

Abstract
Recombination, together with mutation, generates the raw material of evolution, is essential for reproduction and lies at the heart of all genetic analysis. Recent advances in our ability to construct genome-scale, high-resolution recombination maps and new molecular techniques for analysing recombination products have substantially furthered our understanding of this important biological phenomenon in humans and mice: from describing the properties of recombination hot spots in male and female meiosis to the recombination landscape along chromosomes. This progress has been accompanied by the identification of trans-acting systems that regulate the location and relative activity of individual hot spots.
In eukaryotic organisms in which recombination has been studied at sufficiently high resolution and with large enough numbers of progeny, recombination events are not placed randomly along chromosomal DNA but instead are concentrated in highly localized areas of elevated recombination activity. These so-called hot spots are typically 1–2 kb long, are surrounded by much longer regions that are essentially devoid of recombination and largely define the genomic patterns of inheritance in each generation. More recently, studies of hot spot regulation have emerged as models for studying the role of chromatin structure in modulating DNA function. Therefore an understanding of hot spot biology has implications for mechanistic studies of population genetics, evolutionary processes and DNA function.
In this Review we discuss several technical advances, including computational analyses of human population-genetic data, sperm analyses and large-scale crosses in mice, that are propelling current research. We review recent findings on the organization of recombination along chromosomes and on the characteristics of hot spots and their recombination products. This extends to the regulation of crossing over, both on a genome-wide scale and at the level of individual hot spots, including the identification of a new trans-acting factor that has implicated chromatin modification in recombination activation. Finally, we consider several evolutionary aspects of hot spot biology. Throughout we attempt to frame important open questions as well as describe the key experimental findings.
We focus on the biology of hot spots in mammals, as this area has seen important advances in recent years.
We relate these findings to relevant information from other species, especially Saccharomyces cerevisiae and Schizosaccharomyces pombe, in which so many details of the mechanisms of DNA exchange were first discovered. For additional views on the subject, the reader is referred to several recent reviews1-5.
Methodologies
Historical overview
The first mammalian hot spot was discovered in 1982 (REF. 6) in the H2 region of mouse chromosome 17 (Chr 17). This finding emerged from one of the first efforts to relate genetic data to the molecular structure of a chromosome region, in this case a 200-kb segment that was reconstructed by isolating overlapping cosmid clones. The first human hot spots were discovered a few years later in the β-globin7 and insulin regions8 in the course of analysing familial inheritance patterns. The ensuing 25 years saw the number of identified mammalian hot spots increase incrementally, often by chance. The list of recognized hot spots now includes Ath1 (also known as Tnfsf4)9, Scnm1 (REF. 10) and the HS22 region11 in mice, at least four hot spots in the H2 region12, and several human hot spots in the human leukocyte antigen (HLA) region13-20.
The recent introduction of single-nucleotide polymorphism (SNP) genotyping has markedly increased the number of known hot spots and therefore the repertoire of material available for study. Additionally, the field has moved beyond the study of individual hot spots to their distribution across mammalian genomes. These advances have largely relied on several technical developments that are described in the following subsections: statistical analyses of genetic variation data from human populations; large-scale analyses of pedigrees in humans and mice; and molecular analyses of recombination hot spots in sperm samples from both species.
Statistical analyses of genetic variation data
Human population studies have relied partly on the observation that adjacent SNPs tend to form clusters, or haplotype blocks, that are 10–100 kb long. Within these blocks most or all markers are in linkage disequilibrium (LD), and the block boundaries are correlated among populations21. Although the patterns of LD can be shaped by many factors — including mutation, recombination, selection, population demography and genetic drift — computational analyses of LD data with coalescent-based statistical methods have been used to infer probabilities that haplotype boundaries represent historical hot spots. These studies have greatly advanced our knowledge about the genome-wide distribution of recombination events and about the potential cis-acting factors that regulate hot spot activity in humans22-25 (FIG. 1a). This correspondence has been validated by sperm-typing studies (see later section): hot spots in the HLA region identified by sperm typing were also detected by population studies22,23 (FIG. 1 b) and, conversely, the existence of LD-predicted hot spots in humans has been confirmed by sperm typing26.
Figure 1. Hot spot positioning and intensity.
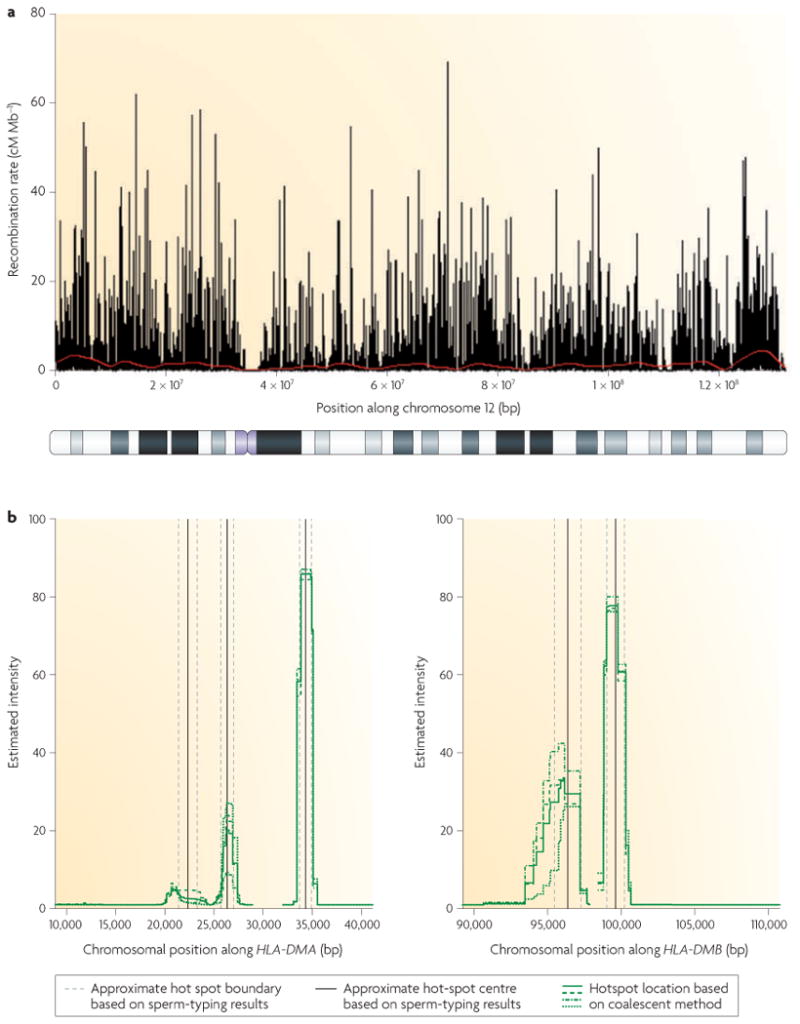
a | Recombination rate variation along human chromosome 12. The graph shows the recombination rate estimated from a genome-wide survey of genetic variation (black bars) and estimated recombination rates from the deCODE genetic map (red line). Recombination hot spots are evident as black spikes surrounded by regions of low or no recombination. Underneath the graph is a cytological banding map of the chromosome. Note the lack of recombination activity near the centromere (purple), b | Estimated locations and intensities of several hot spots in the human major histocompatibility complex (MHC) region inferred by coalescent-based statistical methods (left panel, human leukocyte antigen (HLA)-DMA; right panel, HLA-DMB). Positions along the x-axis are in base pairs according to the consensus map of the human MHC13 The green lines show estimates from the coalescent method22, and the vertical black lines and dashed grey lines show the approximate hot spot centre and the approximate hot spot boundaries, respectively, as estimated from sperm-typing results13. Part a is modified, with permission, from REF. 24 © (2005) American Association for the Advancement of Science. Part b is reproduced, with permission, from Nature Genetics REF. 22
However, several observations suggest that the edges of haplotype blocks are not uniformly coincident with hot spots. The recombination maps of human chromosomes in LD units per Mb pair (which reflects historical recombination) and centimorgans (cM) per Mb pair (which reflects one-generation recombination) are not entirely concordant27. A high-resolution genome-wide association study of recombination in 728 human meioses found that only 60% of detected recombination events coincided with hot spots inferred from LD analysis28. Specific exceptions have also been reported at the level of individual hot spots29.
Statistical analyses based on coalescent approaches are powerful in providing broad-scale, high-resolution recombination maps (FIG. 1a). However, it is important to recognize that they are limited in that they describe sex-averaged, historical recombination among genetically heterogeneous populations.
Large-scale analyses of pedigrees in humans and mice
A more direct approach for studying hot spots that are currently active involves high-resolution mapping of recombination events among the progeny of pedigreed families in humans28,30,31 or of crosses between inbred strains of mice32. For studies of hot spot regulation, animal models provide the advantages of quantitative precision through the use of large-scale crosses between genetically identical parents, the ability to distinguish between male and female recombination and the ability to detect effects of genomic imprinting by comparing recombination in F1 hybrids derived from reciprocal matings. FIG. 2 depicts male and female recombination patterns in a cross between two inbred mouse strains32.
Figure 2. High-resolution sex-specific maps of a mouse chromosome segment.

The plot shows the recombination map of a 27-Mb telomeric region of mouse chromosome 1 in a cross between C57BL/6J and CAST/EiJ strains, highlighting the difference in the recombination rate between the two sexes. The F1 hybrids were either female or male and derived from reciprocal parental combinations (a total of 6,000 progeny). The width of each vertical line indicates the resolution attained. Female recombination rates are in red and male recombination rates are in blue. The arrows show sex-specific hot spots.
The drawback of all large-scale, high-resolution efforts is their cost; collecting and genotyping thousands of offspring, even in the case of mice, is expensive.
Molecular analyses of recombination hot spots using sperm analyses
Studies of human hot spots have focused on the use of sperm as representatives of a large number of individual progeny from individual males; this overcomes the difficulty of obtaining and genotyping large human families and the effect that heterogeneous genetic backgrounds have on the positioning of recombination. In mice, sperm studies have made it possible to characterize the molecular structure of recombination products at individual hot spots and assay for their presence or absence among members of genetically segregating populations of animals.
