google Scholar]
- 7. Cole SJ, Yoon AJ, Faull KF, Diener AC. Host perception of jasmonates promotes infection by Fusarium oxysporum formae speciales that produce isoleucine- and leucine-conjugated jasmonates. Mol Plant Pathol. 2014;15: 589–600. 10.1111/mpp.12117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Thatcher LF, Manners JM, Kazan K. Fusarium oxysporum hijacks COI1-mediated jasmonate signaling to promote disease development in Arabidopsis. Plant J. 2009;58: 927–939. 10.1111/j.1365-313X.2009.03831.x [DOI] [PubMed] [Google Scholar]
- 9. Thatcher LF, Powell JJ, Aitken EA, Kazan K, Manners JM. The lateral organ boundaries domain transcription factor LBD20 functions in Fusarium wilt susceptibility and jasmonate signaling in Arabidopsis. Plant Physiol. 2012;160: 407–18. 10.1104/pp.112.199067 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Pantelides IS, Tjamos SE, Pappa S, Kargakis M, Paplomatas EJ. The ethylene receptor ETR1 is required for Fusarium oxysporum pathogenicity. Plant Pathol. 2013; 62: 1302–1309. [Google Scholar]
- 11. Kidd BN, Kadoo NY, Dombrecht B, Tekeoğlu M, Gardiner DM, Thatcher LF, et al. Auxin signalling and transport promote susceptibility to the root-infecting fungal pathogen Fusarium oxysporum in Arabidopsis. Mol Plant Microbe Interact. 2011;24: 733–748. 10.1094/MPMI-08-10-0194 [DOI] [PubMed] [Google Scholar]
- 12. Anderson JP, Badruzsaufari E, Schenk PM, Manners JM, Desmond OJ, Ehlert C, et al. Antagonistic interaction between abscisic acid and jasmonate-ethylene signalling pathways modulates defense gene expression and disease resistance in Arabidopsis . Plant Cell. 2004;16: 3460–3479 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Edgar CI, McGrath KC, Dombrecht B, Manners JM, Maclean DC, Schenk PM, et al. Salicylic acid mediates resistance to the vascular wilt pathogen Fusarium oxysporum in the model host Arabidopsis thaliana . Aust Plant Pathol. 2006;35: 581–591. [Google Scholar]
- 14. Diener AC, Ausubel FM. Resistance to FUSARIUM OXYSPORUM 1, a dominant Arabidopsis disease-resistance gene, is not race specific. Genetics. 2005;171: 305–321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Trusov Y, Sewelam N, Rookes JE, Kunkel M, Nowak E, Schenk PM, et al. Heterotrimeric G proteins-mediated resistance to necrotrophic pathogens includes mechanisms independent of salicylic acid, jasmonic acid/ethylene and abscisic acid-mediated defense signaling. Plant J. 2009;58: 69–81. 10.1111/j.1365-313X.2008.03755.x [DOI] [PubMed] [Google Scholar]
- 16. Ellis CM, Nagpal P, Young JC, Hagen G, Guilfoyle TJ, Reed JW. AUXIN RESPONSE FACTOR1 and AUXIN RESPONSE FACTOR2 regulates senescence and floral organ abscission in Arabidopsis thaliana . Development. 2005;132: 4563–4574. [DOI] [PubMed] [Google Scholar]
- 17. Okushima Y, Mitina I, Quach HL, Theologis A. AUXIN RESPONSE FACTOR 2 (ARF2): a pleiotropic developmental regulator. Plant J. 2005;43: 29–46 [DOI] [PubMed] [Google Scholar]
- 18. Okushima Y, Overvoorde PJ, Arima K, Alonso JM, Chan A, Chang C, et al. Functional genomic analysis of the AUXIN RESPONSE FACTOR gene family members in Arabidopsis thaliana: unique and overlapping functions of ARF7 and ARF19. Plant Cell. 2005; 17: 444–463 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. McGrath KC, Dombrecht B, Manners JM, Schenk PM, Edgar CI, Maclean DJ, et al. Repressor- and activator-type Ethylene Response Factors functioning in jasmonate signaling and disease resistance identified via a genome-wide screen of Arabidopsis transcription factor gene expression. Plant Physiol. 2005;139: 949–959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Zhu AH, Stephen S, Kazan K, Jin G, Fan L, Taylor J, et al. Characterisation of the defense transcriptome responsive to Fusarium oxysporum infection in Arabidopsis using RNA_seq. Gene 2012;10: 259–66. [DOI] [PubMed] [Google Scholar]
