

Abstract
Obesity, diabetes, and related metabolic disorders are major health issues worldwide. As the epidemic of metabolic disorders continues, the associated medical co-morbidities, including the detrimental impact on reproduction, increase as well. Emerging evidence suggests that the effects of maternal nutrition on reproductive outcomes are likely to be mediated, at least in part, by oocyte metabolism. Well-balanced and timed energy metabolism is critical for optimal development of oocytes. To date, much of our understanding of oocyte metabolism comes from the effects of extrinsic nutrients on oocyte maturation. In contrast, intrinsic regulation of oocyte development by metabolic enzymes, intracellular mediators, and transport systems is less characterized. Specifically, decreased acid transport proteins levels, increased glucose/lipid content and elevated reactive oxygen species in oocytes have been implicated in meiotic defects, organelle dysfunction and epigenetic alteration. Therefore, metabolic disturbances in oocytes may contribute to the diminished reproductive potential experienced by women with metabolic disorders. In-depth research is needed to further explore the underlying mechanisms. This review also discusses several approaches for metabolic analysis. Metabolomic profiling of oocytes, the surrounding granulosa cells, and follicular fluid will uncover the metabolic networks regulating oocyte development, potentially leading to the identification of oocyte quality markers and prevention of reproductive disease and poor outcomes in offspring.
Keywords: Germ cell, Glucose, Lipid, Amino acid, Biomarker
Introduction
Mammalian oocyte development is a complex process regulated by a vast number of intra- and extra-ovarian factors. Oocytes originate from primordial germ cells (PGCs). PGCs migrate to the genital ridge where they proliferate by mitosis and transform from oogonia into primary oocytes [1–3]. Around the time of birth, oocytes are arrested within an ovarian follicle at the diplotene stage of the first meiotic prophase, which is also termed the germinal vesicle (GV) stage. After puberty, oocytes are stimulated to reinitiate meiosis after an endogenous LH surge, as indicated by GV breakdown (GVBD). As the microtubules become organized into a bipolar spindle and all chromosomes align at the spindle equator, the oocytes proceed to metaphase I (MI) and then extrude the first polar body. Oocyte maturation is complete upon arrest at metaphase II (MII), waiting for fertilization [4, 5]. Oocyte growth and maturation are believed to be particularly sensitive to changes in nutritional, chemical, and endocrine environments [6].
Full developmental competence of an oocyte requires synchronous nuclear maturation and cytoplasmic maturation [7, 8]. Indicative of the requirement for energy production and active synthesis during this process, organelles in the ooplasm such as mitochondria, endoplasmic reticulum, and Golgi complexes become more abundant and experience dynamic changes with oocyte growth [9, 10]. A variety of metabolites and metabolism-related enzymes have been demonstrated to play critical roles in multiple cellular events occurring in oocyte maturation [11–14]. The growing mammalian follicle consists of a single oocyte, one or more layers of surrounding somatic cells (granulosa cells), and an outer rim of theca cells. Oocytes are coupled to granulosa cells by gap junctions. These highly specialized membrane connections mediate the transfer of small metabolites, inorganic ions, and second messengers from one cell to another [15]. Metabolic cooperation between oocytes and granulosa cells in the follicle has long been appreciated [16–18]. Granulosa cells provide various energy substrates for nurturing oocytes, while oocytes control metabolic activity in granulosa cells by secretion of paracrine factors [19, 20].
Obesity, diabetes, and related metabolic disorders are major health issues worldwide. As this epidemic of metabolic disorders continues, the associated medical co-morbidities, including those affecting reproduction, increase as well [21]. In particular, women with obesity or poorly controlled diabetes have an increased risk of infertility, miscarriage, obstetric complications, neonatal morbidity and mortality, and birth defects in their offspring. Emerging animal studies and clinical data suggest that metabolic activity in oocytes may be a critical target mediating these reproductive defects.
This review focuses on the recent advances made in the investigation of the metabolic events governing oocyte growth and maturation and the link between maternal nutrition, oocyte metabolism, and reproductive outcomes. In addition, metabolic analysis of oocytes is briefly introduced. Advances in these areas are likely to contribute to the understanding of the mechanisms underlying oocyte development, and as a result, may guide further approaches for the identification of predictors of oocyte quality and for treatment of infertility and poor obstetric and neonatal outcomes. The metabolism during folliculogenesis is not discussed here as it is covered in other recent reviews [22, 23].
Methods
This article provides a comprehensive review with references from both human and animal studies. Relevant research articles published in English up to August 2014 were obtained from PubMed using the following search terms in various combinations: ‘oocyte’, ‘oocyte quality’, ‘metabolism’, ‘metabolic disorder’, ‘maternal nutrition’, ‘obese’, ‘diabetes’, ‘PCOS’, ‘metabolomics’, ‘follicular fluid’, ‘cumulus cell’, ‘embryo’, ‘reproductive outcome’, ‘pregnancy,’ and ‘epigenetics’. Abstracts were evaluated to identify studies appropriate to the review.
Energy metabolism in oocytes
Oocytes cannot be fully understood in isolation from their follicles (specifically granulosa cells), which create an essential niche for oocyte survival, nourishment, and regulation [24]. Oocytes are deficient in their ability to carry out some key metabolic processes and thus depend on the cooperation of granulosa cells to provide metabolites for their development [19, 25]. Despite the oocytes’ commanding role, communication is bidirectional, and follicular cells are interdependent; for detailed reviews see [20, 26].
As Henry Leese noted [27], metabolic control of oocyte development may be considered at two levels: (1) intrinsic control by enzyme amount/activities, intracellular mediators, and plasma membrane transport systems [28–30] and (2) extrinsic control by providing nutrients derived from the follicular niche, female reproductive tract, or in vitro culture environment [31–34]. Overwhelmingly, the focus of research and practice in this area has been on extrinsic nutrients regulating oocyte maturation. In contrast, the potential contribution of intrinsic pathways controlling oocyte development has received little attention. Some intrinsic enzymes related to metabolism have been examined in mammalian oocytes and are summarized in Table 1. Metabolic control of oocyte development, specifically meiosis, epigenetics and organelles, are outlined in Fig. 1.
Table 1.
Summary of the intrinsic enzymes related to metabolism in mammalian oocytes
| Enzyme | Oocyte/species | Functions | References |
|---|---|---|---|
| Carbohydrate metabolism-related enzymes | |||
| HK | Human; mouse; rat | Detectable activity, no functional analysis | [42, 66, 265, 266] |
| GPI | Human; mouse | GPI transcript was detectable; Gpi1−/− null mouse oocytes show normal function | [65, 68, 265, 267] |
| PFK | Human; mouse; rat; bovine | Detectable activity, may be associated with oocyte maturation | [66, 94, 265, 268] |
| G6PDH | Human; mouse; rat; bovine; pig; sheep | Activity is associated with meiotic progression, mitochondrial features and developmental competence of oocytes | [66, 269–274] |
| GAPDH | Mouse | Detectable activity, no functional analysis | [65] |
| GS | Human; Mouse | Detectable activity, no functional analysis | [64, 274] |
| GSK3 | Mouse; Bovine | GSK3b regulates mouse oocyte meiosis, particularly chromosome segregation | [275–277] |
| PK | Human; mouse | Detectable activity, no functional analysis | [64] |
| PDH | Mouse | Pdha1−/− null mouse oocytes are unable to properly complete oogenesis | [12] |
| ADK | Human; mouse | Detectable activity, no functional analysis | [169] |
| T-synthase | Mouse | Mouse fertility is enhanced by oocyte-specific loss of T-synthase | [278] |
| SDH | Rat | Detectable activity, no functional analysis | [279, 280] |
| AR | Mouse; rat | Detectable activity, and may be important for mouse oocyte meiotic resumption | [279–281] |
| CS | Human; mouse | Detectable activity, no functional analysis | [169] |
| MDH | Human; mouse; rat; bovine | Activity increases during oocyte maturation, no functional analysis | [64, 66, 282] |
| NADP-IDH | Bovine | Detectable activity, no functional analysis | [282] |
| LDH | Mouse; pig; cattle | Detectable activity, no functional analysis | [65, 283–286] |
| Lipid metabolism-related enzymes | |||
| Lipase | Bovine | Detectable activity, no functional analysis | [94] |
| GK | Bovine | Transcript was detectable in oocytes, no functional analysis | [287] |
| CPT | Mouse | Essential for oocyte developmental competence and early embryo development | [93, 288] |
| BOAC | Human; mouse | Detectable activity, no functional analysis | [64, 169] |
| ACADL | Pig | Detectable activity, no functional analysis | [286] |
| CT | Human; mouse | Detectable activity, no functional analysis | [64] |
| AC | Human; mouse | Improves the quality of oocytes and embryos and the outcome of IVF | [289, 290] |
| Amino acid metabolism-related enzymes | |||
| GLDH | Human; mouse | [64] | |
| ALT | Human; bovine | Enzymatic units remained constant in oocytes during IVM | [274, 282] |
| AST | Human; mouse; bovine | Enzymatic units remained constant in oocytes during IVM | [64, 274, 282] |
| GPT2 | Mouse | Detectable activity, no functional analysis | [169] |
Unless stated otherwise, the oocytes mentioned above are fully-grown oocytes
HK hexokinase, GPI glucose-6-phosphate isomerase, PFK phosphofructokinase, G6PDH glucose-6-phosphate dehydrogenase, GAPDH glyceraldehyde-3-phosphate dehydrogenase, GS glycogen synthase, GSK3 glycogen synthase kinase 3, PK pyruvate kinase, PDH pyruvate dehydrogenase, ADK adenylate kinase, T-synthase core 1 beta1,3-galactosyltransferase, SDH sorbitol dehydrogenase, AR aldose reductase, CS citrate synthase, MDH malate dehydrogenase, NADP-IDH NADP-dependent isocitrate dehydrogenase, LDH lactate dehydrogenase, GK glycerol kinase, CPT carnitine palmitoyl transferase, BOAC beta-hydroxyacyl-CoA dehydrogenase, ACADL long-chain acyl-CoA dehydrogenase, CT cytoplasmic thiolase, AC acid ceramidase, GLDH glutamate dehydrogenase, ALT alanine aminotransferase, AST aspartate aminotransferase, GPT2 glutamic pyruvate transaminase 2, IVF in vitro fertilization, IVM in vitro maturation
Fig. 1.
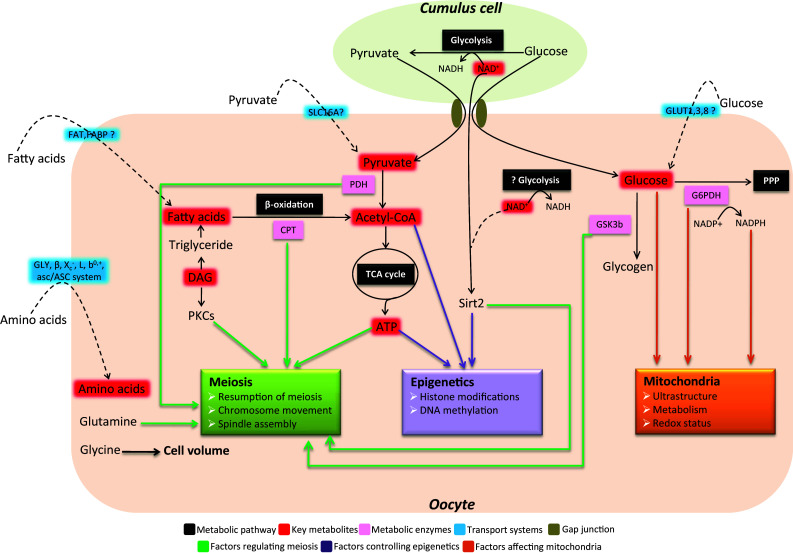
Metabolic control of meiosis, epigenetics and organelles during oocyte development
Glucose metabolism in oocytes
Glucose utilization by cumulus-oocyte complexes (COCs) has been systematically reviewed by Sutton-McDowall et al. [35, 36]. Hence, only a specific discussion on metabolic patterns, transport systems, and critical enzymes in oocytes is presented here.
Metabolic patterns of glucose in oocytes Energy metabolism in primordial germ cells (PGCs) and oogonia remains controversial. Mouse PGCs were originally shown to preferentially oxidize pyruvate over glucose [37]. However, work by Jansen and de Boer [38] suggested that anaerobic glycolysis is the predominant metabolic pattern in PGCs to avoid oxidative damage. Throughout the growth period, pyruvate and oxygen consumption is gradually increased in mouse oocytes [39]. The fully grown oocytes have a limited capacity to utilize glucose [40–43], and glucose first needs to be converted to pyruvate by follicular cells to ensure successful meiotic maturation [18, 44–46]. Pyruvate is transported by the proton-linked monocarboxylate carrier family (MCT) [47], and mouse oocytes are rich in the SLC16A members of MCT [48] (Fig. 1). Nonetheless, important questions about which MCT(s) and how they mediate the pyruvate uptake by oocytes remain unanswered. Highlighting the importance of pyruvate during oocyte maturation, mouse oocyte-specific deletion of Pdhα1 (pyruvate dehydrogenase E1 α) caused severe meiotic defects [12] (Fig. 1; Table 1). Interestingly, dog and pig oocytes seem to utilize glucose as the primary energy substrate at a much higher rate than that of other species [32, 49], implying the species-specific differences in metabolic pattern of glucose in oocytes.
Glucose transport in oocytes Although pyruvate is the preferred substrate, the machinery for glucose metabolism and glucose carrier-mediated uptake have been detected in oocytes [41, 42, 50–54]. Glucose enters cells either actively via sodium-coupled glucose transporters (SGLTs) or through facilitative glucose transporters (GLUTs). Fourteen members of the GLUT family have been identified, including GLUT1–12, the H+-coupled myoinositol transporter, and GLUT14 [29, 55]. GLUTs expression has been reported in mouse [56, 57], cow [58], sheep [59], human [60], and rhesus monkey oocytes [61]. The mouse oocyte contains GLUT1, 3 and 8, but GLUT4 (the primary insulin-sensitive transporter) has not been detected [36, 62] (Fig. 1). Up to now, scarce research has been conducted into the transport mechanisms of glucose in oocytes. Recently, using a non-metabolizable fluorescent glucose derivative, we have revealed an intercellular pathway for glucose transport into mouse oocytes, whereby glucose is taken up in cumulus cells and then transferred into the oocyte through gap junctions [63] (Fig. 1).
Glucose metabolism-related enzymes in oocytes In attempting to discover which metabolic pathways are functional in oocytes, it has been necessary to measure the levels and/or activities of related enzymes. Hexokinase (HK), P-fructokinase (PFK), and pyruvate kinase (PK) were measured in single mouse and human oocytes. Although all of them were detectable, the human levels averaged 3–9 times higher than those in the mouse [64], indicating that they may play species-specific roles. Other enzymes related to glucose metabolism and the tricarboxylic acid cycle (TCA) have also been measured in oocytes from different species using either biochemical or cytochemical methods [65–67]. Studies on the functions of most glucose metabolism-related enzymes during oocyte development have not been carried out (Table 1).
A female mouse chimera composed of wild-type cells and homozygous Gpi −/− null cells (Gpi encodes the glycolytic enzyme glucose phosphate isomerase) was able to produce offspring derived from homozygous Gpi −/− null oocytes [68], supporting the conclusion that mouse oocytes do not require intact endogenous glycolysis for their development. Glucose-6-phosphate dehydrogenase (G6PDH) is a cytosolic enzyme that catalyzes the first step in the pentose phosphate pathway (PPP) and is synthesized within the oocyte during oogenesis (Fig. 1). Since G6PDH can convert the brilliant cresyl blue (BCB) stain from blue to colorless, BCB staining was used to measure G6PDH activity in oocytes. G6PDH has very high activity in the preovulatory oocytes of mice, rats, and cows and experiences dynamic changes during oocyte maturation [65–67]. Numerous reports have shown that low G6PDH activity is associated with a high rate of fertilization and blastocyst formation [69–71], indicating that G6PDH activity may be a good indicator of oocyte quality. Nonetheless, BCB staining has not been used to evaluate the G6PDH activity in human oocytes yet. Moreover, most recently, BCB staining of pig oocytes at the immature stage was found to compromise mitochondrial functions throughout oocyte maturation and, therefore, is not suitable for application in a standard in vitro maturation protocol [72, 73]. Therefore, BCB staining-based oocyte quality evaluation still remains an open issue.
Lipid metabolism in oocytes
Lipid stores in oocytes Intracellular lipid levels within mammalian oocytes vary dramatically among species (4 ng/mouse oocyte, 63 ng/cow oocyte and 161 ng/pig oocyte) [74–76]. Triglycerides are the most abundant lipid in oocytes, constituting over 50 % of all lipid material [77–79] and provide a large potential energy reserve. Lipid droplets accumulate during oocyte growth and undergo temporal and spatial changes during maturation. For example, triglyceride stores sharply decrease over the course of pig and cow oocyte maturation in vitro, despite culturing in the presence of serum [76, 77]. Lipid droplets become larger and more centrally located as the mouse oocyte matures [80]. By contrast, a pronounced peripheral distribution pattern was observed in hamster and pig oocytes [81–83]. It was reported that perlipin-2 and adipose differentiation-related protein (ADRP) surround lipid droplets in mouse and cow oocytes [80, 84], which may control the stabilization of lipid droplets and lipolysis for utilization [85]. Furthermore, using a live cell-imaging method, Sturmey et al. showed the close spatial association between lipid droplets and mitochondria in pig and bovine oocytes [83, 86, 87], indicating the importance of oocyte mitochondria in lipid metabolism.