Sperm assays rely on PCR amplification of DNA from either single sperm or pooled sperm. In the single-sperm assay33,34, individual sperm are subjected to a round of whole-genome amplification to permit multiple loci to be typed from the same sperm. Because of the difficulties of carrying out large numbers of such reactions, single-sperm genotyping is most useful for constructing genetic maps (by detecting reasonably high frequency crossovers (COs) between distant genetic markers) but is less practical for studying recombination events within individual hot spots35.
Pooled-sperm DNA genotyping was developed to overcome the need to examine large numbers of individual sperm and to allow detailed mapping within a hot spot of both CO and non-crossover (NCO) recombination events — the two alternative outcomes of repairing the double-strand break (DSB) that initiates recombination14-16,36,37. To examine the molecular details of individual recombination events, samples are diluted so that aliquots rarely contain two recombinant sperm. To determine recombination rates, concentrations are adjusted so that there is approximately one recombinant sperm per aliquot, and a quantitative estimate of their numbers is obtained based on the fraction of positive samples. Sperm typing has been used to characterize more than thirty hot spots, including TAP2 (REF. 19), the major histocompatibility complex (MHC) region13 and the pseudoautosomal pairing region38 in humans and Psmb9 in mice37,39.
A further development of pooled-sperm genotyping40 allows for unequivocal distinction of single CO and NCO events and a direct estimate of their frequency and molecular structure. The entire hot spot sequence is amplified with primers that are common for all recombinant and non-recombinant products, and the products are cloned into Escherichia coli. Each bacterial colony, which derives from the DNA of a single sperm, is then genotyped.
Pooled-sperm genotyping has the drawback that it is unable to create large-scale recombination maps and sometimes presents difficulties in developing allele-specific amplification based on a single-base-pair difference. These difficulties may be resolved with the advent of high-throughput DNA sequencing41.
Next, we describe the results of the approaches above for characterizing hot spots. These results have been almost entirely consistent between human and mouse studies and provide the context for examining the regulation of hot spot activity, the large-scale organization of recombination along chromosomes and the evolutionary aspects of mammalian recombination.
Properties of hot spots
Recombination at a hot spot (BOX 1 begins with a DSB, which is eventually repaired and resolved as an NCO gene conversion or, less frequently, as a CO that results in the exchange of flanking chromosomal sequences. Despite this extensive DNA repair, recombination is not overtly mutagenic; fine mapping of recombination products has shown that the exchange points of COs within a mammalian hot spot are precise to the base pair. Regulation can be achieved at either the selection of initiation sites or the choice between pathways leading to one of the two products above.
Box 1. Recombination pathways.
Meiosis begins (see figure) with a round of DNA replication (not shown) in which the products stay attached through their centromeres to form pairs of chromatids. Recombination then begins at a hot spot site when the products of trans-acting genes, such as PR domain-containing 9 (Prdm9), locally activate chromatin, permitting the topoisomerase sporulation-specific 11 (SPO11) to catalyse a DNA double-strand break (DSB) on one of the four chromatids. This is followed by resection of the 5′ strand to leave a 3′ overhang, which in turn invades a non-sister chromatid. The resulting strand overlaps to form so-called Holliday junctions, which then migrate outwards away from the original site. This interaction promotes pairing of the non-sister chromatids along their length to form the synaptonemal complex (SC), a linear proteinaceous structure to which the DNA is bound in long loops (BOX 3). The DSBs are subsequently repaired by the process of homologous recombination, yielding either crossovers (COs), with an exchange of flanking markers, or non-crossover (NCO) gene conversions in which the initiating chromatid acquires a short sequence from its homologous partner without the exchange of flanking markers. In either case the site of the original DSB is repaired using the opposite chromatid as a template; when SNPs are available in the middle of the hot spot this fact can be used to determine which chromatid initiated recombination.
Current evidence suggests that the alternative CO and NCO products arise by distinct recombination pathways: DSB repair (DSBR), which yields predominantly COs, and synthesis-dependent strand annealing (SDSA), which yields predominantly NCOs119. Genetic hot spots are the initiation sites for both pathways, as the formation of both CO and NCO products depends on hot spot activity. The SDSA pathway predominates, and only about 10% of original DSBs result in COs.
The crucial controls of recombination activity are the frequency of initiation and the choice of which pathway is followed. Figure modified, with permission, from REF. 119
As described below, we now have a clearer picture of the molecular products of recombination and the sensitivity of hot spots to several influences, including parental imprinting, sequence variation and sex.
Hot spot anatomy
Studies in dense SNP regions show that exchange sites are spread over 500–2,000 bp and form one or two partially overlapping normal distributions around a central point, reflecting the outward migration of Holliday junctions (FIG. 3). By contrast, most NCO gene conversions that have been analysed with sufficient resolution are localized in the central zone and typically involve less than 300 bp, and sometimes as little as tens of base pairs (that is, only a fraction of the total hot spot)14,15,32,36,40. The implication is that although all COs can be detected genetically, many NCOs are not detectable due to a lack of appropriately located SNPs, making it difficult to compare the relative amounts CO and NCO products between hot spots.
Figure 3. Distribution of crossover exchange points in hot spots.

a | Sperm crossover (CO) distribution across the NID1 hot spot. The COs were recovered from a total of 980,000 amplifiable molecules in the sperm DNA of two men. The intervals are defined by SNP markers, and the number of COs is shown above each interval. Exchange sites of crossing over form a single normal distribution, b | Distribution of COs at the Hlx1 hot spot in 6,000 progeny of female and male B6×CAST F1 mouse hybrids. The number of COs in female (red) and male (blue) meiosis is shown above each interval. Exchange sites of crossing over form two overlapping distributions. Part a is modified, with permission, from Nature Genetics REF. 18 © (2005) Macmillan Publishers Ltd. All rights reserved. Part b is modified from REF 32.
Relative rates of non-crossover gene conversions
One of the uncertainties of recombination biology is the fate of the 90% of DSBs that do not become COs; presumably they become NCO conversions, but this has been difficult to prove owing to the considerable variation (from >12:1 to 0.25:1) in the apparent relative frequencies of COs and NCO conversions at human and mouse hot spots14,32,39,42,43. As noted above, this variation probably reflects the difficulty of detecting many NCOs. Measurements of gene conversion frequencies are therefore minimal estimates.
Variation in hot spot activity
Recombination rates at individual hot spots vary over orders of magnitude, from below 0.001 cM to 3 CM11,13,16,32,39,44. In humans23 and mice32, the number of hot spots with different recombination activities follows a simple negative exponential relationship to their activity. A physical model for this has been proposed in which hot spot activity is a function of its DNA-binding characteristics32, which is interesting in light of the physical properties of PR domain-containing 9 (PRDM9, also known as Meisetz), a protein that regulates hot spot activity and is described in a later section.
Imprinting
Imprinting influences recombination in two ways: data suggest that imprinted chromosome regions have higher than average recombination rates45-47 and that the behaviour of chromatids in the recombination process is influenced by their parental origin32,48. In humans, the insulin-like growth factor 2 (IGF2)/H19 and potassium voltage-gated channel subfamily KQT member 1 (KCNQ1) somatically imprinted regions have elevated recombination rates46, and the Prader–Willi syndrome/Angelman syndrome imprinted region shows sex-specific effects on recombination, with alternating domains of recombination that are elevated in males or females47. By comparing multiple hot spots, Lercher and Hurst45 detected elevated female recombination rates in 13 out of 16 human imprinted regions, and Sandovici et al.49 have reported higher historical rates of recombination (measured indirectly by the size of LD domains) and an excess of recombination hot spots in somatically imprinted human chromosome regions.
Imprinting also affects the behaviour of individual chromatids entering meiosis. This seems to contradict our current understanding of how DNA methylation patterns are reset at each generation, as in mice methylation is erased early in gestation. It is re-established before the onset of meiosis in males, leaving both sets of chromatids identically marked, and only after the onset of meiosis in females, again leaving both chromatids identical with respect to methylation50-53 (BOX 2). Despite this, Paigen et al.32 found evidence for parent-of-origin effects on regional rates of recombination along mouse Chr 1, and Ng et al.48 have described a cluster of three imprinted hot spots in the somatically imprinted Kcnq1 region.
Box 2. Sister versus non-sister chromatid recognition.
Mammalian meiosis must include a mechanism for distinguishing sister from non-sister chromatids. Late recombination nodules (which presumably include most, if not all, of the sites of crossing over along the synaptonemal complex that have been identified by MutL homologue 1 (MLH1) and visualized by electron microscopy111) must involve exchanges between non-sister chromatids, otherwise the length of genetic maps would be shorter than predicted by the number of recombination nodules101. We cannot say the same for non-crossovers (NCOs) as these could occur between sister chromatids and be undetectable genetically, or they could occur between non-sister chromatids, in which case they could be detected genetically as conversions, but with only limited efficiency. It is difficult to envision NCOs being repaired by the non-homologous end joining DNA repair process seen in somatic cells, as this would cause deletions after the resection of 5′ strands at the double-strand break (DSB) catalysed by sporulation-specific 11 (SPO11). These considerations require either that all chromatid interactions arising from SPO11-catalysed DSBs are between non-sister chromatids or, alternatively and less likely, that crossovers (COs) can only derive from interactions between non-sister chromatids, whereas NCOs can derive from sister or non-sister interactions.
In Saccharomyces cerevisiae, recombinant molecules form several times more frequently between homologues than between sister chromatids120. Strand invasion between homologues requires a meiosis-specific recombinase, Dmc1 (REFS 121,122), and a trio of other meiosis-specific proteins, Hop1, Red1 and Mek1 (REFS 123-125), which suppress DSB repair between sister chromatids. Although Dmc1 is well conserved and meiosis-specific in mammals, the other proteins are not, which leaves an open question about how the distinction between sister and non-sister chromatids is achieved in this group of organisms. One thing we do know is that it cannot be based on the presence versus absence of DNA sequence differences between chromatids, as the system works perfectly well in inbred strains of animals in which the maternal and paternal genomes are identical. An attractive possibility is that the mechanisms for distinguishing sister and non-sister chromatids involve the detection of hemimethylated sites. If so, this mechanism is likely to be restricted to organisms that methylate their DNA, such as mammals and higher plants, in distinction to those that do not, such as fungi and invertebrates.
