

Abstract
The lack of specificity of traditional cytotoxic drugs has triggered the development of anticancer agents that selectively address specific molecular targets. An intrinsic property of these specialized drugs is their limited applicability for specific patient subgroups. Consequently, the generation of information about tumor characteristics is the key to exploit the potential of these drugs. Currently, cancer stratification relies on three approaches: Gene expression analysis and cancer proteomics, immunohistochemistry and molecular imaging. In order to enable the precise localization of functionally expressed targets, molecular imaging combines highly selective biomarkers and intense signal sources. Thus, cancer stratification and localization are performed simultaneously. Many cancer types are characterized by altered receptor expression, such as somatostatin receptors, folate receptors or Her2 (human epidermal growth factor receptor 2). Similar correlations are also known for a multitude of transporters, such as glucose transporters, amino acid transporters or hNIS (human sodium iodide symporter), as well as cell specific proteins, such as the prostate specific membrane antigen, integrins, and CD20. This review provides a comprehensive description of the methods, targets and agents used in molecular imaging, to outline their application for cancer stratification. Emphasis is placed on radiotracers which are used to identify altered expression patterns of cancer associated markers.
Keywords: cancer stratification, molecular imaging, radiotracers, positron emission tomography (PET), single photon emission computed tomography (SPECT), biomarkers, target structure
1. Introduction
Stratified medicine deploys highly specific drugs for treatment purposes. By virtue of their specificity, these drugs are only applicable in subgroups of patient collectives, which show expression of the required target structure or target isoform. In order to provide patients with the most effective treatment as quickly as possible, clustering of patient subgroups according to target expression patterns is of utmost importance (see Figure 1). The most important methods in the area of patient stratification are cancer proteomics, immunohistochemistry and molecular imaging [1]. Cancer proteomics mainly relies on monitoring serum biomarkers for diagnostic, prognostic or stratification purposes [2] and plays an important role in the discovery of signaling pathways [3] and gene expression profiling [4], which altogether can be used for cancer stratification. The emphasis of cancer proteomics is placed on functional analysis of deregulated cellular pathways and their role in the pathogenesis of a disease [5].
Figure 1.

Cancer stratification by molecular imaging allows patients clustering according to the expected outcome of a therapeutic approach by visualizing an expressed biomarker. The resulting patient subgroups can subsequently be treated with the most promising therapy [6].
Molecular imaging describes in vivo imaging methods that use selective biomarkers in order to gain functional and anatomical information about the patient. For this purpose, many different imaging techniques are currently in use, the most prominent being positron emission tomography (PET) and single photon emission computed tomography (SPECT) [7]. Other important medical imaging methods, such as computed tomography (CT) and magnetic resonance imaging (MRI) require high amounts of contrast agents in order to derive functional information. This limits their applicability for molecular imaging; so far CT and MRI have not been implemented in the clinical molecular imaging routine [8,9].
The basic strategy of molecular imaging is the combination of highly selective biomarkers and efficient signal sources. Especially the use of radioactive substances by PET and SPECT is of particular interest in the field of oncology. This is mainly due to the low detection limit of radioactive decays, which allows functional imaging at high resolution with minimal tracer quantities, compared to CT or MRI [6,7]. Molecular imaging shows several advantages over other stratification methods: It is non-invasive and allows simultaneous, real-time and in vivo cancer detection and localization by visually proving the presence of an expressed biomarker both in the primary tumor and in metastases throughout the body. As molecular imaging relies on highly affine tracer molecules, this also provides an approach for targeted cancer therapy: Substitution of the diagnostic signal source by a cytotoxic moiety results in a therapeutic compound [8].
2. Targets for Cancer Stratification by Molecular Imaging
2.1. Glucose Utilization
The cellular uptake of glucose is mediated by two distinct types of transporters: sodium-dependent glucose transporters (SGLT) and glucose transporters (GLUT). So far, 12 members of the SGLT family are known. They belong to the solute carrier 5 family (SLC5) and function as sodium/glucose symporters [10]. GLUTs, on the other hand, belong to the solute carrier 2 family (SLC2) and allow facilitated diffusion of glucose along its concentration gradient. At present, 14 members of the GLUT family are known [11]. They have been clustered in three different groups depending on sequence homologies [12].
In the absence of sufficient amounts of oxygen, the cellular glucose metabolism changes and glucose is no longer fully oxidized. Under anaerobic conditions, cells rely more or less exclusively on glycolysis for the generation of energy and on the production of lactate, in order to regenerate the amount of NAD+ consumed during glycolysis [13,14]. In order to generate the energy required, cells increase their glucose uptake. There are many organs which are known for their high glucose uptake, such as heart, liver and brain. In addition, inflammations also show high glucose uptake [15,16].
The metabolism of cancer cells differs highly from that of normal cells of the same tissue: Many cancer types show increased glucose uptake and utilization, even under non-hypoxic conditions [11,17]. Depending on the type of cancer observed, glucose uptake can be increased by about 20- to 30-fold when compared to normal tissue and glycolysis was also shown to be performed up to 30-times quicker in cancer cells [18]. Yet, increased glucose uptake is also seen under non-hypoxic conditions in cancer tissue [17]. Nonetheless, hypoxia in cancer correlates with parameters of increased cancer aggressiveness, such as chemotherapy resistance [19] and an increased risk for the formation of metastasis [20]. Recently, it could be shown that the reliance on glycolysis has several advantages for cancer cells. First of all, it allows the generation of side products by removing intermediates from the citric cycle [21]. Secondly, the production of lactate itself provides a powerful tool for cancer cells: The secretion of lactate to the surrounding tissue leads to an acidification, to which most host cells cannot adapt and therefore die [22,23]. Additionally, an angiogenetic effect of lactate has been shown recently [24]. Many cancer types show an overexpression of GLUT1 [25]. This observation correlates with many parameters employed to determine cancer aggressiveness, such as high potential to invade surrounding tissues, high risk for the generation of metastasis and chemotherapy resistance [26,27].
2-Deoxy-2-(18F)fluoro-glucose ([18F]-FDG; see Figure 2) is one of the most well-known agents for molecular imaging. Because of its structural and chemical properties, it is taken up like unmodified glucose and is phosphorylated by hexokinase subsequently, leading to intracellular retention. However, it is not further metabolized afterwards due to the missing 2-hydroxyl-group [28]. Since glucose-6-phosphate and structural derivatives, such as 2-deoxy-2-(18F)fluoro-glucose-6-phosphate, have no GLUT substrate anymore, this leads to a tracer accumulation in tissues with high glucose uptake, such as cancer tissues, brain and liver [29]. This accumulation is also due to the differential regulation of other proteins of the glucose metabolism, such as a down-regulation of glucose-6-phophatase [30] and increased glucose uptake triggered by the overall up-regulation of downstream glycolytic enzymes [31]. These effects lead to increased glucose uptake and FDG accumulation in tissues with high enzymatic activity. FDG-PET imaging subsequently allows the detection of tissues showing high glucose accumulation, by monitoring the incorporated β+ emitting 18F.
Figure 2.
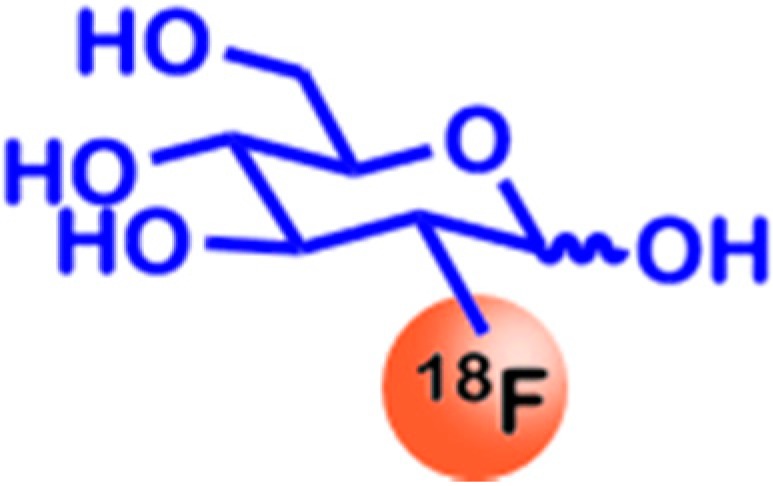
Chemical structure of 2-Deoxy-2-(18F)fluoro-glucose ([18F]-FDG). [18F]-FDG is taken up by the glucose transport system. As many cancer types show higher metabolic glucose turnover, [18F]-FDG uptake is increased, leading to its accumulation in cancer tissue. Subsequently, imaging with positron emission tomography (PET) is possible due to the β+ emitting radionuclide 18F.