β-Oxidation of fatty acid in oocytes Lipase enzymes cleave triglycerides into a glycerol backbone and fatty acids with different chain lengths and saturation degrees. Fatty acid uptake is primarily via fatty acid protein transporters on the cell surface, including fatty acid translocase (FAT), tissue-specific fatty acid transport proteins (FATP), and plasma membrane bound fatty acid binding protein (FABPpm) (Fig. 1). Once inside the cell, fatty acids are transported across the outer mitochondrial membrane by carnitine-palmitoyl transferase (CPT) and then couriered across the inner mitochondrial membrane by carnitine. In mitochondria, the long chain acyl-CoA enters the fatty acid β-oxidation pathway, releasing acetyl-CoA. This acetyl-CoA then enters the TCA cycle to produce ATP. CPT1 is believed to be the rate-limiting step in fatty acid oxidation [88] (Fig. 1). In comparison with other macronutrient classes (carbohydrates and protein), fatty acids yield the most ATP by the β-oxidation pathway.
The role of fatty acids during oocyte development in different animal models has been excellently reviewed by Dunning et al. [89] and Mckeegan and Sturmey [90]; therefore, only a brief account of β-oxidation is given here. Palmitic, stearic, oleic, and linoleic acids are the most abundant intracellular fatty acids in pig, cattle, and human oocytes [78, 79, 84]. By employing inhibitors of CPT1, it has been shown that inhibiting β-oxidation of fatty acids during cow and pig oocyte maturation led to a fall in viability of subsequent embryos [76, 91]. Interestingly, although with low lipid reserves, inhibition of β-oxidation during mouse oocyte maturation blocked AMPK-mediated meiotic resumption [92]. In addition, lipase activity during bovine oocyte maturation and CPT1 expression in mouse oocytes following hormone superovulation were found to be up-regulated [93, 94]. Recent studies have shown that supplementation of l-carnitine (a cofactor of CPT1) to cultured follicles enhanced β-oxidation and improved both oocyte nuclear and cytoplasmic maturation in mice and cows [95–97]. Similarly, l-carnitine enrichment of in vitro maturation (IVM) medium increased the concentration of glutathione (an antioxidant) and promoted pig oocyte development [98]. The importance of fatty acid β-oxidation in nuclear maturation has also been demonstrated in mice, cows, and pigs [99]. These findings collectively suggest that metabolism of fatty acids, specifically β-oxidation pathway, is essential for maintaining developmental competence of mammalian oocytes.
Fatty acids in cellular signaling transduction In addition to the energy supply for cells, fatty acids and their precursors/metabolites have also been reported to be required for the non-metabolic processes, particularly cellular signaling transduction [100, 101]. Diacylglycerol (DAG), an intermediate in glycerolipid metabolism, is historically the first known lipid second messenger [102–104]. DAG is generated by the lipid hydrolysis of phosphatidylinositol (4,5)-bisphosphate, and protein kinase C (PKC) is its major effector. The binding of DAG to a conserved C1 domain in PKC leads to PKC activation, modulating the cell cycle, cellular survival, malignant transformation, and apoptosis [105]. It is worth noting that PKC isozymes have been implicated in multiple key steps during oocyte development, such as meiotic resumption, spindle organization, and activation [106–108] (Fig. 1). Ceramide, the immediate lipid product of sphingomyelin hydrolysis, was demonstrated to behave as a bioeffector for meiotic cycle progression in Xenopus oocytes [109, 110]. Fatty acids also bind nuclear receptors and transcription factors, such as peroxisome-proliferator activated receptors (PPAR) and sterol-regulatory element binding protein (SREBP) [111]. PPAR and SREBP1 have been detected in mouse and bovine oocytes and may be associated with embryo development and female fertility [112, 113]. Although promising, the role of lipids serving as secondary messengers in the control of mammalian oocyte development is currently an almost unexplored research area.
Amino acid metabolism in oocytes
Amino acids are transported into cells by an array of specialized systems and play important roles in cellular function, including protein synthesis, energy production, organic osmolytes, and intracellular buffer [114–116]. In the past few decades, amino acids in preimplantation embryos have been extensively investigated. In contrast, much less is known about amino acid transport and utilization during mammalian oocyte growth and development.
Transport systems of amino acids in oocytes Amino acid transport systems were defined classically by the range of their substrate specificities and the presence or absence of obligate Na+ cotransport [117, 118]. Colonna et al. first examined the amino acid transport systems in isolated mouse oocytes throughout oogenesis. While fully grown GV oocytes were lacking the A-transport system, they demonstrated utilization of the l- and ASC-systems [119]. Van Winkle et al. later showed that glycine transport was mainly via system GLY and cysteine/glutamate transport was via system X−c in immature oocytes [120, 121]. Similarly, system β, L, GLY, X−c, and b0,+ were successively detected in matured oocytes [121–124]. Recently, Pelland et al. [117] measured the transport characteristics of nine amino acids and determined the activity of 11 classically defined amino acid transport systems in mouse oocytes. GLY, β, and X−c had low activity in growing oocytes and were strongly up-regulated during meiotic maturation. L, b0,+, and asc/ASC seemed to be constitutively active throughout oocyte growth and maturation. Consistent with this, oocytes exhibited distinct patterns for amino acid transport [117].
It has been reported that when mouse cumulus cell-enclosed- or cumulus cell-denuded oocytes were cultured in medium containing [14C] l-alanine, the amount of radioactivity in oocytes was higher in cumulus cell-enclosed group [125]. Slc38a3 (a transcript encoding a sodium-coupled neutral amino acid transporter that has high substrate preference for alanine) was abundantly expressed in cumulus cells as compared to oocytes in mice [126]. To date, the enhanced transport rates of glycine, alanine, taurine, and lysine into oocytes have been demonstrated in the presence of granulosa cells [117, 120, 125]. These findings strongly imply that metabolic cooperation between oocytes and follicular cells involves the uptake of amino acids. Surprisingly, follicular cells did not confer additional amino acid transport capabilities on the enclosed oocyte, and in some cases, they appeared to inhibit the uptake of leucine by oocytes in mice [117].
Utilization of amino acid in oocytes The presence of numerous transport systems in oocytes indicates their capacity to utilize amino acids from the external environment. Glutamine has been suggested to be an efficient energy substrate to support oocyte development. For example, using glutamine as a sole energy source is sufficient to initiate meiotic resumption of mouse oocytes within COC, despite the inability to progress through the MII stage [127]. Addition of glutamine to the culture medium promoted oocyte maturation in cow, hamster, dog, rabbit, and rhesus monkey [115, 128–131] (Fig. 1). Glutamine, aspartate and valine also can prevent polyspermic fertilization of pig oocytes [132].
Animal cells control their volume by actively adjusting the cytoplasmic concentrations of osmotically active solutes, thus altering intracellular osmotic pressure [133]. Tartia et al. [134] have done elegant work to examine volume regulation in mouse oocytes. They found that GLYT1, a glycine transporter, is quiescent in immature GV oocytes, which also contain very little endogenous glycine. However, within several hours of the initiation of ovulation, GLYT1-mediated glycine transport becomes activated in oocytes. Concomitantly, oocytes become able to use glycine to regulate their volume. These observations indicate that the glycine-dependent cell-volume-regulatory mechanism exists in oocytes (Fig. 1).
Oocyte metabolism, maternal nutrition, and reproductive outcomes: connecting the dots
Evidence is growing that excessive body fat has a detrimental effect on female fertility and pregnancy. Obese women take longer to conceive, even if cycling regularly, and have a high risk of miscarriage, preeclampsia, gestational diabetes, and congenital defects in the offspring [135–137]. Likewise, women with poorly controlled diabetes often suffer from a series of reproductive problems, such as miscarriage, neonatal morbidity and mortality, and congenital malformations [138–140]. Moreover, despite the glycemic control throughout pregnancy, these women still experience a three to fivefold higher incidence of the pregnancy complications [141], indicating that maternal diabetes may have permanent and irreversible effects on female reproduction. Meanwhile, animal studies also have demonstrated that deficiencies or excesses in a range of macro- and micronutrients are associated with significant impairments in reproductive performance, fertility, fetal development, and long-term offspring health [142–144].
Maternal nutrition and reproduction
In the past 30 years, most studies examining the effects of maternal nutrition on reproduction in humans and animal models have been focused on early embryo and offspring development. We will not include a discussion of metabolic fuel for female reproduction and the effects on fetus/offspring, which have been the subject of other recent reviews [142, 145–147].
Maternal hyperglycemia and obesity have been shown to adversely affect progression from a one-cell zygote to a blastocyst in rodent models [148–153]. Recently, we found that a one-cell zygote transfer from diabetic to non-diabetic mice still results in significantly increased congenital malformations and growth retardation in the offspring [154]. Similar phenotypes were also observed in mice fed a high-fat diet (HFD) [153]. These findings indicate that exposure to disturbed metabolic conditions during oogenesis and fertilization was enough to permanently program the fetus to develop morphological changes.
Data from human clinics have shown that implantation and pregnancy rates are [155, 156] lower in obese women than those of normal weight [157, 158]. Oocyte donation models have been used to determine whether oocyte/embryo quality or uterine receptivity is the main cause of decreased pregnancy rates. There are several systematic reviews illustrating this topic, and conflicting results were produced [159–165]. Nonetheless, the majority of retrospective studies imply that body mass index (BMI) is less likely to negatively affect endometrial receptivity, and it is more likely that poor-quality oocytes or resulting embryos are the cause of reproductive problems experienced by overweight women [161, 162]. Most recently, a very large analysis of 45,000 assisted reproductive technology (ART) cycles concluded that increasing obesity correlated with decreasing likelihood of achieving pregnancy. This effect was only observed when autologous oocytes were used. If donor oocytes from a lean woman were used, the effect was mitigated [166], supporting the idea that the source of the oocyte determines the reproductive outcome.
Together, it is attractive to propose that maternal metabolic disorders have detrimental effects as early as the oocyte stage, which further predisposes them to embryo developmental abnormalities and even metabolic diseases in the offspring. Direct causative relationships between maternal nutrition, oocyte quality, and reproductive outcomes have not been unequivocally established. Nevertheless, evidence for such relationships continues to solidify. Herein, we will summarize the data from both experimental animal models and infertility clinics related to the effects of maternal nutrition on oocyte development, with a specific perspective on oocyte metabolism.
Nutritional effects on oocyte metabolism
Upon leaving the primordial resting pool and during the follicular growth phase, oocytes are particularly sensitive to changes in the follicular environment, especially those stressors induced by nutrient changes [167]. The follicular environment is a reflection of the physiologic status of the animal. Thus, variance in maternal nutrition is likely to have a significant effect on the metabolic activity of oocytes. As outlined above, inappropriate metabolism of oocytes inevitably compromises oocyte quality and the resultant embryo development.
Diabetes mellitus is a metabolic condition characterized by elevated blood glucose levels. Using a type I diabetic mouse model, we found that maternal diabetes markedly results in structural and spatial alterations of mitochondria in oocytes [168]. Consistent with this, the levels of ATP and TCA cycle metabolites including citrate, malate, and aspartate are decreased in preovulatory and ovulated oocytes from diabetic mice [155, 156, 168, 169]. Similar phenomenon was also seen in HFD mice and mice with hyperinsulinemia [153, 170]. Variations in the ATP content have been suggested to affect oocyte quality, embryonic development, and the implantation process [171, 172]. Specifically, microtubule assembly and chromosome movement during meiosis require ATP [173]. Injury of mitochondria in mouse oocytes could decrease ATP content and disrupt the meiotic spindle [174]. Pdhα1-deficient mouse oocytes show inadequate ATP levels along with chromatin and microtubular abnormalities [12]. Oocytes from senescence-accelerated mice demonstrated spindle defects and disturbances in chromosome alignment associated with mitochondrial dysfunction [175]. Recently, association of mitochondria with spindle poles was found to facilitate spindle alignment in Schizosaccharomyces pombe [176]. Furthermore, low mtDNA and ATP content also may contribute to the absence of birefringent spindles imaged with the PolScope in human oocytes [177]. Hence, it is hypothesized that disrupted mitochondrial metabolism may be associated with the spindle defects and chromosome misalignment observed in oocytes from diabetic, obese, and insulin-resistant mice.
Formation of reactive oxygen species (ROS) is a by-product of oxidative phosphorylation in the mitochondria [178, 179]. Igosheva et al. [180] reported increased NAD(P)H content and reduced FAD autofluorescence in oocytes from obese mice, indicating an elevation of ROS generation. Wakefield et al. [181] found that feeding of a diet enriched with long-chain n−3 polyunsaturated fatty acids (PUFA) leads to mitochondrial dysfunction and elevated ROS levels in mouse oocytes, with reduced fertilization capacity and blastocyst formation. These data suggest that excess maternal nutrition could adversely impact mitochondrial status in oocytes, particularly energy production and oxidative stress, and ultimately disrupt oocyte metabolism.
Polycystic ovary syndrome (PCOS) is also associated with metabolic disturbance. Women with PCOS and hyperandrogenism are typically characterized by increased number of oocytes retrieved during in vitro fertilization (IVF). However, these oocytes are often of poor quality, leading to lower fertilization/implantation rates and a higher miscarriage rate [182]. By feeding mice chow supplemented with dehydroepiandrosterone (DHEA), we used a hyperandrogen model to show that citrate levels, G6PDH activity, and lipid content in oocytes from DHEA-exposed mice were significantly lower than those in controls [183], suggesting abnormal TCA and PPP metabolism. Inconsistent with the results derived from animal models, oocytes from PCOS patients show elevated glucose/pyruvate consumption and lower intracellular NAD(P)H content compared to oocytes from control patients [184]. This discrepancy may be explained by the following: (1) although various animal models have been shown to mimic key phenotypes of women with obesity or PCOS, no one model provides complete replication of the complex clinical disorders [185–187]; or (2) the data from human oocytes are sometimes variable because of the limited number of control oocytes from young and healthy women.
HFD mice have higher levels of lipid within oocytes of preovulatory COCs [188–190]. Likewise, both an in vivo hyperglycemic environment and an in vitro high-glucose culture increase free glucose levels in mouse oocytes [63]. Glycogen content is also higher in immature oocytes of diabetic mice, whereas fructose-1, 6-phosphate (FBP) levels are elevated with the resumption of meiosis [169], indicating that a hyperglycemic environment leads to accumulation of glucose stored as glycogen before maturation in oocytes. In contrast, the activity of two enzymes that are major representatives of fatty acid metabolism (hydroxyacyl-CoA dehydrogenase; Hadh2) and amino acid metabolism (glutamic pyruvate transaminase; Gpt2) are lower in diabetic oocytes than controls. In addition, diabetic rat models showed altered prostaglandin (PGE2) production in both ovulated and immature COCs isolated from ovaries [191, 192]. Some of these conditions are thought to contribute to the disrupted meiotic behavior in oocytes from diabetic animals.
Ruminant livestock species are also important models to reveal the significance of periconceptional nutritional status and its effects on reproductive performance. In dairy heifers, it was shown that a high-starch diet had negative effects on oocyte developmental competence, and these were avoided when leucine intake was increased [193]. Crude protein overfeeding also compromises the subsequent capacity of bovine oocytes to develop to blastocysts, probably via direct toxic effects of elevated ammonia and urea concentration in the follicular environment [194, 195]. Nevertheless, the nutritional effects on oocyte metabolism in ruminant species remain poorly evaluated. The relationships between metabolic disturbance and follicle/embryo metabolism of these animals have been extensively reviewed [142, 196, 197].
Potential mechanisms: oocyte metabolism as a link between maternal nutrition and reproductive outcomes
The effects of the metabolic condition on oocyte metabolism and development are summarized in Table 2. A schematic diagram for the proposed pathways through which maternal nutrition exerts its effects on oocyte metabolism and reproductive outcomes is shown in Fig. 2.
Table 2.
Effects of maternal nutritional status on oocyte metabolism and development
| Maternal nutrition | Affected organelles or events in oocytes | Changes in oocyte metabolism | References |
|---|---|---|---|
| Obesity | |||
| Mouse model | Mitochondria | mtDNA content↑; Citrate levels ↓; NADPH levels↑; ROS production↑; Lipid content↑ | [153, 180, 188, 220] |
| Endoplasmic reticulum | |||
| DNA methylation | |||
| Spindle assembly/chromosome alignment | |||
| Women | Spindle assembly/chromosome alignment | Not determined | [214] |
| Diabetes | |||
| Mouse model | Mitochondria |
mtDNA content↑; ATP levels ↓; Citrate levels ↓; Malate levels ↓; Aspartate levels ↓; ROS production↑; Glucose levels↑; Glycogen levels↑; FBP levels↑; Hadh2 activity ↓; GPT2 activity ↓ |
[156, 168, 169, 213, 218, 219, 291] |
| Endoplasmic reticulum | |||
| Meiotic resumption | |||
| Epigenetic modifications | |||
| Spindle assembly/chromosome alignment | |||
| Rat model | Meiotic resumption | Prostaglandin E production in COCs↑ | [192, 292, 293] |
| PCOS | |||
| Hyperandrogenism mouse model | Pentose phosphate pathway metabolism | Citrate levels ↓; Lipid content ↓; G6PDH activity ↓ | [183] |
| Hyperinsulinemia mouse model | Mitochondria | mtDNA content↑; ATP levels ↓; GSH levels↓; GSH/GSSG ratio ↓; ROS production↑ | [170] |
| Spindle assembly/chromosome alignment | |||
| Women | Spindle/chromosome organization |
Glucose consumption↑; Pyruvate consumption↑; NAD(P)H content↑ |
[184, 294] |
| Dietary supply or deficiency of nutrients | |||
| Mouse fed high PUFA diet | Mitochondria | ROS production↑; Expression of SOD2 for scavenging ROS↑; Expression of COX2 for producing prostaglandins ↑ | [181, 295] |
| Epigenetic modifications | |||
| Mouse fed low protein diet | LPD fed exclusively during mouse oocyte maturation leads to behavioral and cardiovascular abnormalities in offspring | Not determined | [296] |
| Pig fed high fiber diet | Oocyte maturity | Not determined | [297, 298] |
| Mouse fed zinc deficient diet | Meiotic resumption | Not determined | [299] |
Unless stated otherwise, the oocytes mentioned above are fully-grown oocytes
mtDNA mitochondrial DNA, NADPH nicotinamide adenine dinucleotide phosphate, ROS reactive oxygen species, FBP fructose-1,6-phosphate, Hadh2 hydroxyacyl-CoA dehydrogenase, GPT2 glutamic pyruvate transaminase 2, COCs cumulus-oocyte complexes, PCOS polycystic ovary syndrome, G6PDH glucose-6-phosphate dehydrogenase, GSH glutathione, GSSG glutathione disulfide, PUFA polyunsaturated fatty acid, SOD2 superoxide dismutase 2, COX2 cyclooxygenase 2, LPD low protein diet
↑ indicates an increase in content; ↓ indicates a decrease in content
Fig. 2.