DNA methylation offers a possible mechanism for how this distinction between sister and non-sister chromatids is achieved, but we must consider the two sexes separately. In males, remethylation of DNA occurs prior to meiosis (see main text). If the methylation of new DNA strands synthesized during the S phase of meiosis I in males was delayed until after recombination was initiated, the various chromatids could tell sister from non-sister by which strand was methylated and which was not, either globally or at specific sites. A requirement that both members of a pair of chromatids engaging in recombination be methylated on the same strand would militate against any sister chromatid exchanges. Applying this mechanism to female meiosis requires that the demethylation prior to meiosis be incomplete, as remethylation does not occur until after recombination has been completed (see main text).
There are two possible explanations for the apparent contradiction between the recombination data and the evidence for erasure and re-establishment of methylation patterns. First, the erasure of the original methylation pattern might be incomplete, leaving some regions with a parental imprint (which suggests the existence of transgenerational imprinting)54-60. Second, the methylation mark may be totally erased but leave a historical residue in the local pattern of histone modifications, which can survive DNA replication and subsequently influence DNA methylation patterns61,62. It is worth pointing out that although the three phenomena — the correlation between transcriptional imprinting and recombination rates, direct imprinting of recombination activity and sex specificity of recombination — are ostensibly distinct, they may all reflect differences in local chromatin structure caused by DNA and histone modifications.
The recent expansion in our understanding of mammalian hot spots has highlighted several questions, such as what determines the location and relative activity of hot spots? How do chromatids recognize whether a partner is a sister or non-sister chromatid (BOX 2)? And how is the choice made between the pathways leading to CO and NCO outcomes? The first question is discussed in more detail below, and addressing the other questions should become possible by applying new molecular and genetic technologies to the increasing repertoire of hot spots that are available for study.
Regulation of hot spot activity
The location and relative activity of hot spots is determined both by their own DNA sequence, which acts in a cis manner on each chromatid, and by transacting factors that directly or indirectly interact with these sequences. Understanding how regulation is achieved requires identifying both sets of elements and determining how they interact.
Cis regulation
In mice, cis regulation is apparent at the Psmb9 (REF. 39) and Hlx1 (REF. 32) hot spots, in which DNA sequence differences among strains affect both sex specificity and absolute recombination rates on each chromatid. Similarly, the activity of the hot spot NID1 (REF 15) is influenced by a single SNP in the sequence motif CCCCACCCC24,25, which is causally associated with the activity of human recombination hot spots.
Finding these determinative mammalian sequences has been a challenge. Several genomic properties show some correlation with hot spot locations, including GC content, repetitive elements and gene density; however, none of them can be used reliably to predict hot spot locations. The first strong indication that a consensus sequence element might define a substantial number of hot spots was provided by Myers et al.24. They found that a heptameric sequence (CCTCCCT) is enriched in human hot spots but only if it is embedded in the transposon-like human element long terminal repeat 1B (THE1B) repetitive element. This finding was subsequently refined25 to show that a degenerate 13-bp sequence (CCNCCNTNNCCNC) is present in 41% of human hot spots. However, this sequence does not by itself confer hot spot activity as it is also present in regions that do not show hot spot activity. This finding is reminiscent of the cyclic AMP response element in S. pombe, which provides better activation of hot spot activity when embedded in an 18-bp consensus element63. No equivalent consensus element has been identified for mouse hot spots.
Trans regulation
The first indication that DNA sequences external to a mammalian hot spot might be involved in determining hot spot activity came with the finding that sequences flanking the Psmb9 hot spot in the H2 region of mouse Chr 17 strongly influence both its absolute and sex-specific activity12. Similarly, Neumann and Jeffreys20 found that men who shared an identical 10-kb haplotype surrounding the MSTM1b hot spot nevertheless showed substantial differences in hot spot activity, again implicating a role for external elements in determining hot spot activity.
Trans regulation by Prdm9
Using a genetic screen designed to detect trans-acting regulatory factors, Parvanov et al.64 identified a trans-acting regulator of recombination located on Chr 17 that is required for crossing over at multiple hot spots on Chr 1. At the same time, Grey et al.65 reported that the flanking element of the Psmb9 hot spot that controls crossing over at that hot spot is located at some distance from the hot spot, again acting in trans. Both groups showed that the Chr 17 trans- regulators control the formation of COs and NCO gene conversions at sensitive hot spots, indicating that they determine the activation of the recombination process, almost certainly by affecting the location of the initiating DSB.
PRDM9 is the trans-acting factor that controls hot spot activation in mice as well as the factor that specifically binds to the 13-bp consensus motif that is common to many human hot spots. It is also the hybrid sterility gene that prevents gene flow between subspecies of Mus musculus66, which suggests an additional, important role in evolutionary speciation. PRDM9 contains a KRAB protein-protein binding domain67, a PR/SET domain that can trimethylate histone 3 lysine 4 (H3K4)68 and an array of 8–16 zinc fingers. It is expressed only during early meiosis, and deficiency of the protein results in abnormal meiosis with aberrant location of DSBs68. The PR/SET domain is important in that H3K4 trimethylation precedes recombination at the Hlx1 and Psmb9 hot spots69 and potentiates hot spot activity in S. cerevisiae70.
In mice, PRDM9 activates Psmb9 and the hot spots on Chr 1 (REF 71), and the human PRDM9 protein shows in vitro DNA-binding specificity for the 13-bp consensus motif, described above, that is present in 41% of human hot spots72. Further evidence for the role of Prdm9 comes from computational analyses of its predicted DNA-binding sequences: it is the only primate C2H2 zinc-finger protein for which the human, but not chimpanzee, allele can bind the 13-bp motif73, which is not present in chimpanzee hot spots. Additionally, variant human Prdm9 alleles support recombination at different populations of hot spots72.
The existence of a protein such as PRDM9 provides a resolution to the ‘hot spot paradox’: because the initiating chromatid acquires the DNA sequence of its opposite partner, there should exist strong evolutionary selection against hot spot survival as more active sequences are replaced by less active ones16,74,75. The paradox lies in the observation that hot spots are in fact abundant74. It is now clear that mutations in Prdm9 can create whole new populations of hot spots in a single step, providing a simple means of replacing lost hot spots. Indeed, this probably happened 1-2 million years ago for the common Caucasian allele of Prdm9 (REE 73). It is not surprising then that PRDM9 is highly polymorphic in mice and humans71,72 and in several other vertebrate and invertebrate species76. Variation is concentrated in the three amino acids of each finger that determine DNA binding71-73,76 and seems to be under strong positive selection76,77. In addition to coding sequence changes, minisatellite shuffling has added, subtracted and rearranged the order of the zinc fingers72.
Prdm9 haplotype specificity
The allele of Prdm9 from the CAST mouse strain activates the Hlx1 hot spot of the B6 strain several times more efficiently than it does the CAST Hlx1 allele32; a similar effect is seen at the Psmb9 hot spot39. These effects are probably a consequence of the hot spot paradox74; the CAST Hlx1 sequence has probably been acquiring mutations that diminish its activity in Mus musculus castaneus, whereas the B6 sequence has probably been conserved due to lack of recombination in Mus musculus domesticus, in which the Prdm9 allele does not activate this hot spot.
Other trims-acting controls
Hot spot activation is not the only aspect of hot spot function under distant control. Prdm9 can diminish recombination in some chromosomal regions65, although it is unclear whether this is a direct suppression of recombination or a secondary consequence of enhanced activation elsewhere coupled with a fixed limit on the total amount of recombination on each chromosome. Direct suppression of individual hot spots by CAST alleles of distant loci in B6×CAST crosses has been observed, as has quantitative modulation of hot spot activity71. It is unclear whether these suppression and modulation effects are mediated by Prdm9. It is also not known whether the same factors determine both the location and relative activity of hot spots, or whether two systems exist, one controlling location and the other relative activity (see BOX 3 for a discussion of trans-acting controls in yeast).
Box 3. Hot spot trans-regulation in yeast.
The most extensively characterized example of trans-regulation of a yeast hot spot concerns the ade6-M26 hot spot in the fission yeast Schizosaccharomyces pombe. Here, the presence of a 7-bp cyclic AMP response element (CRE) adjacent to the hot spot is required for the activity of the hot spot; activity is also dependent on the heterodimer formed by two transcription factors, Atf1 and Pcr1 (REF. 126). Further studies have identified several additional trans-acting factors that are required for activation, including several ATP-dependent chromatin-remodelling factors (including Gcn5, Snf22, Ada2 and Mst2, which activate, and Hrp1 and Hrp3, which act antagonistically) and the transcriptional repressors Tup11 and Tup12, which suppress recombination127,128. Further analysis has revealed that the CRE element ATGACGT is appreciably more effective when it resides in an 18-bp consensus element, GNVTATGACGTCATNBNC63.
In the budding yeast Saccharomyces cerevisiae, recombination at the His4 hot spot requires the proteins Rap1, Bas1 and Bas2 (REF. 129) and is suppressed by Set2 (a histone methylase) and Rpd3 (a histone deacetylase), the latter probably acting to deacetylate histone 3 lysine 27 (H3K27), close chromatin and suppress recombination130. Also in S. cerevisiae, recombination between ribosomal DNA repeats is suppressed by Sir2 (REF. 131), and a majority of recombination hot spots are dependent on histone H3K4 trimethylation catalysed by Set1 (REF. 70).
We now turn to the issue of how the location and activity of hot spots affect the larger pattern of recombination along chromosomes.
Recombination at the chromosomal level
The overall pattern of recombination in an organism reflects the aggregate behaviour of its individual hot spots, the activities of which are affected by the multiplicity of effects described above. Additionally, this pattern is affected by a set of interacting factors operating at the chromosomal level, including sex, genetic interference and the positioning of hot spots relative to centromeres and telomeres.
Sex-specific differences in recombination activity
In many organisms, the sexes differ in their overall recombination activity, which is reflected in the lengths of their genetic maps. Female maps are longer than male maps in humans, mice, dogs, pigs and Arabidopsis thaliana32,78-82. In cows the two maps are approximately the same length83, and in sheep females have the shorter map84. Moreover, the sex preference can vary from one chromosome to another.