[18F]-FDG-PET has been successfully used for detection and tumor staging of different cancer types over the last 20 years, such as lung cancer [32], breast cancer [33], renal cell carcinomas [34] and many others, as reviewed by Gallamini et al. [35] and Hawkins et al. [36].
2.2. Amino Acid Utilization
Due to their charge, amino acids are unable to permeate cell membranes. The carriers responsible for amino acid uptake belong to the solute carrier families SLC3 and SLC7 [37]. SLC7 carriers can be divided into two groups: cationic amino acid transporter (CAT) and l-type amino acid transporters (LAT). CATs perform sodium independent l-type amino acid uptake by facilitated diffusion [38,39,40]. LATs, on the other hand, mainly work as amino acid antiporters, by exporting non-essential amino acids in exchange for essential amino acids [37,41,42].
LATs always occur as heterodimers with members of the SLC3 family. The heterodimeric construct is then called HAT (heterodimeric amino acid transporter). There are only two known members of the SLC3 family: 4F2hc and rBAt, which belong to the group of type II transmembrane N-glycoproteins [43]. The SLC3 members are responsible for the cellular trafficking of the receptor towards the plasma membrane [44]. The substrate specificity of HATs mainly depends on the LAT incorporated in the heterodimer [37].
4F2hc/LAT1(l-type amino acid transporter 1) is overexpressed in many kinds of tumors, such as cervical carcinoma [45], gastric carcinoma [46] and different forms of leukemia—to name only a few [41,47]. High LAT1 expression is associated with parameters of high cancer aggressiveness, such as lymph node metastasis and angiogenesis [48,49]. Another amino acid transporter that is often overexpressed in cancer is ASCT2 (ASC amino-acid transporter 2), which is responsible for the sodium dependent uptake of neutral amino acids [50,51]. It is overexpressed in prostate cancer and hepatocellular cancer and is also associated with high cancer aggressiveness [52].
Similar to the imaging with [18F]-FDG, radiolabeled amino acids can be used to image tissues with increased amino acid throughput. This is especially the case for cancer cells, which usually replicate very quickly and therefore require higher amount of amino acids [51].
The main application of amino acid radiotracers is the detection of tumors characterized by varying glucose uptake, which makes them impossible to monitor reliably and reproducibly with [18F]-FDG. One such example is neuroendocrine tumors [53,54].
As amino acid transporters are ubiquitously expressed, a high selectivity of the tracer molecule is of major importance. In the last years a huge amount of amino acid radiotracers has been developed for PET or SPECT [55]. Here, we only focus on a few selected tracer molecules.
A few years ago, it could be shown that the SPECT tracer l-3-[123I]-iodo-α-methyl-tyrosine shows high LAT1 specificity, compared to other uptake mechanisms [56]. The major disadvantage of this tracer is that it has to be monitored with SPECT, which has a significantly lower resolution than PET. Thus, 18F-labeled compounds, such as l-[3-18F]-α-methyl-tyrosine ([l8F]-FAMT) have been developed (see Figure 3), in order to benefit from the high PET resolution [57]. This development only became possible upon the discovery that the α-methyl group is responsible for LAT1 specificity [58]. [l8F]-FAMT is currently one of the most specific and most often used PET tracers for the imaging of cancers with low glucose uptake rates [59,60].
Figure 3.
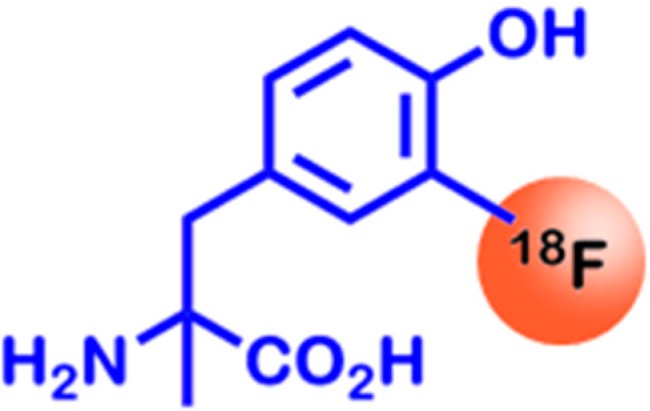
Chemical structure of l-[3-18F]-α-methyl-tyrosine, a PET tracer specifically taken up via amino acid transporters, which are overexpressed in many types of cancer.
2.3. Somatostatin Receptors
Somatostatin receptors (SSTRs) play an important role in the inhibition of growth hormone secretion [61], as well as the production and secretion of gastrointestinal hormones, such as gastrin, renin, glucagon and insulin [62]. SSTRs are known to internalize upon ligand binding [63]. The ligand, somatostatin, is a peptide hormone that is secreted by different tissues of the gastrointestinal tract and stomach. Somatostatin is activated by protease cleavage and occurs in two versions—somatostatin-14 and somatostatin-28 [64].
SSTR2 is the most abundant receptor isoform and is overexpressed in many tumors; especially in neuroendocrine tumors [65]. Additionally, it shows the highest SSTR internalization rate, which makes it a valuable target for molecular imaging and targeted therapy [63,66]. As somatostatin has a short biological half-life (about 3 min) [67], more stable synthetic versions have been developed for clinical applications [64,68]. In 1978, Vale et al. published a comparative analysis of a series of somatostatin analogues, which have been improved further since then [69]. Currently, octreotide is one of the most often used somatostatin analogues. It shows high binding affinity to SSTR2 [65] and exhibits a 19-fold stronger inhibitory effect on the growth hormone secretion than native somatostatin [70].
One of the most well-known octreotide derivatives for therapeutic and imaging purposes is DOTATOC (DOTA(0)-Phe(1)-Tyr(3))octreotide) (see Figure 4). Chemically, it consists of the chelator DOTA (1,4,7,10-tetraazacyclododecane-1,4,7,10-tetraacetic acid) which is linked to octreotide via a peptide bond. The octreotide employed was further modified by substituting phenylalanine at position three with tyrosine, in order to increase the stability of the compound [71]. DOTA is able to chelate many different radionuclides, such as 68Ga, 111In or 90Y. While 68Ga and 111In are used for molecular imaging and cancer stratification via PET and SPECT, 90Y can be used to subsequently treat patients with SSTR-overexpressing cancers [72].
Figure 4.

Chemical structure of [68Ga]-DOTATOC (DOTA(0)-Phe(1)-Tyr(3))octreotide). It shows high affinity to SSTR2 (Somatostatin Receptor 2)—a receptor overexpressed in neuroendocrine tumors. The blue portion of the molecule is responsible for the SSTR2 specificity, whereas the radionuclide 68Ga contained in the chelator DOTA (red) acts as the signal source for PET imaging. For reasons of clarity, chelation bonds are omitted.
In a comparative study, it could be shown recently that the radionuclide incorporated in the tracer has a considerable influence on the target specificity. Gallium chelated octreotide derivatives showed an increased SSTR2 specificity by up to factor five, when compared to the same tracer carrying 111In. Additionally the tumor uptake was 2.5-times higher for gallium carrying tracer molecules. Kidney uptake was also shown to be decreased compared to 111In-octreotide tracers [73,74].
Recently, a next-generation somatostatin analogue (SOM230, Signifor®, Novartis, Switzerland) has been developed for therapeutic purposes. Signifor® was granted an orphan drug status for therapy of the Cushing’s disease by both EMA in 2009 and FDA in 2012. SOM230 distinguishes from its predecessors through its high affinity to all five SSTRs. Moreover, its potency is 3- to 4-fold higher than that of octreotide and elimination from the circulation is prolonged about 10-times to a physical half-life of 23 h [75]. So far, SOM230 is not used as a radiotracer for molecular imaging. Nonetheless, this application could play an important role in the future, due to the high importance of octreotide based tracers for the clinical molecular imaging routine and the improved performance of SOM230 as compared to other SSTR-tracers.
2.4. Integrins
Integrins are a family of heterodimeric receptors and play an important role in cell adhesion and cell signaling. So far, 19 α- and 8 β-integrin-subunits are known, which occur in 24 different arrangements in most vertebrates [76,77]. Receptor specificity mainly depends upon the α-subunit incorporated in the receptor [78]. The 24 members of the vertebrate integrin family are able to interact with several different molecules, such as collagen, laminin and RGD-motif (Arg-Gly-Asp) containing molecules, e.g., fibronectin and vitronectin [79]. Upon ligand binding, integrins induce different intracellular signaling pathways [80]. They play an important role in a multitude of processes that are crucial for tumor progression such as up-regulation and recruitment of tissue-metalloproteases, migration and angiogenesis [81,82]. The most common and well-studied integrins playing an important role in cancer, especially in promoting angiogenesis, are those containing an αv-subunit. [83]. Overexpression of intergin αvβ3 is most often seen in glioblastomas and melanomas [84,85]. Since the α-subunit mainly determines the receptor specificity, most αv containing integrins can be targeted with similar tracers containing an RGD-motif.