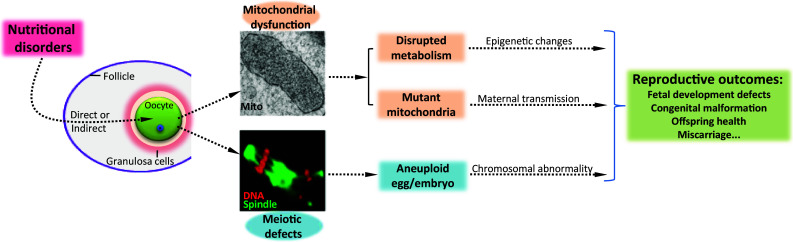
Schematic diagram for the proposed pathways through which maternal nutritional status exerts its effects on oocyte metabolism and, therefore, reproductive outcomes
Metabolic effects on oocyte mitochondria and meiosis Mitochondria are double-membrane organelles composed of an outer membrane, inner membrane, intermembrane space, and matrix. A large number of enzymes involved in different metabolic pathways exist in the mitochondrial matrix, serving as the major site for energy generation in cells. Mitochondria are also characterized by having their own, maternally inherited genome, mitochondrial DNA (mtDNA). As the most abundant and prominent organelle in oocytes, mitochondria undergo stage-specific changes in ultrastructure and distribution during oocyte maturation in response to differing energy requirements [198]. ATP synthesis by oxidative phosphorylation (OXPHOS) is the primary function associated with mitochondria. Most ROS are produced when electrons leak from the mitochondrial respiratory chain. Although mitochondria are major sites of ROS production, excessive ROS can damage mitochondrial functions in oocytes [199]. Mitochondrial function in the oocyte has been extensively reviewed [10, 167, 179, 200].
Follicular fluid is derived from blood in the thecal capillaries. As follicular development progresses, fluid accumulates in the antrum of the follicle, bathing the oocyte and providing the milieu needed for oocyte development [201]. Data from animal models and human clinics have suggested that metabolic disorders are reflected in the microenvironment of pre-ovulatory follicles [182, 202, 203].
By enzymatically measuring intracellular free glucose in individual oocytes, glucose content among oocytes from diabetic mice was shown to be higher than that from control mice [63]. Mitochondrial dysfunction resulting from high glucose has been reported in various cell types [204, 205]. In line with this, altered mitochondrial properties, specifically mitochondrial swelling and increased ROS generation, were observed in diabetic mouse oocytes [168] (Fig. 1). Associated with elevated levels of follicular free fatty acid, obvious lipid accumulation was detected in oocytes from HFD mice [188]. When exposed to a high-lipid environment, cells accumulate triglyceride droplets and free fatty acids, causing significant damages to other cellular organelles, such as mitochondria and endoplasmic reticulum (ER); this process is termed lipotoxicity [206]. Mitochondrial dysfunction and ER stress in oocytes from obese and insulin-resistant mice have been reported by different groups [153, 170, 180, 188, 189, 200]. Collectively, disrupted energy metabolism of oocytes in response to nutritional cues could result in reduced potential for meiotic maturation and fertilization, as well as impaired preimplantation embryonic development. Additionally, these abnormal oocyte mitochondria may be maternally transmitted to the embryo and then be propagated during embryogenesis and fetal development, leading to the reproductive problems experienced by females with metabolic disorders (Fig. 2).
In addition to the direct action of the follicular environment on oocyte metabolism, the deleterious effects on granulosa cells may disrupt trophic and signaling interactions with the oocyte and indirectly contribute to oocyte incompetence and thus poor pregnancy outcomes. Oocytes are deficient in their ability to use glucose as an energy substrate and require cumulus cell-provided glycolytic byproducts for their own development. Pyruvate and ATP, as related products of glycolysis by cumulus cells, can be transferred to oocytes [20, 207]. It is worth noting that abnormal metabolism increased apoptosis, and diminished mitochondrial function are detected in granulosa cells from obese and diabetic mice [156, 188, 208–210]. In particular, glucose uptake is dramatically reduced in cumulus cells of diabetic mice and shows a strong correlation with the lowered ATP levels in the enclosed oocyte [211]. After the resumption of meiosis, oocytes become transcriptionally inactive. At this stage, ATP is only needed to sustain basal metabolism and spindle formation in oocytes [212]. Consistent with this notion, mouse models have revealed that maternal diabetes, obesity, and hyperinsulinemia induce diverse, malformed spindles and misaligned chromosomes in oocytes [153, 168, 170, 213]. Similarly, Machtinger et al. have directly assessed a possible association between severe obesity (class II and III obesity) and oocyte characteristics in women. In severely obese patients, they have shown a high prevalence of cytoskeletal abnormalities in oocytes that failed to fertilize [214]. Any error in the spindle assembly and chromosome movement could result in oocyte chromosomal imbalance, which is the leading genetic cause of miscarriage and congenital birth defects [14, 215, 216]. It is, therefore, possible that the limitation of energy substrates in granulosa cells disrupts the metabolic homeostasis of the oocyte, producing an aneuploid germ cell and contributing to the reproductive problems induced by metabolic disorders.
Metabolic regulation of epigenetics As discussed above, maternal metabolic disorders adversely affect oocyte quality leading to fetal growth retardation and developmental defects, even when removed from the stressful milieu and transferred into a normal uterine environment. Since these fetal abnormalities occur long after the environmental insult, the nutritional effect on the oocyte is likely to be propagated by an epigenetic pathway. Imprinting is an epigenetic mechanism where genes are expressed in a parent of origin specific manner. The chromosomal basis of the epigenetic inheritance of imprinting is methylation of CpG sequences. Imprinting establishment occurs in the germline of the parental generation and maintenance of the parental imprinting commences at the being of embryonic development [217]. Methylation status of several imprinted genes during oocyte development of diabetic mice has been examined. Results showed that the methylation pattern of Peg3 differential methylation regions (DMR) was altered in a time-dependent manner by maternal diabetes, with no significant changes in H19 and Snrpn DMRs [218].
In addition, the differences in acetylation patterns of histone H3 and H4 have also been observed during oocyte maturation in diabetic mice [219]. In striking contrast, DNA methylation of imprinted genes in oocytes was not altered in either obese dams or their offspring. However, DNA methylation of metabolism-related genes was changed. In oocytes of obese mice, the DNA methylation level of the Leptin promoter was significantly increased and that of the PPAR-α promoter was reduced [220]. Interestingly, rosiglitazone was able to normalize oocyte developmental competence from obese mice [221], indicating that its target, PPAR-γ, may be a key regulator of metabolic mechanisms controlling oocyte quality. Failure to establish or maintain germ cell methylation patterns may result in abnormal fetal growth, placental failure, and human diseases [222].
A less studied but recently emerging concept is the integration of information about a cell’s metabolic state into the regulation of epigenetics [223]. Various enzymes responsible for adding or removing epigenetic modifications have been identified. These enzymes that play important roles in epigenetic gene regulation utilize substrates or cofactors generated by cellular metabolism, thereby providing a potential link between nutrition, metabolism, and epigenetic modifications [224]. Most cytosolic acetyl-CoA comes from citrate exported from mitochondria in mammalian cells. Tracing experiments with radiolabeled glucose revealed that about half of the acetyl groups on histone H4K16 came from glucose-derived acetyl-CoA [225] (Fig. 1). Similarly, abrogation of acetyl-CoA generating enzymes leads to global histone hypoacetylation [226]. S-Adenosyl methionine (SAM) is derived from the essential amino acid methionine in an ATP-dependent manner. Both histone and DNA methylation require SAM as the high-energy methyl donor [227]. In response to a low ATP/AMP ratio, AMPK can translocate to chromatin and phosphorylate histone H2B serine 36 [228] (Fig. 1). There is also evidence that changes in glucose and glucose metabolism can leave lasting epigenetic marks [229]. Of particular note, metabolic disorders in oocytes, such as reduced citrate levels, ATP/AMP ratio, and glucose/lipid accumulation have been discovered under nutrition-deficient conditions. The findings outlined herein provide a mechanistic framework for understanding how nutritional effects on oocyte metabolism could impact epigenetic modifications and, therefore, embryo/offspring development where epigenetic alterations play a role (Fig. 2). Future focus should be directed towards identifying how specific metabolites and metabolic co-substrates interact with chromatin to epigenetically alter gene expression.
In addition, accumulating evidence suggests that the metabolic mTOR/PTEN/PI3K signaling pathways play crucial roles during follicular growth and oocyte development in mice [230, 231]. Genetic data have shown that all primordial follicles become depleted in early adulthood of mice lacking PTEN in oocytes, causing premature ovarian failure [232]. PI3K-AKT-mTOR pathway is involved in somatic regulation of oocyte mRNA translation, affecting its developmental competence [233]. It is interesting to note that HFD-induced obesity accelerates the ovarian follicular development and the rate of follicle loss through activating mTOR signaling in rats [234]. mTOR has also recently been indicated to function in the PCOS mouse ovary [235]. However, whether these metabolic pathways in oocytes mediate the nutritional effects on pregnancy outcomes remains unknown. This topic would be a valuable research direction and deserves more attention in the future. Additionally, in response to the changes in maternal nutrition, the fluctuation of a series of extra- and intra-ovarian factors, such as epidermal growth factor (EGF), insulin-like growth factors (IGFs), and leptin, have been identified [182, 236, 237]. Whether, and if so how, they influence oocyte quality and reproductive outcomes requires further clarification and will not be discussed here as it is beyond the scope of this review.
In summary, increasing evidence supports a model where maternal nutrition imbalance disrupts energy metabolism in oocytes. This disruption further contributes to reproductive problems via the metabolic control of meiosis, mitochondria, or epigenetic modifications. As a consequence, meiotic defects result in nondisjunction inducing embryonic aneuploidy. The dysfunctional complements of mitochondria are maternally transmitted and epigenetic changes are likely to be propagated during embryogenesis, leading to the developmental abnormalities of the offspring. Further clarification is needed to exclude other possible metabolism-related pathways in oocytes that mediate the maternal nutritional effects on reproductive outcomes.
Metabolic analysis and oocyte quality
Metabolite measurement in oocytes
Metabolic analysis of mammalian oocytes started in the 1980s, which mainly used radiolabeled substrates to define the metabolites that oocytes require for their growth and maturation. A series of fundamental findings in this area have been discovered successively, such as utilization of glucose and pyruvate, uptake and metabolism of ribonucleoside and hypoxanthine, and transport of amino acids during oocyte development [40, 52, 117, 238–242]. Alternatively, we have developed a technique to measure metabolites and enzyme activities in single oocytes or preimplantation embryos, using not radioactivity but enzymatic cycling reactions that are based on the amplification of a fluorescent signal of pyridine nucleotide [243]. NADH and NADPH have identical absorption bands near UV light, which peak at 340 nm. NAD+ and NADP+ do not absorb at this wavelength. Therefore, changes in oxidation or reduction can be measured in the spectrophotometer. According to this principle, separate assays are developed for each metabolite and are designed to link reactions ending with NAD/NADH or NADP/NADPH, which then are enzymatically amplified in a cycling reaction, and a byproduct of the amplification step is measured in a fluorometric assay. The detailed assay conditions have been described in our previous papers [64, 169, 244].
Metabolomic search for markers of oocyte quality
Very recent attempts are looking beyond a single substance to the totality of the biochemical constituents in the follicular environment, which is referred to as metabolomics. Metabolomic analysis seeks to identify and quantify the entire collection of intracellular and extracellular metabolites. Conceptually, there are two basic analytical methodologies used in metabolomics: target analysis (analysis restricted to single or selectively defined metabolites) and metabolite profiling (study of selective groups of target compounds and their metabolic intermediates using a single analytical technique) [245]. The application of metabolomics has been widely pursued. Nevertheless, it is currently still an enormous challenge for the direct metabolomic analysis of the oocyte itself, due to the requirement of large amounts of material. Thus, development of a technique for metabolic profiling of nano- and femtomolar amounts of sample down to even the single-cell level [246–248] would greatly advance our understanding of oocyte metabolism and improve in vitro maturation systems.
As a key factor in the reproductive technologies, oocyte quality profoundly affects fertilization, early embryonic survival, pregnancy maintenance, and even the offspring health. Thus, one of the important puzzles for reproductive biologists today is the search for the reliable noninvasive predictors of oocyte quality for both human and animal reproduction. In general, the use of morphological characteristics as predictors of oocyte quality is still controversial because of its subjectivity and inaccuracies. The follicle provides a very important microenvironment for the development of COC. This makes factors in follicular cells and those secreted in follicular fluids to be an ideal source of non-invasive and operational predictors of oocyte quality. Global assessment strategies involving genomic, transcriptomic, proteomic, or metabolomic profiling of follicular fluid, cumulus cells, or culture media have been applied to assisted reproduction [249]. Metabolomics allows the understanding of biological processes to be completed, as it is capable of providing biological end-point markers of the cellular processes that occur as a result of the disease, drug exposure, or nutritional alteration [250].
Currently, assessment of the metabolite profile is based on spectroscopic/spectrometric and chromatographic techniques such as nuclear magnetic resonance (NMR) spectroscopy, mass spectrometry (MS; often combined with chromatography), and optical spectroscopy. Both NMR and MS can generate information on multiple compounds with a sample and have applied to analysis of culture media and follicular fluid [251–255]. For the first time, NMR was used to document a change in lipoprotein concentration in follicular fluid during follicular development [256]. NMR analysis of granulosa cells from rats has shown that follicle-stimulating hormone (FSH) increases the activity of the pentose phosphate pathway [257]. Using gas chromatography mass spectrometry (GC/MS)-based metabolomics, Bender et al. [258] found higher concentrations of saturated fatty acids in follicular fluid from cows and higher docosahexaenoic acid levels in follicular fluid from heifers. The elevated content of detrimental saturated fatty acids in cows will have a negative impact on oocyte maturation and early embryo development, which may explain the differences in fertility between heifers and lactating cows.
Liquid chromatography-tandem mass spectrometric (LC-MS/MS) analysis of follicular fluid revealed an increased aromatic amino acid content in PCOS patients independent of obesity, whereas the levels of branched-chain amino acid, glutamic acid, phenylalanine, alanine, and arginine are elevated with body mass index [259]. The disruptions in specific amino acids in the follicular fluid might impair oocyte quality, accounting for the inferior pregnancy outcome in obese and PCOS women. Analysis with high-performance LC also indicates that non-invasive amino acid profiling can be used to measure bovine oocyte developmental competence [260]. The most popular types of optical spectroscopy used in metabolic analysis are near infrared (NIR) and Raman spectroscopy. Although metabolite analysis obtained by these two approaches was successfully correlated with the developmental potential of embryos [254, 261, 262], opposing results have been also reported [263, 264]. Overall, there is a lack of the direct metabolomic analysis of oocytes and the surrounding cumulus cells. Data from this field will undoubtedly provide an essential network for metabolic regulation of oocyte development.
Concluding remarks
In conclusion, this review summarizes the metabolic pathways essential for oocyte development. The focus of this field has been overwhelmingly on extrinsic nutrients, and hence the intrinsic regulation of oocyte growth and development by transport systems and metabolic enzymes has been relatively neglected. Metabolic disorders, such as obesity and diabetes, have major adverse effects on fertility, pregnancy, and the health of offspring. In the present review, we highlight the importance of endogenous oocyte metabolism linking maternal nutrition and reproductive outcomes, with emphasis on the metabolic control of meiosis, mitochondria, and epigenetic modifications.
Clearly, further research is needed to uncover mechanistic details for understanding how nutritional effects on oocyte metabolism are propagated during embryogenesis and offspring development. Global assessment strategy via metabolomic profiling of follicular fluid, cumulus cells, and culture media has been applied to assisted reproduction. Data from metabolomics may serve as a proxy to predict oocyte quality and thereby prevent reproductive defects, but its application is still in the infantile stages.
Acknowledgments
This work was funded by National Key Scientific Research Projects (2014CB943200), Natural Science Foundation of the Jiangsu Higher Education Institutions (13KJA310001), Fundamental Research Funds for the Central Universities (0606J0451) of China, the American Diabetes Association (KHM), and the NIH-NICHD (R01).
Conflict of interest
The authors declare that they have no conflict of interest.
Contributor Information
Ling Gu, Email: lgu@njau.edu.cn.
Qiang Wang, Phone: +86-25-86862151, Email: qwang2012@njmu.edu.cn.