- 21. Trapnell C, Williams BA, Pertea G, Mortazavi AM, Kwan G, van Baren MJ, et al. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nature Biotech. 2010;28: 511–515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Du Z, Zhou X, Ling Y, Zhang Z, Su Z. agriGO: a GO analysis toolkit for the agricultural community. Nucl Acids Res. 2010;38: W64–W70. 10.1093/nar/gkq310 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Lyons R, Iwase A, Gänsewig T, Sherstnev A, Duc C, Barton GJ, et al. The RNA-binding protein FPA regulates flg22-triggered defense responses and transcription factor activity by alternative polyadenylation. 2013;Sci Rep 3: 2866 10.1038/srep02866 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Tytgat TO, Verhoeven KJ, Jansen JJ, Raaijmakers CE, Bakx-Schotman T, McIntyre LM, et al. Plants know where it hurts: root and shoot jasmonic acid induction elicit differential responses in Brassica oleracea . Mol Ecol. 2013;22: 6179–96. 10.1111/mec.12555 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Balmer D, de Papajewski DV, Planchamp C, Glauser G, Mauch-Mani B. Induced resistance in maize is based on organ-specific defence responses. Plant J. 2013;74: 213–225. 10.1111/tpj.12114 [DOI] [PubMed] [Google Scholar]
- 26. Chen YC, Wong CL, Muzzi F, Vlaardingerbroek I, Kidd BN, Schenk PM. Root defense analysis against Fusarium oxysporum reveals new regulators to confer resistance. Sci Rep. 2014; 4: 5584 10.1038/srep05584 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Zhao S, Fung-Leung WP, Bittner A, Ngo K, Liu X.. Comparison of RNA-Seq and microarray in transcriptome profiling of activated T cells. PLoS One. 2014;9: e78644 10.1371/journal.pone.0078644 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Nekrasov V, Li J, Batoux M, Roux M, Chu ZH, Lacombe S, et al. Control of the pattern-recognition receptor EFR by an ER protein complex in plant immunity. EMBO J. 2009;28: 3428–3438. 10.1038/emboj.2009.262 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Reitz MU, Bissue JK, Zocher K, Attard A, Hückelhoven R, Becker K, et al. The subcellular localization of Tubby-like proteins and participation in stress signaling and root colonization by the mutualist Piriformospora indica . Plant Physiol. 2012;160: 349–64. 10.1104/pp.112.201319 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Lev-Yadun S, Gould KS. Role of Anthocyanins in Plant Defence 22–28 In: Gould KS, Davies KM and Winefield C, editors. Life’s colorful solutions: the biosynthesis, functions, and applications of anthocyanins. Springer-Verlag, Berlin: 2009. Pp. 22–28. [Google Scholar]
- 31. Owens DK, Alerding AB, Crosby KC, Bandara AB, Westwood JH, Winkel BS. Functional analysis of a predicted flavonol synthase gene family in Arabidopsis. Plant Physiol. 2008;147: 1046–1061. 10.1104/pp.108.117457 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Kitamura S, Shikazono N, Tanaka A. TRANSPARENT TESTA 19 is involved in the accumulation of both anthocyanins and proanthocyanidins in Arabidopsis. Plant J. 2004;37: 104–14. [DOI] [PubMed] [Google Scholar]
- 33. Wang H, Fan W, Li H, Yang J, Huang J, Zhang P. Functional characterization of dihydroflavonol-4-reductase in anthocyanin biosynthesis of purple sweet potato underlies the direct evidence of anthocyanins function against abiotic stresses. PLoS One. 2013;8: e78484 10.1371/journal.pone.0078484 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Adio AM, Casteel CL, De Vos M, Kim JH, Joshi V, Li B, et al. Biosynthesis and defensive function of Nδ-acetylornithine, a jasmonate-induced Arabidopsis metabolite. Plant Cell. 2011;23: 3303–18. 10.1105/tpc.111.088989 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Kazan K, Lyons R. Intervention of phytohormone pathways by pathogen effectors. Plant Cell. 2014;26: 2285–2309 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Lim CW, Luan S, Lee SC. A prominent role for RCAR3-mediated ABA signaling in response to Pseudomonas syringae pv. tomato DC3000 infection in Arabidopsis. Plant Cell Physiol. 2014;55: 1691–703. 10.1093/pcp/pcu100 [DOI] [PubMed] [Google Scholar]