The first linear RGD-tracers were developed in 2001 [86] and later on modified by developing more stable cyclic RGD-tracers. In order to decrease protease digestibility of the compound, d-amino acids have been incorporated. There are numerous different RGD-tracers nowadays, as reviewed by Cai et al. [87]. Cyclic RGDfV (Cyclo [Arg-Gly-Asp-d-Phe-Val]) is the most prominent lead structure to target integrin αvβ3 [88,89]. In order to generate a theranostic compound, radiolabeling with both diagnostic and therapeutic radionuclides has to be possible. For this purpose, coupling of chelators is very useful, because they allow quick and variable radionuclide usage (see Figure 5).
Figure 5.

Chemical structure of [68Ga]-NOTA-c(RGDyK) (NOTA-Cyclo [Arg-Gly-Asp-d-Tyr-Lys]), which is used for the molecular imaging of αvβ3 integrin overexpressing tumors. 68Ga functions as the signal source for molecular imaging and is chelated to NOTA (red) to prevent displacement of the imaging moiety. The RGD-motif (Arg-Gyl-Asp, blue) is responsible for the integrin interaction. Insertion of d-tyrosine and cyclization reduce the protease digestibility of the compound, while lysine enables the correct positioning of the imaging moiety. For reasons of clarity, chelation bonds are omitted.
Recently, it was observed that Ga3+ ions are too small to perfectly fit into DOTA [90]. Additionally, DOTA-conjugated RGD-tracers showed high blood pool activity due to unspecific interactions with plasma proteins. Thus, smaller chelators, such as NOTA (1,4,7-triazacyclononane-triacetic acid) became more important. However, coupling of chelators to c(RGDfV) was not possible, because all chemically addressable groups could not be modified without influencing the pharmacophore. For this reason, valine was substituted by lysine in order to incorporate an addressable amino functionality. Additional modifications, such as other d-amino acids, were mainly introduced in order to influence pharmacokinetic parameters and increase the serum stability by preventing protease cleavage [89,91]. NOTA-c(RGDyK) showed high integrin αvβ3 affinity (Ki = 3.6 nM) and decreased blood pool activity [92].
2.5. Folate Receptors
Folic acid (pteroylmono-glutamic acid) plays an important role in the biosynthesis of amino acids, purines and thymidylates by acting as a C1-group donor [93]. Mammals are unable to synthesize folate and therefore rely on their diet to keep their folate levels constant [94].
So far, three different folate uptake mechanisms are known: reduced folate carriers, proton coupled folate transporters and folate receptors (FR). Reduced folate carriers are bidirectional antiporters that import folate in exchange for negatively charged biomolecules, such as adenosine phosphates [95]. Proton coupled folate transporters, on the other hand, are proton/folate symporters that deploy energy derived from the transmembrane proton gradient to import folate against its concentration gradient into the cell [96]. Both of the previously mentioned transport mechanisms show low folate binding affinity in comparison to the FR (KD ≈ 10−10 M) [97]. The FR exists in three different isoforms (FRα, FRβ and FRγ) and leads to folate internalization by receptor mediated endocytosis [98]. FR overexpression has been detected in different tumor entities, mainly arising from epithelial tissue, as summarized in Table 1. Additionally FR overexpression correlates with poor prognosis, due to high cancer aggressiveness and chemotherapy resistance [99].
Table 1.
Folate receptor overexpression in different cancer types [100]. FR, folate receptors.
| Cancer Type | Rate of FR Overexpression |
|---|---|
| Ovarian | 93% |
| Endometrial | 90% |
| Renal | 50% |
| Lung | 33% |
| Colorectal | 22% |
| Breast | 21% |
One of the most promising approaches in targeting FR overexpressing tumors is the utilization of folate derivatives, which feature high binding affinities and clear, thus patentable, structures [101]. Vintafolide/etarfolatide, two compounds developed by Endocyte Inc. (West Lafayette, IN, USA) and Merck & Co (Kenilworth, NJ, USA), provide an example for this group of compounds. As shown in Figure 6, both compounds consist of folic acid, which is responsible for the targeting, and either a diagnostic or a therapeutic moiety. The therapeutic component, vintafolide, contains a hydrophilic peptide spacer, a cleavable disulfide-linker and the therapeutic agent deacetylvinblastine monohydrazide, which is a microtubule inhibiting agent [102]. Etarfolatide, on the other hand, contains a non-cleavable peptide bond to prevent displacement of the chelator together with the imaging compound 99mTc [103,104,105]. Early clinical trials proved good results for vintafolide and etarfolatide, however, both compounds have been withdrawn from the European market for the indication of “FR-positive, platinum-resistant ovarian cancer in adult women”, since no significant improvement compared to the current therapy could be shown. Nonetheless, vintafolide is still in clinical trials for its application in “FR-positive, recurrent non-small cellular lung carcinoma”. Most recently, it could be shown that combination therapy with docetaxel leads to increased overall survival rates [106].
Figure 6.

(a) Chemical structure of etarfolatide. It is used to image folate receptor positive cancers with SPECT. It contains folate (blue) to target the compound to the FR and chelated 99mTc in order to allow imaging (red); (b) Chemical structure of vintafolide, which is used to treat folate receptor positive ovarian cancer. It also contains folate as targeting moiety (blue). Additionally, a hydrophilic peptide linker with a cleavable disulfide bond (green) is incorporated to allow the release of deacetylvinblastine monohydrazide (red), which exhibits the cytotoxic effect. For reasons of clarity, chelation bonds are omitted.
2.6. CD20
CD20 is an integral membrane protein exclusively expressed on B cells, with the exception of fully differentiated plasma cells, and may thus be used as a B-cell marker [107,108]. It plays an important role in B-cell differentiation and activation [109], cell cycle progression [110] and Ca2+ uptake [111,112]. Additionally, CD20 is significantly involved in altering gene expression patterns [113] by activating down-stream signaling cascades, such as the c-Myc pathway [114].
The natural ligand of CD20 is not known so far [115,116]. CD20 is expressed on the surface of most malignant B-cells, even though expression levels may vary significantly [108]. Aggressive B-cell lymphomas very often show high expression levels, correlating with the cancer associated tissue dedifferentiation, which leads to CD20 up-regulation [117].
Zevalin® (ibritumomab-tiuxetan, Bayer Healthcare, Leverkusen, Germany) is a monoclonal anti-human-CD20 antibody that is covalently coupled to the chelator molecule tiuxetan (see Figure 7). In combination with the radionuclides 90Y (therapeutic nuclide) and 111In (diagnostic nuclide) it is approved for the treatment of “relapsed or refractory low-grade, follicular or transformed B-cell non-Hodgkin’s lymphoma”.
Figure 7.

111In-ibritumomab-tiuxetan (Zevalin®). The monoclonal antibody ibritumomab (grey) is highly specific for CD20, which is overexpressed by many different lymphomas. The chelator tiuxetan (red) is able to bind 111In, as well as 90Y. This allows molecular imaging and the subsequent therapy of CD20 positive lymphomas. For reasons of clarity, chelation bonds are omitted.
In the US, molecular imaging using SPECT with 111In-ibritumomab-tiuxetan is mandatory in order to verify the presence of CD20 prior to applying the therapeutic antibody. Additionally, correct biodistribution of the therapeutic compound can be confirmed by post-therapeutic imaging [118].
After having blocked CD20 on non-cancerous B-cells using 250 mg/m2 unlabeled rituximab, 185 MBq of 111In-Zevalin® is applied four hours later, followed by subsequent SPECT imaging.
In a comparative study the overall response rate (ORR) of therapeutic Zevalin® was 80% while the non-radiolabeled Rituximab only showed an ORR of 56%. The total time to progression was greater than 12 months in 37% of patients treated with Zevalin® [119,120]. It is very likely that this increase is, at least in part, also due to the previous cancer stratification by molecular imaging. Thus, only patients with an expected positive outcome are treated.
2.7. Her2
Her2 (human epidermal growth factor receptor 2) is a transmembrane glycoprotein belonging to the epidermal growth factor receptor family [121]. Members of this family are receptor tyrosine kinases: receptor dimers are formed upon ligand binding, which induces intracellular tyrosine side chain phosphorylation and leads to downstream signaling. Epidermal growth factor receptors play an important role in cell growth, cell differentiation and cell survival [122].
In contrast to other epidermal growth factor receptors, Her2 is constitutively activated and thus leads to permanent intracellular signaling. Her2 is overexpressed in many different cancer types, such as gastric cancer, ovarian cancer or non-small-cellular lung carcinomas [123]. It is also overexpressed in a significant portion (about 20%) of breast cancers. Her2 overexpression is associated with increased cell proliferation rates and chemotherapy resistance [124].
