

Abstract
A patient recently presented with long-standing severe itching originating from lichen sclerosus et atrophicus at the vulva area. We successively treated her using a topical formulation of TRPM8 agonist which produces a cooling sensation. The cooling sensation, an afferent sensory perception in various skin neuronal pathways, could be a useful mechanism to relieve an itchy sensation in various skin disorders. Mechanoreceptors are related to touch vibration and pressure sensations and have a special morphology where the nerve endings are optimized to receive sensory inputs. However, unmyelinated nerve fibers are believed to transfer nociception such as pain, itching, stinging and burning derived from chemical or thermal stimuli. Among them, the function of transient receptor potential (TRP) receptors is very unique because they transfer the signal not only in the neuronal perception pathway but also in the cellular signal pathway where it appears as an ion channel. This review explains the cooling sensation of skin which has not been evaluated thoroughly, and provides insights for further clinical applications.
Keywords: Skin, Thermosensation, TRPs, Cooling, TRPM8
INTRODUCTION
The skin forms a protective layer around the body against physical, chemical and thermal environmental challenges. In addition to providing a physical barrier, the skin is considered a sensory organ that allows the body to perceive harmful external stimuli and appropriate behavior or body movement can be initiated thereafter.
The peripheral nervous system governs the broadly defined sense of touch perception, communicating with the central nervous system regarding the external environment, allowing for conscious sensations of balance and coordination, pressure and vibration, pain and temperature [1,2]. Sensory nerves are innervated in the entire skin tissue as well as subcutaneous fat layer [1]. Most nerve fibers and endings are found in the mid-dermis and the papillary dermis. In the epidermis, sensory nerves are linked to keratinocytes, melanocytes, Langerhans cells (LC) and Merkel cells. The epidermal nerves consist of free nerve endings or nerve organs. In the dermis, there are free sensory nerve endings such as Pinkus discs, Ruffini, Meissner, Krause and Vater-Pacini corpuscles [1–4].
Peripheral sensory nerves can be classified on the basis of the degree of myelination and the velocity at which action potentials travel through afferent fibers with the fast conducting, myelinated (large) Aα fibers, Aβ fibers, Aδ fibers and the slow conducting, unmyelinated (small) C fibers. Among them, Aβ and Aδ fibers are mostly mechanical sensitive afferents (type I) localized on hairy and glabrous skin. A subpopulation of Aδ fibers on hairy skin are believed to be mechanically insensitive (type II). Aδ fibers constitute approximately 80% of primary sensory nerves sprouting from the dorsal root ganglion (DRG), whereas C fibers constitute approximately 20% of the primary afferents [3–5]. C fibers are either polymodal nociceptors which can respond to chemical or mechanical stimuli and temperature changes, or more specialized perception. Among human peripheral nerves, 45% of the cutaneous afferent nerves belong to a subset of sensory nerves that are both mechano- and heat-responsive C fibers. The C fibers are the predominant nerve pathway for thermoperception of warmth. A certain subpopulation of Aδ fibers respond to gentle cooling, whereas selective C fibers become activated during noxious cold. When we lightly touch with our fingertips, the Aδ fibers respond because they are associated with a variety of cells including keratinocytes, Merkel cells and hair follicles. In addition, the C fibers are known for nociceptor action terminated at the target tissue as a free nerve ending. Similarly, specific receptor distribution and biochemical diversity of different sensory nerve subtypes exists, which is important for various perceptions [6,7].
The information detected from the peripheral sensory nerves is transmitted to the DRG and cranial nerve ganglia such as the trigeminal ganglion. DRG are a cluster of sensory neuronal cell bodies located in the vertebral column lateral to the spinal cord. A functionally specialized zone exists at the DRG neurons that can be partitioned according to specific sensory modalities. Based on the stimulus detected, DRG neurons are functionally classified as proprioceptors, low-threshold mechanoreceptors, and cells that sense pain, itch and/or temperature. The DRG neuronal branch relays information through the dorsal horn of the spinal cord to the central nervous system to recognize the type of sensation [6–9].
THERMOSENSORY PERCEPTION
Thermosensation is a sensory modality of the skin. Thermal stimuli applied to the skin induce a variable degree of sensory perception. When decreasing the stimuli temperature, the quality of sensation may change from cool to cold, from icy cold to pain. Similarly, increasing the stimuli temperature can cause a change in the sensations from warmth, to heat then to sharp or dull pain. The reported temperatures that cause these different sensations vary considerably. This variability can be explained by differences in the characteristics of the stimulus used, experimental conditions employed, and individual background differences (previous experiences of pain, ethnicity, sex, social status or religion).