References
- 1.Krisher RL. Oocyte physiology and development in domestic animals. 1. New York: Wiley-Blackwell; 2013. [Google Scholar]
- 2.Jagarlamudi K, Rajkovic A. Oogenesis: transcriptional regulators and mouse models. Mol Cell Endocrinol. 2012;356(1–2):31–39. doi: 10.1016/j.mce.2011.07.049. [DOI] [PubMed] [Google Scholar]
- 3.Sanchez F, Smitz J. Molecular control of oogenesis. Biochim Biophys Acta. 2012;1822(12):1896–1912. doi: 10.1016/j.bbadis.2012.05.013. [DOI] [PubMed] [Google Scholar]
- 4.Polanski Z. Spindle assembly checkpoint regulation of chromosome segregation in mammalian oocytes. Reprod Fertil Dev. 2013;25(3):472–483. doi: 10.1071/RD12145. [DOI] [PubMed] [Google Scholar]
- 5.Dumont J, Desai A. Acentrosomal spindle assembly and chromosome segregation during oocyte meiosis. Trends Cell Biol. 2012;22(5):241–249. doi: 10.1016/j.tcb.2012.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hunt PA, Hassold TJ. Human female meiosis: what makes a good egg go bad? Trends in genetics: TIG. 2008;24(2):86–93. doi: 10.1016/j.tig.2007.11.010. [DOI] [PubMed] [Google Scholar]
- 7.Eppig JJ. Coordination of nuclear and cytoplasmic oocyte maturation in eutherian mammals. Reprod Fertil Dev. 1996;8(4):485–489. doi: 10.1071/rd9960485. [DOI] [PubMed] [Google Scholar]
- 8.Krisher RL. The effect of oocyte quality on development. J Anim Sci. 2004;82:E14–E23. doi: 10.2527/2004.8213_supplE14x. [DOI] [PubMed] [Google Scholar]
- 9.Peters H, McNatty KP. The ovary. New York: Granada; 1980. [Google Scholar]
- 10.Van Blerkom J. Mitochondrial function in the human oocyte and embryo and their role in developmental competence. Mitochondrion. 2011;11(5):797–813. doi: 10.1016/j.mito.2010.09.012. [DOI] [PubMed] [Google Scholar]
- 11.Tatone C, Heizenrieder T, Di Emidio G, Treffon P, Amicarelli F, Seidel T, Eichenlaub-Ritter U. Evidence that carbonyl stress by methylglyoxal exposure induces DNA damage and spindle aberrations, affects mitochondrial integrity in mammalian oocytes and contributes to oocyte ageing. Hum Reprod. 2011;26(7):1843–1859. doi: 10.1093/humrep/der140. [DOI] [PubMed] [Google Scholar]
- 12.Johnson MT, Freeman EA, Gardner DK, Hunt PA. Oxidative metabolism of pyruvate is required for meiotic maturation of murine oocytes in vivo. Biol Reprod. 2007;77(1):2–8. doi: 10.1095/biolreprod.106.059899. [DOI] [PubMed] [Google Scholar]
- 13.Zuelke KA, Jones DP, Perreault SD. Glutathione oxidation is associated with altered microtubule function and disrupted fertilization in mature hamster oocytes. Biol Reprod. 1997;57(6):1413–1419. doi: 10.1095/biolreprod57.6.1413. [DOI] [PubMed] [Google Scholar]
- 14.Eichenlaub-Ritter U, Wieczorek M, Luke S, Seidel T. Age related changes in mitochondrial function and new approaches to study redox regulation in mammalian oocytes in response to age or maturation conditions. Mitochondrion. 2011;11(5):783–796. doi: 10.1016/j.mito.2010.08.011. [DOI] [PubMed] [Google Scholar]
- 15.Buccione R, Schroeder AC, Eppig JJ. Interactions between somatic cells and germ cells throughout mammalian oogenesis. Biol Reprod. 1990;43(4):543–547. doi: 10.1095/biolreprod43.4.543. [DOI] [PubMed] [Google Scholar]
- 16.Sugiura K, Pendola FL, Eppig JJ. Oocyte control of metabolic cooperativity between oocytes and companion granulosa cells: energy metabolism. Dev Biol. 2005;279(1):20–30. doi: 10.1016/j.ydbio.2004.11.027. [DOI] [PubMed] [Google Scholar]
- 17.Li R, Albertini DF. The road to maturation: somatic cell interaction and self-organization of the mammalian oocyte. Nat Rev Mol Cell Biol. 2013;14(3):141–152. doi: 10.1038/nrm3531. [DOI] [PubMed] [Google Scholar]
- 18.Leese HJ, Barton AM. Production of pyruvate by isolated mouse cumulus cells. J Exp Zool. 1985;234(2):231–236. doi: 10.1002/jez.1402340208. [DOI] [PubMed] [Google Scholar]
- 19.Sugiura K, Eppig JJ, Society for Reproductive Biology Founders’ Lecture 2005 Control of metabolic cooperativity between oocytes and their companion granulosa cells by mouse oocytes. Reprod Fertil Dev. 2005;17(7):667–674. doi: 10.1071/rd05071. [DOI] [PubMed] [Google Scholar]
- 20.Su YQ, Sugiura K, Eppig JJ. Mouse oocyte control of granulosa cell development and function: paracrine regulation of cumulus cell metabolism. Semin Reprod Med. 2009;27(1):32–42. doi: 10.1055/s-0028-1108008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Grindler NM, Moley KH. Maternal obesity, infertility and mitochondrial dysfunction: potential mechanisms emerging from mouse model systems. Mol Hum Reprod. 2013;19(8):486–494. doi: 10.1093/molehr/gat026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Collado-Fernandez E, Picton HM, Dumollard R. Metabolism throughout follicle and oocyte development in mammals. Int J Dev Biol. 2012;56(10–12):799–808. doi: 10.1387/ijdb.120140ec. [DOI] [PubMed] [Google Scholar]
- 23.Hernandez-Medrano JH, Campbell BK, Webb R. Nutritional influences on folliculogenesis. Reprod Domest Anim. 2012;47(Suppl 4):274–282. doi: 10.1111/j.1439-0531.2012.02086.x. [DOI] [PubMed] [Google Scholar]
- 24.Gosden R, Lee B. Portrait of an oocyte: our obscure origin. J Clin Investig. 2010;120(4):973–983. doi: 10.1172/JCI41294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Matzuk MM, Burns KH, Viveiros MM, Eppig JJ. Intercellular communication in the mammalian ovary: oocytes carry the conversation. Science. 2002;296(5576):2178–2180. doi: 10.1126/science.1071965. [DOI] [PubMed] [Google Scholar]
- 26.Gilchrist RB, Lane M, Thompson JG. Oocyte-secreted factors: regulators of cumulus cell function and oocyte quality. Hum Reprod Update. 2008;14(2):159–177. doi: 10.1093/humupd/dmm040. [DOI] [PubMed] [Google Scholar]
- 27.Leese HJ. Metabolic control during preimplantation mammalian development. Hum Reprod Update. 1995;1(1):63–72. doi: 10.1093/humupd/1.1.63. [DOI] [PubMed] [Google Scholar]
- 28.Kurus M, Karakaya C, Karalok MH, To G, Johnson J. The control of oocyte survival by intrinsic and extrinsic factors. Adv Exp Med Biol. 2013;761:7–18. doi: 10.1007/978-1-4614-8214-7_2. [DOI] [PubMed] [Google Scholar]
- 29.Purcell SH, Moley KH. Glucose transporters in gametes and preimplantation embryos. Trends Endocrinol Metab. 2009;20(10):483–489. doi: 10.1016/j.tem.2009.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Baltz JM, Zhou C. Cell volume regulation in mammalian oocytes and preimplantation embryos. Mol Reprod Dev. 2012;79(12):821–831. doi: 10.1002/mrd.22117. [DOI] [PubMed] [Google Scholar]
- 31.Fragouli E, Lalioti MD, Wells D. The transcriptome of follicular cells: biological insights and clinical implications for the treatment of infertility. Hum Reprod Update. 2014;20(1):1–11. doi: 10.1093/humupd/dmt044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Krisher RL, Brad AM, Herrick JR, Sparman ML, Swain JE. A comparative analysis of metabolism and viability in porcine oocytes during in vitro maturation. Anim Reprod Sci. 2007;98(1–2):72–96. doi: 10.1016/j.anireprosci.2006.10.006. [DOI] [PubMed] [Google Scholar]
- 33.Anckaert E, De Rycke M, Smitz J. Culture of oocytes and risk of imprinting defects. Hum Reprod Update. 2013;19(1):52–66. doi: 10.1093/humupd/dms042. [DOI] [PubMed] [Google Scholar]
- 34.Van Hoeck V, Bols PE, Binelli M, Leroy JL. Reduced oocyte and embryo quality in response to elevated non-esterified fatty acid concentrations: a possible pathway to subfertility? Anim Reprod Sci. 2014 doi: 10.1016/j.anireprosci.2014.07.015. [DOI] [PubMed] [Google Scholar]
- 35.Thompson JG. The impact of nutrition of the cumulus oocyte complex and embryo on subsequent development in ruminants. J Reprod Dev. 2006;52(1):169–175. doi: 10.1262/jrd.17089. [DOI] [PubMed] [Google Scholar]
- 36.Sutton-McDowall ML, Gilchrist RB, Thompson JG. The pivotal role of glucose metabolism in determining oocyte developmental competence. Reproduction. 2010;139(4):685–695. doi: 10.1530/REP-09-0345. [DOI] [PubMed] [Google Scholar]
- 37.Brinster RL, Harstad H. Energy metabolism in primordial germ cells of the mouse. Exp Cell Res. 1977;109(1):111–117. doi: 10.1016/0014-4827(77)90050-7. [DOI] [PubMed] [Google Scholar]
- 38.Jansen RP, de Boer K. The bottleneck: mitochondrial imperatives in oogenesis and ovarian follicular fate. Mol Cell Endocrinol. 1998;145(1–2):81–88. doi: 10.1016/s0303-7207(98)00173-7. [DOI] [PubMed] [Google Scholar]
- 39.Harris SE, Leese HJ, Gosden RG, Picton HM. Pyruvate and oxygen consumption throughout the growth and development of murine oocytes. Mol Reprod Dev. 2009;76(3):231–238. doi: 10.1002/mrd.20945. [DOI] [PubMed] [Google Scholar]
- 40.Biggers JD, Whittingham DG, Donahue RP. The pattern of energy metabolism in the mouse oocyte and zygote. Proc Natl Acad Sci USA. 1967;58(2):560–567. doi: 10.1073/pnas.58.2.560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Rieger D, Loskutoff NM. Changes in the metabolism of glucose, pyruvate, glutamine and glycine during maturation of cattle oocytes in vitro. J Reprod Fertil. 1994;100(1):257–262. doi: 10.1530/jrf.0.1000257. [DOI] [PubMed] [Google Scholar]
- 42.Saito T, Hiroi M, Kato T. Development of glucose utilization studied in single oocytes and preimplantation embryos from mice. Biol Reprod. 1994;50(2):266–270. doi: 10.1095/biolreprod50.2.266. [DOI] [PubMed] [Google Scholar]
- 43.Zuelke KA, Brackett BG. Effects of luteinizing hormone on glucose metabolism in cumulus-enclosed bovine oocytes matured in vitro. Endocrinology. 1992;131(6):2690–2696. doi: 10.1210/endo.131.6.1446610. [DOI] [PubMed] [Google Scholar]
- 44.Downs SM, Mastropolo AM. The participation of energy substrates in the control of meiotic maturation in murine oocytes. Dev Biol. 1994;162(1):154–168. doi: 10.1006/dbio.1994.1075. [DOI] [PubMed] [Google Scholar]
- 45.Eppig JJ. Analysis of mouse oogenesis in vitro. Oocyte isolation and the utilization of exogenous energy sources by growing oocytes. The Journal of experimental zoology. 1976;198(3):375–382. doi: 10.1002/jez.1401980311. [DOI] [PubMed] [Google Scholar]
- 46.Fagbohun CF, Downs SM. Requirement for glucose in ligand-stimulated meiotic maturation of cumulus cell-enclosed mouse oocytes. J Reprod Fertil. 1992;96(2):681–697. doi: 10.1530/jrf.0.0960681. [DOI] [PubMed] [Google Scholar]
- 47.Halestrap AP, Price NT. The proton-linked monocarboxylate transporter (MCT) family: structure, function and regulation. Biochem J. 1999;343(Pt 2):281–299. [PMC free article] [PubMed] [Google Scholar]
- 48.Herubel F, El Mouatassim S, Guerin P, Frydman R, Menezo Y. Genetic expression of monocarboxylate transporters during human and murine oocyte maturation and early embryonic development. Zygote. 2002;10(2):175–181. doi: 10.1017/s096719940200223x. [DOI] [PubMed] [Google Scholar]
- 49.Songsasen N, Wesselowski S, Carpenter JW, Wildt DE. The ability to achieve meiotic maturation in the dog oocyte is linked to glycolysis and glutamine oxidation. Mol Reprod Dev. 2012;79(3):186–196. doi: 10.1002/mrd.22011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Brinster RL. Hexokinase activity in the preimplantation mouse embryo. Enzymologia. 1968;34(5):304–308. [PubMed] [Google Scholar]
- 51.Brinster RL. Oxidation of pyruvate and glucose by oocytes of the mouse and rhesus monkey. J Reprod Fertil. 1971;24(2):187–191. doi: 10.1530/jrf.0.0240187. [DOI] [PubMed] [Google Scholar]
- 52.Downs SM, Utecht AM. Metabolism of radiolabeled glucose by mouse oocytes and oocyte-cumulus cell complexes. Biol Reprod. 1999;60(6):1446–1452. doi: 10.1095/biolreprod60.6.1446. [DOI] [PubMed] [Google Scholar]
- 53.Rushmer RA, Brinster RL. Carbon dioxide production from pyruvate and glucose by bovine oocytes. Exp Cell Res. 1973;82(2):252–254. doi: 10.1016/0014-4827(73)90338-8. [DOI] [PubMed] [Google Scholar]
- 54.Urner F, Sakkas D. Characterization of glycolysis and pentose phosphate pathway activity during sperm entry into the mouse oocyte. Biol Reprod. 1999;60(4):973–978. doi: 10.1095/biolreprod60.4.973. [DOI] [PubMed] [Google Scholar]
- 55.Joost HG, Bell GI, Best JD, Birnbaum MJ, Charron MJ, Chen YT, Doege H, James DE, Lodish HF, Moley KH, Moley JF, Mueckler M, Rogers S, Schurmann A, Seino S, Thorens B. Nomenclature of the GLUT/SLC2A family of sugar/polyol transport facilitators. Am J Physiol Endocrinol Metab. 2002;282(4):E974–E976. doi: 10.1152/ajpendo.00407.2001. [DOI] [PubMed] [Google Scholar]
- 56.Aghayan M, Rao LV, Smith RM, Jarett L, Charron MJ, Thorens B, Heyner S. Developmental expression and cellular localization of glucose transporter molecules during mouse preimplantation development. Development. 1992;115(1):305–312. doi: 10.1242/dev.115.1.305. [DOI] [PubMed] [Google Scholar]
- 57.Morita Y, Tsutsumi O, Hosoya I, Taketani Y, Oka Y, Kato T. Expression and possible function of glucose transporter protein GLUT1 during preimplantation mouse development from oocytes to blastocysts. Biochem Biophys Res Commun. 1992;188(1):8–15. doi: 10.1016/0006-291x(92)92342-u. [DOI] [PubMed] [Google Scholar]
- 58.Augustin R, Pocar P, Navarrete-Santos A, Wrenzycki C, Gandolfi F, Niemann H, Fischer B. Glucose transporter expression is developmentally regulated in in vitro derived bovine preimplantation embryos. Mol Reprod Dev. 2001;60(3):370–376. doi: 10.1002/mrd.1099. [DOI] [PubMed] [Google Scholar]
- 59.Pisani LF, Antonini S, Pocar P, Ferrari S, Brevini TA, Rhind SM, Gandolfi F. Effects of pre-mating nutrition on mRNA levels of developmentally relevant genes in sheep oocytes and granulosa cells. Reproduction. 2008;136(3):303–312. doi: 10.1530/REP-07-0394. [DOI] [PubMed] [Google Scholar]
- 60.Dan-Goor M, Sasson S, Davarashvili A, Almagor M. Expression of glucose transporter and glucose uptake in human oocytes and preimplantation embryos. Hum Reprod. 1997;12(11):2508–2510. doi: 10.1093/humrep/12.11.2508. [DOI] [PubMed] [Google Scholar]
- 61.Zheng P, Vassena R, Latham KE. Effects of in vitro oocyte maturation and embryo culture on the expression of glucose transporters, glucose metabolism and insulin signaling genes in rhesus monkey oocytes and preimplantation embryos. Mol Hum Reprod. 2007;13(6):361–371. doi: 10.1093/molehr/gam014. [DOI] [PubMed] [Google Scholar]
- 62.Purcell SH, Moley KH. The impact of obesity on egg quality. J Assist Reprod Genet. 2011;28(6):517–524. doi: 10.1007/s10815-011-9592-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Wang Q, Chi MM, Schedl T, Moley KH. An intercellular pathway for glucose transport into mouse oocytes. Am J Phys Endocrinol Metab. 2012;302(12):E1511–E1518. doi: 10.1152/ajpendo.00016.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Chi MM, Manchester JK, Yang VC, Curato AD, Strickler RC, Lowry OH. Contrast in levels of metabolic enzymes in human and mouse ova. Biol Reprod. 1988;39(2):295–307. doi: 10.1095/biolreprod39.2.295. [DOI] [PubMed] [Google Scholar]
- 65.De Schepper GG, Van Noorden CJ, Koperdraad F. A cytochemical method for measuring enzyme activity in individual preovulatory mouse oocytes. J Reprod Fertil. 1985;74(2):709–716. doi: 10.1530/jrf.0.0740709. [DOI] [PubMed] [Google Scholar]
