

Abstract
Evidence suggests that eosinophils play a significant role in promoting several gastrointestinal diseases, and animal models are the significant tools to understand the pathogenesis of eosinophil-associatd inflammatory disorders. The focus of this review is on the significance of mouse models that mimic the characteristics of human eosinophilic gastrointestinal diseases. Eosinophils are the important leukocytes with diverse functions in the gastrointestinal tract, such as excretion of intestinal parasites and promoting the pathogenesis of a numerous allergic gastrointestinal disorders like food allergy, parasitic infection, allergic gastroenteritis, allergic colitis, and eosinophilic esophagitis. Among these gastrointestinal diseases, the eosinophilic esophagitis is the most recently recognized disease and the mouse models are proven to be an effective tool to understand the pathophysiology of disease and to test novel treatment strategies. Based on patients allergic conditions and the gene overexpressed in human EGID, a number of gene overexpressed and allergen-challenged mouse models of gastrointestinal disorders were developed. These models were utilized to explore the mechanism(s) that promotes the eosinophil-mediated gastrointestinal diseases including the role of the eosinophil responsive cytokines and chemokines. Herein, we have provided a detailed overviews of the mouse models of gastrointestinal disorders that mimic the human eosinophilic gastrointestinal diseases and can be utilized as a tool for understanding the diseases pathogenesis and developing novel therapeutic targets.
Keywords: Allergen, Eosinophil, Gene-deficient, Interleukin, Eotaxin, Transgenic, Mice
Introduction
Eosinophils are a small proportion of peripheral blood leukocytes, but they can selectively accumulate in peripheral blood or tissue in certain inflammatory disorders, and they play an important role in host defense as well as in the regulation of immune system[1,2]. There are numerous human inflammatory diseases that are characterized by tissue or blood eosinophilia, like asthma and food allergy eosinophilic dermatitis, eosinophilic esophagitis, gastroesophageal reflux, numerous eosinophilic gastrointestinal disorders (celiac disease, inflammatory bowl disease and allergic colitis), and parasitic infections. In some instances, eosinophils play an important role in host defense and immune system regulation[2,3] and are necessary for the eradication of parasitic infections or malignancies. In other cases, such as in asthma or allergic diseases, eosinophilic gastrointestinal disease, or hypereosinophilic syndromes, they selectively accumulate in peripheral blood or in tissues in response to the numerous stimului. Eosinophils release mediators that are pro-inflammatory and destructive to the tissues. The eosinophilia in the number of inflammatory diseases has led researchers/scientists to understand the s mechanisms involved in the induction and prgression of eosinophilic inflammation. Two molecules that appear to be critical in the regulation of selective eosinophilia are interleukin (IL)-5 and eotaxin-1, -2 and -3. IL-5 is a critical regulator of the selective growth and differentiation of eosinophil progenitor cells. IL-5 regulates the post-mitotic survival and activation of mature eosinophils, and promotes the human diseases with eosinophilia. Eotaxin-1,-2 and -3 are critical in regulating eosinophil accumulation in healthy and following allergen exposure in disease states. It has also been hypothesized that eotaxins work in concert with IL-5 to promote the tissue localization of eosinophils in diseased conditions. In addition, T lymphocytes and antigen presenting cells play a key role in orchestrating eosinophilic inflammation via the production of a number of other cytokines like, IL-5, IL-13, IL-15, IL-18, and IL-33.
Eosinophilic disorders occur when eosinophils, number increases in above the normal amounts in various parts of the body by food, parasite or due to any genetic conditions[4-7]. In healthy state the eosinophils reside in the gastrointestinal tract[8,9]; however, during allergic (food or aeroallergen) responses, a large number of eosinophils are generated from their precursors in response to the stimulus and move into the different segments of gastrointestinal segments and release a variety of toxins that can cause chronic gastrointestinal inflammation[4-7], resulting in tissue damage, repair and functional abnormalities[10,11]. The gastrointestinal disorders promoted by the eosinophils includes, eosinophilic esophagitis, characterized by elevated levels of eosinophils in the esophagus; eosinophilic gastritis, characterized by elevated eosinophils in the stomach; eosinophilic enteritis, characterized by elevated eosinophils in the small intestine and eosinophilic colitis, characterized by excessive eosinophils.
Selecting an Animal Models for Human Diseases
Several animal models have been used to understand the mechanistic pathways of a number of human diseases, including flies, zebra fish, mice, rats, cats, pigs, dogs and some non-human primates[12]. These animal species share various common genes and metabolic pathways with humans. Researchers have used these animal species as an experimental model to understand the human biology in health and disease. Experimentation with large animals is extremely expensive and might involve ethical issues; whereas, mice have become more suitable species because of their small size, cheaper maintenance costs, shorter breeding and generation time for the researchers throughout the world. Most importantly, mouse shares 99% identical genome with humans and is the only mammal, after the human, whose genome has been sequenced[13]. Therefore, mice are considered a relevant model for medical research and are widely used in understanding the mechanisms of disease pathogenesis. In addition, mice are the important species to use in medical research for many other reasons, first, they are genetically homogeneous; second, a large number of mice can be used for screening within a single experiment; third, inbred mice allow the study of identical cohorts and fourth, resources for mouse research are commercially and radially available. The use of the mouse model has led to the discovery of numerous unknown pathways in disease pathogenesis and the treatment strategy of several diseases like cancer, diabetes, gastrointestinal disorders and cardiovascular abnormalities. Mouse models have been used for the experiments for decades and much is now known concerning mouse biology and genetic. A large number of tools and techniques including gene manipulation were developed to understand the genes function in human diseases. The gene manipulation technology is not yet developed and available for other organisms. The evidence indicates that genetically engineered mice has become very common in experimental studies, mouse genes were replaced by the human orthologous or to create a mouse with a mutated version of human genes associated with specific human diseases including gastrointestinal disorders. Additionally, several mouse models are developed to study the mechanism of the induction of food or aeroallergen-induced gastrointestinal inflammatory diseases that mimic the human gastrointestinal diseases. This review mainly focuses on the significance of mouse models that provides much-needed mechanistic information of the development of human eosinophilic gastrointestinal diseases (EGID) and in the designing the therapeutic strategies.