Cutaneous thermosensation is mediated by a variety of primary afferent nerve fibers that transduce, encode and transmit thermal information [7–11]. There are 2 populations of thermosensory fibers, one responding to warmth and the other to cold. Specific cutaneous cold and warm receptors have been defined as slowly conducting units that exhibit a steady-state discharge at constant skin temperature and a dynamic response to temperature changes [11]. Cold-specific and warm-specific receptors can be distinguished from nociceptors that respond to noxious low and high temperatures (<20°C and >45°C), and also from thermo-sensitive mechanoreceptors [11,12].
In general, the perception of non-painful, cool temperature occurs when the skin is cooled as little as 1°C from normal body temperature. In a fiber recording study, temperatures in the range of 30-15°C can activate both Aδ and C fibers. These cold-sensitive afferents fire continuously at normal body temperature, but under cold stimuli the rate of firing increases, while warm temperatures reduce (or antagonize) this activity. However, the peak responsiveness can vary when the temperatures fall below either 25 or 15°C. Once the temperature approaches 15°C, the perception of cold as well as pain is evoked, described as burning, aching, and pricking sensations [13]. Further discussion regarding each innocuous and noxious cold sensation is described below. This delicate thermosensing process is also related to specific receptor proteins that are located within the free nerve endings in the skin, known as transient receptor potential (TRP) [14].
1. TRP
Advances in understanding the temperature sensation have emerged from the cloning and characterization of TRP ion channels whose activity depends on the temperature of their environment. Activation of TRP channels depolarize cells from the resting membrane potential and shortens action potential duration. Most TRP channels are cation-non-selective and permeable to the signal transduction element Ca2+. Each of these receptors operates over a specific temperature range, thereby providing a potential molecular basis for thermosensation. These specialized thermal receptors are embedded in the terminals of afferent fibers which end as free nerve endings in the skin [14,15].
TRP cation channels are unique cellular sensors characterized by a promiscuous activation mechanism. TRPs are classified according to their primary amino acid sequence rather than selectivity or ligand affinity, because their properties are heterogenous and their regulation is complex. TRP channels are membrane proteins with 6 putative trans-membrane (TM) spans and a cation-permeable pore region formed by a short hydrophobic stretch between TM5 and TM6 (Fig. 1). TRP proteins are essentially cation-permeable ion channels sensitive to a remarkable range of stimuli [15].
Fig. 1.
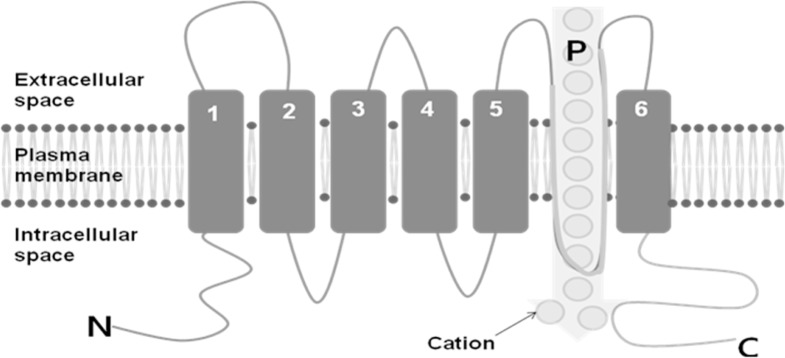
Schematic image of the six transmembrane spans (TM 1-6) of a transient receptor potential (TRP) protein. The pore loop (P) is located between TM5 and 6, and the intracellular amino (N) and carboxy (C) termini are also seen.
In humans, 28 different TRP channels have been identified and are grouped into 6 families. Of these, members of 3 families, i.e. the vanilloid TRP channels (TRPV), the melastatin or long TRP channels (TRPM), and the ankyrin transmembrane protein channels (TRPA) are of particular interest as thermoreceptors. Representative mammalian TRP channels are gated by heat and capsaicin (>43°C; TRPV1), noxious heat (>52°C; TRPV2), and cooling (<22°C; TRPM8) [15] (Table 1).
Table 1.