- 66.Tsutsumi O, Satoh K, Taketani Y, Kato T. Determination of enzyme activities of energy metabolism in the maturing rat oocyte. Mol Reprod Dev. 1992;33(3):333–337. doi: 10.1002/mrd.1080330315. [DOI] [PubMed] [Google Scholar]
- 67.Ferrandi B, Cremonesti F, Geiger R, Consiglio AL, Carnevali A, Porcelli F. Quantitative cytochemical study of some enzymatic activities in preovulatory bovine oocytes after in vitro maturation. Acta Histochem. 1993;95(1):89–96. doi: 10.1016/S0065-1281(11)80394-6. [DOI] [PubMed] [Google Scholar]
- 68.Kelly A, West JD. Survival and normal function of glycolysis-deficient mouse oocytes. Reproduction. 2002;124(4):469–473. doi: 10.1530/rep.0.1240469. [DOI] [PubMed] [Google Scholar]
- 69.Pujol M, Lopez-Bejar M, Paramio MT. Developmental competence of heifer oocytes selected using the brilliant cresyl blue (BCB) test. Theriogenology. 2004;61(4):735–744. doi: 10.1016/s0093-691x(03)00250-4. [DOI] [PubMed] [Google Scholar]
- 70.Alm H, Torner H, Lohrke B, Viergutz T, Ghoneim IM, Kanitz W. Bovine blastocyst development rate in vitro is influenced by selection of oocytes by brillant cresyl blue staining before IVM as indicator for glucose-6-phosphate dehydrogenase activity. Theriogenology. 2005;63(8):2194–2205. doi: 10.1016/j.theriogenology.2004.09.050. [DOI] [PubMed] [Google Scholar]
- 71.El Shourbagy SH, Spikings EC, Freitas M, St John JC. Mitochondria directly influence fertilisation outcome in the pig. Reproduction. 2006;131(2):233–245. doi: 10.1530/rep.1.00551. [DOI] [PubMed] [Google Scholar]
- 72.Pawlak P, Warzych E, Chabowska A, Lechniak D. Differences in cytoplasmic maturation between the BCB + and control porcine oocytes do not justify application of the BCB test for a standard IVM protocol. J Reprod Dev. 2014;60(1):28–36. doi: 10.1262/jrd.2013-092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Santos EC, Sato D, Lucia T, Iwata H (2013) Brilliant cresyl blue staining negatively affects mitochondrial functions in porcine oocytes. Zygote:1–8. doi:10.1017/S0967199413000610 [DOI] [PubMed]
- 74.Loewenstein JE, Cohen AI. Dry mass, lipid content and protein content of the intact and zona-free mouse ovum. J Embryol Exp Morphol. 1964;12:113–121. [PubMed] [Google Scholar]
- 75.McEvoy TG, Coull GD, Broadbent PJ, Hutchinson JS, Speake BK. Fatty acid composition of lipids in immature cattle, pig and sheep oocytes with intact zona pellucida. J Reprod Fertil. 2000;118(1):163–170. [PubMed] [Google Scholar]
- 76.Sturmey RG, Leese HJ. Energy metabolism in pig oocytes and early embryos. Reproduction. 2003;126(2):197–204. doi: 10.1530/rep.0.1260197. [DOI] [PubMed] [Google Scholar]
- 77.Ferguson EM, Leese HJ. Triglyceride content of bovine oocytes and early embryos. J Reprod Fertil. 1999;116(2):373–378. doi: 10.1530/jrf.0.1160373. [DOI] [PubMed] [Google Scholar]
- 78.Kim JY, Kinoshita M, Ohnishi M, Fukui Y. Lipid and fatty acid analysis of fresh and frozen-thawed immature and in vitro matured bovine oocytes. Reproduction. 2001;122(1):131–138. [PubMed] [Google Scholar]
- 79.Homa ST, Racowsky C, McGaughey RW. Lipid analysis of immature pig oocytes. J Reprod Fertil. 1986;77(2):425–434. doi: 10.1530/jrf.0.0770425. [DOI] [PubMed] [Google Scholar]
- 80.Yang X, Dunning KR, Wu LL, Hickey TE, Norman RJ, Russell DL, Liang X, Robker RL. Identification of perilipin-2 as a lipid droplet protein regulated in oocytes during maturation. Reprod Fertil Dev. 2010;22(8):1262–1271. doi: 10.1071/RD10091. [DOI] [PubMed] [Google Scholar]
- 81.Guraya SS. Histochemical study of lipids in the developing ovarian oocyte of the golden hamster (Mesocricetus auratus) Acta Anat. 1969;74(1):65–75. doi: 10.1159/000143363. [DOI] [PubMed] [Google Scholar]
- 82.Guraya SS. Histochemical observations on the juxtanuclear complex of organelles in the hamster oocyte. Acta Anat. 1975;93(3):335–343. doi: 10.1159/000144509. [DOI] [PubMed] [Google Scholar]
- 83.Sturmey RG, O’Toole PJ, Leese HJ. Fluorescence resonance energy transfer analysis of mitochondrial:lipid association in the porcine oocyte. Reproduction. 2006;132(6):829–837. doi: 10.1530/REP-06-0073. [DOI] [PubMed] [Google Scholar]
- 84.Aardema H, Vos PL, Lolicato F, Roelen BA, Knijn HM, Vaandrager AB, Helms JB, Gadella BM. Oleic acid prevents detrimental effects of saturated fatty acids on bovine oocyte developmental competence. Biol Reprod. 2011;85(1):62–69. doi: 10.1095/biolreprod.110.088815. [DOI] [PubMed] [Google Scholar]
- 85.Brasaemle DL. Thematic review series: adipocyte biology. The perilipin family of structural lipid droplet proteins: stabilization of lipid droplets and control of lipolysis. J Lipid Res. 2007;48(12):2547–2559. doi: 10.1194/jlr.R700014-JLR200. [DOI] [PubMed] [Google Scholar]
- 86.Fleming WN, Saacke RG. Fine structure of the bovine oocyte from the mature graafian follicle. J Reprod Fertil. 1972;29(2):203–213. doi: 10.1530/jrf.0.0290203. [DOI] [PubMed] [Google Scholar]
- 87.Van Praag D, Farber SJ. Biosynthesis of cyclooxygenase and lipoxygenase metabolites of arachidonic acid by rabbit renal microsomes. Prostaglandins Leukot Med. 1983;12(1):29–47. doi: 10.1016/0262-1746(83)90065-3. [DOI] [PubMed] [Google Scholar]
- 88.Lopaschuk GD, Ussher JR, Folmes CD, Jaswal JS, Stanley WC. Myocardial fatty acid metabolism in health and disease. Physiol Rev. 2010;90(1):207–258. doi: 10.1152/physrev.00015.2009. [DOI] [PubMed] [Google Scholar]
- 89.Dunning KR, Russell DL, Robker RL. Lipids and oocyte developmental competence: the role of fatty acids and beta-oxidation. Reproduction. 2014;148(1):R15–R27. doi: 10.1530/REP-13-0251. [DOI] [PubMed] [Google Scholar]
- 90.McKeegan PJ, Sturmey RG. The role of fatty acids in oocyte and early embryo development. Reprod Fertil Dev. 2011;24(1):59–67. doi: 10.1071/RD11907. [DOI] [PubMed] [Google Scholar]
- 91.Ferguson EM, Leese HJ. A potential role for triglyceride as an energy source during bovine oocyte maturation and early embryo development. Mol Reprod Dev. 2006;73(9):1195–1201. doi: 10.1002/mrd.20494. [DOI] [PubMed] [Google Scholar]
- 92.Downs SM, Mosey JL, Klinger J. Fatty acid oxidation and meiotic resumption in mouse oocytes. Mol Reprod Dev. 2009;76(9):844–853. doi: 10.1002/mrd.21047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Dunning KR, Cashman K, Russell DL, Thompson JG, Norman RJ, Robker RL. Beta-oxidation is essential for mouse oocyte developmental competence and early embryo development. Biol Reprod. 2010;83(6):909–918. doi: 10.1095/biolreprod.110.084145. [DOI] [PubMed] [Google Scholar]
- 94.Cetica P, Pintos L, Dalvit G, Beconi M. Activity of key enzymes involved in glucose and triglyceride catabolism during bovine oocyte maturation in vitro. Reproduction. 2002;124(5):675–681. [PubMed] [Google Scholar]
- 95.Dunning KR, Akison LK, Russell DL, Norman RJ, Robker RL. Increased beta-oxidation and improved oocyte developmental competence in response to l-carnitine during ovarian in vitro follicle development in mice. Biol Reprod. 2011;85(3):548–555. doi: 10.1095/biolreprod.110.090415. [DOI] [PubMed] [Google Scholar]
- 96.Hashimoto S. Application of in vitro maturation to assisted reproductive technology. J Reprod Dev. 2009;55(1):1–10. doi: 10.1262/jrd.20127. [DOI] [PubMed] [Google Scholar]
- 97.Somfai T, Kaneda M, Akagi S, Watanabe S, Haraguchi S, Mizutani E, Dang-Nguyen TQ, Geshi M, Kikuchi K, Nagai T. Enhancement of lipid metabolism with L-carnitine during in vitro maturation improves nuclear maturation and cleavage ability of follicular porcine oocytes. Reprod Fertil Dev. 2011;23(7):912–920. doi: 10.1071/RD10339. [DOI] [PubMed] [Google Scholar]
- 98.Wu GQ, Jia BY, Li JJ, Fu XW, Zhou GB, Hou YP, Zhu SE. L-carnitine enhances oocyte maturation and development of parthenogenetic embryos in pigs. Theriogenology. 2011;76(5):785–793. doi: 10.1016/j.theriogenology.2011.04.011. [DOI] [PubMed] [Google Scholar]
- 99.Paczkowski M, Silva E, Schoolcraft WB, Krisher RL. Comparative importance of fatty acid beta-oxidation to nuclear maturation, gene expression, and glucose metabolism in mouse, bovine, and porcine cumulus oocyte complexes. Biol Reprod. 2013;88(5):111. doi: 10.1095/biolreprod.113.108548. [DOI] [PubMed] [Google Scholar]
- 100.Huang YH, Sauer K. Lipid signaling in T-cell development and function. Cold Spring Harb Perspect Biol. 2010;2(11):a002428. doi: 10.1101/cshperspect.a002428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Griner EM, Kazanietz MG. Protein kinase C and other diacylglycerol effectors in cancer. Nat Rev Cancer. 2007;7(4):281–294. doi: 10.1038/nrc2110. [DOI] [PubMed] [Google Scholar]
- 102.Bell RM, Coleman RA. Enzymes of glycerolipid synthesis in eukaryotes. Annu Rev Biochem. 1980;49:459–487. doi: 10.1146/annurev.bi.49.070180.002331. [DOI] [PubMed] [Google Scholar]
- 103.Bell RM. Protein kinase C activation by diacylglycerol second messengers. Cell. 1986;45(5):631–632. doi: 10.1016/0092-8674(86)90774-9. [DOI] [PubMed] [Google Scholar]
- 104.Nishizuka Y. The role of protein kinase C in cell surface signal transduction and tumour promotion. Nature. 1984;308(5961):693–698. doi: 10.1038/308693a0. [DOI] [PubMed] [Google Scholar]
- 105.Wang QJ. PKD at the crossroads of DAG and PKC signaling. Trends Pharmacol Sci. 2006;27(6):317–323. doi: 10.1016/j.tips.2006.04.003. [DOI] [PubMed] [Google Scholar]
- 106.Kalive M, Faust JJ, Koeneman BA, Capco DG. Involvement of the PKC family in regulation of early development. Mol Reprod Dev. 2010;77(2):95–104. doi: 10.1002/mrd.21112. [DOI] [PubMed] [Google Scholar]
- 107.Yeboah D, Kalabis GM, Sun M, Ou RC, Matthews SG, Gibb W. Expression and localisation of breast cancer resistance protein (BCRP) in human fetal membranes and decidua and the influence of labour at term. Reprod Fertil Dev. 2008;20(2):328–334. doi: 10.1071/rd07133. [DOI] [PubMed] [Google Scholar]
- 108.Sun QY, Miao YL, Schatten H. Towards a new understanding on the regulation of mammalian oocyte meiosis resumption. Cell Cycle. 2009;8(17):2741–2747. doi: 10.4161/cc.8.17.9471. [DOI] [PubMed] [Google Scholar]
- 109.Ghosh S, Strum JC, Bell RM. Lipid biochemistry: functions of glycerolipids and sphingolipids in cellular signaling. FASEB J. 1997;11(1):45–50. doi: 10.1096/fasebj.11.1.9034165. [DOI] [PubMed] [Google Scholar]
- 110.Strum JC, Swenson KI, Turner JE, Bell RM. Ceramide triggers meiotic cell cycle progression in Xenopus oocytes. A potential mediator of progesterone-induced maturation. J Biol Chem. 1995;270(22):13541–13547. doi: 10.1074/jbc.270.22.13541. [DOI] [PubMed] [Google Scholar]
- 111.Sampath H, Ntambi JM. Polyunsaturated fatty acid regulation of genes of lipid metabolism. Annu Rev Nutr. 2005;25:317–340. doi: 10.1146/annurev.nutr.25.051804.101917. [DOI] [PubMed] [Google Scholar]
- 112.Cui Y, Miyoshi K, Claudio E, Siebenlist UK, Gonzalez FJ, Flaws J, Wagner KU, Hennighausen L. Loss of the peroxisome proliferation-activated receptor gamma (PPARgamma) does not affect mammary development and propensity for tumor formation but leads to reduced fertility. J Biol Chem. 2002;277(20):17830–17835. doi: 10.1074/jbc.M200186200. [DOI] [PubMed] [Google Scholar]
- 113.Al Darwich A, Perreau C, Petit MH, Papillier P, Dupont J, Guillaume D, Mermillod P, Guignot F. Effect of PUFA on embryo cryoresistance, gene expression and AMPKalpha phosphorylation in IVF-derived bovine embryos. Prostaglandins Other Lipid Mediat. 2010;93(1–2):30–36. doi: 10.1016/j.prostaglandins.2010.06.002. [DOI] [PubMed] [Google Scholar]
- 114.Sutton ML, Gilchrist RB, Thompson JG. Effects of in vivo and in vitro environments on the metabolism of the cumulus-oocyte complex and its influence on oocyte developmental capacity. Hum Reprod Update. 2003;9(1):35–48. doi: 10.1093/humupd/dmg009. [DOI] [PubMed] [Google Scholar]
- 115.Bae IH, Foote RH. Utilization of glutamine for energy and protein synthesis by cultured rabbit follicular oocytes. Exp Cell Res. 1975;90(2):432–436. doi: 10.1016/0014-4827(75)90333-x. [DOI] [PubMed] [Google Scholar]
- 116.Van Winkle LJ. Amino acid transport regulation and early embryo development. Biol Reprod. 2001;64(1):1–12. doi: 10.1095/biolreprod64.1.1. [DOI] [PubMed] [Google Scholar]
- 117.Pelland AM, Corbett HE, Baltz JM. Amino Acid transport mechanisms in mouse oocytes during growth and meiotic maturation. Biol Reprod. 2009;81(6):1041–1054. doi: 10.1095/biolreprod.109.079046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Christensen HN. Role of amino acid transport and countertransport in nutrition and metabolism. Physiol Rev. 1990;70(1):43–77. doi: 10.1152/physrev.1990.70.1.43. [DOI] [PubMed] [Google Scholar]
- 119.Colonna R, Cecconi S, Buccione R, Mangia F. Amino acid transport systems in growing mouse oocytes. Cell Biol Int Rep. 1983;7(12):1007–1015. doi: 10.1016/0309-1651(83)90005-x. [DOI] [PubMed] [Google Scholar]
- 120.Haghighat N, Van Winkle LJ. Developmental change in follicular cell-enhanced amino acid uptake into mouse oocytes that depends on intact gap junctions and transport system Gly. J Exp Zool. 1990;253(1):71–82. doi: 10.1002/jez.1402530110. [DOI] [PubMed] [Google Scholar]
- 121.Van Winkle LJ, Mann DF, Wasserlauf HG, Patel M. Mediated Na(+)-independent transport of l-glutamate and l-cystine in 1- and 2-cell mouse conceptuses. Biochim Biophys Acta. 1992;1107(2):299–304. doi: 10.1016/0005-2736(92)90416-j. [DOI] [PubMed] [Google Scholar]
- 122.Van Winkle LJ, Patel M, Wasserlauf HG, Dickinson HR, Campione AL. Osmotic regulation of taurine transport via system beta and novel processes in mouse preimplantation conceptuses. Biochim Biophys Acta. 1994;1191(2):244–255. doi: 10.1016/0005-2736(94)90175-9. [DOI] [PubMed] [Google Scholar]
- 123.Van Winkle LJ, Haghighat N, Campione AL, Gorman JM. Glycine transport in mouse eggs and preimplantation conceptuses. Biochim Biophys Acta. 1988;941(2):241–256. doi: 10.1016/0005-2736(88)90185-x. [DOI] [PubMed] [Google Scholar]
- 124.Van Winkle LJ, Campione AL, Gorman JM, Weimer BD. Changes in the activities of amino acid transport systems b0, + and L during development of preimplantation mouse conceptuses. Biochim Biophys Acta. 1990;1021(1):77–84. doi: 10.1016/0005-2736(90)90387-4. [DOI] [PubMed] [Google Scholar]
- 125.Colonna R, Mangia F. Mechanisms of amino acid uptake in cumulus-enclosed mouse oocytes. Biol Reprod. 1983;28(4):797–803. doi: 10.1095/biolreprod28.4.797. [DOI] [PubMed] [Google Scholar]
- 126.Eppig JJ, Pendola FL, Wigglesworth K, Pendola JK. Mouse oocytes regulate metabolic cooperativity between granulosa cells and oocytes: amino acid transport. Biol Reprod. 2005;73(2):351–357. doi: 10.1095/biolreprod.105.041798. [DOI] [PubMed] [Google Scholar]
- 127.Downs SM, Hudson ED. Energy substrates and the completion of spontaneous meiotic maturation. Zygote. 2000;8(4):339–351. doi: 10.1017/s0967199400001131. [DOI] [PubMed] [Google Scholar]