Mouse Models to Understand the Initiation and Progression of Human EGID
Eosinophils have very diverse functions. They are involved in parasite excretion at healthy state, as well as in inflammatory responses at disease state[14-16]. Eosinophils reside in all segments of the gastrointestinal tract, except the esophagus from the prenatal to adult stage in both mice and humans at healthy state[8,9]. Eosinophil number increases in the gastrointestinal tract or in other tissues during inflammatory conditions[7,17-19]. Mouse and human eosinophil biology is very similar except in the shape of their nuclei; mouse eosinophils are multi-lobed or doughnut-shaped compared to the bi-lobed nuclei of human eosinophils. Both, human and mouse eosinophil possess all four major granular toxic proteins, termed as major basic proteins, eosinophilic cationic proteins, eosinophilic peroxidase and eosinophil-derived neurotoxins[16,19]. The induced gastrointestinal eosinophilia is observed in a number of human gastrointestinal diseases, such as celiac disease (CE), eosinophilic colitis (EC), EoE, eosinophilic gastroenteritis (EG) and gastro-esophageal reflux disease (GERD). In past years, several mouse models were developed to understand the mechanistic pathways of human gastrointestinal diseases[4,6,7,10,11,20-22]. These experimental mouse models of gastrointestinal diseases were developed either by challenging mice with food allergen, aeroallergens or by the overexpression of interested candidate genes. The specific gene overexpressed mice were created by either constitutive transgene overexpression, intestine-specific overexpression or comprising gene-inducible and gene-deficient mice to study the several human eosinophilic gastrointestinal disorders[6,10]. Interestingly, most of the developed mouse models mimic the disease characteristics observed in human. However, some disadvantages were also noticed in using the mouse models for human diseases like, the disease response differ among mouse strains. Therefore, more than one strain has to be examined. Additionally, to study any particular gastrointestinal segment abnormality, several mice have to be sacrificed. The tissue biopsy technology to evaluate disease initiation and progression for murine upper gastrointestinal tract are not yet developed. Currently, the mouse gastrointestinal biopsy technology is only available for mouse colon and is in use to evaluate the disease progression. Taken together, the use of mouse models in medical research has led to major advances in our understanding of the initiation, progression and proposing a treatment strategies for a number of eosinophilic gastrointestinal diseases.
Mouse Models that Mimic Human EOE Pathogenesis
EoE is a painful and sometimes devastating inflammatory disease that often leads to problems in swallowing, food refusal, food intolerance in infants, dysphagia and food impactions in adolescents and adults[1,23,24]. EoE is an emerging entity throughout the world, as documented by recent cases from a number of developed and developing countries[25-34]. EoE is an allergen-induced T-cell-mediated disease and is distinguished from GERD by the magnitude of mucosal eosinophilia, intraepithelial eosinophils and the lack of response to acid suppression[35,36]. Earlier, a number of food and aeroallergen-induced EoE models were reported[4,7,21,22,37]; the first EoE model was developed in 2001 by intranasally challenging the mice with Aspergillus fumigatus extract. The aeroallergen-induced EoE mouse model showed almost all the characteristic features of human EoE[7], like intraepithelial eosinophilia, extracellular eosinophilic granules, basal cell hyperplasia and eosinophilic microabscesses. The pathological similarities of mouse and human EoE pathological characteristics are shown in figure 1. Apart from the Aspergillus-induced EoE mouse model, the oral intragastric OVA beads[4] or OVA sensitized and challenged mouse models were also developed[37]. However, in both these OVA mouse models a low magnitude of esophageal eosinophilia and the absence of a number of human EoE characteristics, such as, intraepithelial eosinophils, eosinophilic microabscess, basal cell hyperplasia, and extracellular eosinophilic granules in the esophageal epithelium are not shown or reported. Further, a mouse model of a food allergen (i.e. peanut or corn) sensitized and challenged EoE mouse model was also reported that indicated a critical role of T cell subset, iNKT cells, in EoE pathogenesis[22]. Both peanut and corn allergens promoted a large number of intraepithelial esophageal eosinophilia, eosinophilic microabscess and extracellular eosinophilic granules in the esophageal epithelium that are shown here in figure 2 A-D. In addition, using the mouse models, indoor insect allergens like cockroaches and dust mites are also implicated for the EoE pathogenesis[21]. All these models dissected the significant role of eosinophil active cytokines such as IL-5, IL-13 and chemokines like eotaxin in EoE pathogenesis[21,38]. In summary, a number of EoE models are now available to study the EoE pathogenesis and to develop a treatment strategy for human EoE. Importantly, the mouse models of EoE provided the evidence of iNKT cell involvement in EoE and indicated that targeting iNKT cells may be a future novel therapy for EoE.
Figure 1.
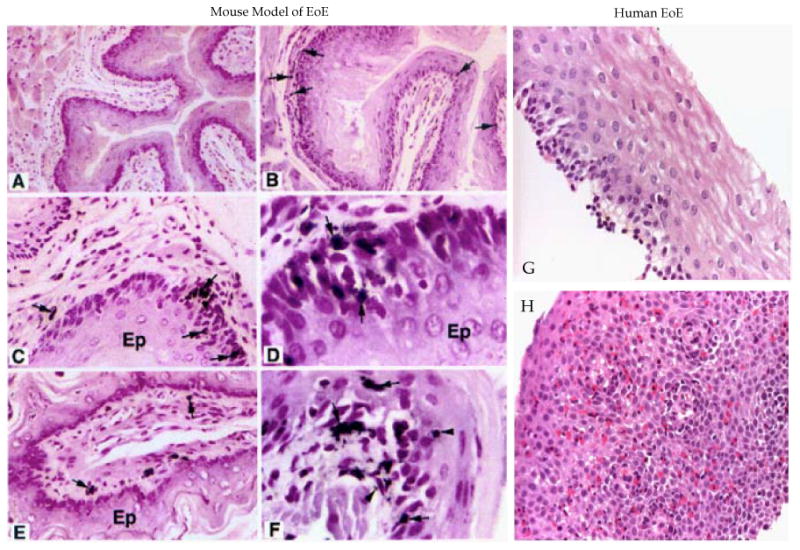
Comparision of mouse and human EoE pathological characteristics. Mouse esophageal cross-section photomicrographs of naive mouse and Aspergillus induced EoE in mice are stained with anti-MBP and is shown (A, B). The photomicrographs show esophageal microabsess and epithelial cell proliferation (C, D), laminopropria eosinophilia (E) and extracellular eosinophilic granules (F). The photomicrograph of human normal individual (G) and EoE patient (H) esophageal biopsies (G) are stained with hematoxylin and eosin is shown. No eosinophils and basal layer expansion in the epithelial mucosa in the esophageal biopsy of normal individual are detected; whereas, the EoE patients esophageal biopsies show intraepithelial eosinophils, extracellular eosinophilic granules, basal cell hyperplasia and eosinophilic microabscess. [The photomicrographs of EoE mouse model with their details are earlier published in J. Clin. Invst. 2001; 107(1):83-90 doi:10.1172/JCI10224 (Mishra et al., entitled “An ethiological role for aeroallergens and eosinophils in experimental esophagitis”].
Figure 2.