Sex differences in the cumulative rate of crossing over along an entire chromosome are intimately tied to the phenomenon of genetic interference. Although the physical basis for interference is uncertain, it could involve the release of mechanical stresses associated with chromatid twisting85,86. In mice, nearly all COs are separated by a minimum distance87. In the case of mammalian meioses, male and female interference distances are the same when measured in microns of length along the synaptonemal complex (SC)87, but shorter in females when measured in megabases of DNA sequence. This is because both mouse88 and human89 female chromatids are less compacted and hence longer than their male counterparts, with the result that there are fewer megabases of DNA per micron of length in females (BOX 4).
Box 4. Loops and interference.
The physical structure of chromatids offers a unitary model for how sex differences in recombination, interference and chromatid compaction relate to each other (see figure).
Long loops of DNA, tens to hundreds of kilobases in length132, are attached to the axial elements at the leptotene stage of prophase I through short interstitial stretches of DNA in a manner very similar to classical lampbrush chromosomes. These loops are shorter and hence greater in number in females (left panels) than in males89 (right panels). We also know from cytological studies that the DNA double-strand breaks (DSBs)133,134 (yellow circles in the upper panels) and late recombination nodules marking crossovers (COs)110,135 (red circles in the lower panels) are located along the synaptonemal complex (SC) ratherthan in the loops.
Based on these facts, the interference distance in males will be the same as in females when measured in microns of SC length (lower panel of the figure) but longer when measured in megabases if: the number of DSBs is proportional to the number of attachment points and loops; a modified version of the counting model of interference136-139 pertains (in which we postulate that COs are separated by a fixed number of loops); and the compacted length of chromatids at the pachytene stage is proportional to the number of loops and their attachment points.
Support for this model comes from the finding that loops are shorter and hence more numerous in the subtelomeric region, in which male recombination is enhanced132 (see the right-hand side of each panel; centromeres are marked with blue circles on the left-hand sides of each panel), the finding that interference distances are not dependent on the total number of DSBs91, 109, and from the work of Kleckner et al.86, which indicates that interference is related to the physical structure of chromatids.
We can also conclude that because the number of loops far exceeds the number of DSBs, only about 1–3% of attachment regions will suffer a DSB in any single meiosis.
The consequence is a greater opportunity to fit multiple COs along female chromatids and hence a longer genetic map. A similar phenomenon is seen in higher plants82.
Interestingly, there is evidence that the locations of recombination-initiating DSBs are also subject to interference, but necessarily at a much shorter distance88. Curiously, in the yeast S. cerevisiae, interference is seen between COs and between COs and NCOs but not between NCOs90. In this regard, Baudat and de Massy42 have suggested that the regulation of NCO formation may be fundamentally different in yeast and mammals.
The existence of interference indicates that the fate of a DSB is determined by the fate of adjacent DSBs as well as by its own properties. Clues as to how this communication is achieved come from mutations that disrupt interference, such as the loss in mice of the serine/threonine kinase ataxia telangiectasia mutated (ATM), which has an ill-defined role in maintaining meiotic recombination91, and the loss in Caenorhabditis elegans of any of the subunits of condensin I, a protein component of the SC92,93. This condensin I mutation and the dependence on chromatid compaction described above suggest an important role for the physical organization of the SC (BOX 4).
Sex differences in the distribution of recombination
In addition to the presence of sex-specific hot spots (BOX 5) and the role of interference, there is considerable variation in the sex ratio of activity along the chromosome31,32 (FIG. 2). This is apparent in the extent to which crossing over occurs near telomeres in males, which is likely to reflect the tendency for recombination to initiate more frequently near telomeres in males owing to the shorter time allotted in that sex for bouquet formation94-95. In turn, this enhances the possibility of a second CO on the same chromosome and partially overcomes the impact of differential chromosome compaction. By contrast, female recombination is distributed relatively evenly along chromosomes30-32,81,87, albeit with alternating domains of higher and lower activity from centromere to telomere. The reasons for these regional differences are unclear.
Box 5. Sex-specific hot spots.
Adding to the complexities of understanding hot spot behaviour is the fact that some hot spots show considerable sex specificity, with marked differences in activity such that recombination may only be detected in one sex12,32,39. Interestingly, the existence of regional and local sex-specific differences in recombination activity seem to reflect distinct mechanisms, as sex-specific hot spots can occur in regions in which recombination in the other sex predominates — for example, male-specific hot spots in a region of predominantly female recombination, and vice versa.
How sex specificity is achieved is completely unknown at this time. The most obvious explanation — that regulatory proteins are differentially transcribed in male and female meiosis — does not apply to a regulatory protein such as PR domain-containing 9 (PRDM9), as among the hot spots activated by this protein there are a number that do not show sex-specific differences in activity64.
Regulation of total recombination activity
Known genetic influence on number of chiasmata
There is considerable individual variation in overall rates of recombination both in men and in women30,31,97-100. The first indication that such variation is at least in part genetically determined came with the observation101 that the average number of chiasmata per male meiosis (counted as MutL homologue 1 foci (MLH1 foci)) ranges from 21.5 to 24.9 among four strains of inbred mice; this was later confirmed by the finding of heritable variation in genomic recombination rates in a stock of genetically heterogeneous mice102. The existence of such differences agrees with the ability to select for quantitative changes in recombination rates in Drosophila species103-105. Genome-wide association studies in human populations have begun to identify some of the molecular components of the regulatory system that controls rates of crossing over. Stefansson et al.106 found a 900-kb inversion on human Chr 17 that correlates with increased recombination rates in female carriers in the Icelandic population, and Kong et al.107, studying the same population, found two polymorphisms near the ring finger protein 212 (RNF212) locus that influence recombination frequencies in both sexes, albeit in opposite directions; this latter identification has been replicated independently, with the suggestion of additional loci108. Although these polymorphisms explain only a fraction of the variation in recombination rates among individuals, the ability to map such loci provides a potential means of identifying the molecular components of the control machinery. Proteins that contribute to the intensity of interference may well be the sites of such variation given the importance of this phenomenon in deciding how many COs can be present along a meiotic chromatid.
Relationship between numbers of double-strand breaks and crossovers
In addition to these known genetic effects, ‘crossover homeostasis’ describes the fact that a reduction in the number of initiating DSBs does not result in a corresponding reduction in the number of COs, which implies the existence of a regulatory system that maintains sufficient crossing over in the face of variation in the number of DSBs. This phenomenon was first seen in yeast mutants with reduced sporulation-specific 11 (Spo 11) activity109, and it was subsequently reported in mice carrying only one functional Spo11 allele91.
In mice, the number of DSBs initiating recombination (estimated from staining for RAD51 foci) is about ten times larger110 than the number of COs (estimated from staining for MLH1 foci)101 (see also REF. 111). This order of magnitude difference between the number of DSBs and the number of COs raises several important questions: what happens to the rest of the DSBs? (They must be repaired; see the section on NCO gene conversions.) How is the tight regulation of CO number achieved? And what is the evolutionary significance of the fact that across many species the number of COs is only slightly larger than the number of chromosome arms112,113?
We should note that although the rule of one obligate chiasma per chromosome arm is widely accepted in the literature, the rule for humans now seems to be one chiasma per chromosome rather than per arm, with some meioses having no chiasmata on smaller chromosomes114. In mice, the evidence favours the reverse situation, with one chiasma per arm rather than per chromosome, as strains with as few as 11 chromosomes — but with the usual 20 arms as a result of centromeric fusions — do not have a reduced number of chiasmata115.
Evolution
Rates of hot spot evolution
Hot spot sequences and the macromolecular systems that recognize them must evolve in synchrony. The available evidence suggests that this is quite rapid. To the extent they have been tested, hot spots are not shared between humans and chimpanzees116,117. There is also strong evidence that hot spots are evolving rapidly in mice; there is only a weak correlation between local recombination rates in crosses between different mouse strains9,32.
The best example we have of evolution at a specific hot spot is the work of Jeffreys and Neumann17, which describes a human hot spot that is estimated to have arisen through a single-base-pair substitution approximately 70,000 years ago (range 7,000–270,000 years ago). In this hot spot the activating mutation is distinctly undertransmitted, and computer simulations suggest that the hot spot will inevitably die.
Although hot spot usage is not well conserved among mouse strains, broader scale regional recombination rates are conserved32, suggesting that the factors controlling regional rates of recombination operate independently from the regulatory systems controlling the location and behaviour of individual hot spots.
Variation in genome-wide rates of recombination
The variation in overall recombination rates among mammals, expressed as the rate of crossing over per unit length of DNA (cM Mb−1), seems to be the product of neutral evolution118. Expressed in this dimension, higher rates are correlated with an increase in the number of chromosome arms (possibly increasing the number of obligate COs per genome) and/or a decrease in the interference distance (permitting multiple COs on the same arm). Relating rates of recombination to numbers of chromosome arms4, 118 suggests that among rodents (mice and rats) the change in overall recombination rates is related to the number of chromosome arms, whereas among primates (humans, baboons and macaques), horses, cats, dogs, pigs, sheep and cows the change is related to interference, emphasizing the part that interference plays.
Selection minimizes crossing over
Despite its obvious biological value in generating population diversity and promoting efficient selection against deleterious mutations, levels of meiotic crossing over among species are not much greater than what is required to assure at least one chiasma per chromosome arm, suggesting a counterintuitive evolutionary selection against additional crossing over. The existence of a much larger and variable number of DSBs initiating recombination, with their risk of imprecise DNA repair, carries the surprising implication that selection is directed at limiting genetic exchanges as much as against the possibility of DNA damage from DSBs. It is conceivable that the DSB repair pathway that gives rise to COs carries some hitherto unrecognized genomic danger. Given the importance of interference in spacing chiasmata, the presently unknown molecular basis for interference is the likely target for such evolutionary selection. These observations raise still another conundrum: given that the total amount of genetic recombination is related to the number of chromosome arms and that there is likely to be selection against excessive crossing over, why are mammalian species so variable in their numbers of chromosomes and chromosome arms?