- 37. Fujita Y, Nakashima K, Yoshida T, Katagiri T, Kidokoro S, Kanamori N, et al. Three SnRK2 protein kinases are the main positive regulators of abscisic acid signaling in response to water stress in Arabidopsis. Plant Cell Physiol. 2009;50: 2123–32. 10.1093/pcp/pcp147 [DOI] [PubMed] [Google Scholar]
- 38. Seo S, Gomi K, Kaku H, Abe H, Seto H, Nakatsu S, et al. Identification of natural diterpenes that inhibit bacterial wilt disease in tobacco, tomato and Arabidopsis. Plant Cell Physiol. 2012;53: 1432–1444. 10.1093/pcp/pcs085 [DOI] [PubMed] [Google Scholar]
- 39. Sun X, Feng P, Xu X, Guo H, Ma J, Chi W, et al. A chloroplast envelope-bound PHD transcription factor mediates chloroplast signals to the nucleus. Nat Commun. 2011;20: 477–486. [DOI] [PubMed] [Google Scholar]
- 40. Barre A, Bourne Y, Van Damme EJM, Peumans WJ, Rougé P. Mannose-binding plant lectins: different structural scaffolds for a common sugar-recognition process. Biochimie. 2001;83: 645–651. [DOI] [PubMed] [Google Scholar]
- 41. Hwang BK, Hwang IS. The pepper mannose-binding lectin gene CaMBL1 is required to regulate cell death and defense responses to microbial pathogens. Plant Physiol. 2011;155: 447–463. 10.1104/pp.110.164848 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Guidarelli M, Zoli L, Orlandini A, Bertolini P, Baraldi E. The mannose-binding lectin gene FaMBL1 is involved in the resistance of unripe strawberry fruits to Colletotrichum acutatum . Mol Plant Pathol. 2014;15: 832–840. 10.1111/mpp.12143 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Miyakawa T, Hatano K, Miyauchi Y, Suwa Y, Sawano Y, Tanokura M. A secreted protein with plant-specific cysteine-rich motif functions as a mannose-binding lectin that exhibits antifungal activity. Plant Physiol. 2014;166: 766–78. 10.1104/pp.114.242636 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Wang XC, Bauw G, Van Damme EJM, Peumans WJ, Chen ZL, Van Montagu M, et al. Gastrodianin-like mannose-binding proteins: a novel class of plant proteins with antifungal properties. Plant J. 2001;25: 651–66. [DOI] [PubMed] [Google Scholar]
- 45. Araujo-Filho JH, Vasconcelos IM, Martins-Miranda AS, Gondim DM, Oliveira JT. A ConA-like lectin from Dioclea guianensis Benth. has antifungal activity against Colletotrichum gloeosporioides, unlike its homologues, ConM and ConA. J. Agric. Food Chem. 2010;58: 4090–4096. 10.1021/jf903254b [DOI] [PubMed] [Google Scholar]
- 46. Cox KD, Layne DR, Scorza R, Schnabel G. Gastrodia anti-fungal protein from the orchid Gastrodia elata confers disease resistance to root pathogens in transgenic tobacco. Planta. 2006;224: 1373–1383. [DOI] [PubMed] [Google Scholar]
- 47. Plancot B, Santaella C, Jaber R, Kiefer-Meyer MC, Follet-Gueye ML, Leprince J, et al. Deciphering the responses of root border-like cells of Arabidopsis and flax to pathogen-derived elicitors. Plant Physiol. 2013;4: 1584–97. 10.1104/pp.113.222356 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Torres MA. ROS in biotic interactions. Physiol Plant. 2010;138: 414–429. 10.1111/j.1399-3054.2009.01326.x [DOI] [PubMed] [Google Scholar]