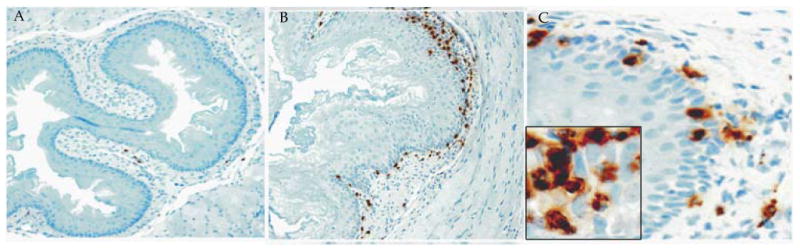
Eosinophil analysis in the esophagus of peanut sensitized and challenged mice. A, esohageal sections of saline challenged mice stained with anti-MBP show few lamina propria eosinophils (original magnification, 100×). B, esohageal sections of peanut extract challenged mice stained with anti-MBP show a large number of eosinophils in the epithelial mucosa and lamina propria (original magnification, 100×). C, the peanut extract challenged esophageal sections in high magnification (original magnification, 400×) show intraepithelial eosinophilia and degranulated eosinophils. The inset photograph shows eosinophil microabscesses with extracellular eosinophilic granules (original magnification, ×1000). [All these figures were previously published in “AJP-Gastroenterology and Liver Disease 2012; 302: G645-G654”]
Mouse Models of Esophageal Functional Abnormalities that Mimic Human Disease
The allergen-induced EoE models also show esophageal remodeling and fibrosis that are commonly observed in chronic human EoE[10,39]. Tissue remodeling or tissue fibrosis is the result of an imbalance in tissue regeneration and repair[40,41] and leukocyte infiltration is believed to contribute to the remodeling process, including eosinophils and mast cells[42,43]. Tissue remodeling involves epithelial cell hyperplasia and thickening of the epithelial mucosa, with the accumulation of extracellular matrix proteins (collagen) beneath the basement membrane[44,45]. The Masson trichrome staining of mouse esophageal section of experimental EoE (figure 3 A, B) and esophageal biopsies of human EoE (Figure 4 A) shows a low level of lamina propria collagen. In contrast, an increased lamina propria collagen accumulation together with elongation of the stromal papillae and thickened basal epithelial layer of Aspergillus-induced murine EoE was observed (Figure 3 C-F) as well as in the esophageal biopsies of EoE patients (Figure 4 B). Further, esophageal remodeling leads to develop a number of esophageal disorders like esophageal stricture (narrowing), peristaltic dysfunction, esophageal rings, tissue scars and epithelial and muscle cell hyperplasia/hypertrophy, which are commonly observed in human EoE[42-52]. Therefore, to study the functional esophageal abnormalities in EoE, a chronic mouse model of EoE was examined that showed very high eosinophil levels in each segments of the esophagus. These mice overexpressed IL-5 under control of the T-cell promoter, CD2. The eosinophils and collagen accumulation in these mice were found distributed in all the segments of esophagus from external layers of the loose connective tissue, the sub-mucosa and muscularis region[10,20]. The IL-5 chronic EoE mouse model showed a similar esophageal fibrosis as observed in human (Figure 5 A, C). Of note, the mice overexpresses IL-5 under the control of the FABPi, intestinal promoter, in which IL-5 is specifically expressed in the small intestine, showed some esophageal eosinophilia, but no induced basal layer thickness nor the accumulation of lamina propria collagen[6]. These observations highlighted the significance of local IL-5 induction in the esophagus, not systemic IL-5 for developing esophageal fibrosis in EoE. The CD2-IL-5 transgenic mice also developed a similar esophageal stricture frequently observed in human EoE (Figure 6 A, B). The development of esophageal stricture is reported to be specific for eosinophilic inflammation and is not dependent on the IL-5 transgene. A similar esophageal stricture was also reported in doxycycline (DOX) inducible IL-13 bitransgenic mouse model of EoE[11]. However, it is still not clear whether IL-13-induced esophageal eosinophilia, esophageal remodeling and the development of esophageal stricture are directly related to IL-13 or IL-13-induced IL-5. Most recent report indicates that at least allergen-induced experimental EoE is independent of IL-13[43]. Furthermore, the mouse model of chronic EoE was very useful in defining the abnormalities in esophageal abnormalities like esophageal motility. Using the mouse model, it is reported that esophageal motility is independent of the eosinophilic inflammation, and possibly might be related to the induced esophageal mast cells. The increased numbers of mast cells were reported in both human and experimental EoE[21,43,46]. The most recent report show that mast cells promote muscle cell hyperplasia and hypertrophy in allergen-induced EoE[46]. In summary, these chronic mouse models were proven to be a useful tool in defining that the development of esophageal stricture is dependent on eosinophilic inflammation and esophageal peristaltic dysfunction is independent to eosinophilic inflammation in EoE.
Figure 3.
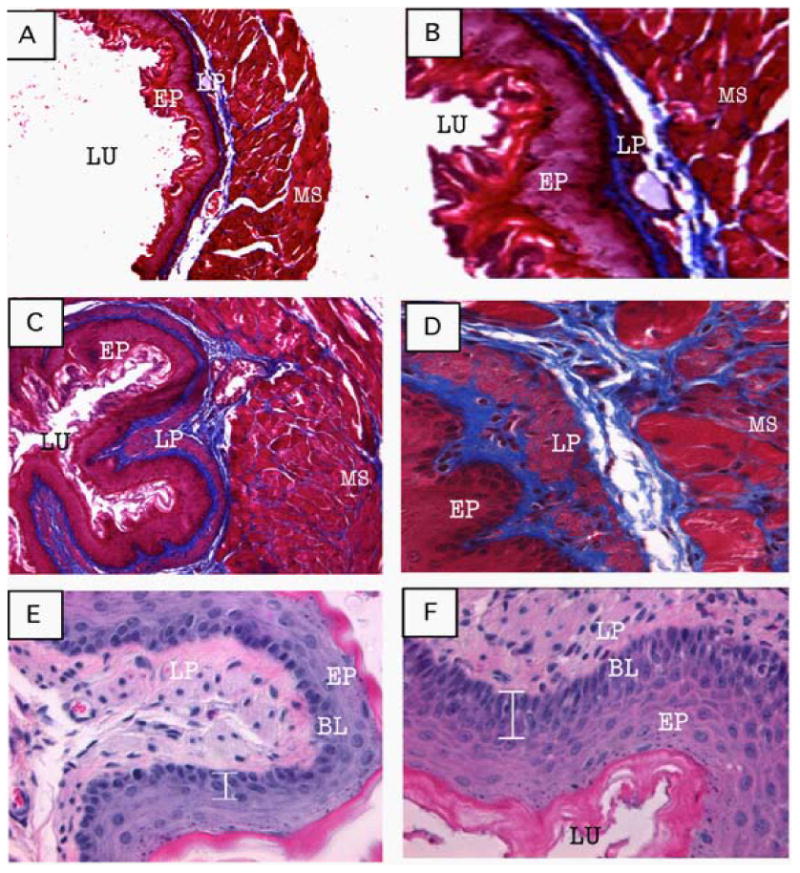
Tissue remodeling in allergen-induced EoE. Representative photomicrographs of Masson's trichrome stained tissue sections of a saline-challenged mouse shows normal lamina propria collagen (A, original magnification 100× and B, 400×) and induced collagen accumulation in lamina propria, muscularis mucosa and extended stromal papillae following allergen challenge (C, original magnification 100× and D, 400× D). The photomicrograph of the H&E stained tissue section shows single or two cell layers thickening of basal layer in saline-challenged mice (E, original magnification 400×) compared to 4-5 cell thickening in allergen-challenged mice (F, original magnification 400×). [All these figures were previously published in J Gastroenterology 2008; 134: 204-2014].
Figure 4.