- 128.Bilodeau-Goeseels S. Effects of culture media and energy sources on the inhibition of nuclear maturation in bovine oocytes. Theriogenology. 2006;66(2):297–306. doi: 10.1016/j.theriogenology.2005.11.014. [DOI] [PubMed] [Google Scholar]
- 129.Kito S, Bavister BD. Gonadotropins, serum, and amino acids alter nuclear maturation, cumulus expansion, and oocyte morphology in hamster cumulus-oocyte complexes in vitro. Biol Reprod. 1997;56(5):1281–1289. doi: 10.1095/biolreprod56.5.1281. [DOI] [PubMed] [Google Scholar]
- 130.Songsasen N, Wildt DE. Oocyte biology and challenges in developing in vitro maturation systems in the domestic dog. Anim Reprod Sci. 2007;98(1–2):2–22. doi: 10.1016/j.anireprosci.2006.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Zheng P, Bavister BD, Ji WZ. Amino acid requirements for maturation of rhesus monkey (Macacca mulatta) oocytes in culture. Reproduction. 2002;124(4):515–522. doi: 10.1530/rep.0.1240515. [DOI] [PubMed] [Google Scholar]
- 132.Hong J, Lee E. Intrafollicular amino acid concentration and the effect of amino acids in a defined maturation medium on porcine oocyte maturation, fertilization, and preimplantation development. Theriogenology. 2007;68(5):728–735. doi: 10.1016/j.theriogenology.2007.06.002. [DOI] [PubMed] [Google Scholar]
- 133.Hoffmann EK, Lambert IH, Pedersen SF. Physiology of cell volume regulation in vertebrates. Physiol Rev. 2009;89(1):193–277. doi: 10.1152/physrev.00037.2007. [DOI] [PubMed] [Google Scholar]
- 134.Tartia AP, Rudraraju N, Richards T, Hammer MA, Talbot P, Baltz JM. Cell volume regulation is initiated in mouse oocytes after ovulation. Development. 2009;136(13):2247–2254. doi: 10.1242/dev.036756. [DOI] [PubMed] [Google Scholar]
- 135.Gesink Law DC, Maclehose RF, Longnecker MP. Obesity and time to pregnancy. Hum Reprod. 2007;22(2):414–420. doi: 10.1093/humrep/del400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Rich-Edwards JW, Goldman MB, Willett WC, Hunter DJ, Stampfer MJ, Colditz GA, Manson JE. Adolescent body mass index and infertility caused by ovulatory disorder. Am J Obstet Gynecol. 1994;171(1):171–177. doi: 10.1016/0002-9378(94)90465-0. [DOI] [PubMed] [Google Scholar]
- 137.Jungheim ES, Moley KH. Current knowledge of obesity’s effects in the pre- and periconceptional periods and avenues for future research. Am J Obstet Gynecol. 2010;203(6):525–530. doi: 10.1016/j.ajog.2010.06.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Becerra JE, Khoury MJ, Cordero JF, Erickson JD. Diabetes mellitus during pregnancy and the risks for specific birth defects: a population-based case-control study. Pediatrics. 1990;85(1):1–9. [PubMed] [Google Scholar]
- 139.Greene MF. Spontaneous abortions and major malformations in women with diabetes mellitus. Semin Reprod Endocrinol. 1999;17(2):127–136. doi: 10.1055/s-2007-1016220. [DOI] [PubMed] [Google Scholar]
- 140.Sadler TW, Hunter ES, 3rd, Balkan W, Horton WE., Jr Effects of maternal diabetes on embryogenesis. Am J Perinatol. 1988;5(4):319–326. doi: 10.1055/s-2007-999717. [DOI] [PubMed] [Google Scholar]
- 141.Doblado M, Moley KH. Glucose metabolism in pregnancy and embryogenesis. Curr Opin Endocrinol Diabetes Obes. 2007;14(6):488–493. doi: 10.1097/MED.0b013e3282f1cb92. [DOI] [PubMed] [Google Scholar]
- 142.Sinclair KD, Watkins AJ. Parental diet, pregnancy outcomes and offspring health: metabolic determinants in developing oocytes and embryos. Reprod Fertil Dev. 2013;26(1):99–114. doi: 10.1071/RD13290. [DOI] [PubMed] [Google Scholar]
- 143.Dupont J, Reverchon M, Bertoldo MJ, Froment P. Nutritional signals and reproduction. Mol Cell Endocrinol. 2014;382(1):527–537. doi: 10.1016/j.mce.2013.09.028. [DOI] [PubMed] [Google Scholar]
- 144.Walsh SW, Williams EJ, Evans AC. A review of the causes of poor fertility in high milk producing dairy cows. Anim Reprod Sci. 2011;123(3–4):127–138. doi: 10.1016/j.anireprosci.2010.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Rinaudo P, Wang E. Fetal programming and metabolic syndrome. Annu Rev Physiol. 2012;74:107–130. doi: 10.1146/annurev-physiol-020911-153245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Seli E, Babayev E, Collins SC, Nemeth G, Horvath TL (2014) Minireview: metabolism of female reproduction: regulatory mechanisms and clinical implications. Mol Endocrinol 28(6):790–804. doi:10.1210/me.2013-1413 [DOI] [PMC free article] [PubMed]
- 147.Mircea CN, Lujan ME, Pierson RA. Metabolic fuel and clinical implications for female reproduction. J Obstet Gynaecol Can. 2007;29(11):887–902. doi: 10.1016/S1701-2163(16)32661-5. [DOI] [PubMed] [Google Scholar]
- 148.Diamond MP, Moley KH, Pellicer A, Vaughn WK, DeCherney AH. Effects of streptozotocin- and alloxan-induced diabetes mellitus on mouse follicular and early embryo development. J Reprod Fertil. 1989;86(1):1–10. doi: 10.1530/jrf.0.0860001. [DOI] [PubMed] [Google Scholar]
- 149.Lea RG, McCracken JE, McIntyre SS, Smith W, Baird JD. Disturbed development of the preimplantation embryo in the insulin-dependent diabetic BB/E rat. Diabetes. 1996;45(11):1463–1470. doi: 10.2337/diab.45.11.1463. [DOI] [PubMed] [Google Scholar]
- 150.Moley KH, Chi MM, Mueckler MM. Maternal hyperglycemia alters glucose transport and utilization in mouse preimplantation embryos. Am J Physiol. 1998;275(1 Pt 1):E38–E47. doi: 10.1152/ajpendo.1998.275.1.E38. [DOI] [PubMed] [Google Scholar]
- 151.Moley KH, Vaughn WK, DeCherney AH, Diamond MP. Effect of diabetes mellitus on mouse pre-implantation embryo development. J Reprod Fertil. 1991;93(2):325–332. doi: 10.1530/jrf.0.0930325. [DOI] [PubMed] [Google Scholar]
- 152.Moley KH, Vaughn WK, Diamond MP. Manifestations of diabetes mellitus on mouse preimplantation development: effect of elevated concentration of metabolic intermediates. Hum Reprod. 1994;9(1):113–121. doi: 10.1093/oxfordjournals.humrep.a138298. [DOI] [PubMed] [Google Scholar]
- 153.Luzzo KM, Wang Q, Purcell SH, Chi M, Jimenez PT, Grindler N, Schedl T, Moley KH. High fat diet induced developmental defects in the mouse: oocyte meiotic aneuploidy and fetal growth retardation/brain defects. PLoS ONE. 2012;7(11):e49217. doi: 10.1371/journal.pone.0049217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Wyman A, Pinto AB, Sheridan R, Moley KH. One-cell zygote transfer from diabetic to nondiabetic mouse results in congenital malformations and growth retardation in offspring. Endocrinology. 2008;149(2):466–469. doi: 10.1210/en.2007-1273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Colton SA, Pieper GM, Downs SM. Altered meiotic regulation in oocytes from diabetic mice. Biol Reprod. 2002;67(1):220–231. doi: 10.1095/biolreprod67.1.220. [DOI] [PubMed] [Google Scholar]
- 156.Colton SA, Humpherson PG, Leese HJ, Downs SM. Physiological changes in oocyte-cumulus cell complexes from diabetic mice that potentially influence meiotic regulation. Biol Reprod. 2003;69(3):761–770. doi: 10.1095/biolreprod.102.013649. [DOI] [PubMed] [Google Scholar]
- 157.Maheshwari A, Stofberg L, Bhattacharya S. Effect of overweight and obesity on assisted reproductive technology–a systematic review. Hum Reprod Update. 2007;13(5):433–444. doi: 10.1093/humupd/dmm017. [DOI] [PubMed] [Google Scholar]
- 158.Wang JX, Davies M, Norman RJ. Body mass and probability of pregnancy during assisted reproduction treatment: retrospective study. BMJ. 2000;321(7272):1320–1321. doi: 10.1136/bmj.321.7272.1320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Brewer CJ, Balen AH. The adverse effects of obesity on conception and implantation. Reproduction. 2010;140(3):347–364. doi: 10.1530/REP-09-0568. [DOI] [PubMed] [Google Scholar]
- 160.Cardozo E, Pavone ME, Hirshfeld-Cytron JE. Metabolic syndrome and oocyte quality. Trends Endocrinol Metab. 2011;22(3):103–109. doi: 10.1016/j.tem.2010.12.002. [DOI] [PubMed] [Google Scholar]
- 161.Styne-Gross A, Elkind-Hirsch K, Scott RT., Jr Obesity does not impact implantation rates or pregnancy outcome in women attempting conception through oocyte donation. Fertil Steril. 2005;83(6):1629–1634. doi: 10.1016/j.fertnstert.2005.01.099. [DOI] [PubMed] [Google Scholar]
- 162.Wattanakumtornkul S, Damario MA, Stevens Hall SA, Thornhill AR, Tummon IS. Body mass index and uterine receptivity in the oocyte donation model. Fertil Steril. 2003;80(2):336–340. doi: 10.1016/s0015-0282(03)00595-8. [DOI] [PubMed] [Google Scholar]
- 163.Zenke U, Chetkowski RJ. Transfer and uterine factors are the major recipient-related determinants of success with donor eggs. Fertil Steril. 2004;82(4):850–856. doi: 10.1016/j.fertnstert.2004.03.057. [DOI] [PubMed] [Google Scholar]
- 164.Bellver J, Melo MA, Bosch E, Serra V, Remohi J, Pellicer A. Obesity and poor reproductive outcome: the potential role of the endometrium. Fertil Steril. 2007;88(2):446–451. doi: 10.1016/j.fertnstert.2006.11.162. [DOI] [PubMed] [Google Scholar]
- 165.Bellver J, Rossal LP, Bosch E, Zuniga A, Corona JT, Melendez F, Gomez E, Simon C, Remohi J, Pellicer A. Obesity and the risk of spontaneous abortion after oocyte donation. Fertil Steril. 2003;79(5):1136–1140. doi: 10.1016/s0015-0282(03)00176-6. [DOI] [PubMed] [Google Scholar]
- 166.Luke B, Brown MB, Stern JE, Missmer SA, Fujimoto VY, Leach R. Female obesity adversely affects assisted reproductive technology (ART) pregnancy and live birth rates. Hum Reprod. 2011;26(1):245–252. doi: 10.1093/humrep/deq306. [DOI] [PubMed] [Google Scholar]
- 167.Ramalho-Santos J, Varum S, Amaral S, Mota PC, Sousa AP, Amaral A. Mitochondrial functionality in reproduction: from gonads and gametes to embryos and embryonic stem cells. Hum Reprod Update. 2009;15(5):553–572. doi: 10.1093/humupd/dmp016. [DOI] [PubMed] [Google Scholar]
- 168.Wang Q, Ratchford AM, Chi MM, Schoeller E, Frolova A, Schedl T, Moley KH. Maternal diabetes causes mitochondrial dysfunction and meiotic defects in murine oocytes. Mol Endocrinol. 2009;23(10):1603–1612. doi: 10.1210/me.2009-0033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Ratchford AM, Chang AS, Chi MM, Sheridan R, Moley KH. Maternal diabetes adversely affects AMP-activated protein kinase activity and cellular metabolism in murine oocytes. Am J Physiol Endocrinol Metab. 2007;293(5):E1198–E1206. doi: 10.1152/ajpendo.00097.2007. [DOI] [PubMed] [Google Scholar]
- 170.Ou XH, Li S, Wang ZB, Li M, Quan S, Xing F, Guo L, Chao SB, Chen Z, Liang XW, Hou Y, Schatten H, Sun QY. Maternal insulin resistance causes oxidative stress and mitochondrial dysfunction in mouse oocytes. Hum Reprod. 2012;27(7):2130–2145. doi: 10.1093/humrep/des137. [DOI] [PubMed] [Google Scholar]
- 171.Van Blerkom J, Davis PW, Lee J. ATP content of human oocytes and developmental potential and outcome after in vitro fertilization and embryo transfer. Hum Reprod. 1995;10(2):415–424. doi: 10.1093/oxfordjournals.humrep.a135954. [DOI] [PubMed] [Google Scholar]
- 172.Quinn P, Wales RG. The relationships between the ATP content of preimplantation mouse embryos and their development in vitro during culture. J Reprod Fertil. 1973;35(2):301–309. doi: 10.1530/jrf.0.0350301. [DOI] [PubMed] [Google Scholar]
- 173.Wordeman L. How kinesin motor proteins drive mitotic spindle function: lessons from molecular assays. Semin Cell Dev Biol. 2010;21(3):260–268. doi: 10.1016/j.semcdb.2010.01.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Zhang X, Wu XQ, Lu S, Guo YL, Ma X. Deficit of mitochondria-derived ATP during oxidative stress impairs mouse MII oocyte spindles. Cell Res. 2006;16(10):841–850. doi: 10.1038/sj.cr.7310095. [DOI] [PubMed] [Google Scholar]
- 175.Liu L, Trimarchi JR, Smith PJ, Keefe DL. Mitochondrial dysfunction leads to telomere attrition and genomic instability. Aging Cell. 2002;1(1):40–46. doi: 10.1046/j.1474-9728.2002.00004.x. [DOI] [PubMed] [Google Scholar]
- 176.Kruger N, Tolic-Norrelykke IM. Association of mitochondria with spindle poles facilitates spindle alignment. Curr Biol. 2008;18(15):R646–R647. doi: 10.1016/j.cub.2008.06.069. [DOI] [PubMed] [Google Scholar]
- 177.Zeng HT, Ren Z, Yeung WS, Shu YM, Xu YW, Zhuang GL, Liang XY. Low mitochondrial DNA and ATP contents contribute to the absence of birefringent spindle imaged with PolScope in in vitro matured human oocytes. Hum Reprod. 2007;22(6):1681–1686. doi: 10.1093/humrep/dem070. [DOI] [PubMed] [Google Scholar]
- 178.Balaban RS, Nemoto S, Finkel T. Mitochondria, oxidants, and aging. Cell. 2005;120(4):483–495. doi: 10.1016/j.cell.2005.02.001. [DOI] [PubMed] [Google Scholar]
- 179.Dumollard R, Carroll J, Duchen MR, Campbell K, Swann K. Mitochondrial function and redox state in mammalian embryos. Semin Cell Dev Biol. 2009;20(3):346–353. doi: 10.1016/j.semcdb.2008.12.013. [DOI] [PubMed] [Google Scholar]
- 180.Igosheva N, Abramov AY, Poston L, Eckert JJ, Fleming TP, Duchen MR, McConnell J. Maternal diet-induced obesity alters mitochondrial activity and redox status in mouse oocytes and zygotes. PLoS ONE. 2010;5(4):e10074. doi: 10.1371/journal.pone.0010074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Wakefield SL, Lane M, Schulz SJ, Hebart ML, Thompson JG, Mitchell M. Maternal supply of omega-3 polyunsaturated fatty acids alter mechanisms involved in oocyte and early embryo development in the mouse. Am J Physiol Endocrinol Metab. 2008;294(2):E425–E434. doi: 10.1152/ajpendo.00409.2007. [DOI] [PubMed] [Google Scholar]
- 182.Qiao J, Feng HL. Extra- and intra-ovarian factors in polycystic ovary syndrome: impact on oocyte maturation and embryo developmental competence. Hum Reprod Update. 2011;17(1):17–33. doi: 10.1093/humupd/dmq032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Jimenez PT, Frolova AI, Chi MM, Grindler NM, Willcockson AR, Reynolds KA, Zhao Q, Moley KH. DHEA-mediated inhibition of the pentose phosphate pathway alters oocyte lipid metabolism in mice. Endocrinology. 2013;154(12):4835–4844. doi: 10.1210/en.2012-2140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Harris SE, Maruthini D, Tang T, Balen AH, Picton HM. Metabolism and karyotype analysis of oocytes from patients with polycystic ovary syndrome. Hum Reprod. 2010;25(9):2305–2315. doi: 10.1093/humrep/deq181. [DOI] [PubMed] [Google Scholar]
- 185.Walters KA, Allan CM, Handelsman DJ. Rodent models for human polycystic ovary syndrome. Biol Reprod. 2012;86(5):149. doi: 10.1095/biolreprod.111.097808. [DOI] [PubMed] [Google Scholar]
- 186.van Houten EL, Visser JA. Mouse models to study polycystic ovary syndrome: a possible link between metabolism and ovarian function? Reprod Biol. 2014;14(1):32–43. doi: 10.1016/j.repbio.2013.09.007. [DOI] [PubMed] [Google Scholar]
- 187.King AJ. The use of animal models in diabetes research. Br J Pharmacol. 2012;166(3):877–894. doi: 10.1111/j.1476-5381.2012.01911.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Wu LL, Dunning KR, Yang X, Russell DL, Lane M, Norman RJ, Robker RL. High-fat diet causes lipotoxicity responses in cumulus-oocyte complexes and decreased fertilization rates. Endocrinology. 2010;151(11):5438–5445. doi: 10.1210/en.2010-0551. [DOI] [PubMed] [Google Scholar]