Summary
We now possess a reasonably comprehensive view of how mammalian hot spots are distributed along chromosomes and a description of many of the properties of hot spots. We now also know of the existence of regulatory systems that control chiasma numbers, the activation of specific hot spots and the quantitative modulation of hot spot activity, although many of the features of these regulatory systems remain unknown. These findings have also clarified the existence of substantial gaps in our understanding and have raised several experimental challenges. Some of these are described below.
The identity of the mammalian mechanisms that decide which hot spots go on to become COs and which become NCOs remains obscure.
We do not understand the nature of the factors that control the regional distribution of recombination along chromosomes, the mechanisms that control chromatid compaction and interference nor the interactions between these factors and mechanisms. These may be related by their dependence on the magnitude of DNA looping along chromosomes, although as yet we have no direct evidence of this. Moreover, if looping is a factor, we do not understand why loop sizes vary along the chromosome and what determines them.
We are still uncertain about the identity of the DNA sequences that define hot spots; we have only a partial answer for the human genome and no clear answer yet for mice. We also need to identify the proteins that interact with these sequences to initiate and/or regulate the recombination process; PRDM9 is the only such protein to be identified, but there are probably others.
We can only guess how sister and non-sister chromatids are distinguished; we have suggested that this may depend on the methylation state of DNA strands when recombination initiates, but there is no direct evidence of this. A related phenomenon is meiotic imprinting, which implies the persistence of parental chromosome marking through germ cell maturation and entry into meiosis.
Finally, we do not understand multiple aspects of sex specificity in meiotic recombination (particularly the varying ratios of activity along chromosomes) nor the impact telomere adjacency has on elevating male recombination, and the mechanism by which sex-specific hot spots are generated is still up for debate.
Acknowledgments
The authors are thankful to M. A. Handel for her thoughtful comments. This work was supported by US National Institutes of Health grants GM078643 and GM083408 to K.P., grant GM078452 to P.P., project grant GM076468 (part of which is managed by P. P.) to G. Churchill and grant CA34196 to The Jackson Laboratory.
Glossary
- Cosmid
A bacterial recombination vector that contains long inserted DNA sequences.
- Single-nucleotide polymorphism
Single-nucleotide polymorphisms (SNPs) distinguish the chromosomes of two individuals or mouse strains. There are millions of SNPs in mammalian genomes, and they have become the preferred markers for genetic studies.
- Haplotype
A set of genetic markers that are present on a single chromosome and that show complete or nearly complete linkage disequilibrium — that is they are inherited through generations without being changed by crossing over or other recombination mechanisms.
- Linkage disequilibrium
Preferential association of allelic combinations among groups of nearby genes.
- Genetic drift
The random fluctuations in allele frequencies over time that are due to chance alone.
- Coalescent-based statistical methods
Methods of reconstructing population history by simulating the genealogy of genes back to the most recent common ancestor of all alleles currently in the population.
- Genome-wide association study
An examination of common genetic variation across the genome that is designed to identify associations with traits, such as common diseases.
- Imprinting
The epigenetic marking of a gene on the basis of parental origin, which in somatic tissues results in monoallelic expression.
- Crossover
A reciprocal exchange of DNA along chromatids such that the proximal end of one homologue becomes attached to the distal end of the other.
- Double-strand break
A cut made across a DNA molecule in which both strands are broken. In recombination the cut is made by the enzyme sporulation-specific 11 (SPO11).
- Pseudoautosomal
A region on a sex chromosome that is homologous between the X chromosome and the Y chromosome. Successful meiosis in males requires a crossover in this pseudoautosomal region.
- Gene conversion
The process during recombination in which a short segment of DNA on the initiating chromatid is replaced by the DNA sequence of its partner without the exchange of flanking markers.
- Holliday junction
The point at which the strands of two dsDNA molecules exchange partners as an intermediate step in crossing over. Typically, two Holliday junctions are formed in the recombination pathway that gives rise to crossovers.
- Topoisomerase
An ATP-dependent enzyme that normally creates transient breaks in both strands of the DNA sugar-phosphate backbone, then passes one strand through the other and reseals the break. In the case of the topoisomerase sporulation-specific 11 (SPO 11), which initiates recombination, the breaks are not immediately reseated because the 5′ strand on each end is rapidly resected leaving a free 3′ overhang.
- Chromatid
The product of chromosome replication in meiosis I. Chromatids are distinguished from chromosomes by the fact that the two daughter chromatids of one chromosome remain attached at their centromeres through meiosis I cell division.
- Recombination nodules
The early, visible manifestations of sites of chiasmata and crossing over. They are recognized by immunochemical staining, typically for the protein MutL homologue 1, which is a component of late recombination nodules.
- Zinc finger
A protein loop in which cysteine or cysteine-histidine residues coordinate a zinc ion to form the base of the loop. Three of the amino acids in the loop cooperate to recognize three base pairs of DNA, and a tandem array of zinc fingers can show considerable DNA-binding specificity.
- Positive selection
A process by which natural selection favours a single beneficial genotype over other genotypes and may drive this genotype to a high frequency in a population.
- Minisatellite
A region of DNA in which repeat units of 10–50 bp are tandemly arranged in arrays 0.5–30 kb in length.
- Genetic interference
The presence of a recombinational event in one region that affects the occurrence of recombinational events in adjacent regions. Positive interference, which is seen in eukaryotes, reduces the probability of using nearby hot spots in the same meiosis and causes a more even spacing of crossovers than would occur by chance.
- Synaptonemal complex
A linear protein complex that forms the backbone of each chromatid during prophase I of meiosis and promotes genetic recombination. The DNA of the chromatid is attached to the complex in long loops. The name is derived from the word synapsis, which has been used to describe chromatid pairing.
- Bouquet formation
The clustering of telomeres together on the nuclear membrane early in meiosis.
- Chiasmata
A chiasma (plural chiasmata) is the cytologically visible physical connection between homologous chromatids during meiosis that corresponds to the sites of genetic crossing over.
- MutL homologue 1 foci
Sites of staining for MutL homologue 1 that identify sites of genetic crossing over.
- RAD51
The human homologue of bacterial RecA. RAD51 is required for homologous recombination, during which it promotes strand invasion forming nucleoprotein filaments around ssDNA. Immunohistochemical staining of RAD51 foci identifies sites of DNA double-strand breaks.
Footnotes
Competing interests statement: The authors declare no competing financial interests.
Contributor Information
Kenneth Paigen, Email: ken.paigen@jax.org, The Jackson Laboratory, 600 Main Street, Bar Harbor, Maine 04609, USA.
Petko Petkov, Email: petko.petkov@jax.org, The Jackson Laboratory, 600 Main Street, Bar Harbor, Maine 04609, USA.
References
- 1.Arnheim N, Calabrese P, Tiemann-Boege I. Mammalian meiotic recombination hot spots. Annu Rev Genet. 2007;41:369–399. doi: 10.1146/annurev.genet.41.110306.130301. [DOI] [PubMed] [Google Scholar]
- 2.Buard J, de Massy B. Playing hide and seek with mammalian meiotic crossover hotspots. Trends Genet. 2007;23:301–309. doi: 10.1016/j.tig.2007.03.014. [DOI] [PubMed] [Google Scholar]
- 3.de Massy B. Distribution of meiotic recombination sites. Trends Genet. 2003;19:514–522. doi: 10.1016/S0168-9525(03)00201-4. [DOI] [PubMed] [Google Scholar]
- 4.Coop G, Przeworski M. An evolutionary view of human recombination. Nature Rev Genet. 2007;8:23–34. doi: 10.1038/nrg1947. [DOI] [PubMed] [Google Scholar]
- 5.Kauppi L, Jeffreys AJ, Keeney S. Where the crossovers are: recombination distributions in mammals. Nature Rev Genet. 2004;5:413–424. doi: 10.1038/nrg1346. [DOI] [PubMed] [Google Scholar]
- 6.Steinmetz M, et al. A molecular map of the immune response region from the major histocompatibility complex of the mouse. Nature. 1982;300:35–42. doi: 10.1038/300035a0. The paper in which a recombination hot spot was first described and in which the term ‘hot spot’ was first used. [DOI] [PubMed] [Google Scholar]
- 7.Chakravarti A, et al. Nonuniform recombination within the human β-globin gene cluster. Am J Hum Genet. 1984;36:1239–1258. [PMC free article] [PubMed] [Google Scholar]
- 8.Chakravarti A, Elbein SC, Permutt MA. Evidence for increased recombination near the human insulin gene: implication for disease association studies. Proc Natl Acad Sci USA. 1986;83:1045–1049. doi: 10.1073/pnas.83.4.1045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kelmenson PM, et al. A torrid zone on mouse chromosome 1 containing a cluster of recombinational hotspots. Genetics. 2005;169:833–841. doi: 10.1534/genetics.104.035063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Buchner DA, Trudeau M, George AL, Jr, Sprunger LK, Meisler MH. High-resolution mapping of the sodium channel modifier Scnm 1 on mouse chromosome 3 and identification of a 1.3-kb recombination hot spot. Genomics. 2003;82:452–459. doi: 10.1016/s0888-7543(03)00152-6. [DOI] [PubMed] [Google Scholar]