Induced collagen accumulation and basal layer thickening in human EoE. A representative photomicrograph of a normal esophageal biopsy shows low levels of collagen (A, original magnification 100×). Representative photomicrograph of a EoE patient shows impressive collagen in the lamina propria and papillae (B, original magnification 100×). [Both these figures were previously published in J Gastroenterology 2008; 134: 204-2014].
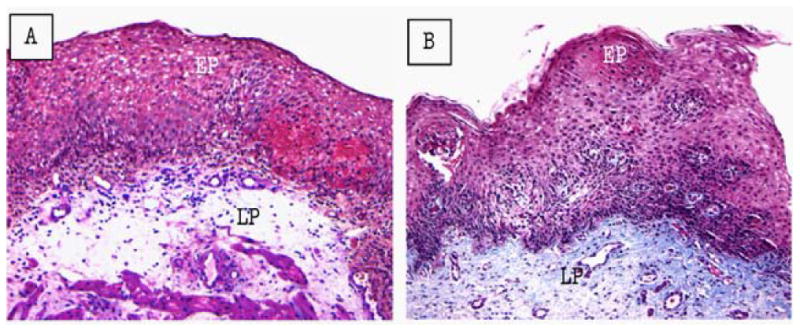
Figure 5.
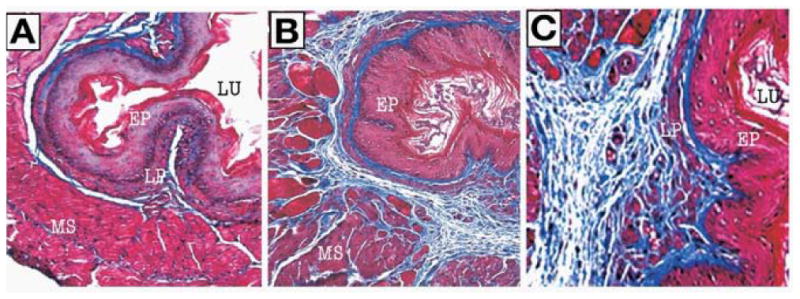
IL-5 transgene-induced esophageal tissue remodeling. Histopathological analysis was performed on the esophageal tissue sections of WT and CD2-IL-5 transgenic mice following Masson's trichrome staining and a representative photomicrograph of WT esophagus highlighting its basal layer thickness and lamina propria collagen level (A, original magnification, 100×). The CD2-IL-5 transgenic mice demonstrate thickened basal layer and collagen accumulation in the lamina propria and muscularis mucosa (B, original magnification, 100×) and induced accumulated collagen in the lamina propria and elongated papillae (C, original magnification, 400×). [All these figures were previously published in J Gastroenterology 2008; 134: 204-2014].
Figure 6.
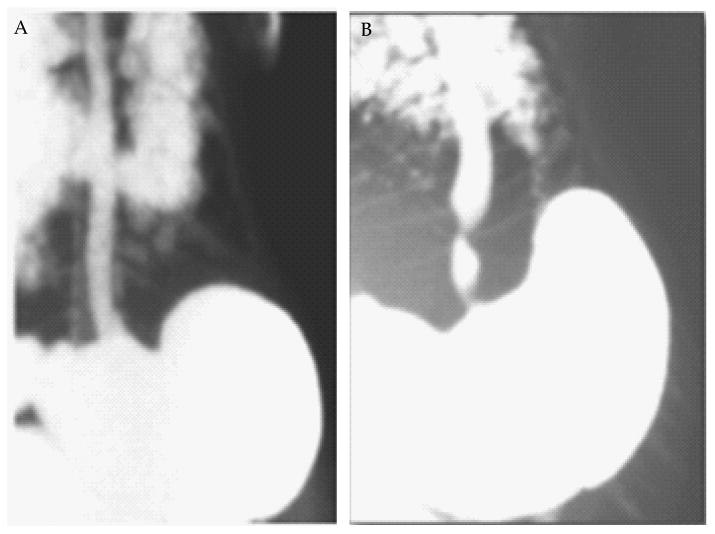
Esophageal stricture in CD2-I.L-5 transgenic mice. A radiological barium-swallowing photograph indicated esophageal stricture in CD2-IL-5 mice develops in response to IL-5-induced chronic eosinophilic inflammation in the esophagus. The barium swallowed esophageal X-ray photograph of a wild type mice (A) and CD2-IL-5 transgenic mice (B) is shown. [Both these figure were previously published in our manuscript in “AJP-Gastroenterology and Liver Disease 2012; 302: G1347-G1355”].
Mouse Model of Eosinophilic Gastroenteritis that Mimic the Human Disease
Several eosinophilic intestinal disorders including celiac disease, eosinophilic ileitis, protein-induced enteropathy and parasitic infections are associated with substantial eosinophil accumulation in the different intestinal segments[47,48]. Therefore, to dissect the signaling processes involved in induction of eosinophilic intestinal disorders, several mouse models of intestinal eosinophilia were developed by inducing the allergic responses or transgene insertion in the mouse intestine. The study of mouse models have revealed that eosinophils normally reside in the lamina propria at healthy state like humans[4,5] and in allergic conditions their number increases in both lamina propria as well as in epithelial mucosa. Further, it is well established that the eosinophil active chemokines, eotaxin, cooperate with the cytokine, interleukin (IL)-5[7,8]. IL-5 promotes eosinophil growth, development, survival, and responsiveness to chemoattractants in health and disease[7,8] and in promoting intestinal eosinophilia. Notably, elevated levels of IL-5 was earlier reported in the biopsy specimens of patients with food allergies and eosinophilic gastroenteritis[9,10] and mice overexpressing IL5 have increased levels of eotaxin-dependent eosinophilia in the gastrointestinal tract[5]. Further to dissect the mechanism of allergen-induced eosinophilic gastroenteritis, an antigeninduced eosinophilic gastrointestinal mouse model was developed by oral allergen sensitization. The allergen was given in the form of enteric-coated beads that dissolved only in acidic pH. This model resembles many characteristics of human eosinophilic gastroenteritis including a marked induction of allergen-specific IgG1, IgE, cytokine-like IL-4, IL-5 and intestinal eosinophilia[4,49]. In the model, eotaxin-1 gene deficiency showed complete protection; whereas, IL-5 deficiency showed a partial protection of the eosinophil accumulation in the intestine[4,49]. Taken together, the orally sensitized mouse model provided the mechanism that accumulation of intestinal eosinophilia is independent to IL-5 but dependent on eotaxin-1[4,49].