Transient receptor potential (TRP) channels involved in thermosensation
| Name | Other name | Temperature sensitivity | Non thermal agonists | Blockers | Tissue distribution |
|---|---|---|---|---|---|
| Trpv1 | Vr1 | ≥42°C | Capsaicin, lipoxygenase, acidic PH, resiniferatoxin, NADA, anandamide and ethanol | Ruthenium red, capsazepine | PNS, brain, spinal cord, skin, tongue, bladder |
| Trpv2 | Vr1 | ≥52°C | Growth factors (mouse) | Ruthenium red | PNS, brain, spinal cord, widely expressed |
| Trpv3 | Vr3 | >33°C | Hypotonic, 4-α phorbol | Ruthenium red | Skin, PNS (human) |
| Trpv4 | OTRPC4 VR-OAC Trp12 Vrl2 |
∼27-42°C | Menthol, icilin, eucalyptol | Ruthenium red, gadolinium | Kidney, PNS, skin, inner ear, brain, liver, trachea, heart, skin, hypothalamus, fat, endothelium |
| Trpm8 | CMR1 | ≤25°C | Icilin | PNS*, prostate (human) | |
| TRPA1 (Anktm1) | ≤17°C | Ruthenium red | PNS |
PNS*: peripheral nervous system.
TRPV1, vanilloid receptor 1, is the most well-known and characterized TRP channel which is activated by noxious stimuli such as capsaicin and excessive heat and pH below 5. TRPV1 is predominantly expressed in sensory neurons and believed to play a crucial role in temperature sensing and nociception [16]. Therefore, they were used as a molecular target for pain treatment and pruritus over the last several decades.
While TRPV1 and TRPV2 are established as neuronal thermosensors for heat sensation, other TRPV subfamily members, TRPV3 and TRPV4 are also involved in the sensation of warmth. In regards to cool and cold sensation, TRPA1 and transient receptor potential melastatin 8 (TRPM8) perform a critical role. TRPA1 is activated to mediate noxious cold sensation while TRPM8 mediates cool and fresh mint-like sensation. Recently, the cooling sensation generated especially by TRPM8 and/or cold and menthol receptor 1 (CMR1) has attracted attention for its potential usefulness.
2. Innocuous cold sensation and TRPM8
At steady state temperatures, cold fibers have a characteristic stimulus response function which is bell-shaped, with a maximal steady activity between 20 and 30oC and less activity at either lower or higher temperatures. The reported temperature range of cold fiber actions varies. At constant temperatures above 40°C or below 17°C, cold fibers maintain a very low frequency discharge or become silent. However, some cold fibers can also be activated by high temperatures in the noxious range, and this may be the basis for the paradoxical cold sensation. Cold afferents respond vigorously when the skin is actively cooled down. However, these dynamic responses are rather transient [18–20].
C fibers responding to innocuous cold temperatures have been found in human skin [20,21]. In addition, high-threshold cold receptors (HCRs) have previously been identified. These afferents are believed to be sensitive to cold temperatures below 27°C, but do not respond to mechanical or heat stimuli and have conduction velocities in the low range of Aδ and C fibers [17]. Rapid cooling of the skin is also able to activate unmyelinated, low threshold mechanoreceptors (CLTMs). These C fibers are non-responsive to intense heat stimuli and respond vigorously to gentle touch [21]. This response to cooling is only transient, and relatively rapid decreasing temperatures are required for long effective activation. Several studies suggest cold sensitivity can also be found in large myelinated afferents which respond vigorously to mechanical stimuli [20–22].
TRP channels sense the innocuous cold through the TRPM8 ion channel [23], which is specifically expressed in the DRG and trigeminal ganglion neurons with small nerve fibers, probably C fibers. Based on mice studies, the role of TRPM8 in thermosensation is well-established [22]. TRPM8 is activated by temperatures below 26°C, and is expressed in approximately 15% of small DRG neurons [23]. TRPM8 is activated by agents known to produce the sensation of cold in humans; the well-known agonists are menthol, eucalyptol and icilin. TRPM8 channels also have a relationship with somatosensation and nociception. As injury impacts thermosensation, sensory reaction showed a heightened sensitivity to temperatures in the innocuous range (allodynia) and in the already painful range (hyperalgesia) [24,25]. Colburn et al. assessed the involvement of TRPM8 in injury-evoked hypersensitivity to cold, and suggested that TRPM8 cooling sensation may modulate the majority of allodynia and hyperalgesia [26]. In addition, numerous studies have demonstrated TRPM8 expressing afferents are related to analgesic effects. The analgesic role mediated by TRPM8 was confirmed with TRPM8-null mice [27]. TRPM8 is also expressed in the prostate, lung and bladder tissues [25].