- 49. Jin J, Hewezi T, Baum TJ. Arabidopsis peroxidase AtPRX53 influences cell elongation and susceptibility to Heterodera schachtii . Plant Signal Behav. 2011;6: 1778–86. 10.4161/psb.6.11.17684 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Daudi A, Cheng Z, O'Brien JA, Mammarella N, Khan S, Ausubel FM, et al. The apoplastic oxidative burst peroxidase in Arabidopsis is a major component of pattern-triggered immunity. Plant Cell. 2012;24: 275–87. 10.1105/tpc.111.093039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. O’Brien JA, Daudi A, Finch P, Butt VS, Whitelegge JP, Souda P, et al. A peroxidase-dependent apoplastic oxidative burst in cultured Arabidopsis cells functions in MAMP-elicited defense. Plant Physiol. 2012;158: 2013–2027. 10.1104/pp.111.190140 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Bindschedler LV, Dewdney J, Blee KA, Stone JM, Asai T, Plotnikov J, et al. Peroxidase-dependent apoplastic oxidative burst in Arabidopsis required for pathogen resistance. Plant J. 2006;47: 851–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Mammarella ND, Cheng Z, Fu ZQ, Daudi A, Bolwell GP, Dong X, et al. Apoplastic peroxidases are required for salicylic acid-mediated defense against Pseudomonas syringae . Phytochem. 2014; 10.1016/j.phytochem.2014.07.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Kiirika LM, Bergmann HF, Schikowsky C, Wimmer D, Korte J, Schmitz U, et al. Silencing of the Rac1GTPase MtROP9 in M. truncatula stimulates early mycorrhizial and oomycete root colonizations but negatively affects rhizobial infection. Plant Physiol. 2012;159: 501–516. 10.1104/pp.112.193706 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Kobayashi M, Yoshioka M, Asai S, Nomura H, Kuchimura K, Mori H, et al. StCDPK5 confers resistance to late blight pathogen but increases susceptibility to early blight pathogen in potato via reactive oxygen species burst. New Phytol. 2012;196: 223–237. 10.1111/j.1469-8137.2012.04226.x [DOI] [PubMed] [Google Scholar]
- 56. Asai S, Yoshioka H. Nitric oxide as a partner of reactive oxygen species participates in disease resistance to necrotrophic pathogen Botrytis cinerea in Nicotiana benthamiana . Mol Plant Microbe Interact. 2009;22: 619–629. 10.1094/MPMI-22-6-0619 [DOI] [PubMed] [Google Scholar]
- 57. Manzano C, Pallero-Baena M, Casimiro I, De Rybel B, Orman-Ligeza B, Van Isterdael G, et al. The emerging role of reactive oxygen species signaling during lateral root development Plant Physiol. 2014;165: 1105–1119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Jing HC, Hebeler R, Oeljeklaus S, Sitek B, Stuhler K, Meyer HE, et al. Early leaf senescence is associated with an altered cellular redox balance in Arabidopsis cpr5/old1 mutants. Plant Biol. 2008;1: 85–98. [DOI] [PubMed] [Google Scholar]
- 59. Passardi F, Tognolli M, De Meyer M, Penel C, Dunand C. Two cell wall associated peroxidases from Arabidopsis influence root elongation. Planta. 2006;223: 965–974. [DOI] [PubMed] [Google Scholar]
- 60. Ranjan A, Sawant S. Genome-wide transcriptomic comparison of cotton (Gossypium herbaceum) leaf and root under drought stress. 3 Biotech. 2014; 10.1007/s13205-014-0257-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Ali MA, Abbas A, Kreil DP, Bohlmann H. Overexpression of the transcription factor RAP2.6 leads to enhanced callose deposition in syncytia and enhanced resistance against the beet cyst nematode Heterodera schachtii in Arabidopsis roots. BMC Plant Biol. 2013;13: 47 10.1186/1471-2229-13-47 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Wang Z, Xing S, Birkenbihl RP, Zachgo S. Conserved functions of Arabidopsis and rice CC-type glutaredoxins in flower development and pathogen response. Mol Plant. 2009;2: 323–35. 10.1093/mp/ssn078 [DOI] [PubMed] [Google Scholar]
- 63. Liu PP, Yang Y, Pichersky E, Klessig DF. Altering expression of Benzoic Acid/Salicylic Acid Carboxyl Methyltransferase 1 compromises systemic acquired resistance and PAMP-triggered immunity in Arabidopsis. Mol Plant Microbe Interact. 2010;23: 82–90. 10.1094/MPMI-23-1-0082 [DOI] [PubMed] [Google Scholar]