Human Intestinal Enterocytes Overexpresses Eotaxin-1
Intestinal enterocytes have been shown to be a chief source of selected cytokines and chemokines (e.g. IL-8, MCP-1, ENA-78) in inflammatory lesions in patients with diverse gastrointestinal inflammatory disorders[31,39,40]. Although, intestinal enterocytes express diverse chemokines, the specific involvement of enterocyte-derived chemokines in the gastro-intestinal immune system and in the pathogenesis of disease is not largely understood. The critical role of eotaxin-1 was shown in the mouse model of eosinophilic gastroenteritis; however, it was not clear that this chemokine had a similar role in human gastroenteritis. Therefore, eotaxin-1 expression was examined in the duodenal biopsies of gastroenteritis patients along with normal individuals. The analysis indicated that intestinal enterocytes had the capacity to produce eotaxin-1 in disease state, as no significant staining of anti-eotaxin-1 in duodenal biopsy was detected in a normal individual (Figure 7A); however, anti-eotaxin-1 stained duodenal biopsy of gastroenteritis revealed that eotaxin-1 protein expressed in high levels in the intestinal epithelial cells and some submucosal cells (Figure 7B). This study indicated that small intestinal enterocytes are the source of eotaxin-1 in human at disease states and provided the evidence that mouse models of eosinophilic gastroenteritis mimic the human disease. Furthermore, to establish the role of eotaxin-1 and IL-5 in gastrointestinal eosinophilic inflammation, a transgenic mouse was developed by utilizing the intestine-specific promoter of the rat fatty acid binding protein (FABPi) gene[28,29]. The FABPi promoter was shown that it specifically expresses in enterocytes of the small intestine[28,29] and eotaxin-1 and IL-5 overexpression in enterocytes promoted eosinophil trafficking in small intestine (Figure 8 A-C). Furthermore, to establish the distinct role of eotaxin-1, a genetic rescue mouse, was developed and examined. These mice resolve the eosinophil deficiency observed in eotaxin-deficient mice and provided the opportunity to establish the role of eotaxin-1 in regulating gastrointestinal eosinophilia[6]. The mice overexpressing IL-5 and eotaxin-1 in the intestinal enterocytes mimic the characteristics observed in the human eosinophilic gastrointestinal disease. In short, these described mouse models are proven to be a potential tool to understand the role of eotaxin-1 in regulating eosinophils in the human gastrointestinal tract at disease states.
Figure 7.
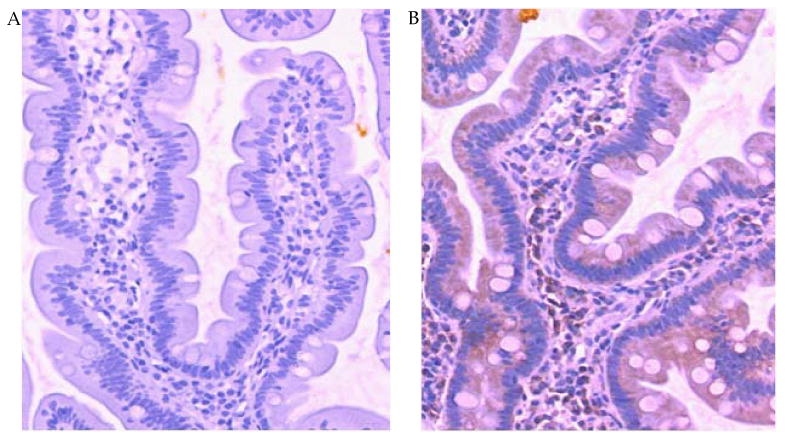
Eotaxin expression by the human enterocytes. Eotaxin protein expression was determined by immunohistochemical staining of duodenal biopsy samples from patients with eosinophilic gastroenteritis. Representative immunostaining with control IgG (A) and anti-eotaxin (B) is shown. The eotaxin staining is represented by brown color.
Figure 8.
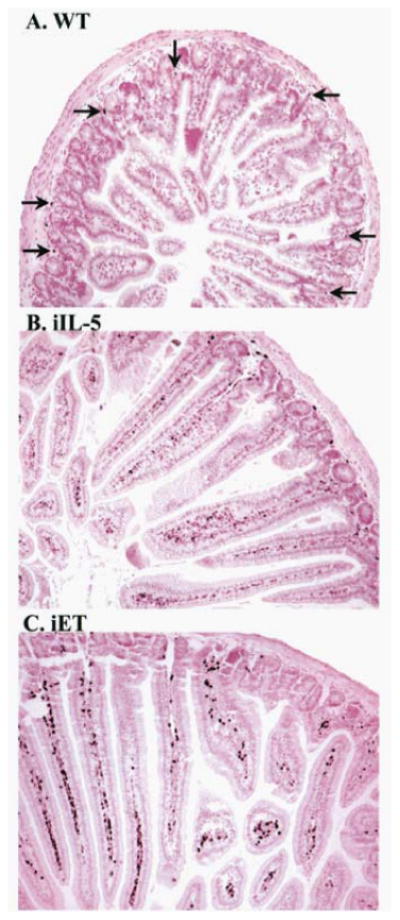
Immunohistochemical staining of jejunum sections from wild-type and transgenic mice. The jejunum from wild-type (WT; A), FABPii-IL-5 transgenic (iIL-5; B), or FABPi-eotaxin transgenic (iET; C) mice was stained with anti-MBP. Representative tissue sections are shown, and representative eosinophils, recognized by anti-MBP stained black color cells.staining, are indicated by the arrows. Original magnification, ×125. (These figures were original published in J. Biol. Chem 277; (6), 4406-4412, 2002 @ the American Society for Biochemistry and Molecular Biology).
Mouse Model of Eosinophilic Colitis that Mimic the Human Disease
Researchers developed a chronic eosinophilic colitis mouse model by either providing dextran sodium sulfate (DSS) via drinking water or by intra-rectal delivery of tri-nitrobenzene sulfonate (TNBS) intra-rectally. Both these mouse models show a large number of eosinophils in the mouse colon that mimicked the human disease, including weight loss, diarrhea, and rectal bleeding[5,50,51]. Diarrhea in these models was identified by mucus–fecal material adhering to anal fur or was confirmed by examination of the colon following the termination of the experiment. The appearance of rectal bleeding was diagnosed as diarrhea containing visible blood, mucus, or both. The intestinal pathology was examined in these models by haematoxylin and eosin and Masson trichrome staining and found to be similar to pathological characteristics as observed in human[5,50,51]. The most common characteristics observed in these models that mimic the human disease were edema, erosion/ulceration, crypt loss and infiltration of mononuclear and polymorphonuclear cells (neutrophils and eosinophils) in the lamina propria and submucosa[5,50,51]. Furthermore, murine colonoscopy techniques similar to the human were also reported. Anesthetizing the mouse like human and by using a small-animal rigid telescope connected to a camera, the colon biopsies were obtained[5,50,51]. The colonoscopy technique in mouse was found very useful for studying the mechanism of disease and treatment progression. Taken together, the mouse model of eosinophilic colitis and newly developed colonoscopy techniques provided a tool to better understand the mechanism of the progression of eosinophilic colitis in humans.