- 189.Wu LL, Russell DL, Norman RJ, Robker RL. Endoplasmic reticulum (ER) stress in cumulus-oocyte complexes impairs pentraxin-3 secretion, mitochondrial membrane potential (DeltaPsi m), and embryo development. Mol Endocrinol. 2012;26(4):562–573. doi: 10.1210/me.2011-1362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Wu LL, Norman RJ, Robker RL. The impact of obesity on oocytes: evidence for lipotoxicity mechanisms. Reprod Fertil Dev. 2011;24(1):29–34. doi: 10.1071/RD11904. [DOI] [PubMed] [Google Scholar]
- 191.Jawerbaum A, Gonzalez ET, Carolina P, Debora S, Christian P, Gimeno MA. Diminished levels of prostaglandin E in type I diabetic oocyte-cumulus complexes. Influence of nitric oxide and superoxide dismutase. Reprod Fertil Dev. 1999;11(2):105–110. doi: 10.1071/rd99033. [DOI] [PubMed] [Google Scholar]
- 192.Jawerbaum A, Gonzalez ET, Faletti A, Novaro V, Vitullo A, Gimeno MA. Altered prostanoid production by cumulus-oocyte complexes in a rat model of non-insulin-dependent diabetes mellitus. Prostaglandins. 1996;52(3):209–219. doi: 10.1016/s0090-6980(96)00098-6. [DOI] [PubMed] [Google Scholar]
- 193.Rooke JA, Ainslie A, Watt RG, Alink FM, McEvoy TG, Sinclair KD, Garnsworthy PC, Webb R. Dietary carbohydrates and amino acids influence oocyte quality in dairy heifers. Reprod Fertil Dev. 2009;21(3):419–427. doi: 10.1071/rd08193. [DOI] [PubMed] [Google Scholar]
- 194.Sinclair KD, Kuran M, Gebbie FE, Webb R, McEvoy TG. Nitrogen metabolism and fertility in cattle: II. Development of oocytes recovered from heifers offered diets differing in their rate of nitrogen release in the rumen. J Anim Sci. 2000;78(10):2670–2680. doi: 10.2527/2000.78102670x. [DOI] [PubMed] [Google Scholar]
- 195.De Wit AA, Cesar ML, Kruip TA. Effect of urea during in vitro maturation on nuclear maturation and embryo development of bovine cumulus-oocyte-complexes. J Dairy Sci. 2001;84(8):1800–1804. doi: 10.3168/jds.S0022-0302(01)74618-8. [DOI] [PubMed] [Google Scholar]
- 196.Leroy JL, Van Soom A, Opsomer G, Goovaerts IG, Bols PE. Reduced fertility in high-yielding dairy cows: are the oocyte and embryo in danger? Part II. Mechanisms linking nutrition and reduced oocyte and embryo quality in high-yielding dairy cows. Reprod Domest Anim. 2008;43(5):623–632. doi: 10.1111/j.1439-0531.2007.00961.x. [DOI] [PubMed] [Google Scholar]
- 197.Leroy JL, Rizos D, Sturmey R, Bossaert P, Gutierrez-Adan A, Van Hoeck V, Valckx S, Bols PE. Intrafollicular conditions as a major link between maternal metabolism and oocyte quality: a focus on dairy cow fertility. Reprod Fertil Dev. 2011;24(1):1–12. doi: 10.1071/RD11901. [DOI] [PubMed] [Google Scholar]
- 198.Van Blerkom J. Mitochondria in human oogenesis and preimplantation embryogenesis: engines of metabolism, ionic regulation and developmental competence. Reproduction. 2004;128(3):269–280. doi: 10.1530/rep.1.00240. [DOI] [PubMed] [Google Scholar]
- 199.Agarwal A, Aponte-Mellado A, Premkumar BJ, Shaman A, Gupta S. The effects of oxidative stress on female reproduction: a review. Reprod Biol Endocrinol. 2012;10:49. doi: 10.1186/1477-7827-10-49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Turner N, Robker RL. Developmental programming of obesity and insulin resistance: does mitochondrial dysfunction in oocytes play a role? Mol Hum Reprod. 2014 doi: 10.1093/molehr/gau042. [DOI] [PubMed] [Google Scholar]
- 201.Rodgers RJ, Irving-Rodgers HF. Formation of the ovarian follicular antrum and follicular fluid. Biol Reprod. 2010;82(6):1021–1029. doi: 10.1095/biolreprod.109.082941. [DOI] [PubMed] [Google Scholar]
- 202.Robker RL, Akison LK, Bennett BD, Thrupp PN, Chura LR, Russell DL, Lane M, Norman RJ. Obese women exhibit differences in ovarian metabolites, hormones, and gene expression compared with moderate-weight women. J Clin Endocrinol Metab. 2009;94(5):1533–1540. doi: 10.1210/jc.2008-2648. [DOI] [PubMed] [Google Scholar]
- 203.Valckx SD, De Pauw I, De Neubourg D, Inion I, Berth M, Fransen E, Bols PE, Leroy JL. BMI-related metabolic composition of the follicular fluid of women undergoing assisted reproductive treatment and the consequences for oocyte and embryo quality. Hum Reprod. 2012;27(12):3531–3539. doi: 10.1093/humrep/des350. [DOI] [PubMed] [Google Scholar]
- 204.Russell JW, Golovoy D, Vincent AM, Mahendru P, Olzmann JA, Mentzer A, Feldman EL. High glucose-induced oxidative stress and mitochondrial dysfunction in neurons. FASEB J. 2002;16(13):1738–1748. doi: 10.1096/fj.01-1027com. [DOI] [PubMed] [Google Scholar]
- 205.Yu T, Sheu SS, Robotham JL, Yoon Y. Mitochondrial fission mediates high glucose-induced cell death through elevated production of reactive oxygen species. Cardiovasc Res. 2008;79(2):341–351. doi: 10.1093/cvr/cvn104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Graier WF, Malli R, Kostner GM. Mitochondrial protein phosphorylation: instigator or target of lipotoxicity? Trends Endocrinol Metab. 2009;20(4):186–193. doi: 10.1016/j.tem.2009.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Downs SM. The influence of glucose, cumulus cells, and metabolic coupling on ATP levels and meiotic control in the isolated mouse oocyte. Dev Biol. 1995;167(2):502–512. doi: 10.1006/dbio.1995.1044. [DOI] [PubMed] [Google Scholar]
- 208.Chang AS, Dale AN, Moley KH. Maternal diabetes adversely affects preovulatory oocyte maturation, development, and granulosa cell apoptosis. Endocrinology. 2005;146(5):2445–2453. doi: 10.1210/en.2004-1472. [DOI] [PubMed] [Google Scholar]
- 209.Ratchford AM, Esguerra CR, Moley KH. Decreased oocyte-granulosa cell gap junction communication and connexin expression in a type 1 diabetic mouse model. Mol Endocrinol. 2008;22(12):2643–2654. doi: 10.1210/me.2007-0495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Wang Q, Frolova AI, Purcell S, Adastra K, Schoeller E, Chi MM, Schedl T, Moley KH. Mitochondrial dysfunction and apoptosis in cumulus cells of type I diabetic mice. PLoS ONE. 2010;5(12):e15901. doi: 10.1371/journal.pone.0015901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Wang Q, Chi MM, Moley KH. Live imaging reveals the link between decreased glucose uptake in ovarian cumulus cells and impaired oocyte quality in female diabetic mice. Endocrinology. 2012;153(4):1984–1989. doi: 10.1210/en.2011-1815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Baird DT, Collins J, Egozcue J, Evers LH, Gianaroli L, Leridon H, Sunde A, Templeton A, Van Steirteghem A, Cohen J, Crosignani PG, Devroey P, Diedrich K, Fauser BC, Fraser L, Glasier A, Liebaers I, Mautone G, Penney G, Tarlatzis B. Fertility and ageing. Hum Reprod Update. 2005;11(3):261–276. doi: 10.1093/humupd/dmi006. [DOI] [PubMed] [Google Scholar]
- 213.Cheng PP, Xia JJ, Wang HL, Chen JB, Wang FY, Zhang Y, Huang X, Zhang QJ, Qi ZQ. Islet transplantation reverses the effects of maternal diabetes on mouse oocytes. Reproduction. 2011;141(4):417–424. doi: 10.1530/REP-10-0370. [DOI] [PubMed] [Google Scholar]
- 214.Machtinger R, Combelles CM, Missmer SA, Correia KF, Fox JH, Racowsky C. The association between severe obesity and characteristics of failed fertilized oocytes. Hum Reprod. 2012;27(11):3198–3207. doi: 10.1093/humrep/des308. [DOI] [PubMed] [Google Scholar]
- 215.Jones KT, Lane SI. Chromosomal, metabolic, environmental, and hormonal origins of aneuploidy in mammalian oocytes. Exp Cell Res. 2012;318(12):1394–1399. doi: 10.1016/j.yexcr.2012.02.012. [DOI] [PubMed] [Google Scholar]
- 216.Hassold T, Hunt P. To err (meiotically) is human: the genesis of human aneuploidy. Nat Rev Genet. 2001;2(4):280–291. doi: 10.1038/35066065. [DOI] [PubMed] [Google Scholar]
- 217.Ferguson-Smith AC. Genomic imprinting: the emergence of an epigenetic paradigm. Nat Rev Genet. 2011;12(8):565–575. doi: 10.1038/nrg3032. [DOI] [PubMed] [Google Scholar]
- 218.Ge ZJ, Liang XW, Guo L, Liang QX, Luo SM, Wang YP, Wei YC, Han ZM, Schatten H, Sun QY. Maternal diabetes causes alterations of DNA methylation statuses of some imprinted genes in murine oocytes. Biol Reprod. 2013;88(5):117. doi: 10.1095/biolreprod.112.105981. [DOI] [PubMed] [Google Scholar]
- 219.Ding L, Pan R, Huang X, Wang JX, Shen YT, Xu L, Zhang Y, Liu Y, He XQ, Yang XJ, Qi ZQ, Wang HL. Changes in histone acetylation during oocyte meiotic maturation in the diabetic mouse. Theriogenology. 2012;78(4):784–792. doi: 10.1016/j.theriogenology.2012.03.026. [DOI] [PubMed] [Google Scholar]
- 220.Ge ZJ, Luo SM, Lin F, Liang QX, Huang L, Wei YC, Hou Y, Han ZM, Schatten H, Sun QY. DNA methylation in oocytes and liver of female mice and their offspring: effects of high-fat-diet-induced obesity. Environ Health Perspect. 2014;122(2):159–164. doi: 10.1289/ehp.1307047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Minge CE, Bennett BD, Norman RJ, Robker RL. Peroxisome proliferator-activated receptor-gamma agonist rosiglitazone reverses the adverse effects of diet-induced obesity on oocyte quality. Endocrinology. 2008;149(5):2646–2656. doi: 10.1210/en.2007-1570. [DOI] [PubMed] [Google Scholar]
- 222.Li L, Zheng P, Dean J. Maternal control of early mouse development. Development. 2010;137(6):859–870. doi: 10.1242/dev.039487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223.Lu C, Thompson CB. Metabolic regulation of epigenetics. Cell Metab. 2012;16(1):9–17. doi: 10.1016/j.cmet.2012.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Kaelin WG, Jr, McKnight SL. Influence of metabolism on epigenetics and disease. Cell. 2013;153(1):56–69. doi: 10.1016/j.cell.2013.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Morrish F, Noonan J, Perez-Olsen C, Gafken PR, Fitzgibbon M, Kelleher J, VanGilst M, Hockenbery D. Myc-dependent mitochondrial generation of acetyl-CoA contributes to fatty acid biosynthesis and histone acetylation during cell cycle entry. J Biol Chem. 2010;285(47):36267–36274. doi: 10.1074/jbc.M110.141606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Takahashi H, McCaffery JM, Irizarry RA, Boeke JD. Nucleocytosolic acetyl-coenzyme a synthetase is required for histone acetylation and global transcription. Mol Cell. 2006;23(2):207–217. doi: 10.1016/j.molcel.2006.05.040. [DOI] [PubMed] [Google Scholar]
- 227.Feil R, Fraga MF. Epigenetics and the environment: emerging patterns and implications. Nat Rev Genet. 2011;13(2):97–109. doi: 10.1038/nrg3142. [DOI] [PubMed] [Google Scholar]
- 228.Bungard D, Fuerth BJ, Zeng PY, Faubert B, Maas NL, Viollet B, Carling D, Thompson CB, Jones RG, Berger SL. Signaling kinase AMPK activates stress-promoted transcription via histone H2B phosphorylation. Science. 2010;329(5996):1201–1205. doi: 10.1126/science.1191241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229.Pirola L, Balcerczyk A, Okabe J, El-Osta A. Epigenetic phenomena linked to diabetic complications. Nat Rev Endocrinol. 2010;6(12):665–675. doi: 10.1038/nrendo.2010.188. [DOI] [PubMed] [Google Scholar]
- 230.Sobinoff AP, Sutherland JM, McLaughlin EA. Intracellular signalling during female gametogenesis. Mol Hum Reprod. 2013;19(5):265–278. doi: 10.1093/molehr/gas065. [DOI] [PubMed] [Google Scholar]
- 231.Siddappa D, Kalaiselvanraja A, Bordignon V, Dupuis L, Gasperin BG, Roux PP, Duggavathi R. Mechanistic target of rapamycin (MTOR) signaling during ovulation in mice. Mol Reprod Dev. 2014;81(7):655–665. doi: 10.1002/mrd.22333. [DOI] [PubMed] [Google Scholar]
- 232.Reddy P, Liu L, Adhikari D, Jagarlamudi K, Rajareddy S, Shen Y, Du C, Tang W, Hamalainen T, Peng SL, Lan ZJ, Cooney AJ, Huhtaniemi I, Liu K. Oocyte-specific deletion of Pten causes premature activation of the primordial follicle pool. Science. 2008;319(5863):611–613. doi: 10.1126/science.1152257. [DOI] [PubMed] [Google Scholar]
- 233.Chen J, Torcia S, Xie F, Lin CJ, Cakmak H, Franciosi F, Horner K, Onodera C, Song JS, Cedars MI, Ramalho-Santos M, Conti M. Somatic cells regulate maternal mRNA translation and developmental competence of mouse oocytes. Nat Cell Biol. 2013;15(12):1415–1423. doi: 10.1038/ncb2873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Wang N, Luo LL, Xu JJ, Xu MY, Zhang XM, Zhou XL, Liu WJ, Fu YC. Obesity accelerates ovarian follicle development and follicle loss in rats. Metab, Clin Exp. 2014;63(1):94–103. doi: 10.1016/j.metabol.2013.09.001. [DOI] [PubMed] [Google Scholar]
- 235.Yaba A, Demir N. The mechanism of mTOR (mammalian target of rapamycin) in a mouse model of polycystic ovary syndrome (PCOS) J Ovar Res. 2012;5(1):38. doi: 10.1186/1757-2215-5-38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Niu Z, Lin N, Gu R, Sun Y, Feng Y (2014) Associations between insulin resistance, free fatty acids, and oocyte quality in polycystic ovary syndrome during in vitro fertilization. J Clin Endocrinol Metab:jc20133942. doi:10.1210/jc.2013-3942 [DOI] [PMC free article] [PubMed]
- 237.Ola SI, Sun QY. Factors influencing the biochemical markers for predicting mammalian oocyte quality. J Reprod Dev. 2012;58(4):385–392. doi: 10.1262/jrd.11-084h. [DOI] [PubMed] [Google Scholar]
- 238.Heller DT, Schultz RM. Ribonucleoside metabolism by mouse oocytes: metabolic cooperativity between the fully grown oocyte and cumulus cells. J Exp Zool. 1980;214(3):355–364. doi: 10.1002/jez.1402140314. [DOI] [PubMed] [Google Scholar]
- 239.Fourcroy JL. RNA synthesis in immature mouse oocyte development. J Exp Zool. 1982;219(2):257–266. doi: 10.1002/jez.1402190214. [DOI] [PubMed] [Google Scholar]
- 240.Downs SM, Coleman DL, Eppig JJ. Maintenance of murine oocyte meiotic arrest: uptake and metabolism of hypoxanthine and adenosine by cumulus cell-enclosed and denuded oocytes. Dev Biol. 1986;117(1):174–183. doi: 10.1016/0012-1606(86)90359-3. [DOI] [PubMed] [Google Scholar]
- 241.Racowsky C. The releasing action of calcium upon cyclic AMP-dependent meiotic arrest in hamster oocytes. J Exp Zool. 1986;239(2):263–275. doi: 10.1002/jez.1402390214. [DOI] [PubMed] [Google Scholar]
- 242.Bleil JD, Wassarman PM. Synthesis of zona pellucida proteins by denuded and follicle-enclosed mouse oocytes during culture in vitro. Proc Natl Acad Sci USA. 1980;77(2):1029–1033. doi: 10.1073/pnas.77.2.1029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.Chi MM, Moley KH. Single-embryo measurement of basal- and insulin-stimulated glucose uptake. Methods Mol Med. 2003;83:171–178. doi: 10.1385/1-59259-377-1:171. [DOI] [PubMed] [Google Scholar]
- 244.Chi MM, Hoehn A, Moley KH. Metabolic changes in the glucose-induced apoptotic blastocyst suggest alterations in mitochondrial physiology. Am J Phys Endocrinol Metab. 2002;283(2):E226–E232. doi: 10.1152/ajpendo.00046.2002. [DOI] [PubMed] [Google Scholar]
- 245.Nielsen J, Oliver S. The next wave in metabolome analysis. Trends Biotechnol. 2005;23(11):544–546. doi: 10.1016/j.tibtech.2005.08.005. [DOI] [PubMed] [Google Scholar]