3. Noxious temperatures
Cold temperatures can produce the pain sensation with temperatures between 10 and 15°C in glabrous skin and approximately 18°C in hairy skin [28]. Noxious cold stimuli cause distinct sensations such as pricking, burning, aching and even heat [29], suggesting different fiber populations exist and are activated in the cold sensation. Mechanosensitive Aδ afferent fibers (A-MSA), including those sensitive to heat stimuli, can be also activated by noxious cold [30]. Instead of signaling the intensity of a cold stimulus, A-MSA fibers might encode the pricking sensation associated with noxious cold stimuli; cold-induced pricking sensation is largely reduced when Aδ fibers are blocked [31].
In addition to Aδ fibers, a subset of C fiber nociceptors can be excited by noxious cold and may therefore contribute to the sensation of cold pain [32]. Similar to Aδ fibers, most of the cold-sensitive C fibers are believed to be poly-modal in nature. Although C fibers are also responsive to mechanical and heat stimuli, their threshold temperatures for activation vary over a wide range [33,34]. During the block of an Aδ fiber, when C afferents are still active, the perception of noxious cold stimulus is experienced as burning or heat. This suggests that input from the Aδ fibers that signal the ‘cool’ sensation either blocks or modifies the C fiber input into the central nervous system. Therefore, the sensory experience induced by noxious cold stimuli depends on the integration of neuronal activity from both small myelinated and unmyelinated afferents.
In noxious cold, around 17oC or below, TRPA1 (ankyrin transmemebrane protein 1; Anktm) is activated and known to be insensitive to menthol and capsaicin but activated by icilin. TRPA1 is specifically expressed in a subset of sensory neurons that express the nociceptive markers CGRP and substance P, and may operate as peripheral mechanosensors [23,25]. TRPA1 are receptors for isothiocyanates, the pungent ingredients in wasabi and yellow mustard, cinnamon, mustard oil and inflammatory peptides and bradykinin.
The expression pattern in nociceptors suggests that TRPA1 is involved in nociceptive signaling as well as mediating the distinct pungent sensations. Moreover, TRPA1 has been postulated to play an important role in inflammatory hypersensitivity through receptor-operated mechanism by pro-algesic or pro-inflammatory mediators [28]. However, recently, whether TRPA1 is involved in cold sensation needs to be reconciled because data are conflicting regarding TRPA1 functioning as a cold sensor in vivo [29–34]. Additional studies and experimental methods are necessary to verify the function of this receptor.
CLINICAL POTENTIAL OF COOLING SENSATION
A cool and gentle breeze is known to make humans feel better and relieve discomfort and pain. The chemical sensory/cooling agents that can mimic these sensations without a change in tissue temperatures are promising as a new treatment for various disorders. Menthol, a cyclic terpene alcohol found in the leaves of the genus Mentha is representative of these agents and has been used in a variety of products, such as toothpaste, candy and even tobacco. When applied at low concentrations to the skin or mouth, menthol elicits a pleasant cool sensation. The cooling sensation of menthol arises from a molecular site of the cold receptor, TRPM8, as mentioned above [35,36]. However, menthol can evoke pain (or irritation) through activation and sensitization of C fibers with activation thresholds within the noxious range of temperatures. Therefore, in the case of menthol, both pain (or discomfort) and cold sensation appear closely linked [16,28]. Notably, low concentrations of TRPM8 agonists such as menthol or icilin were found to be anti-hyperalgesic in certain conditions [37,38]. In addition, cold patches have traditionally been used to induce analgesia or pain relief caused by traumatic injuries. Most recently, TRPM8 antagonists were shown to be effective in reversing established pain in neuropathic and visceral pain models [39].
Pruritus is a condition where the cooling sensation can actively intervene. Previously, Carstens et al. suggested that lowering the skin temperature reduces pruritus via a central inhibitory pathway [40]. Yosipovitch also suggested menthol can be used as a counterirritant for pruritus [41]. In addition to menthol, other cooling agents such as icilin, which is considered more efficacious and potent than menthol, activates the TRPM8 channel [37]. We previously showed the effect of topical application of icilin in the management of paroxysmal itching sensation over the vulvae area [42]. A TRPM8 agonist also showed efficacy in the histamine-induced itching model in vivo [43].