Her2 positive breast cancer is commonly treated by administration of the monoclonal antibody trastuzumab (Herceptin®, Genentech, San Francisco, CA, USA) or the antibody-drug-conjugate ado-trastuzumab emtansine (Kadcyla®, Roche, Basel, Switzerland) [125]. Ado-trastuzumab emtansine contains on average 3.5 covalently coupled DM1 molecules per antibody. DM1 induces cell death by tubuline inhibition [126].
In order to ensure a therapeutic effect, tumor staging is inevitable. Currently, radiotracers for in vivo imaging are being developed, in order to facilitate diagnosis and stratification as compared to the currently applied ex vivo assays, such as immunohistochemistry or fluorescence in situ hybridization. Due to their high affinity, many Her2 antibodies serve as pharmaceutical leads for the development of new Her2 tracer molecules, which holds one major disadvantage, though: having very long physical half-lives, antibodies impede the performance of molecular imaging using common radionuclides. [125]. Current approaches rely on the incorporation of long-lived radionuclides, such as 89Zr which is characterized by its long physical half-life and its biologic inertness [127]. Nonetheless, 89Zr leads to a disadvantageous 2.5-fold higher radiation exposure when compared to conventional FDG-PET [128,129].
In order to decrease the radiation dose, alternative systems have been developed, such as [64Cu]-DOTA-trastuzumab. The utilization of 64Cu leads to significantly lower radiation doses, as compared to 89Zr. Yet, high blood pool activity was observed, making it impossible to detect tumors or metastasis localized next to organs with high blood perfusion, such as heart and liver [130].
Another approach to reduce the radiation exposure is the utilization of alternative binders, such as affibodies [131]. Affibody molecules are small (6.5 kDa) single domain proteins derived from an engineered B-domain of the Staphylococcus aureus protein A, which is then called Z-domain [132,133]. It is characterized by high thermal and chemical stability, rapid folding and high solubility [134]. Affibody scaffolds do not contain any cysteine. The introduction of a single cysteine therefore enables site-specific modifications [135].
In 2010, the first Her2-specific Affibody molecule, ZHer2:342-pep2 (ABY-002, Affibody AB, Solna, Sweden), was tested in human beings. Two differently radiolabeled variants ([111In]-ABY-002 and [68Ga]-ABY-002) were applied. Both compounds showed specific tumor uptake and allowed for high contrast imaging of Her2-positive tissue [136]. Nonetheless, a comparative mouse study revealed that [111In]-ABY-002 shows high blood pool activity, as well as lung and spleen accumulation [137].
In the last years, the Affibody scaffold of ABY-002 was further modified, in order to increase specificity, stability and to allow site-specific modifications. One of the most promising variants is ABY-025 (maleimide-monamide-DOTA-Cys61-ZHer2:2891-Cys) [134]. In preclinical animal studies, [111In]-ABY-025 could be shown to be neither toxic nor immunogenic [138]. Additionally, it showed high Her2-specificity and affinity (KD ≈ 76 pM) [134]. First in-human studies revealed high tolerability of the compound and specific uptake in Her2-positive cancer tissues. High background-uptake was observed for kidneys, liver and spleen [139].
2.8. hNIS
The human sodium iodide symporter (hNIS) is responsible for cellular iodide uptake deploying the physiological sodium gradient. This leads to an intracellular iodide accumulation with concentrations that are 20- to 40-fold higher than in the physiological extracellular environment [140].
hNIS belongs to the group of sodium/solute symporters of the solute carrier 5 family [141,142]. It is mainly expressed in the thyroid, where it imports iodide for the synthesis of the thyroid hormones tri-iodothyronine (T3) and thyroxine (T4). Among other effects, these hormones play an important role in the development of the nervous system and the lungs as well as the skeleton and the muscles [142]. High expression levels of hNIS are also seen in the breast tissue of pregnant and lactating women, especially in the lactating mammary gland. The physiological function of hNIS therin is to secrete iodide into the milk, in order to supply the newborn with sufficient amounts of iodide for the synthesis of thyroid hormones [143,144]. In accordance with these findings, hNIS was also found in the placenta, where it ensures the iodide supply of the unborn [145]. Lower, yet detectable, expression levels of hNIS can be found in various tissues, such as the salivary gland [146,147], the testis [148] and the intestines [149].
hNIS is mainly overexpressed in thyroid cancer [150] and estrogen receptor positive breast cancer [151]. Yet, it must be stated that changes in the intracellular trafficking lead to accumulation of hNIS in the cytosol [152]. Thus, neither thyroid cancer nor breast cancer show higher extracellular presence of hNIS when compared to non-cancerous tissue of the same origin. Nonetheless, radioiodine therapy is a powerful tool to reduce hNIS positive diffuse or metastatic thyroid cancer tissue [153]. Additionally, non-cancerous breast tissue is rarely affected by radioiodine therapy or imaging, because hNIS in the breast is only expressed during pregnancy and lactation [154]. In this period of time, radiotherapy is not in compliance with current health care standards [153].
Radioiodine therapy relies on 131I in order to generate a therapeutic effect. It is applied in the form of sodium iodide [155]. SPECT imaging of hNIS overexpressing tissues is possible by using 123I, instead of the therapeutic radionuclide. 123I imaging is mainly applied for whole body imaging of thyroid cancers [156,157]. Recently, the superiority of 123I over 131I for SPECT imaging of the thyroid could also be shown [158]. Another approach for the imaging of hNIS overexpressing tissues in thyroid and breast relies on the utilization of 99mTc-pertechnetate, which also functions as a substrate of hNIS [159,160].
2.9. Prostate Specific Membrane Antigen
The prostate specific membrane antigen (PSMA) is a type II transmembrane glycoprotein and zinc metalloprotease with glutamate-carboxypeptidase functionality, which cleaves the glutamine fraction of folate [161]. PSMA is expressed in low levels on the surface of most prostate cells. Prostate cancer often shows PSMA overexpression [162,163,164]. PSMA expression is inversely correlated to androgen levels and overexpression is thus seen most often in androgen independent prostate cancer [165,166]. Additionally, PSMA is expressed on the vascular epithelium of different cancer types [167,168].
The exact function of PSMA is yet unknown, but it is thought to play an important role in cell signaling and nutrient uptake [169]. It is also known that PSMA internalizes upon ligand binding via clathrin-dependent endocytosis, which makes it a favorable target for imaging and therapy [170,171].
In the beginning, PSMA was visualized in biopsy and tissue samples by the antibody 111In-capromab pendetide (ProstaScint®, Cytogen, Princeton, NJ, USA) [172]. In vivo imaging using radiolabeled ProstaScint® is not possible, because it recognizes an epitope that is localized intracellularly in living cells [173]. Additionally, antibodies are more difficult to handle for in vivo imaging purposes, because they show high blood circulation times compared to small molecules. This long physical half-life prolongs the circulation time of the radioisotopes, which thereby damage untargeted tissue [125].
Molecular imaging and targeted therapy finally became possible upon the discovery of structural and, later on, functional correlations of PSMA and the N-acetylaspartylglutamat peptidase NAALADASE [174,175]. This led to the development of PSMA tracers derived from inhibitors of NAALADASE [176,177]. Currently, most tracer molecules contain the targeting sequence Glu-urea-Lys, where the ε-amino group of lysine can be used for site specific modifications.
Recently, a new PSMA tracer was published by Eder et al. [178], carrying the acyclic chelator HBED-CC via an aminocaproic acid linker, which ensures reduced interaction between the chelator and the binding pocket of PSMA (see Figure 8). The utilization of HBED-CC has several benefits over other gallium chelators: firstly, it allows quick and stable chelation with 68Ga at low temperatures [179] and secondly, it adds an aromatic moiety to the tracer, which is useful for strong PSMA-tracer interactions and internalization [180]. This property makes PSMA-HBED-CC a valuable tracer for molecular imaging of PSMA-overexpressing cancers.
Figure 8.

Chemical structure of [68Ga]-PSMA-HBED-CC. The Glu-urea-Lys sequence (blue) is responsible for the targeting properties of the tracer. It is linked to the 68Ga-labeledchelator HBED-CC (red) via an aminocaproic acid linker (green). For reasons of clarity, chelation bonds are omitted.
3. Methods for Molecular Imaging—An Overview
3.1. Computed Tomography (CT)
CT is used to gain information about anatomical structures by monitoring X-ray absorption. The observed differences of intensity depend on the interaction between electromagnetic radiation and solid matter [181]. Additionally, photoelectric absorption correlates with increasing ordinal numbers, which requires the application of contrast agents containing heavy atoms. The most prominent examples are iodine containing CT contrast agents, such as iodixanol (Visipaque®, GE Healthcare, Buckinghamshire, UK) [182]. In order to generate functional information deploying CT, high amounts of contrast agents have to be applied, which commonly increases side effects [183]. In consequence, CT is currently not considered to become an imaging technique for molecular imaging purposes (see Table 2).