- 11.Bois PR. A highly polymorphic meiotic recombination mouse hot spot exhibits incomplete repair. Mol Cell Biol. 2007;27:7053–7062. doi: 10.1128/MCB.00874-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Shiroishi T, Sagai T, Hanzawa N, Gotoh H, Moriwaki K. Genetic control of sex-dependent meiotic recombination in the major histocompatibility complex of the mouse. EMBO J. 1991;10:681–686. doi: 10.1002/j.1460-2075.1991.tb07997.x. The initial demonstration that sequences outside the hot spot itself can determine hot spot activity and sex specificity. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Jeffreys AJ, Kauppi L, Neumann R. Intensely punctate meiotic recombination in the class II region of the major histocompatibility complex. Nature Genet. 2001;29:217–222. doi: 10.1038/ng1001-217. [DOI] [PubMed] [Google Scholar]
- 14.Jeffreys AJ, May CA. Intense and highly localized gene conversion activity in human meiotic crossover hot spots. Nature Genet. 2004;36:151–156. doi: 10.1038/ng1287. [DOI] [PubMed] [Google Scholar]
- 15.Jeffreys AJ, Neumann R. Factors influencing recombination frequency and distribution in a human meiotic crossover hotspot. Hum Mol Genet. 2005;14:2277–2287. doi: 10.1093/hmg/ddi232. [DOI] [PubMed] [Google Scholar]
- 16.Jeffreys AJ, Neumann R. Reciprocal crossover asymmetry and meiotic drive in a human recombination hot spot. Nature Genet. 2002;31:267–271. doi: 10.1038/ng910. This paper and reference 13 are excellent examples of the use of sperm assays to define the molecular details of recombination products at human hot spots. [DOI] [PubMed] [Google Scholar]
- 17.Jeffreys AJ, Neumann R. The rise and fall of a human recombination hot spot. Nature Genet. 2009;41:625–629. doi: 10.1038/ng.346. An intriguing paper that analysed the evolutionary appearance of a new hot spot and documented its eventual decline. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Jeffreys AJ, Neumann R, Panayi M, Myers S, Donnelly P. Human recombination hot spots hidden in regions of strong marker association. Nature Genet. 2005;37:601–606. doi: 10.1038/ng1565. [DOI] [PubMed] [Google Scholar]
- 19.Jeffreys AJ, Ritchie A, Neumann R. High resolution analysis of haplotype diversity and meiotic crossover in the human TAP2 recombination hotspot. Hum Mol Genet. 2000;9:725–733. doi: 10.1093/hmg/9.5.725. [DOI] [PubMed] [Google Scholar]
- 20.Neumann R, Jeffreys AJ. Polymorphism in the activity of human crossover hotspots independent of local DNA sequence variation. Hum Mol Genet. 2006;15:1401–1411. doi: 10.1093/hmg/ddl063. [DOI] [PubMed] [Google Scholar]
- 21.Gabriel SB, et al. The structure of haplotype blocks in the human genome. Science. 2002;296:2225–2229. doi: 10.1126/science.1069424. [DOI] [PubMed] [Google Scholar]
- 22.Crawford DC, et al. Evidence for substantial fine-scale variation in recombination rates across the human genome. Nature Genet. 2004;36:700–706. doi: 10.1038/ng1376. [DOI] [PubMed] [Google Scholar]
- 23.McVean GA, et al. The fine-scale structure of recombination rate variation in the human genome. Science. 2004;304:581–584. doi: 10.1126/science.1092500. [DOI] [PubMed] [Google Scholar]
- 24.Myers S, Bottolo L, Freeman C, McVean G, Donnelly P. A fine-scale map of recombination rates and hotspots across the human genome. Science. 2005;310:321–324. doi: 10.1126/science.1117196. [DOI] [PubMed] [Google Scholar]
- 25.Myers S, Freeman C, Auton A, Donnelly P, McVean G. A common sequence motif associated with recombination hot spots and genome instability in humans. Nature Genet. 2008;40:1124–1129. doi: 10.1038/ng.213. [DOI] [PubMed] [Google Scholar]
- 26.Tiemann-Boege I, Calabrese P, Cochran DM, Sokol R, Arnheim N. High-resolution recombination patterns in a region of human chromosome 21 measured by sperm typing. PLoS Genet. 2006;2:e70. doi: 10.1371/journal.pgen.0020070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Tapper W, et al. A map of the human genome in linkage disequilibrium units. Proc Natl Acad Sci USA. 2005;102:11835–11839. doi: 10.1073/pnas.0505262102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Coop G, Wen X, Ober C, Pritchard JK, Przeworski M. High-resolution mapping of crossovers reveals extensive variation in fine-scale recombination patterns among humans. Science. 2008;319:1395–1398. doi: 10.1126/science.1151851. [DOI] [PubMed] [Google Scholar]
- 29.Kauppi L, Stumpf MP, Jeffreys AJ. Localized breakdown in linkage disequilibrium does not always predict sperm crossover hot spots in the human MHC class II region. Genomics. 2005;86:13–24. doi: 10.1016/j.ygeno.2005.03.011. [DOI] [PubMed] [Google Scholar]
- 30.Kong A, et al. A high-resolution recombination map of the human genome. Nature Genet. 2002;31:241–247. doi: 10.1038/ng917. [DOI] [PubMed] [Google Scholar]
- 31.Broman KW, Murray JC, Sheffield VC, White RL, Weber JL. Comprehensive human genetic maps: individual and sex-specific variation in recombination. Am J Hum Genet. 1998;63:861–869. doi: 10.1086/302011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Paigen K, et al. The recombinational anatomy of a mouse chromosome. PLoS Genet. 2008;4:e1000119. doi: 10.1371/journal.pgen.1000119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Li HH, et al. Amplification and analysis of DNA sequences in single human sperm and diploid cells. Nature. 1988;335:414–417. doi: 10.1038/335414a0. [DOI] [PubMed] [Google Scholar]
- 34.Cullen M, Perfetto SP, Klitz W, Nelson G, Carrington M. High-resolution patterns of meiotic recombination across the human major histocompatibility complex. Am J Hum Genet. 2002;71:759–776. doi: 10.1086/342973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Arnheim N, Calabrese P, Nordborg M. Hot and cold spots of recombination in the human genome: the reason we should find them and how this can be achieved. Am J Hum Genet. 2003;73:5–16. doi: 10.1086/376419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Guillon H, Baudat F, Grey C, Liskay RM, de Massy B. Crossover and Noncrossover Pathways in Mouse Meiosis. Molecular Cell. 2005;20:563–573. doi: 10.1016/j.molcel.2005.09.021. [DOI] [PubMed] [Google Scholar]
- 37.Guillon H, de Massy B. An initiation site for meiotic crossing-over and gene conversion in the mouse. Nature Genet. 2002;32:296–299. doi: 10.1038/ng990. [DOI] [PubMed] [Google Scholar]
- 38.May CA, Shone AC, Kalaydjieva L, Sajantila A, Jeffreys AJ. Crossover clustering and rapid decay of linkage disequilibrium in the Xp/Yp pseudoautosomal gene SHOX. Nature Genet. 2002;31:272–275. doi: 10.1038/ng918. [DOI] [PubMed] [Google Scholar]
- 39.Baudat F, de Massy B. Cis- and trans-acting elements regulate the mouse Psmb9 meiotic recombination hotspot. PLoS Genet. 2007;3:e100. doi: 10.1371/journal.pgen.0030100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Ng SH, Parvanov E, Petkov PM, Paigen K. A quantitative assay for crossover and noncrossover molecular events at individual recombination hotspots in both male and female gametes. Genomics. 2008;92:204–209. doi: 10.1016/j.ygeno.2008.06.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Huang X, et al. High-throughput genotyping by whole-genome resequencing. Genome Res. 2009;19:1068–1076. doi: 10.1101/gr.089516.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Baudat F, de Massy B. Regulating double-stranded DNA break repair towards crossover or non-crossover during mammalian meiosis. Chromosome Res. 2007;15:565–577. doi: 10.1007/s10577-007-1140-3. [DOI] [PubMed] [Google Scholar]
- 43.Holloway K, Lawson VE, Jeffreys AJ. Allelic recombination and de novo deletions in sperm in the human β-globin gene region. Hum Mol Genet. 2006;15:1099–1111. doi: 10.1093/hmg/ddl025. [DOI] [PubMed] [Google Scholar]
- 44.Jeffreys AJ, et al. Meiotic recombination hot spots and human DNA diversity. Phil Trans R Soc Lond B. 2004;359:141–152. doi: 10.1098/rstb.2003.1372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Lercher MJ, Hurst LD. Imprinted chromosomal regions of the human genome have unusually high recombination rates. Genetics. 2003;165:1629–1632. doi: 10.1093/genetics/165.3.1629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Paldi A, Gyapay G, Jami J. Imprinted chromosomal regions of the human genome display sex-specific meiotic recombination frequencies. Curr Biol. 1995;5:1030–1035. doi: 10.1016/s0960-9822(95)00207-7. [DOI] [PubMed] [Google Scholar]
- 47.Robinson WP, Lalande M. Sex-specific meiotic recombination in the Prader–Willi/Angelman syndrome imprinted region. Hum Mol Genet. 1995;4:801–806. doi: 10.1093/hmg/4.5.801. [DOI] [PubMed] [Google Scholar]
- 48.Ng SH, et al. Parental origin of chromosomes influences crossover activity within the Kcnq1 transcriptionally imprinted domain of Mus musculus. BMC Mol Biol. 2009;10:43. doi: 10.1186/1471-2199-10-43. This study provides direct evidence that the parental origin of chromatids affects their recombinatorial behaviour. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Sandovici I, et al. Human Imprinted Chromosomal Regions Are Historical Hot-Spots of Recombination. PLoS Genetics. 2006;2:e101. doi: 10.1371/journal.pgen.0020101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Lucifero D, Mann MR, Bartolomei MS, Trasler JM. Gene-specific timing and epigenetic memory in oocyte imprinting. Hum Mol Genet. 2004;13:839–849. doi: 10.1093/hmg/ddh104. [DOI] [PubMed] [Google Scholar]
- 51.Kono T, Obata Y, Yoshimzu T, Nakahara T, Carroll J. Epigenetic modifications during oocyte growth correlates with extended parthenogenetic development in the mouse. Nature Genet. 1996;13:91–94. doi: 10.1038/ng0596-91. [DOI] [PubMed] [Google Scholar]