Mouse Models: A Useful Tool for Preclinical Therapeutic Drug Trials for Gastrointestinal Disorders
The choice of an appropriate animal model for a pre-clinical trial depends on the disease and drug to be tested. The selection of correct animal model is important along with good laboratory practice, basic research ethics, veterinarians, documentarians, a regulatory research coordinator, physician and a basic research scientist. It is always debatable whether animal models, in particular, the mouse are suitable models for conducting clinical trials. Notably, mouse models are widely used for conducting preclinical trials in academic and medical environment, as well as by pharmaceutical companies. Mice have occasionally been shown not to be reliable as pre-clinical models for all human diseases; drugs are effective in the mouse but are ineffective in human clinical trials. Alternatively, for some diseases the mouse modes has been proven to be very useful[52,53], for example, knocking out the leptin hormone gene in mice showed that it regulates appetite and prevents obesity[54,55]. Leptin is currently used to treat people suffering from certain types of obesity[56,57]. Similarly, anti-IL-5 therapy tested in mice was poor in controlling eosinophilic pulmonary disorder but showed a significant improvement in initial clinical trials in human EoE[58]. In addition, the mouse models were also successfully used to develop a treatment strategy for human Crohn's disease and irritable bowl diseases[59]. Furthermore, the mouse models are critical tool for preclinical trials, as they will be easily manipulated for optimization of the dosage regimes for any new treatments or treatment combinations. Taken together, the mouse models are proven effective for a number of diseases including EGID and are needed for testing any new drug for the treatment of human gastrointestinal disorders/diseases or any other disease.
Conclusions
The overwhelming conclusion is that mouse models contribute and provide in depth information to our understanding on the disease pathogenesis of EGID and on the development of new treatment strategies including the testing of new drugs. Although, mouse models raise some concerns; but still most academicians, medical researchers and drug companies use mouse models to understand the induction and progression of complex disease mechanisms and design the treatment strategies of a number of human gastrointestinal disorders like EoE, IBD and EC. A large-scale phenotyping of genetically engineered mice provides valuable information in dissecting the mechanism of a number of diseases. Importantly, without the use of mouse models, the development and performance of preclinical studies of new drugs to treat human diseases might not be possible. Mouse models have been successfully used to validate drug targets and to determine efficacious and safe usage schemes for combination treatment in humans. In conclusion, the tremendous evidence indicates that the mouse models are vastly used in dissecting and understanding the human diseases; even though the questions continue to be raised on their significance for mimicking the human diseases.
Acknowledgments
This work was supported in part by the grants NIH R01 DK067255 (AM), NIH R01 AI080581 (AM). The help and guidance of Dr. Marc Rothenberg, MD, PhD, (Director, Division of Allergy and Immunology, Cincinnati Children's Hospital Medical Center, Cincinnati, Ohio) is greatly appreciated for successfully completing a number of studies reported in this review. We further thank Drs. L . Lee Hamm and Joseph A. Lasky for providing the facility to continue our eosinophil associated disorders research work at Tulane University School of Medicine.
Footnotes
Peer reviewer: ANDREW STEWART DAY, Department of Paediatrics, University of Otago, Christchurch, Christchurch, 8140, NEW ZEALAND.
References
- 1.Markowitz JE, Liacouras CA. Eosinophilic esophagitis. Gastroenterol Clin North Am. 2003;32:949–966. doi: 10.1016/s0889-8553(03)00047-5. [DOI] [PubMed] [Google Scholar]
- 2.Bjerke K, Halstensen TS, Jahnsen F, Pulford K, Brandtzaeg P. Distribution of macrophages and granulocytes expressing L1 protein (calprotectin) in human Peyer's patches compared with normal ileal lamina propria and mesenteric lymph nodes. Gut. 1993;34:1357–1363. doi: 10.1136/gut.34.10.1357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Rothenberg ME. Eosinophilic gastrointestinal disorders (EGID) J Allergy Clin Immunol. 2004;113:11–28. doi: 10.1016/j.jaci.2003.10.047. quiz 29. [DOI] [PubMed] [Google Scholar]
- 4.Hogan SP, Mishra A, Brandt EB, Royalty MP, Pope SM, Zimmermann N, Foster PS, Rothenberg ME. A pathological function for eotaxin and eosinophils in eosinophilic gastrointestinal inflammation. Nat Immunol. 2001;2:353–360. doi: 10.1038/86365. [DOI] [PubMed] [Google Scholar]
- 5.Hogan SP, Seidu L, Blanchard C, Groschwitz K, Mishra A, Karow ML, Ahrens R, Artis D, Murphy AJ, Valenzuela DM, Yancopoulos GD, Rothenberg ME. Resistin-like molecule beta regulates innate colonic function: barrier integrity and inflammation susceptibility. J Allergy Clin Immunol. 2006;118:257–268. doi: 10.1016/j.jaci.2006.04.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Mishra A, Hogan SP, Brandt EB, Wagner N, Crossman MW, Foster PS, Rothenberg ME. Enterocyte expression of the eotaxin and interleukin-5 transgenes induces compartmentalized dysregulation of eosinophil trafficking. J Biol Chem. 2002;277:4406–4412. doi: 10.1074/jbc.M110424200. [DOI] [PubMed] [Google Scholar]
- 7.Mishra A, Hogan SP, Brandt EB, Rothenberg ME. An etiological role for aeroallergens and eosinophils in experimental esophagitis. J Clin Invest. 2001;107:83–90. doi: 10.1172/JCI10224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Mishra A, Hogan SP, Lee JJ, Foster PS, Rothenberg ME. Fundamental signals that regulate eosinophil homing to the gastrointestinal tract. J Clin Invest. 1999;103:1719–1727. doi: 10.1172/JCI6560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.DeBrosse CW, Case JW, Putnam PE, Collins MH, Rothenberg ME. Quantity and distribution of eosinophils in the gastrointestinal tract of children. Pediatr Dev Pathol. 2006;9:210–218. doi: 10.2350/11-05-0130.1. [DOI] [PubMed] [Google Scholar]