- 64. Baudry A, Ito S, Song YH, Strait AA, Kiba T, Lu S, et al. F-box proteins FKF1 and LKP2 act in concert with ZEITLUPE to control Arabidopsis clock progression. Plant Cell. 2010;22: 606–622 10.1105/tpc.109.072843 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Hu Y, Jiang L, Wang F, Yu D. Jasmonate regulates the inducer of CBF EXPRESSION-C-REPEAT BINDING FACTOR/DRE BINDING FACTOR 1 CASCADE and freezing tolerance in Arabidopsis. Plant Cell. 2013;25: 2907–2924. 10.1105/tpc.113.112631 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Seo E, Lee H, Jeon J, Park H, Kim J, Noh YS, et al. Crosstalk between cold response and flowering in Arabidopsis is mediated through the flowering-time gene SOC1 and its upstream negative regulator FLC . Plant Cell. 2009;21: 3185–3197. 10.1105/tpc.108.063883 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Korves TM, Bergelson J. A developmental response to pathogen infection in Arabidopsis. Plant Physiol. 2003;133: 339–347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Lee SH, Sakuraba Y, Lee T, Kim KW, An G, Lee HY, et al. Mutation of Oryza sativa CORONATINE INSENSITIVE 1b (OsCOI1b) delays leaf senescence. J Integr Plant Biol. 2014; 10.1111/jipb.12276 [DOI] [PubMed] [Google Scholar]
- 69. Chen L, Qu XQ, Hou BH, Sosso D, Osorio S, Fernie AR, et al. Sucrose efflux mediated by SWEET proteins as a key step for phloem transport. Science. 2012;335: 207–211. 10.1126/science.1213351 [DOI] [PubMed] [Google Scholar]
- 70. Tauzin AS, Giardina T. Sucrose and invertases, a part of the plant defense response to the biotic stresses. Front Plant Sci. 2014;23: 293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Morkunas I, Ratajczak L. The role of sugar signaling in plant defense responses against fungal pathogens, Acta Physiol Plant. 2014;36: 1607–1619. [Google Scholar]
- 72. Lohman KN, Gan S, John MC, Amasino RM. Molecular analysis of natural leaf senescence in Arabidopsis thaliana . Physiol Plant. 1994;92: 322–328. [Google Scholar]
- 73. Zhang K, Halitschke R, Yin C, Liu CJ, Gan SS. Salicylic acid 3-hydroxylase regulates Arabidopsis leaf longevity by mediating salicylic acid catabolism. PNAS. 2013;3: 14807–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Kariola T, Brader G, Li J, Palva ET. Chlorophyllase 1, a damage control enzyme, affects the balance between defense pathways in plants. Plant Cell. 2004;17: 282–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Kazan K, Manners JM. Linking development to defense: auxin in plant-pathogen interactions. Trends Plant Sci. 2009;14: 373–82. 10.1016/j.tplants.2009.04.005 [DOI] [PubMed] [Google Scholar]
- 76. Czymmek KJ, Fogg M, Powell DH, Sweigard J, Park SY, Kang S. In vivo time-lapse documentation using confocal and multi-photon microscopy reveals the mechanisms of invasion into the Arabidopsis root vascular system by Fusarium oxysporum . Fungal Genet Biol. 2007;44:1011–1023. [DOI] [PubMed] [Google Scholar]
- 77. Okushima Y, Fukaki H, Onoda M, Theologis A, Tasaka M. ARF7 and ARF19 regulate lateral root formation via direct activation of LBD/ASL genes in Arabidopsis . Plant Cell. 2007;19: 118–130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Rogg LE, Lasswell J, Bartel B. A gain-of-function mutation in iaa28 suppresses lateral root development. Plant Cell. 2001;13: 465–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Hermans C, Porco S, Verbruggen N, Bush DR. Chitinase-like protein CTL1 plays a role in altering root system architecture in response to multiple environmental conditions. Plant Physiol. 2010; 152: 904–17. 10.1104/pp.109.149849 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Vatén A, Dettmer J, Wu S, Stierhof YD, Miyashima S, Yadav SR, et al. Callose biosynthesis regulates symplastic trafficking during root development. Dev Cell. 2011;21: 1144–1155. 10.1016/j.devcel.2011.10.006 [DOI] [PubMed] [Google Scholar]
- 81. Chilley PM, Casson SA, Tarkowski P, Hawkins N, Wang KL, Hussey PJ, et al. The POLARIS peptide of Arabidopsis regulates auxin transport and root growth via effects on ethylene signaling. Plant Cell. 2006;18: 3058–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Hagen G, Guilfoyle T. Auxin-responsive gene expression: genes, promoters, and regulatory factors. Plant Mol Biol. 2002;49: 373–385. [PubMed] [Google Scholar]