Table 2.
| Method | Spatial Resolution | Temporal Resolution | Sensitivity [mol/L] | Costs | Advantages | Drawbacks |
|---|---|---|---|---|---|---|
| CT | 50–200 µm | Minute | – | Low | Generation of anatomical images | It is difficult to generate functional, non-quantitative |
| MRI | 25–100 µm | Minute to hour | 10−3–10−5 | Very high | High spatial resolution, non-radioactive tracers | Low sensitivity |
| PET | 2–5 mm | Second to minute | 10−11–10−12 | Very high | The most sensitive imaging method, quantitative method, allows to use biologically relevant radionuclides | Imaging of large areas is expensive, low spatial resolution |
| SPECT | 7.5–10 mm | Minute | 10−10–10−11 | High | Simultaneous multi-probe imaging is possible | Low spatial resolution |
CT: Computed Tomography; MRI: Magnetic Resonance Imaging; PET: Positron Emission Tomography; SPECT: Single Photon Emission Computed Tomography.
3.2. Magnetic Resonance Imaging (MRI)
MRI represents an additional imaging technique for the visualization of anatomical structures, utilizing the proton density and the respective chemical environment of an object [186,187]. Imaging contrast can be increased by using contrast agents carrying paramagnetic or super-paramagnetic substances, which influence the T1 relaxation (transversal relaxation) or the T2 relaxation (longitudinal relaxation) respectively. The most commonly used MRI contrast agents contain gadolinium [188], manganese (both paramagnetic substances) [189], or iron oxide crystals (super-paramagnetic contrast agents) [190]. As in CT, MRI usually requires high amounts of contrast agents in order to gain functional information. This again limits its applicability for molecular imaging purposes [183] (see Table 2).
3.3. Positron Emission Tomography (PET)
PET is the most important method for molecular imaging in oncology [191]. It depends on the detection of gamma rays in order to gain anatomical, as well as functional, information. PET tracers are labeled with radionuclides undergoing β+ decay and thereby emitting positrons, passing the surrounding tissue for a characteristic distance. Interaction with existent electrons leads to annihilation, resulting in the emission of two collinear γ photons traveling in opposite directions. Thus, the detection of two separate signals at different detectors localized opposite one another is enabled within a very small timeframe. Only such coincident events at opposite positions are monitored and allow the determination of the trajectory on which the annihilation occurred. Monitoring many of these coincident events allows to determine the spatial distribution of the tracer molecule [7].
Additionally, modern PET instruments monitor the time difference between the detected events. This time of flight analysis facilitates a more precise localization of the site of decay [192]. In the last years, multimodality imaging, such as PET/CT became increasingly important, because of their high sensitivity and the possibility to simultaneously generate functional and anatomical data [193,194]. Positron emission is triggered in vivo by using tracers containing β+ emitting radionuclides. Common representatives are 11C, 13N, 15O, 18F, 68Ga and 124I [8,184] (see Table 2).
3.4. Single Photon Emission Computed Tomography (SPECT)
SPECT is a method for molecular imaging that relies on monitoring low energy γ-rays by gamma cameras in order to collect anatomical and functional information. There are three main differences between PET and SPECT: Firstly, PET and SPECT rely on different radionuclides for imaging due to their different imaging processes. In comparison to positron emission tomography, the radiation energy emitted during SPECT is substantially lower (see Table 3). Moreover the emission is measured directly by using single photon emitting radionuclides such as 99mTc and 111In; whereas PET scanners detect coincident events driven by photon pairs [8]. Secondly, PET and SPECT differ in their maximum resolutions, and thirdly in the area they are able to monitor simultaneously [184,195] (see Table 2).
Table 3.
Radionuclides used in oncology.
| Radionuclide | Decay | Half-Life | Energy | Application | Source |
|---|---|---|---|---|---|
| 11C | β+ | 20 min | 511 keV | Diagnosis (PET) | [7] |
| 13N | β+ | 10 min | 511 keV | Diagnosis (PET) | [7] |
| 18F | β+ | 110 min | 511 keV | Diagnosis (PET) | [7] |
| 64Cu | β+ | 12.7 h | 511 keV | Diagnosis (PET) | [7] |
| 67Ga | γ | 78 h | 93 keV | Diagnosis (SPECT) | [196] |
| 68Ga | β+ | 68 min | 511 keV | Diagnosis (PET) | [7] |
| 86Y | β+ | 14.7 h | 511 keV | Diagnosis (PET) | [197] |
| 89Zr | β+ | 3.3 days | 511 keV | Diagnosis (PET) | [127] |
| 90Y | β− | 2.7 days | 2.28 MeV | Therapy | [198] |
| 99mTc | γ | 6 h | 141 keV | Diagnosis (SPECT) | [199] |
| 111In | γ | 2.8 days | 171 keV | Diagnosis (SPECT) | [199] |
| 123I | γ | 13.2 h | 159 keV | Diagnosis (SPECT) | [200] |
| 124I | β+ | 4.18 days | 511 keV | Diagnosis (PET) | [7] |
| 131I | β− | 8 days | 0.61 MeV | Therapy | [201] |
| 177Lu | β− | 6.7 days | 0.5 MeV | Therapy | [198] |
| 201Tl | γ | 73 h | 80 keV | Diagnosis (SPECT) | [202] |
4. Conclusions
This article provides an overview of current and future applications of molecular imaging in oncology for the purpose of cancer stratification. Molecular imaging enables non-invasive, simultaneous cancer stratification and localization in the patient. Consequently, its potential differs fundamentally from alternative stratification modalities, such as cancer proteomics and immunohistochemistry. Molecular imaging enables to visually prove the presence of target structures that can be deployed for treatment with highly specific drugs. As radioactive decays can be detected with high sensitivity and accuracy, radiotracers are the most important imaging compounds in modern oncology.
Based on the description of the most commonly used imaging methods in oncology (CT, MRI, PET and SPECT), tumor specific cellular structures and cancer phenotypes are elucidated. For each of these structures, the physiological role, the mode of action, as well as the specific role in cancer development and progression is given to outline their respective prognostic value. Subsequently, specific tracer molecules that target or employ the aforementioned structures are introduced.
A multitude of tracers are already in use for diverse target structures, such as receptors (e.g., somatostatin receptors, folate receptors or Her2), transporters (e.g., glucose transporters, amino acid transporters or hNIS), and cell specific target structures (e.g., PSMA, integrins, and CD20). Moreover, molecular imaging provides information on the functional expression of specific targets which is the basis for the application of theranostics. Examples are Zevalin®, DOTATOC or the radiotracer-therapeutic pair etarfolatide/vintafolide. In conclusion, molecular imaging plays a steadily increasing role in cancer stratification and contributes significantly to the current development of personalized medicine in cancer therapy.
Author Contributions
Uwe Haberkorn and Walter Mier wrote the vital passages and designed and revised the manuscript. Justus Weber composed the main part of the manuscript and composed the figures.
Conflicts of Interest
The authors declare no conflict of interest.