- 10.Mishra A, Wang M, Pemmaraju VR, Collins MH, Fulkerson PC, Abonia JP, Blanchard C, Putnam PE, Rothenberg ME. Esophageal Remodeling Develops as a Consequence of Tissue Specific IL-5-Induced Eosinophilia. Gastroenterology. 2008;134:204–214. doi: 10.1053/j.gastro.2007.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mavi P, Rajavelu P, Rayapudi M, Paul RJ, Mishra A. Esophageal functional impairments in experimental eosinophilic esophagitis. Am J Physiol Gastrointest Liver Physiol. 2012;302:G1347–1355. doi: 10.1152/ajpgi.00013.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Rust JH. Animal models for human diseases. Perspect Biol Med. 1982;25:662–672. doi: 10.1353/pbm.1982.0067. [DOI] [PubMed] [Google Scholar]
- 13.Bradley A. Mining the mouse genome. Nature. 2002;420:512–514. doi: 10.1038/420512a. [DOI] [PubMed] [Google Scholar]
- 14.Abu-Ghazaleh RI, Kita H, Gleich GJ. Eosinophil activation and function in health and disease. Immunology Series. 1992;57:137–67. [PubMed] [Google Scholar]
- 15.Capron M. Eosinophils and parasites. Annales de Parasitologie Humaine et Comparee. 1991;66:41–45. [PubMed] [Google Scholar]
- 16.Rothenberg ME, Hogan SP. The eosinophil. Annu Rev Immunol. 2006;24:147–174. doi: 10.1146/annurev.immunol.24.021605.090720. [DOI] [PubMed] [Google Scholar]
- 17.Boyce JA. The pathobiology of eosinophilic inflammation. Allergy Asthma Proc. 1997;18:293–300. doi: 10.2500/108854197778590489. [DOI] [PubMed] [Google Scholar]
- 18.Desreumaux P, Capron M. Eosinophils in allergic reactions. Curr Opin Immunol. 1996;8:790–795. doi: 10.1016/s0952-7915(96)80006-9. 1996. [DOI] [PubMed] [Google Scholar]
- 19.Gleich GJ, Ottesen EA, Leiferman KM, Ackerman SJ. Eosinophils and human disease. International Archives of Allergy & Applied Immunology. 1989;88:1–2. doi: 10.1159/000234749. [DOI] [PubMed] [Google Scholar]
- 20.Mishra A, Hogan SP, Brandt EB, Rothenberg ME. IL-5 promotes eosinophil trafficking to the esophagus. J Immunol. 2002;168:2464–2469. doi: 10.4049/jimmunol.168.5.2464. [DOI] [PubMed] [Google Scholar]
- 21.Rayapudi M, Mavi P, Zhu X, Pandey AK, Abonia JP, Rothenberg ME, Mishra A. Indoor insect allergens are potent inducers of experimental eosinophilic esophagitis in mice. J Leukoc Biol. 2010;88:337–346. doi: 10.1189/jlb.0110025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Rajavelu RM, Zhu X, Narayanan S, Mattner J, Collins MH, Kaul A, Putnam PE, Rothenberg ME, Mishra A. Induction and activation of invariant natural killer T cells is critical in the pathogenesis of eosinophilic esophagitis Mucosa Immunology. Am J Physiol Gastrointest Liver Physiol. 2012;302(7):645–654. [Google Scholar]
- 23.Liacouras CA, Spergel JM, Ruchelli E, Verma R, Mascarenhas M, Semeao E, Flick J, Kelly J, Brown-Whitehorn T, Mamula P, Markowitz JE. Eosinophilic esophagitis: a 10-year experience in 381 children. Clin Gastroenterol Hepatol. 2005;3:1198–1206. doi: 10.1016/s1542-3565(05)00885-2. [DOI] [PubMed] [Google Scholar]
- 24.Liacouras CA, Furuta GT, Hirano I, Atkins D, Attwood SE, Bonis PA, Burks AW, Chehade M, Collins MH, Dellon ES, Dohil R, Falk GW, Gonsalves N, Gupta SK, Katzka DA, Lucendo AJ, Markowitz JE, Noel RJ, Odze RD, Putnam PE, Richter JE, Romero Y, Ruchelli E, Sampson HA, Schoepfer A, Shaheen NJ, Sicherer SH, Spechler S, Spergel JM, Straumann A, Wershil BK, Rothenberg ME, Aceves SS. Eosinophilic esophagitis: Updated consensus recommendations for children and adults. J Allergy Clin Immunol. 2011;128(1):3–20. doi: 10.1016/j.jaci.2011.02.040. 2011. [DOI] [PubMed] [Google Scholar]
- 25.Croese J, Fairley SK, Masson JW, Chong AK, Whitaker DA, Kanowski PA, Walker NI. Clinical and endoscopic features of eosinophilic esophagitis in adults. Gastrointest Endosc. 2003;58:516–522. doi: 10.1067/s0016-5107(03)01870-4. [DOI] [PubMed] [Google Scholar]
- 26.Cury EK, Schraibman V, Faintuch S. Eosinophilic infiltration of the esophagus: gastroesophageal reflux versus eosinophilic esophagitis in children--discussion on daily practice. J Pediatr Surg. 2004;39:e4–7. doi: 10.1016/j.jpedsurg.2003.10.028. [DOI] [PubMed] [Google Scholar]
- 27.Attwood SE, Smyrk TC, Demeester TR, Jones JB. Esophageal eosinophilia with dysphagia. A distinct clinicopathologic syndrome. Dig Dis Sci. 1993;38:109–116. doi: 10.1007/BF01296781. [DOI] [PubMed] [Google Scholar]
- 28.Cantu P, Velio P, Prada A, Penagini R. Ringed oesophagus and idiopathic eosinophilic oesophagitis in adults: an association in two cases. Dig Liver Dis. 2005;37:129–134. doi: 10.1016/j.dld.2004.04.008. [DOI] [PubMed] [Google Scholar]
- 29.Fujiwara H, Morita A, Kobayashi H, Hamano K, Fujiwara Y, Hirai K, Yano M, Naka T, Saeki Y. Infiltrating eosinophils and eotaxin: their association with idiopathic eosinophilic esophagitis. Ann Allergy Asthma Immunol. 2002;89:429–432. doi: 10.1016/S1081-1206(10)62047-9. [DOI] [PubMed] [Google Scholar]
- 30.Munitiz V, Martinez de Haro LF, Ortiz A, Pons JA, Bermejo J, Serrano A, Molina J, Parrilla P. Primary eosinophilic esophagitis. Dis Esophagus. 2003;16:165–168. doi: 10.1046/j.1442-2050.2003.00319.x. [DOI] [PubMed] [Google Scholar]
- 31.Lucendo AJ, Carrion G, Navarro M, Pascual JM, Gonzalez P, Castillo P, Erdozain JC. Eosinophilic esophagitis in adults: an emerging disease. Dig Dis Sci. 2004;49:1884–1888. doi: 10.1007/s10620-004-9588-x. [DOI] [PubMed] [Google Scholar]
- 32.Straumann A, Spichtin HP, Bucher KA, Heer P, Simon HU. Eosinophilic esophagitis: red on microscopy, white on endoscopy. Digestion. 2004;70:109–116. doi: 10.1159/000080934. [DOI] [PubMed] [Google Scholar]
- 33.Straumann A. What is your diagnosis? Primary eosinophilic esophagitis. Schweiz Rundsch Med Prax. 2004;93:795–796. doi: 10.1024/0369-8394.93.19.795. [DOI] [PubMed] [Google Scholar]
- 34.Orenstein SR, Shalaby TM, Di Lorenzo C, Putnam PE, Sigurdsson L, Kocoshis SA. The spectrum of pediatric eosinophilic esophagitis beyond infancy: a clinical series of 30 children. Am J Gastroenterol. 2000;95:1422–1430. doi: 10.1111/j.1572-0241.2000.02073.x. [DOI] [PubMed] [Google Scholar]
- 35.Furuta GT, Liacouras CA, Collins MH, Gupta SK, Justinich C, Putnam PE, Bonis P, Hassall E, Straumann A, Rothenberg ME. Eosinophilic esophagitis in children and adults: a systematic review and consensus recommendations for diagnosis and treatment. Gastroenterology. 2007;133:1342–1363. doi: 10.1053/j.gastro.2007.08.017. [DOI] [PubMed] [Google Scholar]