- 83. Furutani M, Sakamoto N, Yoshida S, Kajiwara T, Robert HS, Friml J, et al. Polar-localized NPH3-like proteins regulate polarity and endocytosis of PIN-FORMED auxin efflux carriers. Development. 2011;138: 2069–78. 10.1242/dev.057745 [DOI] [PubMed] [Google Scholar]
- 84. Tiwari SB, Wang XJ, Hagen G, Guilfoyle TJ. AUX/IAA proteins are active repressors, and their stability and activity are modulated by auxin. Plant Cell. 2001;13: 2809–2822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85. Marin E, Jouannet V, Herz A, Lokerse AS, Weijers D, Vaucheret H, et al. miR390, Arabidopsis TAS3 tasiRNAs, and their AUXIN RESPONSE FACTOR targets define an autoregulatory network quantitatively regulating lateral root growth. Plant Cell. 2010;22: 1104–1117. 10.1105/tpc.109.072553 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Mueller-Roeber B, Balazadeh S. Auxin and its role in plant senescence. J Plant Growth Regul. 2014; 10.1007/s00344-013-9398-5 [DOI] [Google Scholar]
- 87. Lim PO, Lee IC, Kim J, Kim HJ, Ryu JS, Woo HR, et al. Auxin response factor 2 (ARF2) plays a major role in regulating auxin-mediated leaf longevity. J Exp Bot. 2010;61: 1419–1430. 10.1093/jxb/erq010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88. Robert-Seilaniantz A, MacLean D, Jikumaru Y, Hill L, Yamaguchi S, Kamiya Y, et al. The microRNA miR393 re-directs secondary metabolite biosynthesis away from camalexin and towards glucosinolates. Plant J. 2011;67: 218–31. 10.1111/j.1365-313X.2011.04591.x [DOI] [PubMed] [Google Scholar]
- 89. Stotz HU, Jikumaru Y, Shimada Y, Sasaki E, Stingl N, Mueller MJ, et al. Jasmonate-dependent and COI1-independent defense responses against Sclerotinia sclerotiorum in Arabidopsis thaliana: auxin is part of COI1-independent defense signaling. Plant Cell Physiol. 2011;52: 1941–56. 10.1093/pcp/pcr127 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The mean fold induction or repression of genes that were regulated in (A) both timepoints (middle circles, Fig 2B) or in (B) both tissues (each middle circle, Fig 2D) was compared using pairwise comparisons. Data shown are mean fold change and standard error observations. Asterisk indicates significant difference (P<0.05) in a paired T-test.
(TIF)
Proportion of genes showing tissue-specific induction at one timepoint (indicated above Venn diagram) that show tissue-specific regulation throughout the time-course. The number of DEGs that are only ever regulated by F. oxysporum in the designated tissue are shown in parenthesis.
(TIF)
Table A, Novel F. oxysporum-regulated genes identified in 2 week old seedlings [16] and their regulation in this study. Table B, Genes regulated by F. oxysporum in the roots of A. thaliana soil-grown plants at 2 dpi [17] and their regulation in this study. Table C, Results of gene ontology (GO) singular enrichment analysis (SEA comparing the top five most highly overrepresented functional gene ontology categories between tissues for each timepoint. Table D, The ten most strongly induced or repressed genes in the roots at 1 dpi. Highlighted values indicate a fold change >2. Table E, The ten most strongly induced or repressed genes in the roots at 6 dpi. Highlighted values indicate a fold change >2. Table F, Photosynthesis associated genes differentially regulated by F. oxysporum. Highlighted values indicate a fold change >2. Table G, The ten most strongly induced or repressed genes in the leaves at 1dpi. Highlighted values indicate a fold change >2. Table H, The ten most strongly induced or repressed genes in the leaves at 6dpi. Highlighted values indicate a fold change >2. Table I, SMALL AUXIN UPREGULATED (SAUR) and SAUR-like genes regulated > 2 fold by F. oxysporum. Highlighted values indicate a fold change >2.
(XLSX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.