- 246.Ferreira CR, Saraiva SA, Catharino RR, Garcia JS, Gozzo FC, Sanvido GB, Santos LF, Lo Turco EG, Pontes JH, Basso AC, Bertolla RP, Sartori R, Guardieiro MM, Perecin F, Meirelles FV, Sangalli JR, Eberlin MN. Single embryo and oocyte lipid fingerprinting by mass spectrometry. J Lipid Res. 2010;51(5):1218–1227. doi: 10.1194/jlr.D001768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Koek MM, Bakels F, Engel W, van den Maagdenberg A, Ferrari MD, Coulier L, Hankemeier T. Metabolic profiling of ultrasmall sample volumes with GC/MS: from microliter to nanoliter samples. Anal Chem. 2010;82(1):156–162. doi: 10.1021/ac9015787. [DOI] [PubMed] [Google Scholar]
- 248.Ferreira CR, Eberlin LS, Hallett JE, Cooks RG. Single oocyte and single embryo lipid analysis by desorption electrospray ionization mass spectrometry. J Mass Spectrom. 2012;47(1):29–33. doi: 10.1002/jms.2022. [DOI] [PubMed] [Google Scholar]
- 249.Seli E, Robert C, Sirard MA. OMICS in assisted reproduction: possibilities and pitfalls. Mol Hum Reprod. 2010;16(8):513–530. doi: 10.1093/molehr/gaq041. [DOI] [PubMed] [Google Scholar]
- 250.Lindon JC, Holmes E, Bollard ME, Stanley EG, Nicholson JK. Metabonomics technologies and their applications in physiological monitoring, drug safety assessment and disease diagnosis. Biomarkers. 2004;9(1):1–31. doi: 10.1080/13547500410001668379. [DOI] [PubMed] [Google Scholar]
- 251.Houghton FD, Hawkhead JA, Humpherson PG, Hogg JE, Balen AH, Rutherford AJ, Leese HJ. Non-invasive amino acid turnover predicts human embryo developmental capacity. Hum Reprod. 2002;17(4):999–1005. doi: 10.1093/humrep/17.4.999. [DOI] [PubMed] [Google Scholar]
- 252.Brison DR, Houghton FD, Falconer D, Roberts SA, Hawkhead J, Humpherson PG, Lieberman BA, Leese HJ. Identification of viable embryos in IVF by non-invasive measurement of amino acid turnover. Hum Reprod. 2004;19(10):2319–2324. doi: 10.1093/humrep/deh409. [DOI] [PubMed] [Google Scholar]
- 253.Katz-Jaffe MG, Gardner DK. Embryology in the era of proteomics. Theriogenology. 2007;68(Suppl 1):S125–S130. doi: 10.1016/j.theriogenology.2007.03.014. [DOI] [PubMed] [Google Scholar]
- 254.Seli E, Sakkas D, Scott R, Kwok SC, Rosendahl SM, Burns DH. Noninvasive metabolomic profiling of embryo culture media using Raman and near-infrared spectroscopy correlates with reproductive potential of embryos in women undergoing in vitro fertilization. Fertil Steril. 2007;88(5):1350–1357. doi: 10.1016/j.fertnstert.2007.07.1390. [DOI] [PubMed] [Google Scholar]
- 255.Frey R, Muck W, Kirschbaum N, Kratzschmar J, Weimann G, Wensing G. Riociguat (BAY 63-2521) and warfarin: a pharmacodynamic and pharmacokinetic interaction study. J Clin Pharmacol. 2011;51(7):1051–1060. doi: 10.1177/0091270010378119. [DOI] [PubMed] [Google Scholar]
- 256.Gosden RG, Sadler IH, Reed D, Hunter RH. Characterization of ovarian follicular fluids of sheep, pigs and cows using proton nuclear magnetic resonance spectroscopy. Experientia. 1990;46(10):1012–1015. doi: 10.1007/BF01940658. [DOI] [PubMed] [Google Scholar]
- 257.Bentley JD, Jentoft JE, Foreman D, Ambrose D. 31P nuclear magnetic resonance (NMR) identification of sugar phosphates in isolated rat ovarian follicular granulosa cells and the effects of follicle-stimulating hormone. Mol Cell Endocrinol. 1990;73(2–3):179–185. doi: 10.1016/0303-7207(90)90131-q. [DOI] [PubMed] [Google Scholar]
- 258.Bender K, Walsh S, Evans AC, Fair T, Brennan L. Metabolite concentrations in follicular fluid may explain differences in fertility between heifers and lactating cows. Reproduction. 2010;139(6):1047–1055. doi: 10.1530/REP-10-0068. [DOI] [PubMed] [Google Scholar]
- 259.Zhang CM, Zhao Y, Li R, Yu Y, Yan LY, Li L, Liu NN, Liu P, Qiao J. Metabolic heterogeneity of follicular amino acids in polycystic ovary syndrome is affected by obesity and related to pregnancy outcome. BMC Pregnancy Childbirth. 2014;14:11. doi: 10.1186/1471-2393-14-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 260.Hemmings KE, Leese HJ, Picton HM. Amino acid turnover by bovine oocytes provides an index of oocyte developmental competence in vitro. Biol Reprod. 2012;86(5):165. doi: 10.1095/biolreprod.111.092585. [DOI] [PubMed] [Google Scholar]
- 261.Ahlstrom A, Wikland M, Rogberg L, Barnett JS, Tucker M, Hardarson T. Cross-validation and predictive value of near-infrared spectroscopy algorithms for day-5 blastocyst transfer. Reprod Biomed Online. 2011 doi: 10.1016/j.rbmo.2011.01.009. [DOI] [PubMed] [Google Scholar]
- 262.Scott R, Seli E, Miller K, Sakkas D, Scott K, Burns DH. Noninvasive metabolomic profiling of human embryo culture media using Raman spectroscopy predicts embryonic reproductive potential: a prospective blinded pilot study. Fertil Steril. 2008;90(1):77–83. doi: 10.1016/j.fertnstert.2007.11.058. [DOI] [PubMed] [Google Scholar]
- 263.Hardarson T, Ahlstrom A, Rogberg L, Botros L, Hillensjo T, Westlander G, Sakkas D, Wikland M. Non-invasive metabolomic profiling of day 2 and 5 embryo culture medium: a prospective randomized trial. Hum Reprod. 2012;27(1):89–96. doi: 10.1093/humrep/der373. [DOI] [PubMed] [Google Scholar]
- 264.Vergouw CG, Kieslinger DC, Kostelijk EH, Botros LL, Schats R, Hompes PG, Sakkas D, Lambalk CB. Day 3 embryo selection by metabolomic profiling of culture medium with near-infrared spectroscopy as an adjunct to morphology: a randomized controlled trial. Hum Reprod. 2012;27(8):2304–2311. doi: 10.1093/humrep/des175. [DOI] [PubMed] [Google Scholar]
- 265.el Mouatassim S, Hazout A, Bellec V, Menezo Y. Glucose metabolism during the final stage of human oocyte maturation: genetic expression of hexokinase, glucose phosphate isomerase and phosphofructokinase. Zygote. 1999;7(1):45–50. doi: 10.1017/s0967199499000386. [DOI] [PubMed] [Google Scholar]
- 266.Tsutsumi O, Yano T, Satoh K, Mizuno M, Kato T. Studies of hexokinase activity in human and mouse oocyte. Am J Obstet Gynecol. 1990;162(5):1301–1304. doi: 10.1016/0002-9378(90)90041-5. [DOI] [PubMed] [Google Scholar]
- 267.Peterson AC, Wong GG. Genetic regulation of glucose phosphate isomerase in mouse oocytes. Nature. 1978;276(5685):267–269. doi: 10.1038/276267a0. [DOI] [PubMed] [Google Scholar]
- 268.Tsutsumi O, Yano T, Taketani Y. Phosphofructokinase activity as a measure of maturation of rat oocytes developed in vivo and in vitro. Horm Res. 1994;41(Suppl 1):63–67. doi: 10.1159/000183945. [DOI] [PubMed] [Google Scholar]
- 269.Egerszegi I, Alm H, Ratky J, Heleil B, Brussow KP, Torner H. Meiotic progression, mitochondrial features and fertilisation characteristics of porcine oocytes with different G6PDH activities. Reprod Fertil Dev. 2010;22(5):830–838. doi: 10.1071/RD09140. [DOI] [PubMed] [Google Scholar]
- 270.Silva DS, Rodriguez P, Galuppo A, Arruda NS, Rodrigues JL. Selection of bovine oocytes by brilliant cresyl blue staining: effect on meiosis progression, organelle distribution and embryo development. Zygote. 2013;21(3):250–255. doi: 10.1017/S0967199411000487. [DOI] [PubMed] [Google Scholar]
- 271.Torner H, Ghanem N, Ambros C, Holker M, Tomek W, Phatsara C, Alm H, Sirard MA, Kanitz W, Schellander K, Tesfaye D. Molecular and subcellular characterisation of oocytes screened for their developmental competence based on glucose-6-phosphate dehydrogenase activity. Reproduction. 2008;135(2):197–212. doi: 10.1530/REP-07-0348. [DOI] [PubMed] [Google Scholar]
- 272.Lequarre AS, Grisart B, Moreau B, Schuurbiers N, Massip A, Dessy F. Glucose metabolism during bovine preimplantation development: analysis of gene expression in single oocytes and embryos. Mol Reprod Dev. 1997;48(2):216–226. doi: 10.1002/(SICI)1098-2795(199710)48:2<216::AID-MRD9>3.0.CO;2-V. [DOI] [PubMed] [Google Scholar]
- 273.de Schepper GG, van Noorden CJ, Tas J, James J. Cytochemical determination of glucose-6-phosphate dehydrogenase (G6PDH) activity in mouse oocytes with the use of a polyacrylamide carrier. Cell Biol Int Rep. 1982;6(7):651. doi: 10.1016/0309-1651(82)90126-6. [DOI] [PubMed] [Google Scholar]
- 274.Yazigi RA, Chi MM, Mastrogiannis DS, Strickler RC, Yang VC, Lowry OH. Enzyme activities and maturation in unstimulated and exogenous gonadotropin-stimulated human oocytes. Am J Physiol. 1993;264(4 Pt 1):C951–C955. doi: 10.1152/ajpcell.1993.264.4.C951. [DOI] [PubMed] [Google Scholar]
- 275.Acevedo N, Wang X, Dunn RL, Smith GD. Glycogen synthase kinase-3 regulation of chromatin segregation and cytokinesis in mouse preimplantation embryos. Mol Reprod Dev. 2007;74(2):178–188. doi: 10.1002/mrd.20495. [DOI] [PubMed] [Google Scholar]
- 276.Wang X, Liu XT, Dunn R, Ohl DA, Smith GD. Glycogen synthase kinase-3 regulates mouse oocyte homologue segregation. Mol Reprod Dev. 2003;64(1):96–105. doi: 10.1002/mrd.10213. [DOI] [PubMed] [Google Scholar]
- 277.Uzbekova S, Salhab M, Perreau C, Mermillod P, Dupont J. Glycogen synthase kinase 3B in bovine oocytes and granulosa cells: possible involvement in meiosis during in vitro maturation. Reproduction. 2009;138(2):235–246. doi: 10.1530/REP-09-0136. [DOI] [PubMed] [Google Scholar]
- 278.Williams SA, Stanley P. Mouse fertility is enhanced by oocyte-specific loss of core 1-derived O-glycans. FASEB J. 2008;22(7):2273–2284. doi: 10.1096/fj.07-101709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 279.Iwata N, Inazu N, Satoh T. The purification and properties of aldose reductase from rat ovary. Arch Biochem Biophys. 1990;282(1):70–77. doi: 10.1016/0003-9861(90)90088-g. [DOI] [PubMed] [Google Scholar]
- 280.Kaneko T, Iuchi Y, Takahashi M, Fujii J. Colocalization of polyol-metabolizing enzymes and immunological detection of fructated proteins in the female reproductive system of the rat. Histochem Cell Biol. 2003;119(4):309–315. doi: 10.1007/s00418-003-0516-5. [DOI] [PubMed] [Google Scholar]
- 281.Colton SA, Downs SM. Potential role for the sorbitol pathway in the meiotic dysfunction exhibited by oocytes from diabetic mice. J Exp Zool Part A Comp Exp Biol. 2004;301(5):439–448. doi: 10.1002/jez.a.20070. [DOI] [PubMed] [Google Scholar]
- 282.Cetica P, Pintos L, Dalvit G, Beconi M. Involvement of enzymes of amino acid metabolism and tricarboxylic acid cycle in bovine oocyte maturation in vitro. Reproduction. 2003;126(6):753–763. [PubMed] [Google Scholar]
- 283.Wright RW, Jr, Cupps PT, Watson JG, Chaykin S. Lactate dehydrogenase isoenzyme patterns in oocytes, unfertilized eggs and embryos from mice and cattle. J Anim Sci. 1976;43(3):613–616. doi: 10.2527/jas1976.433613x. [DOI] [PubMed] [Google Scholar]
- 284.Cetica PD, Pintos LN, Dalvit GC, Beconi MT. Effect of lactate dehydrogenase activity and isoenzyme localization in bovine oocytes and utilization of oxidative substrates on in vitro maturation. Theriogenology. 1999;51(3):541–550. doi: 10.1016/s0093-691x(99)00008-4. [DOI] [PubMed] [Google Scholar]
- 285.Mangia F, Epstein CJ. Biochemical studies of growing mouse oocytes: preparation of oocytes and analysis of glucose-6-phosphate dehydrogenase and lactate dehydrogenase activities. Dev Biol. 1975;45(2):211–220. doi: 10.1016/0012-1606(75)90061-5. [DOI] [PubMed] [Google Scholar]
- 286.Yuan Y, Ida JM, Paczkowski M, Krisher RL. Identification of developmental competence-related genes in mature porcine oocytes. Mol Reprod Dev. 2011;78(8):565–575. doi: 10.1002/mrd.21351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 287.Okawara S, Hamano S, Tetsuka M. Bovine oocytes and early embryos express mRNA encoding glycerol kinase but addition of glycerol to the culture media interferes with oocyte maturation. J Reprod Dev. 2009;55(2):177–182. doi: 10.1262/jrd.20103. [DOI] [PubMed] [Google Scholar]
- 288.Gentile L, Monti M, Sebastiano V, Merico V, Nicolai R, Calvani M, Garagna S, Redi CA, Zuccotti M. Single-cell quantitative RT-PCR analysis of Cpt1b and Cpt2 gene expression in mouse antral oocytes and in preimplantation embryos. Cytogenet Genome Res. 2004;105(2–4):215–221. doi: 10.1159/000078191. [DOI] [PubMed] [Google Scholar]
- 289.Eliyahu E, Shtraizent N, Shalgi R, Schuchman EH. Construction of conditional acid ceramidase knockout mice and in vivo effects on oocyte development and fertility. Cell Physiol Biochem. 2012;30(3):735–748. doi: 10.1159/000341453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 290.Eliyahu E, Shtraizent N, Martinuzzi K, Barritt J, He X, Wei H, Chaubal S, Copperman AB, Schuchman EH. Acid ceramidase improves the quality of oocytes and embryos and the outcome of in vitro fertilization. FASEB J. 2010;24(4):1229–1238. doi: 10.1096/fj.09-145508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 291.Zhang CH, Qian WP, Qi ST, Ge ZJ, Min LJ, Zhu XL, Huang X, Liu JP, Ouyang YC, Hou Y, Schatten H, Sun QY. Maternal diabetes causes abnormal dynamic changes of endoplasmic reticulum during mouse oocyte maturation and early embryo development. Reprod Biol Endocrinol. 2013;11:31. doi: 10.1186/1477-7827-11-31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 292.Jawerbaum A, Gonzalez ET, Novaro V, Faletti A, Gimeno MA. Nitric oxide mediates increased prostaglandin E production by oocyte-cumulus complexes in the non-insulin-dependent diabetic rat. Reprod Fertil Dev. 1998;10(2):185–190. doi: 10.1071/r97069. [DOI] [PubMed] [Google Scholar]
- 293.Kim K, Kim CH, Moley KH, Cheon YP. Disordered meiotic regulation of oocytes by duration of diabetes mellitus in BBdp rat. Reprod Sci. 2007;14(5):467–474. doi: 10.1177/1933719107306228. [DOI] [PubMed] [Google Scholar]
- 294.Chattopadhayay R, Ganesh A, Samanta J, Jana SK, Chakravarty BN, Chaudhury K. Effect of follicular fluid oxidative stress on meiotic spindle formation in infertile women with polycystic ovarian syndrome. Gynecol Obstet Invest. 2010;69(3):197–202. doi: 10.1159/000270900. [DOI] [PubMed] [Google Scholar]
- 295.Tian X, Diaz FJ. Acute dietary zinc deficiency before conception compromises oocyte epigenetic programming and disrupts embryonic development. Dev Biol. 2013;376(1):51–61. doi: 10.1016/j.ydbio.2013.01.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 296.Watkins AJ, Wilkins A, Cunningham C, Perry VH, Seet MJ, Osmond C, Eckert JJ, Torrens C, Cagampang FR, Cleal J, Gray WP, Hanson MA, Fleming TP. Low protein diet fed exclusively during mouse oocyte maturation leads to behavioural and cardiovascular abnormalities in offspring. J Physiol. 2008;586(8):2231–2244. doi: 10.1113/jphysiol.2007.149229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 297.Ferguson EM, Slevin J, Hunter MG, Edwards SA, Ashworth CJ. Beneficial effects of a high fibre diet on oocyte maturity and embryo survival in gilts. Reproduction. 2007;133(2):433–439. doi: 10.1530/REP-06-0018. [DOI] [PubMed] [Google Scholar]
- 298.Weaver AC, Kelly JM, Kind KL, Gatford KL, Kennaway DJ, Herde PJ, van Wettere WH. Oocyte maturation and embryo survival in nulliparous female pigs (gilts) is improved by feeding a lupin-based high-fibre diet. Reprod Fertil Dev. 2013;25(8):1216–1223. doi: 10.1071/RD12329. [DOI] [PubMed] [Google Scholar]
- 299.Tian X, Diaz FJ. Zinc depletion causes multiple defects in ovarian function during the periovulatory period in mice. Endocrinology. 2012;153(2):873–886. doi: 10.1210/en.2011-1599. [DOI] [PMC free article] [PubMed] [Google Scholar]