- 36.Rothenberg ME, Mishra A, Collins MH, Putnam PE. Pathogenesis and clinical features of eosinophilic esophagitis. J Allergy Clin Immunol. 2001;108:891–894. doi: 10.1067/mai.2001.120095. [DOI] [PubMed] [Google Scholar]
- 37.Rubinstein E, Cho JY, Rosenthal P, Chao J, Miller M, Pham A, Aceves SS, Varki A, Broide DH. Siglec-F inhibition reduces esophageal eosinophilia and angiogenesis in a mouse model of eosinophilic esophagitis. J Pediatr Gastroenterol Nutr. 2011;53:409–416. doi: 10.1097/MPG.0b013e3182182ff8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Blanchard C, Stucke EM, Rodriguez-Jimenez B, Burwinkel K, Collins MH, Ahrens A, Alexander ES, Butz BK, Jameson SC, Kaul A, Franciosi JP, Kushner JP, Putnam PE, Abonia JP, Rothenberg ME. A striking local esophageal cytokine expression profile in eosinophilic esophagitis. J Allergy Clin Immunol. 2011;127:208–217. 217 e201–207. doi: 10.1016/j.jaci.2010.10.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Aceves SS, Newbury RO, Dohil R, Bastian JF, Broide DH. Esophageal remodeling in pediatric eosinophilic esophagitis. J Allergy Clin Immunol. 119:206–212. doi: 10.1016/j.jaci.2006.10.016. [DOI] [PubMed] [Google Scholar]
- 40.Bousquet J, Jeffery PK, Busse WW, Johnson M, Vignola AM. Asthma. From bronchoconstriction to airways inflammation and remodeling. Am J Respir Crit Care Med. 2000;161:1720–1745. doi: 10.1164/ajrccm.161.5.9903102. [DOI] [PubMed] [Google Scholar]
- 41.Jeffery PK, Wardlaw AJ, Nelson FC, Collins JV, Kay AB. Bronchial biopsies in asthma. An ultrastructural, quantitative study and correlation with hyperreactivity. Am Rev Respir Dis. 1989;140:1745–1753. doi: 10.1164/ajrccm/140.6.1745. [DOI] [PubMed] [Google Scholar]
- 42.Gross TJ, Hunninghake GW. Idiopathic pulmonary fibrosis. N Engl J Med. 2001;345:517–525. doi: 10.1056/NEJMra003200. [DOI] [PubMed] [Google Scholar]
- 43.Niranjan R, R M, Mishra A, Dutt P, Mishra A. Pathogenesis of allergen-induced eosinophilic esophagitis is independent of Interleukin (IL)-13. Immonology and Cell Biology. 2013 doi: 10.1038/icb.2013.21. In Press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Jeffery PK. Remodeling in asthma and chronic obstructive lung disease. Am J Respir Crit Care Med. 2001;164:S28–38. doi: 10.1164/ajrccm.164.supplement_2.2106061. [DOI] [PubMed] [Google Scholar]
- 45.Hoshino M, Nakamura Y, Sim JJ. Expression of growth factors and remodelling of the airway wall in bronchial asthma. Thorax. 1998;53:21–27. doi: 10.1136/thx.53.1.21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Niranjan R, M P, Rayapudi M, Dynda S, Mishra A. Pathogenic role of mast cells in experimental eosinophilic esophagitis. Am J Physiol Gastroenterest LiverPhysiol. 2013;304 doi: 10.1152/ajpgi.00070.2013. In Press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Rothenberg ME, Mishra A, Brandt EB, Hogan SP. Gastrointestinal eosinophils. Immunol Rev. 2001;179:139–155. doi: 10.1034/j.1600-065x.2001.790114.x. [DOI] [PubMed] [Google Scholar]
- 48.Rothenberg ME, Mishra A, Brandt EB, Hogan SP. Gastrointestinal eosinophils in health and disease. Adv Immunol. 2001;78:291–328. doi: 10.1016/s0065-2776(01)78007-8. [DOI] [PubMed] [Google Scholar]
- 49.Hogan SP, Mishra A, Brandt EB, Foster PS, Rothenber ME. A critical role for eotaxin in experimental oral antigen-induced eosinophilic gastrointestinal allergy. Proc Natl Acad Sci U S A. 2000;97:6681–6686. doi: 10.1073/pnas.97.12.6681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Stein J, Ries J, Barrett KE. Disruption of intestinal barrier function associated with experimental colitis: possible role of mast cells. Am J Physiol. 1998;274:G203–209. doi: 10.1152/ajpgi.1998.274.1.G203. [DOI] [PubMed] [Google Scholar]
- 51.Muther H, Kuhlcke K, Gessner A, Abdallah S, Lother H. Homodimeric murine interleukin-3 agonists indicate that ligand dimerization is important for high-affinity receptor complex formation. Growth Factors. 1994;10:17–27. doi: 10.3109/08977199409019600. [DOI] [PubMed] [Google Scholar]
- 52.Sun XY, Hayashi Y, Xu S, Kanou Y, Takagishi Y, Tang YP, Murata Y. Inactivation of the Rcan2 gene in mice ameliorates the age- and diet-induced obesity by causing a reduction in food intake. PLoS One. 2011;6:e14605. doi: 10.1371/journal.pone.0014605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.de Meijer VE, Le HD, Meisel JA, Puder M. Repetitive orogastric gavage affects the phenotype of diet-induced obese mice. Physiol Behav. 2010;100:387–393. doi: 10.1016/j.physbeh.2010.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.White CL, Ishihara Y, Dotson TL, Hughes DA, Bray GA, York DA. Effect of a beta-3 agonist on food intake in two strains of rats that differ in susceptibility to obesity. Physiol Behav. 2004;82:489–496. doi: 10.1016/j.physbeh.2004.04.059. [DOI] [PubMed] [Google Scholar]
- 55.Couturier K, Servais S, Koubi H, Sempore B, Cottet-Emard JM, Guigas B, Lavoie JM, Favier R. Metabolic and hormonal responses to exercise in the anti-obese Lou/C rats. Int J Obes Relat Metab Disord. 2004;28:972–978. doi: 10.1038/sj.ijo.0802717. [DOI] [PubMed] [Google Scholar]
- 56.Hall RS, Thomas SJ, Johnson SK. Australian sweet lupin flour addition reduces the glycaemic index of a white bread breakfast without affecting palatability in healthy human volunteers. Asia Pac J Clin Nutr. 2005;14:91–97. [PubMed] [Google Scholar]
- 57.Chu SC, Chou YC, Liu JY, Chen CH, Shyu JC, Chou FP. Fluctuation of serum leptin level in rats after ovariectomy and the influence of estrogen supplement. Life Sci. 1999;64:2299–2306. doi: 10.1016/s0024-3205(99)00181-2. [DOI] [PubMed] [Google Scholar]
- 58.Stein ML, Collins MH, Villanueva JM, Kushner JP, Putnam PE, Buckmeier BK, Filipovich AH, Assa'ad AH, Rothenberg ME. Anti-IL-5 (mepolizumab) therapy for eosinophilic esophagitis. J Allergy Clin Immunol. 2006;118:1312–1319. doi: 10.1016/j.jaci.2006.09.007. [DOI] [PubMed] [Google Scholar]
- 59.Dooley TP, Curto EV, Reddy SP, Davis RL, Lambert GW, Wilborn TW, Elson CO. Regulation of gene expression in inflammatory bowel disease and correlation with IBD drugs: screening by DNA microarrays. Inflamm Bowel Dis. 2004;10:1–14. doi: 10.1097/00054725-200401000-00001. [DOI] [PubMed] [Google Scholar]