However, limitations may exist with regard to the duration of action because cold receptors must adapt with a prolonged change of membrane potential dependent upon calcium current [13] or because TRPM8 requires the presence of phosphatidylinositol 4,5 bisphosphate (PIP2) for stable activity [44]. Therefore, prolonged TRPM8 activation depletes calcium and PIP2 as well as a certain level of cellular adaptation. Recently, several researchers have suggested TRPM8 can be a target for treating prostate cancer because an androgen-dependent Ca2+ channel is necessary for prostate cancer to survive and grow [45].
As summarized above, cold sensation of peripheral afferent sensory is not well understood but is relevant for further exploration. The cellular and molecular mechanisms involved in the TRP family, especially TRPM8, and TRPA1 are not well recognized, thus further investigation on the interactive neuronal system of cooling and nociception can extend our knowledge for clinical use in various fields.
REFERENCES
- 1.Munger BL, Ide C. The structure and function of cutaneous sensory receptors. Arch Histol Cytol. 1988;51:1–34. doi: 10.1679/aohc.51.1. [DOI] [PubMed] [Google Scholar]
- 2.Tschachler E, Reinisch CM, Mayer C, Paiha K, Lassmann H, Weninger W. Sheet preparations expose the dermal nerve plexus of human skin and render the dermal nerve end organ accessible to extensive analysis. J Invest Dermatol. 2004;122:177–82. doi: 10.1046/j.0022-202X.2003.22102.x. [DOI] [PubMed] [Google Scholar]
- 3.Kelly EJ, Terenghi G, Hazari A, Wiberg M. Nerve fibre and sensory end organ density in the epidermis and papillary dermis of the human hand. Br J Plast Surg. 2005;58:774–9. doi: 10.1016/j.bjps.2004.12.017. [DOI] [PubMed] [Google Scholar]
- 4.Yamada N, Kashima Y, Inoue T. Scanning electron microscopy of the basal surface of the epidermis of human digits. Acta Anat (Basel) 1996;155:242–8. doi: 10.1159/000147812. [DOI] [PubMed] [Google Scholar]
- 5.Lawson SN. Phenotype and function of somatic primary afferent nociceptive neurones with C-, Adelta- or Aalpha/beta-fibres. Exp Physiol. 2002;87:239–44. doi: 10.1113/eph8702350. [DOI] [PubMed] [Google Scholar]
- 6.Alvarez FJ, Fyffe RE. Nociceptors for the 21st century. Curr Rev Pain. 2000;4:451–8. doi: 10.1007/s11916-000-0069-4. [DOI] [PubMed] [Google Scholar]
- 7.Lumpkin EA, Caterina MJ. Mechanism of sensory transduction in the skin. Nature. 2007;445(7130):858–65. doi: 10.1038/nature05662. [DOI] [PubMed] [Google Scholar]
- 8.Tsunozaki M, Bautista DM. Mammalian somatosensory mechanotransduction. Curr Opin Neurobiol. 2009;19:362–9. doi: 10.1016/j.conb.2009.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Delmas P, Hao J, Rodat-Despoix L. Molecular mechanisms of mechanotransduction in mammalian sensory neurons. Nat Rev Neurosci. 2011;12:139–53. doi: 10.1038/nrn2993. [DOI] [PubMed] [Google Scholar]
- 10.Campero M, Serra J, Ochoa JL. C-polymodal nociceptors activated by noxious low temperature in human skin. J Physiol. 1996;497:565–72. doi: 10.1113/jphysiol.1996.sp021789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hensel H. Thermoreception and temperature regulation. Monogr Physiol Soc. 1981;38:1–321. [PubMed] [Google Scholar]
- 12.Clapham DE, Runnels LW, Strübing C. The TRP ion channel family. Nat Rev Neurosci. 2001;2:387–96. doi: 10.1038/35077544. [DOI] [PubMed] [Google Scholar]
- 13.Voets T, Droogmans G, Wissenbach U, Janssens A, Flockerzi V, Nilius B. The principle of temperature-dependent gating in cold- and heat-sensitive TRP channels. Nature. 2004;430:748–54. doi: 10.1038/nature02732. [DOI] [PubMed] [Google Scholar]