References
- 1.Shaw E.C., Johnson P.W.M. Stratified medicine for cancer therapy. Drug Discov. Today. 2012;17:261–268. doi: 10.1016/j.drudis.2012.02.004. [DOI] [PubMed] [Google Scholar]
- 2.Kulasingam V., Diamandis E.P. Strategies for discovering novel cancer biomarkers through utilization of emerging technologies. Nat. Clin. Pract. Oncol. 2008;5:588–599. doi: 10.1038/ncponc1187. [DOI] [PubMed] [Google Scholar]
- 3.Ummanni R., Mundt F., Pospisil H., Venz S., Scharf C., Barett C., Falth M., Kollermann J., Walther R., Schlomm T., et al. Identification of clinically relevant protein targets in prostate cancer with 2D-DIGE coupled mass spectrometry and systems biology network platform. PLoS One. 2011;6:e16833. doi: 10.1371/journal.pone.0016833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Palma P., Cano C., Conde-Muino R., Comino A., Bueno P., Ferron J.A., Cuadros M. Expression profiling of rectal tumors defines response to neoadjuvant treatment related genes. PLoS One. 2014;9:e112189. doi: 10.1371/journal.pone.0112189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Tan H.T., Lee Y.H., Chung M.C. Cancer proteomics. Mass Spectrom. Rev. 2012;31:583–605. doi: 10.1002/mas.20356. [DOI] [PubMed] [Google Scholar]
- 6.Uhl P., Fricker G., Haberkorn U., Mier W. Radionuclides in drug development. Drug Discov. Today. 2014;20:198–208. doi: 10.1016/j.drudis.2014.09.027. [DOI] [PubMed] [Google Scholar]
- 7.Ametamey S.M., Honer M., Schubiger P.A. Molecular imaging with PET. Chem. Rev. 2008;108:1501–1516. doi: 10.1021/cr0782426. [DOI] [PubMed] [Google Scholar]
- 8.Massoud T.F., Gambhir S.S. Molecular imaging in living subjects: Seeing fundamental biological processes in a new light. Genes Dev. 2003;17:545–580. doi: 10.1101/gad.1047403. [DOI] [PubMed] [Google Scholar]
- 9.Weissleder R., Mahmood U. Molecular imaging. Radiology. 2001;219:316–333. doi: 10.1148/radiology.219.2.r01ma19316. [DOI] [PubMed] [Google Scholar]
- 10.Wright E.M., Turk E. The sodium/glucose cotransport family SLC5. Pflugers Arch. 2004;447:510–518. doi: 10.1007/s00424-003-1202-0. [DOI] [PubMed] [Google Scholar]
- 11.Macheda M.L., Rogers S., Best J.D. Molecular and cellular regulation of glucose transporter (GLUT) proteins in cancer. J. Cell. Physiol. 2005;202:654–662. doi: 10.1002/jcp.20166. [DOI] [PubMed] [Google Scholar]
- 12.Joost H.G., Bell G.I., Best J.D., Birnbaum M.J., Charron M.J., Chen Y.T., Doege H., James D.E., Lodish H.F., Moley K.H., et al. Nomenclature of the GLUT/SLC2A family of sugar/polyol transport facilitators. Am. J. Physiol. Endocrinol. Metab. 2002;282:E974–E976. doi: 10.1152/ajpendo.00407.2001. [DOI] [PubMed] [Google Scholar]
- 13.El Sayed S.M., Mahmoud A.A., El Sawy S.A., Abdelaal E.A., Fouad A.M., Yousif R.S., Hashim M.S., Hemdan S.B., Kadry Z.M., Abdelmoaty M.A., et al. Warburg effect increases steady-state ROS condition in cancer cells through decreasing their antioxidant capacities (anticancer effects of 3-bromopyruvate through antagonizing Warburg effect) Med. Hypotheses. 2013;81:866–870. doi: 10.1016/j.mehy.2013.08.024. [DOI] [PubMed] [Google Scholar]
- 14.Jang M., Kim S.S., Lee J. Cancer cell metabolism: Implications for therapeutic targets. Exp. Mol. Med. 2013;45:e45. doi: 10.1038/emm.2013.85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Mamede M., Higashi T., Kitaichi M., Ishizu K., Ishimori T., Nakamoto Y., Yanagihara K., Li M., Tanaka F., Wada H., et al. [18F]FDG uptake and PCNA, Glut-1, and Hexokinase-II expressions in cancers and inflammatory lesions of the lung. Neoplasia. 2005;7:369–379. doi: 10.1593/neo.04577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Meller J., Sahlmann C.O., Scheel A.K. 18F-FDG PET and PET/CT in fever of unknown origin. J. Nucl. Med. 2007;48:35–45. [PubMed] [Google Scholar]
- 17.Horsman M.R. Measurement of tumor oxygenation. Int. J. Radiat. Oncol. Biol. Phys. 1998;42:701–704. doi: 10.1016/S0360-3016(98)00332-0. [DOI] [PubMed] [Google Scholar]
- 18.Ganapathy V., Thangaraju M., Prasad P.D. Nutrient transporters in cancer: Relevance to Warburg hypothesis and beyond. Pharmacol. Ther. 2009;121:29–40. doi: 10.1016/j.pharmthera.2008.09.005. [DOI] [PubMed] [Google Scholar]
- 19.Teicher B.A. Hypoxia and drug resistance. Cancer Metastasis Rev. 1994;13:139–168. doi: 10.1007/BF00689633. [DOI] [PubMed] [Google Scholar]
- 20.Hockel M., Schlenger K., Aral B., Mitze M., Schaffer U., Vaupel P. Association between tumor hypoxia and malignant progression in advanced cancer of the uterine cervix. Cancer Res. 1996;56:4509–4515. [PubMed] [Google Scholar]
- 21.Berg J.M., Tymoczko J.L., L. S. Biochemistry. W.H. Freeman and Company; New York, NY, USA: 2002. Section 17.3, The citric acid cycle is a source of biosynthetic precursors. [Google Scholar]
- 22.Gatenby R.A., Gillies R.J. Why do cancers have high aerobic glycolysis? Nat. Rev. Cancer. 2004;4:891–899. doi: 10.1038/nrc1478. [DOI] [PubMed] [Google Scholar]
- 23.Gatenby R. Altered glucose metabolism and the invasive tumor phenotype. Int. J. Oncol. 1996;8:597–601. [PubMed] [Google Scholar]
- 24.Vegran F., Boidot R., Michiels C., Sonveaux P., Feron O. Lactate influx through the endothelial cell monocarboxylate transporter MCT1 supports an NF-κB/IL-8 pathway that drives tumor angiogenesis. Cancer Res. 2011;71:2550–2560. doi: 10.1158/0008-5472.CAN-10-2828. [DOI] [PubMed] [Google Scholar]
- 25.Yamamoto T., Seino Y., Fukumoto H., Koh G., Yano H., Inagaki N., Yamada Y., Inoue K., Manabe T., Imura H. Over-expression of facilitative glucose transporter genes in human cancer. Biochem. Biophys. Res. Commun. 1990;170:223–230. doi: 10.1016/0006-291X(90)91263-R. [DOI] [PubMed] [Google Scholar]
- 26.Hennipman A., van Oirschot B.A., Smits J., Rijksen G., Staal G.E. Glycolytic enzyme activities in breast cancer metastases. Tumour Biol. 1988;9:241–248. doi: 10.1159/000217568. [DOI] [PubMed] [Google Scholar]
- 27.Brophy S., Sheehan K.M., McNamara D.A., Deasy J., Bouchier-Hayes D.J., Kay E.W. GLUT-1 expression and response to chemoradiotherapy in rectal cancer. Int. J. Cancer. 2009;125:2778–2782. doi: 10.1002/ijc.24693. [DOI] [PubMed] [Google Scholar]
- 28.Smith T.A. FDG uptake, tumour characteristics and response to therapy: A review. Nucl. Med. Commun. 1998;19:97–105. doi: 10.1097/00006231-199802000-00002. [DOI] [PubMed] [Google Scholar]
- 29.Pauwels E.K., Ribeiro M.J., Stoot J.H., McCready V.R., Bourguignon M., Maziere B. FDG accumulation and tumor biology. Nucl. Med. Biol. 1998;25:317–322. doi: 10.1016/S0969-8051(97)00226-6. [DOI] [PubMed] [Google Scholar]
- 30.Nelson C.A., Wang J.Q., Leav I., Crane P.D. The interaction among glucose transport, hexokinase, and glucose-6-phosphatase with respect to 3H-2-deoxyglucose retention in murine tumor models. Nucl. Med. Biol. 1996;23:533–541. doi: 10.1016/0969-8051(96)00037-6. [DOI] [PubMed] [Google Scholar]
- 31.Baumann M., Jezussek D., Lang T., Richter R.T., Brand K. Activities of phosphohexose isomerase and other glycolytic enzymes in normal and tumor tissue of patients with neoplastic diseases: Comparison with serum activities and correlation to tumor staging and grading. Tumour Biol. 1988;9:281–286. doi: 10.1159/000217573. [DOI] [PubMed] [Google Scholar]
- 32.Im H.J., Pak K., Cheon G.J., Kang K.W., Kim S.J., Kim I.J., Chung J.K., Kim E.E., Lee D.S. Prognostic value of volumetric parameters of 18F-FDG PET in non-small-cell lung cancer: A meta-analysis. Eur. J. Nucl. Med. Mol. Imaging. 2014;42:241–251. doi: 10.1007/s00259-014-2903-7. [DOI] [PubMed] [Google Scholar]