- 52.Sato S, Yoshimizu T, Sato E, Matsui Y. Erasure of methylation imprinting of Igf2r during mouse primordial germ-cell development. Mol Reprod Dev. 2003;65:41–50. doi: 10.1002/mrd.10264. [DOI] [PubMed] [Google Scholar]
- 53.Hajkova P, et al. Epigenetic reprogramming in mouse primordial germ cells. Mech Dev. 2002;117:15–23. doi: 10.1016/s0925-4773(02)00181-8. [DOI] [PubMed] [Google Scholar]
- 54.Han Z, Mtango NR, Patel BG, Sapienza C, Latham KE. Hybrid vigor and transgenerational epigenetic effects on early mouse embryo phenotype. Biol Reprod. 2008;79:638–648. doi: 10.1095/biolreprod.108.069096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Rakyan VK, et al. Transgenerational inheritance of epigenetic states at the murine AxinFu allele occurs after maternal and paternal transmission. Proc Natl Acad Sci USA. 2003;100:2538–2543. doi: 10.1073/pnas.0436776100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Morgan HD, Sutherland HG, Martin DI, Whitelaw E. Epigenetic inheritance at the agouti locus in the mouse. Nat Genet. 1999;23:314–318. doi: 10.1038/15490. [DOI] [PubMed] [Google Scholar]
- 57.Sutherland HG, et al. Reactivation of heritably silenced gene expression in mice. Mamm Genome. 2000;11:347–355. doi: 10.1007/s003350010066. [DOI] [PubMed] [Google Scholar]
- 58.Herman H, et al. Trans allele methylation and paramutation-like effects in mice. Nature Genet. 2003;34:199–202. doi: 10.1038/ng1162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Cuzin F, Grandjean V, Rassoulzadegan M. Inherited variation at the epigenetic level: paramutation from the plant to the mouse. Curr Opin Genet Dev. 2008;18:193–196. doi: 10.1016/j.gde.2007.12.004. [DOI] [PubMed] [Google Scholar]
- 60.Rassoulzadegan M, et al. RNA-mediated non-mendelian inheritance of an epigenetic change in the mouse. Nature. 2006;441:469–474. doi: 10.1038/nature04674. [DOI] [PubMed] [Google Scholar]
- 61.Cedar H, Bergman Y. Linking DNA methylation and histone modification: patterns and paradigms. Nature Rev Genet. 2009;10:295–304. doi: 10.1038/nrg2540. [DOI] [PubMed] [Google Scholar]
- 62.Probst AV, Dunleavy E, Almouzni G. Epigenetic inheritance during the cell cycle. Nature Rev Mol Cell Biol. 2009;10:192–206. doi: 10.1038/nrm2640. [DOI] [PubMed] [Google Scholar]
- 63.Steiner WW, Smith GR. Optimizing the nucleotide sequence of a meiotic recombination hotspot in Schizosaccharomyces pombe. Genetics. 2005;169:1973–1983. doi: 10.1534/genetics.104.039230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Parvanov ED, Ng SH, Petkov PM, Paigen K. Trans-regulation of mouse meiotic recombination hotspots by Rcr1. PLoS Biology. 2009;7:e1000036. doi: 10.1371/journal.pbio.1000036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Grey C, Baudat F, de Massy B. Genome-wide control of the distribution of meiotic recombination. PLoS Biol. 2009;7:e35. doi: 10.1371/journal.pbio.1000035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Mihola O, Trachtulec Z, Vlcek C, Schimenti JC, Forejt J. A mouse speciation gene encodes a meiotic histone H3 methyltransferase. Science. 2009;323:373–375. doi: 10.1126/science.1163601. [DOI] [PubMed] [Google Scholar]
- 67.Birtle Z, Ponting CP. Meisetz and the birth of the KRAB motif. Bioinformatics. 2006;22:2841–2845. doi: 10.1093/bioinformatics/btl498. [DOI] [PubMed] [Google Scholar]
- 68.Hayashi K, Yoshida K, Matsui Y. A histone H3 methyltransferase controls epigenetic events required for meiotic prophase. Nature. 2005;438:374–378. doi: 10.1038/nature04112. [DOI] [PubMed] [Google Scholar]
- 69.Buard J, Barthes P, Grey C, de Massy B. Distinct histone modifications define initiation and repair of meiotic recombination in the mouse. Embo J. 2009;28:2616–2624. doi: 10.1038/emboj.2009.207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Borde V, et al. Histone H3 lysine 4 trimethylation marks meiotic recombination initiation sites. Embo J. 2009;28:99–111. doi: 10.1038/emboj.2008.257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Parvanov ED, Petkov PM, Paigen K. Prdm9 controls activation of mammalian recombination hotspots. Science. 2009 Dec 31; doi: 10.1126/science.1181495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Baudat F, et al. PRDM9 is a major determinant of meiotic recombination hotspots in humans and mice. Science. 2009 Dec 31; doi: 10.1126/science.1183439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Myers S, et al. Drive against hotspot motifs in primates implicates the PRDM9 gene in meiotic recombination. Science. 2009 Dec 31; doi: 10.1126/science.1182363. References 71–73 provide evidence that PRDM9 is a major trans-acting regulator of hot spot activity in humans and mice. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Boulton A, Myers RS, Redfield RJ. The hotspot conversion paradox and the evolution of meiotic recombination. Proc Natl Acad Sci USA. 1997;94:8058–8063. doi: 10.1073/pnas.94.15.8058. The first paper to point out the paradox that hot spots are prevalent despite strong selection against their survival from mutations that diminish their activity. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Jeffreys AJ, Neumann R. Factors influencing recombination frequency and distribution in a human meiotic crossover hotspot. Hum Mol Genet. 2005;14:2277–2287. doi: 10.1093/hmg/ddi232. [DOI] [PubMed] [Google Scholar]
- 76.Oliver PL, et al. Accelerated evolution of the Prdm9 speciation gene across diverse metazoan taxa. PLoS Genet. 2009;5:e1000753. doi: 10.1371/journal.pgen.1000753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Thomas JH, Emerson RO, Shendure J. Extraordinary molecular evolution in the PRDM9 fertility gene. PLOS ONE. 2009;4:e8505. doi: 10.1371/journal.pone.0008505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Neff MW, et al. A second-generation genetic linkage map of the domestic dog, Canis familiaris. Genetics. 1999;151:803–820. doi: 10.1093/genetics/151.2.803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Mikawa S, et al. A linkage map of 243 DNA markers in an intercross of Gottingen miniature and Meishan pigs. Anim Genet. 1999;30:407–17. doi: 10.1046/j.1365-2052.1999.00493.x. [DOI] [PubMed] [Google Scholar]
- 80.Lynn A, Ashley T, Hassold T. Variation in human meiotic recombination. Annu Rev Genomics Hum Genet. 2004;5:317–349. doi: 10.1146/annurev.genom.4.070802.110217. [DOI] [PubMed] [Google Scholar]
- 81.Cox A, et al. A new standard genetic map for the laboratory mouse. Genetics. 2009;182:1335–1344. doi: 10.1534/genetics.109.105486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Drouaud J, et al. Sex-Specific crossover distributions and variations in interference level along Arabidopsis thaliana chromosome 4. PLoS Genet. 2007;3:e106. doi: 10.1371/journal.pgen.0030106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Kappes SM, et al. A second-generation linkage map of the bovine genome. Genome Res. 1997;7:235–249. doi: 10.1101/gr.7.3.235. [DOI] [PubMed] [Google Scholar]
- 84.Maddox JF, et al. An enhanced linkage map of the sheep genome comprising more than 1000 loci. Genome Res. 2001;11:1275–1289. doi: 10.1101/gr.135001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Borner GV, Kleckner N, Hunter N. Crossover/noncrossover differentiation, synaptonemal complex formation, and regulatory surveillance at the leptotene/zygotene transition of meiosis. Cell. 2004;117:29–45. doi: 10.1016/s0092-8674(04)00292-2. [DOI] [PubMed] [Google Scholar]
- 86.Kleckner N, et al. A mechanical basis for chromosome function. Proc Natl Acad Sci U S A. 2004;101:12592–12597. doi: 10.1073/pnas.0402724101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Petkov PM, Broman KW, Szatkiewicz JP, Paigen K. Crossover interference underlies sex differences in recombination rates. Trends Genet. 2007;23:539–542. doi: 10.1016/j.tig.2007.08.015. This paper shows that CO interference underlying sex differences in overall recombination rates is a function of the reduced compaction of the SC in females relative to males. [DOI] [PubMed] [Google Scholar]
- 88.de Boer E, Stam P, Dietrich AJ, Pastink A, Heyting C. Two levels of interference in mouse meiotic recombination. Proc Natl Acad Sci U S A. 2006;103:9607–9612. doi: 10.1073/pnas.0600418103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Tease C, Hulten MA. Inter-sex variation in synaptonemal complex lengths largely determine the different recombination rates in male and female germ cells. Cytogenet Genome Res. 2004;107:208–215. doi: 10.1159/000080599. [DOI] [PubMed] [Google Scholar]
- 90.Mancera E, Bourgon R, Brozzi A, Huber W, Steinmetz LM. High-resolution mapping of meiotic crossovers and non-crossovers in yeast. Nature. 2008;454:479–485. doi: 10.1038/nature07135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Barchi M, et al. ATM promotes the obligate XY crossover and both crossover control and chromosome axis integrity on autosomes. PLoS Genet. 2008;4:e1000076. doi: 10.1371/journal.pgen.1000076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Mets DG, Meyer BJ. Condensins regulate meiotic DNA break distribution, thus crossover frequency, by controlling chromosome structure. Cell. 2009;139:73–86. doi: 10.1016/j.cell.2009.07.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Tsai CJ, et al. Meiotic crossover number and distribution are regulated by a dosage compensation protein that resembles a condensin subunit. Genes Dev. 2008;22:194–211. doi: 10.1101/gad.1618508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Roig I, et al. Female-specific features of recombinational double-stranded DNA repair in relation to synapsis and telomere dynamics in human oocytes. Chromosoma. 2004;113:22–33. doi: 10.1007/s00412-004-0290-8. [DOI] [PubMed] [Google Scholar]
- 95.Pfeifer C, Scherthan H, Thomsen PD. Sex-specific telomere redistribution and synapsis initiation in cattle oogenesis. Dev Biol. 2003;255:206–215. doi: 10.1016/s0012-1606(02)00093-3. [DOI] [PubMed] [Google Scholar]
- 96.Tankimanova M, Hulten MA, Tease C. The initiation of homologous chromosome synapsis in mouse fetal oocytes is not directly driven by centromere and telomere clustering in the bouquet. Cytogenet Genome Res. 2004;105:172–181. doi: 10.1159/000078187. [DOI] [PubMed] [Google Scholar]