- 14.Voets T, Nilius B. The pore of TRP channels: trivial or neglected? Cell Calcium. 2003;33:299–302. doi: 10.1016/s0143-4160(03)00044-7. [DOI] [PubMed] [Google Scholar]
- 15.Patapoutian A, Peier AM, Story GM, Viswanath V. Thermotrp channels and beyond: Mechanisms of temperature sensation. Nat Rev Neurosci. 2003;4:529–39. doi: 10.1038/nrn1141. [DOI] [PubMed] [Google Scholar]
- 16.Tominaga M, Caterina MJ. Thermosensation and pain. J Neurobiol. 2004;61:3–12. doi: 10.1002/neu.20079. [DOI] [PubMed] [Google Scholar]
- 17.LaMotte RH, Thalhammer JG. Response properties of high threshold cutaneous cold receptors in the primate. Brain Res. 1982;244:279–87. doi: 10.1016/0006-8993(82)90086-5. [DOI] [PubMed] [Google Scholar]
- 18.Schepers RJ, Ringkamp M. Thermoreceptors and thermosensitive afferents. Neurosci Biobehav Rev. 2010;34:177–84. doi: 10.1016/j.neubiorev.2009.10.003. [DOI] [PubMed] [Google Scholar]
- 19.Dubner R, Sumino R, Wood WI. A peripheral “cold” fiber population responsive to innocuous and noxious thermal stimuli applied to monkey's face. J Neurophysiol. 1975;38:1373–89. doi: 10.1152/jn.1975.38.6.1373. [DOI] [PubMed] [Google Scholar]
- 20.Campero M, Serra J, Bostock H, Ochoa JL. Slowly conducting afferents activated by innocuous low temperature in human skin. J Physiol. 2001;535:855–65. doi: 10.1111/j.1469-7793.2001.t01-1-00855.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kumazawa T, Perl ER. Primate cutaneous sensory units with unmyelinated (C) afferent fibers. J Neurophysiol. 1977;40:1325–38. doi: 10.1152/jn.1977.40.6.1325. [DOI] [PubMed] [Google Scholar]
- 22.Darian-Smith I, Johnson KO, Dykes R. “Cold” fiber population innervating palmar and digital skin of the monkey: responses to cooling pulses. J Neurophysiol. 1973;36:325–46. doi: 10.1152/jn.1973.36.2.325. [DOI] [PubMed] [Google Scholar]
- 23.McKemy DD. How cold is it? TRPM8 and TRPA1 in the molecular logic of cold sensation. Mol Pain. 2005;1:16. doi: 10.1186/1744-8069-1-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mackenzie RA, Burke D, Skuse NF, Lethlean AK. Fibre function and perception during cutaneous nerve block. J Neurol Neurosurg Psychiatry. 1975;38:865–73. doi: 10.1136/jnnp.38.9.865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.McKemy DD, Neuhausser WM, Julius D. Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature. 2002;416:52–8. doi: 10.1038/nature719. [DOI] [PubMed] [Google Scholar]
- 26.Colburn RW, Lubin ML, Stone DJ, Jr, Wang Y, Lawrence D, D'Andrea MR, Brandt MR, Liu Y, Flores CM, Qin N. Attenuated cold sensitivity in TRPM8 null mice. Neuron. 2007;54:379–86. doi: 10.1016/j.neuron.2007.04.017. [DOI] [PubMed] [Google Scholar]
- 27.Dhaka A, Murray AN, Mathur J, Earley TJ, Petrus MJ, Patapoutian A. TRPM8 is required for cold sensation in mice. Neuron. 2007;54:371–8. doi: 10.1016/j.neuron.2007.02.024. [DOI] [PubMed] [Google Scholar]
- 28.Chéry-Croze S. Relationship between noxious cold stimuli and the magnitude of pain sensation in man. Pain. 1983;15:265–9. doi: 10.1016/0304-3959(83)90061-1. [DOI] [PubMed] [Google Scholar]
- 29.Davis KD, Pope GE. Noxious cold evokes multiple sensations with distinct time courses. Pain. 2002;98:179–85. doi: 10.1016/s0304-3959(02)00043-x. [DOI] [PubMed] [Google Scholar]
- 30.Simone DA, Kajander KC. Responses of cutaneous A-fiber nociceptors to noxious cold. J Neurophysiol. 1997;77:2049–60. doi: 10.1152/jn.1997.77.4.2049. [DOI] [PubMed] [Google Scholar]