- 33.Iakovou I.P., Giannoula E. [Nuclear medicine in diagnosis of breast cancer] Hell. J. Nucl. Med. 2014;17:221–227. doi: 10.1967/s002449910150. [DOI] [PubMed] [Google Scholar]
- 34.Win A.Z. Renal cell carcinoma metastasis to the gallbladder detected by FDG-PET/CT. J. Clin Med. Res. 2014;6:482–486. doi: 10.14740/jocmr1886w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Gallamini A., Zwarthoed C., Borra A. Positron emission tomography (PET) in oncology. Cancers (Basel) 2014;6:1821–1889. doi: 10.3390/cancers6041821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Hawkins R.A., Hoh C.K. PET FDG studies in oncology. Nucl. Med. Biol. 1994;21:739–747. doi: 10.1016/0969-8051(94)90045-0. [DOI] [PubMed] [Google Scholar]
- 37.Fotiadis D., Kanai Y., Palacin M. The SLC3 and SLC7 families of amino acid transporters. Mol. Asp. Med. 2013;34:139–158. doi: 10.1016/j.mam.2012.10.007. [DOI] [PubMed] [Google Scholar]
- 38.Closs E.I., Boissel J.P., Habermeier A., Rotmann A. Structure and function of cationic amino acid transporters (CATs) J. Membr. Biol. 2006;213:67–77. doi: 10.1007/s00232-006-0875-7. [DOI] [PubMed] [Google Scholar]
- 39.Closs E.I. Expression, regulation and function of carrier proteins for cationic amino acids. Curr. Opin. Nephrol. Hypertens. 2002;11:99–107. doi: 10.1097/00041552-200201000-00015. [DOI] [PubMed] [Google Scholar]
- 40.Closs E.I., Lyons C.R., Kelly C., Cunningham J.M. Characterization of the third member of the MCAT family of cationic amino acid transporters. Identification of a domain that determines the transport properties of the MCAT proteins. J. Biol. Chem. 1993;268:20796–20800. [PubMed] [Google Scholar]
- 41.Yanagida O., Kanai Y., Chairoungdua A., Kim D.K., Segawa H., Nii T., Cha S.H., Matsuo H., Fukushima J., Fukasawa Y., et al. Human l-type amino acid transporter 1 (LAT1): Characterization of function and expression in tumor cell lines. Biochim. Biophys. Acta. 2001;1514:291–302. doi: 10.1016/S0005-2736(01)00384-4. [DOI] [PubMed] [Google Scholar]
- 42.Kanai Y., Segawa H., Miyamoto K., Uchino H., Takeda E., Endou H. Expression cloning and characterization of a transporter for large neutral amino acids activated by the heavy chain of 4F2 antigen (CD98) J. Biol. Chem. 1998;273:23629–23632. doi: 10.1074/jbc.273.37.23629. [DOI] [PubMed] [Google Scholar]
- 43.Fort J., de la Ballina L.R., Burghardt H.E., Ferrer-Costa C., Turnay J., Ferrer-Orta C., Uson I., Zorzano A., Fernandez-Recio J., Orozco M., et al. The structure of human 4F2hc ectodomain provides a model for homodimerization and electrostatic interaction with plasma membrane. J. Biol. Chem. 2007;282:31444–31452. doi: 10.1074/jbc.M704524200. [DOI] [PubMed] [Google Scholar]
- 44.Palacin M., Kanai Y. The ancillary proteins of HATs: SLC3 family of amino acid transporters. Pflugers Arch. 2004;447:490–494. doi: 10.1007/s00424-003-1062-7. [DOI] [PubMed] [Google Scholar]
- 45.Uno K., Kuwabara H., Terado Y., Kojima K., Kawakami T., Kamma H., Sakurai H., Sakamoto A., Kurata A. Divergent expression of l-type amino acid transporter 1 during uterine cervical carcinogenesis. Hum. Pathol. 2011;42:1660–1666. doi: 10.1016/j.humpath.2011.01.013. [DOI] [PubMed] [Google Scholar]
- 46.Ichinoe M., Mikami T., Yoshida T., Igawa I., Tsuruta T., Nakada N., Anzai N., Suzuki Y., Endou H., Okayasu I. High expression of l-type amino-acid transporter 1 (LAT1) in gastric carcinomas: Comparison with non-cancerous lesions. Pathol. Int. 2011;61:281–289. doi: 10.1111/j.1440-1827.2011.02650.x. [DOI] [PubMed] [Google Scholar]
- 47.Yoon J.H., Kim I.J., Kim H., Kim H.J., Jeong M.J., Ahn S.G., Kim S.A., Lee C.H., Choi B.K., Kim J.K., et al. Amino acid transport system L is differently expressed in human normal oral keratinocytes and human oral cancer cells. Cancer Lett. 2005;222:237–245. doi: 10.1016/j.canlet.2004.09.040. [DOI] [PubMed] [Google Scholar]
- 48.Kaira K., Oriuchi N., Imai H., Shimizu K., Yanagitani N., Sunaga N., Hisada T., Tanaka S., Ishizuka T., Kanai Y., et al. Prognostic significance of l-type amino acid transporter 1 expression in resectable stage I–III nonsmall cell lung cancer. Br. J. Cancer. 2008;98:742–748. doi: 10.1038/sj.bjc.6604235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kaira K., Oriuchi N., Takahashi T., Nakagawa K., Ohde Y., Okumura T., Murakami H., Shukuya T., Kenmotsu H., Naito T., et al. l-type amino acid transporter 1 (LAT1) expression in malignant pleural mesothelioma. Anticancer Res. 2011;31:4075–4082. [PubMed] [Google Scholar]
- 50.Kekuda R., Prasad P.D., Fei Y.J., Torres-Zamorano V., Sinha S., Yang-Feng T.L., Leibach F.H., Ganapathy V. Cloning of the sodium-dependent, broad-scope, neutral amino acid transporter Bo from a human placental choriocarcinoma cell line. J. Biol. Chem. 1996;271:18657–18661. doi: 10.1074/jbc.271.31.18657. [DOI] [PubMed] [Google Scholar]
- 51.Fuchs B.C., Bode B.P. Amino acid transporters ASCT2 and LAT1 in cancer: Partners in crime? Semin. Cancer Biol. 2005;15:254–266. doi: 10.1016/j.semcancer.2005.04.005. [DOI] [PubMed] [Google Scholar]
- 52.Li R., Younes M., Frolov A., Wheeler T.M., Scardino P., Ohori M., Ayala G. Expression of neutral amino acid transporter ASCT2 in human prostate. Anticancer Res. 2003;23:3413–3418. [PubMed] [Google Scholar]
- 53.Sundin A., Garske U., Orlefors H. Nuclear imaging of neuroendocrine tumours. Best Pract. Res. Clin. Endocrinol. Metab. 2007;21:69–85. doi: 10.1016/j.beem.2006.12.003. [DOI] [PubMed] [Google Scholar]
- 54.Huang C., McConathy J. Radiolabeled amino acids for oncologic imaging. J. Nucl. Med. 2013;54:1007–1010. doi: 10.2967/jnumed.112.113100. [DOI] [PubMed] [Google Scholar]
- 55.McConathy J., Yu W., Jarkas N., Seo W., Schuster D.M., Goodman M.M. Radiohalogenated nonnatural amino acids as PET and SPECT tumor imaging agents. Med. Res. Rev. 2012;32:868–905. doi: 10.1002/med.20250. [DOI] [PubMed] [Google Scholar]
- 56.Shikano N., Kanai Y., Kawai K., Inatomi J., Kim D.K., Ishikawa N., Endou H. Isoform selectivity of 3–125I-iodo-α-methyl-l-tyrosine membrane transport in human l-type amino acid transporters. J. Nucl. Med. 2003;44:244–246. [PubMed] [Google Scholar]
- 57.Pauleit D., Floeth F., Tellmann L., Hamacher K., Hautzel H., Muller H.W., Coenen H.H., Langen K.J. Comparison of O-(2–18F-fluoroethyl)-l-tyrosine PET and 3–123I-iodo-α-methyl-l-tyrosine SPECT in brain tumors. J. Nucl. Med. 2004;45:374–381. [PubMed] [Google Scholar]
- 58.Wiriyasermkul P., Nagamori S., Tominaga H., Oriuchi N., Kaira K., Nakao H., Kitashoji T., Ohgaki R., Tanaka H., Endou H., et al. Transport of 3-fluoro-l-α-methyl-tyrosine by tumor-upregulated l-type amino acid transporter 1: A cause of the tumor uptake in PET. J. Nucl. Med. 2012;53:1253–1261. doi: 10.2967/jnumed.112.103069. [DOI] [PubMed] [Google Scholar]
- 59.Kaira K., Oriuchi N., Shimizu K., Ishikita T., Higuchi T., Imai H., Yanagitani N., Sunaga N., Hisada T., Ishizuka T., et al. Correlation of angiogenesis with 18F-FMT and 18F-FDG uptake in non-small cell lung cancer. Cancer Sci. 2009;100:753–758. doi: 10.1111/j.1349-7006.2008.01077.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Imai H., Kaira K., Oriuchi N., Shimizu K., Tominaga H., Yanagitani N., Sunaga N., Ishizuka T., Nagamori S., Promchan K., et al. Inhibition of l-type amino acid transporter 1 has antitumor activity in non-small cell lung cancer. Anticancer Res. 2010;30:4819–4828. [PubMed] [Google Scholar]