- 97.Lynn A, et al. Covariation of synaptonemal complex length and mammalian meiotic exchange rates. Science. 2002;296:2222–2225. doi: 10.1126/science.1071220. [DOI] [PubMed] [Google Scholar]
- 98.Hassold T, et al. Cytological studies of meiotic recombination in human males. Cytogenet Genome Res. 2004;107:249–255. doi: 10.1159/000080602. [DOI] [PubMed] [Google Scholar]
- 99.Sun F, et al. Variation in MLH1 distribution in recombination maps for individual chromosomes from human males. Hum Mol Genet. 2006;15:2376–2391. doi: 10.1093/hmg/ddl162. [DOI] [PubMed] [Google Scholar]
- 100.Laurie DA, Hulten MA. Further studies on bivalent chiasma frequency in human males with normal karyotypes. Ann Hum Genet. 1985;49:189–201. doi: 10.1111/j.1469-1809.1985.tb01693.x. [DOI] [PubMed] [Google Scholar]
- 101.Koehler KE, Cherry JP, Lynn A, Hunt PA, Hassold TJ. Genetic control of mammalian meiotic recombination. I. Variation in exchange frequencies among males from inbred mouse strains. Genetics. 2002;162:297–306. doi: 10.1093/genetics/162.1.297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Dumont BL, Broman KW, Payseur BA. Variation in genomic recombination rates among heterogeneous stock mice. Genetics. 2009;182:1345–1349. doi: 10.1534/genetics.109.105114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Charlesworth B, Charlesworth D. Genetic variation in recombination in Drosophila. I. Responses to selection and preliminary genetic analysis. Heredity. 1985;54:71–83. [Google Scholar]
- 104.Charlesworth B, Charlesworth D. Genetic variation in recombination in Drosophila. II. Genetic analysis of a high recombination stock. Heredity. 1985;54:85–98. [Google Scholar]
- 105.Detlefsen JA, Roberts E. Studies on crossing over. I. The effect of selection on crossover values. J Exp Zoology. 1921;32:333–354. [Google Scholar]
- 106.Stefansson H, et al. A common inversion under selection in Europeans. Nature Genet. 2005;37:129–137. doi: 10.1038/ng1508. [DOI] [PubMed] [Google Scholar]
- 107.Kong A, et al. Sequence variants in the RNF212 gene associate with genome-wide recombination rate. Science. 2008;319:1398–1401. doi: 10.1126/science.1152422. Describes the use of a genome-wide association study to map genes that determine overall recombination rates in humans. [DOI] [PubMed] [Google Scholar]
- 108.Chowdhury R, Bois PR, Feingold E, Sherman SL, Cheung VG. Genetic analysis of variation in human meiotic recombination. PLoS Genet. 2009;5:e1000648. doi: 10.1371/journal.pgen.1000648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Martini E, Diaz RL, Hunter N, Keeney S. Crossover homeostasis in yeast meiosis. Cell. 2006;126:285–295. doi: 10.1016/j.cell.2006.05.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Moens PB, et al. The time course and chromosomal localization of recombination-related proteins at meiosis in the mouse are compatible with models that can resolve the early DNA–DNA interactions without reciprocal recombination. J Cell Sci. 2002;115:1611–1622. doi: 10.1242/jcs.115.8.1611. A detailed description of the physical locations of a number of key molecules relative to the SC during recombination. [DOI] [PubMed] [Google Scholar]
- 111.Moens PB, Marcon E, Shore JS, Kochakpour N, Spyropoulos B. Initiation and resolution of interhomolog connections: crossover and non-crossover sites along mouse synaptonemal complexes. J Cell Sci. 2007;120:1017–1027. doi: 10.1242/jcs.03394. [DOI] [PubMed] [Google Scholar]
- 112.Dutrillaux B. Role of chromosomes in evolution: a new interpretation. Ann Genet. 1986;29:69–75. in French. [PubMed] [Google Scholar]
- 113.Pardo-Manuel de Villena F, Sapienza C. Recombination is proportional to the number of chromosome arms in mammals. Mamm Genome. 2001;12:318–322. doi: 10.1007/s003350020005. [DOI] [PubMed] [Google Scholar]
- 114.Fledel-Alon A, et al. Broad-scale recombination patterns underlying proper disjunction in humans. PLoS Genet. 2009;5:e1000658. doi: 10.1371/journal.pgen.1000658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Dumas D, Britton-Davidian J. Chromosomal rearrangements and evolution of recombination: comparison of chiasma distribution patterns in standard and robertsonian populations of the house mouse. Genetics. 2002;162:1355–1366. doi: 10.1093/genetics/162.3.1355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Ptak SE, et al. Fine-scale recombination patterns differ between chimpanzees and humans. Nature Genet. 2005;37:429–434. doi: 10.1038/ng1529. [DOI] [PubMed] [Google Scholar]
- 117.Winckler W, et al. Comparison of fine-scale recombination rates in humans and chimpanzees. Science. 2005;308:107–111. doi: 10.1126/science.1105322. References 116 and 117 demonstrate the rapid evolution of hot spots among primate species. [DOI] [PubMed] [Google Scholar]
- 118.Dumont BL, Payseur BA. Evolution of the genomic rate of recombination in mammals. Evolution. 2008;62:276–294. doi: 10.1111/j.1558-5646.2007.00278.x. [DOI] [PubMed] [Google Scholar]
- 119.Allers T, Lichten M. Differential timing and control of noncrossover and crossover recombination during meiosis. Cell. 2001;106:47–57. doi: 10.1016/s0092-8674(01)00416-0. [DOI] [PubMed] [Google Scholar]
- 120.Schwacha A, Kleckner N. Identification of joint molecules that form frequently between homologs but rarely between sister chromatids during yeast meiosis. Cell. 1994;76:51–63. doi: 10.1016/0092-8674(94)90172-4. [DOI] [PubMed] [Google Scholar]
- 121.Bishop DK. RecA homologs Dmc1 and Rad51 interact to form multiple nuclear complexes prior to meiotic chromosome synapsis. Cell. 1994;79:1081–1092. doi: 10.1016/0092-8674(94)90038-8. [DOI] [PubMed] [Google Scholar]
- 122.Schwacha A, Kleckner N. Interhomolog bias during meiotic recombination: meiotic functions promote a highly differentiated interhomolog-only pathway. Cell. 1997;90:1123–1135. doi: 10.1016/s0092-8674(00)80378-5. [DOI] [PubMed] [Google Scholar]
- 123.Thompson DA, Stahl FW. Genetic control of recombination partner preference in yeast meiosis. Isolation and characterization of mutants elevated for meiotic unequal sister-chromatid recombination. Genetics. 1999;153:621–641. doi: 10.1093/genetics/153.2.621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Niu H, et al. Mek1 kinase is regulated to suppress double-strand break repair between sister chromatids during budding yeast meiosis. Mol Cell Biol. 2007;27:5456–5467. doi: 10.1128/MCB.00416-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Niu H, et al. Regulation of meiotic recombination via Mek1-mediated Rad54 phosphorylation. Mol Cell. 2009;36:393–404. doi: 10.1016/j.molcel.2009.09.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Kon N, Krawchuk MD, Warren BG, Smith GR, Wahls WP. Transcription factor Mts1/Mts2 (Atf1/Pcr1, Gad7/Pcr1) activates the M26 meiotic recombination hotspot in Schizosaccharomyces pombe. Proc Natl Acad Sci U S A. 1997;94:13765–13770. doi: 10.1073/pnas.94.25.13765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Hirota K, Mizuno K, Shibata T, Ohta K. Distinct chromatin modulators regulate the formation of accessible and repressive chromatin at the fission yeast recombination hotspot ade6-M26. Mol Biol Cell. 2008;19:1162–1173. doi: 10.1091/mbc.E07-04-0377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Pryce DW, McFarlane RJ. The meiotic recombination hotspots of Schizosaccharomyces pombe. Genome Dyn. 2009;5:1–13. doi: 10.1159/000166614. [DOI] [PubMed] [Google Scholar]
- 129.White MA, Dominska M, Petes TD. Transcription factors are required for the meiotic recombination hotspot at the HIS4 locus in Saccharomyces cerevisiae. Proc Natl Acad Sci U S A. 1993;90:6621–6625. doi: 10.1073/pnas.90.14.6621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Merker JD, et al. The histone methylase Set2p and the histone deacetylase Rpd3p repress meiotic recombination at the HIS4 meiotic recombination hotspot in Saccharomyces cerevisiae. DNA Repair (Amst) 2008;7:1298–1308. doi: 10.1016/j.dnarep.2008.04.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Gottlieb S, Esposito RE. A new role for a yeast transcriptional silencer gene, SIR2, in regulation of recombination in ribosomal DNA. Cell. 1989;56:771–776. doi: 10.1016/0092-8674(89)90681-8. [DOI] [PubMed] [Google Scholar]
- 132.Heng HH, et al. Regulation of meiotic chromatin loop size by chromosomal position. Proc Natl Acad Sci U S A. 1996;93:2795–2800. doi: 10.1073/pnas.93.7.2795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Moens PB, et al. Rad51 immunocytology in rat and mouse spermatocytes and oocytes. Chromosoma. 1997;106:207–215. doi: 10.1007/s004120050241. [DOI] [PubMed] [Google Scholar]
- 134.Anderson LK, Hooker KD, Stack SM. The distribution of early recombination nodules on zygotene bivalents from plants. Genetics. 2001;159:1259–1269. doi: 10.1093/genetics/159.3.1259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Anderson LK, Reeves A, Webb LM, Ashley T. Distribution of crossing over on mouse synaptonemal complexes using immunofluorescent localization of MLH1 protein. Genetics. 1999;151:1569–1579. doi: 10.1093/genetics/151.4.1569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Foss E, Lande R, Stahl FW, Steinberg CM. Chiasma interference as a function of genetic distance. Genetics. 1993;133:681–691. doi: 10.1093/genetics/133.3.681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Foss EJ, Stahl FW. A test of a counting model for chiasma interference. Genetics. 1995;139:1201–1209. doi: 10.1093/genetics/139.3.1201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.McPeek MS, Speed TP. Modeling interference in genetic recombination. Genetics. 1995;139:1031–1044. doi: 10.1093/genetics/139.2.1031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Broman KW, Weber JL. Characterization of human crossover interference. Am J Hum Genet. 2000;66:1911–1926. doi: 10.1086/302923. [DOI] [PMC free article] [PubMed] [Google Scholar]