- 31.Davis KD. Cold-induced pain and prickle in the glabrous and hairy skin. Pain. 1998;75:47–57. doi: 10.1016/S0304-3959(97)00203-0. [DOI] [PubMed] [Google Scholar]
- 32.Bessou P, Perl ER. Response of cutaneous sensory units with unmyelinated fibers to noxious stimuli. J Neurophysiol. 1969;32:1025–43. doi: 10.1152/jn.1969.32.6.1025. [DOI] [PubMed] [Google Scholar]
- 33.Cain DM, Khasabov SG, Simone DA. Response properties of mechanoreceptors and nociceptors in mouse glabrous skin: an in vivo study. J Neurophysiol. 2001;85:1561–74. doi: 10.1152/jn.2001.85.4.1561. [DOI] [PubMed] [Google Scholar]
- 34.Campero M, Serra J, Ochoa JL. C-polymodal nociceptors activated by noxious low temperature in human skin. J Physiol. 1996;497:565–72. doi: 10.1113/jphysiol.1996.sp021789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wasner G, Schattschneider J, Binder A, Baron R. Topical menthol--a human model for cold pain by activation and sensitization of C nociceptors. Brain. 2004;127:1159–71. doi: 10.1093/brain/awh134. [DOI] [PubMed] [Google Scholar]
- 36.Peier AM, Moqrich A, Hergarden AC, Reeve AJ, Andersson DA, Story GM, Earley TJ, Dragoni I, McIntyre P, Bevan S, Patapoutian A. A TRP channel that senses cold stimuli and menthol. Cell. 2002;108:705–15. doi: 10.1016/s0092-8674(02)00652-9. [DOI] [PubMed] [Google Scholar]
- 37.Chuang HH, Neuhausser WM, Julius D. The super-cooling agent icilin reveals a mechanism of coincidence detection by a temperature-sensitive TRP channel. Neuron. 2004;43:859–69. doi: 10.1016/j.neuron.2004.08.038. [DOI] [PubMed] [Google Scholar]
- 38.Proudfoot CJ, Garry EM, Cottrell DF, Rosie R, Anderson H, Robertson DC, Fleetwood-Walker SM, Mitchell R. Analgesia mediated by the TRPM8 cold receptor in chronic neuropathic pain. Curr Biol. 2006;16:1591–605. doi: 10.1016/j.cub.2006.07.061. [DOI] [PubMed] [Google Scholar]
- 39.Lashinger ES, Steiginga MS, Hieble JP, Leon LA, Gardner SD, Nagilla R, Davenport EA, Hoffman BE, Laping NJ, Su X. AMTB, a TRPM8 channel blocker: evidence in rats for activity in overactive bladder and painful bladder syndrome. Am J Physiol Renal Physiol. 2008;295:F803–10. doi: 10.1152/ajprenal.90269.2008. [DOI] [PubMed] [Google Scholar]
- 40.Carstens E, Jinks SL. Skin cooling attenuates rat dorsal horn neuronal responses to intracutaneous histamine. Neuroreport. 1998;9:4145–9. doi: 10.1097/00001756-199812210-00027. [DOI] [PubMed] [Google Scholar]
- 41.Yosipovitch G, Szolar C, Hui XY, Maibach H. Effect of topically applied menthol on thermal, pain and itch sensations and biophysical properties of the skin. Arch Arch Dermatol Res. 1996;288:245–8. doi: 10.1007/BF02530092. [DOI] [PubMed] [Google Scholar]
- 42.Han JH, Choi HK, Kim SJ. Topical TRPM8 agonist(icilin) relieved vulva pruritus originating from lichen sclerosus et atrophicus. Acta Derm Venereol. 2012;92:561–2. doi: 10.2340/00015555-1244. [DOI] [PubMed] [Google Scholar]
- 43.Bromm B, Scharein E, Darsow U, Ring J. Effects of menthol and cold on histamine-induced itch and skin reactions in man. Neurosci Lett. 1995;187:157–60. doi: 10.1016/0304-3940(95)11362-z. [DOI] [PubMed] [Google Scholar]
- 44.Liu B, Qin F. Functional control of cold- and menthol-sensitive TRPM8 ion channels by phosphatidylinositol 4,5-bisphosphate. J Neurosci. 2005;25:1674–81. doi: 10.1523/JNEUROSCI.3632-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Zhang L, Barritt GJ. Evidence that TRPM8 is an androgen-dependent Ca2+ channel required for the survival of prostate cancer cells. Cancer Res. 2004;64:8365–73. doi: 10.1158/0008-5472.CAN-04-2146. [DOI] [PubMed] [Google Scholar]