- 61.Brazeau P., Vale W., Burgus R., Ling N., Butcher M., Rivier J., Guillemin R. Hypothalamic polypeptide that inhibits the secretion of immunoreactive pituitary growth hormone. Science. 1973;179:77–79. doi: 10.1126/science.179.4068.77. [DOI] [PubMed] [Google Scholar]
- 62.Reichlin S. Secretion of somatostatin and its physiologic function. J. Lab. Clin. Med. 1987;109:320–326. [PubMed] [Google Scholar]
- 63.Reubi J.C., Waser B., Liu Q., Laissue J.A., Schonbrunn A. Subcellular distribution of somatostatin sst2A receptors in human tumors of the nervous and neuroendocrine systems: Membranous versus intracellular location. J. Clin. Endocrinol. Metab. 2000;85:3882–3891. doi: 10.1210/jcem.85.10.6864. [DOI] [PubMed] [Google Scholar]
- 64.Lüllmann H., Mohr K., Hein L. Hormonsystem. Pharmakologie und Toxikologie. 2010;Volume 17:395–451. [Google Scholar]
- 65.Reubi J.C., Waser B., Schaer J.C., Laissue J.A. Somatostatin receptor sst1–sst5 expression in normal and neoplastic human tissues using receptor autoradiography with subtype-selective ligands. Eur. J. Nucl. Med. 2001;28:836–846. doi: 10.1007/s002590100541. [DOI] [PubMed] [Google Scholar]
- 66.Nouel D., Gaudriault G., Houle M., Reisine T., Vincent J.P., Mazella J., Beaudet A. Differential internalization of somatostatin in COS-7 cells transfected with SST1 and SST2 receptor subtypes: A confocal microscopic study using novel fluorescent somatostatin derivatives. Endocrinology. 1997;138:296–306. doi: 10.1210/endo.138.1.4834. [DOI] [PubMed] [Google Scholar]
- 67.Reichlin S. Somatostatin. N. Engl. J. Med. 1983;309:1495–1501. doi: 10.1056/NEJM198312153092406. [DOI] [PubMed] [Google Scholar]
- 68.Delaunoit T., Rubin J., Neczyporenko F., Erlichman C., Hobday T.J. Somatostatin analogues in the treatment of gastroenteropancreatic neuroendocrine tumors. Mayo Clin. Proc. 2005;80:502–506. doi: 10.4065/80.4.502. [DOI] [PubMed] [Google Scholar]
- 69.Vale W., Rivier J., Ling N., Brown M. Biologic and immunologic activities and applications of somatostatin analogs. Metabolism. 1978;27:1391–1401. doi: 10.1016/0026-0495(78)90081-1. [DOI] [PubMed] [Google Scholar]
- 70.Bauer W., Briner U., Doepfner W., Haller R., Huguenin R., Marbach P., Petcher T.J., Pless J. SMS 201–995: A very potent and selective octapeptide analogue of somatostatin with prolonged action. Life Sci. 1982;31:1133–1140. doi: 10.1016/0024-3205(82)90087-X. [DOI] [PubMed] [Google Scholar]
- 71.Modlin I.M., Pavel M., Kidd M., Gustafsson B.I. Review article: Somatostatin analogues in the treatment of gastroenteropancreatic neuroendocrine (carcinoid) tumours. Aliment. Pharmacol. Ther. 2010;31:169–188. doi: 10.1111/j.1365-2036.2009.04174.x. [DOI] [PubMed] [Google Scholar]
- 72.Frilling A., Weber F., Saner F., Bockisch A., Hofmann M., Mueller-Brand J., Broelsch C.E. Treatment with (90)Y- and (177)Lu-DOTATOC in patients with metastatic neuroendocrine tumors. Surgery. 2006;140:968–976. doi: 10.1016/j.surg.2006.07.030. [DOI] [PubMed] [Google Scholar]
- 73.Antunes P., Ginj M., Zhang H., Waser B., Baum R.P., Reubi J.C., Maecke H. Are radiogallium-labelled DOTA-conjugated somatostatin analogues superior to those labelled with other radiometals? Eur. J. Nucl. Med. Mol. Imaging. 2007;34:982–993. doi: 10.1007/s00259-006-0317-x. [DOI] [PubMed] [Google Scholar]
- 74.Froidevaux S., Eberle A.N., Christe M., Sumanovski L., Heppeler A., Schmitt J.S., Eisenwiener K., Beglinger C., Macke H.R. Neuroendocrine tumor targeting: Study of novel gallium-labeled somatostatin radiopeptides in a rat pancreatic tumor model. Int. J. Cancer. 2002;98:930–937. doi: 10.1002/ijc.10295. [DOI] [PubMed] [Google Scholar]
- 75.Bruns C., Lewis I., Briner U., Meno-Tetang G., Weckbecker G. SOM230: A novel somatostatin peptidomimetic with broad somatotropin release inhibiting factor (SRIF) receptor binding and a unique antisecretory profile. Eur. J. Endocrinol. 2002;146:707–716. doi: 10.1530/eje.0.1460707. [DOI] [PubMed] [Google Scholar]
- 76.Takada Y., Ye X., Simon S. The integrins. Genome Biol. 2007;8:215. doi: 10.1186/gb-2007-8-5-215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Hynes R.O. Integrins: Bidirectional, allosteric signaling machines. Cell. 2002;110:673–687. doi: 10.1016/S0092-8674(02)00971-6. [DOI] [PubMed] [Google Scholar]
- 78.Luo B.H., Carman C.V., Springer T.A. Structural basis of integrin regulation and signaling. Annu. Rev. Immunol. 2007;25:619–647. doi: 10.1146/annurev.immunol.25.022106.141618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Barczyk M., Carracedo S., Gullberg D. Integrins. Cell. Tissue Res. 2010;339:269–280. doi: 10.1007/s00441-009-0834-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Guo W., Giancotti F.G. Integrin signalling during tumour progression. Nat. Rev. Mol. Cell. Biol. 2004;5:816–826. doi: 10.1038/nrm1490. [DOI] [PubMed] [Google Scholar]
- 81.Miranti C.K., Brugge J.S. Sensing the environment: A historical perspective on integrin signal transduction. Nat. Cell. Biol. 2002;4:E83–E90. doi: 10.1038/ncb0402-e83. [DOI] [PubMed] [Google Scholar]
- 82.Hsu M.Y., Shih D.T., Meier F.E., van Belle P., Hsu J.Y., Elder D.E., Buck C.A., Herlyn M. Adenoviral gene transfer of β3 integrin subunit induces conversion from radial to vertical growth phase in primary human melanoma. Am. J. Pathol. 1998;153:1435–1442. doi: 10.1016/S0002-9440(10)65730-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Cai W., Niu G., Chen X. Imaging of integrins as biomarkers for tumor angiogenesis. Curr. Pharm. Des. 2008;14:2943–2973. doi: 10.2174/138161208786404308. [DOI] [PubMed] [Google Scholar]
- 84.Gladson C.L., Cheresh D.A. Glioblastoma expression of vitronectin and the αvβ3 integrin. Adhesion mechanism for transformed glial cells. J. Clin. Investig. 1991;88:1924–1932. doi: 10.1172/JCI115516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Albelda S.M., Mette S.A., Elder D.E., Stewart R., Damjanovich L., Herlyn M., Buck C.A. Integrin distribution in malignant melanoma: Association of the β3 subunit with tumor progression. Cancer Res. 1990;50:6757–6764. [PubMed] [Google Scholar]
- 86.Sutcliffe-Goulden J.L., O’Doherty M.J., Marsden P.K., Hart I.R., Marshall J.F., Bansal S.S. Rapid solid phase synthesis and biodistribution of 18F-labelled linear peptides. Eur. J. Nucl. Med. Mol. Imaging. 2002;29:754–759. doi: 10.1007/s00259-001-0756-3. [DOI] [PubMed] [Google Scholar]
- 87.Cai H., Conti P.S. RGD-based PET tracers for imaging receptor integrin αvβ3 expression. J. Label. Compd. Radiopharm. 2013;56:264–279. doi: 10.1002/jlcr.2999. [DOI] [PubMed] [Google Scholar]
- 88.Aumailley M., Gurrath M., Muller G., Calvete J., Timpl R., Kessler H. Arg-Gly-Asp constrained within cyclic pentapeptides. Strong and selective inhibitors of cell adhesion to vitronectin and laminin fragment P1. FEBS Lett. 1991;291:50–54. doi: 10.1016/0014-5793(91)81101-D. [DOI] [PubMed] [Google Scholar]
- 89.Haubner R.H., Wester H.J., Weber W.A., Schwaiger M. Radiotracer-based strategies to image angiogenesis. Q. J. Nucl. Med. 2003;47:189–199. [PubMed] [Google Scholar]
- 90.Wadas T.J., Wong E.H., Weisman G.R., Anderson C.J. Coordinating radiometals of copper, gallium, indium, yttrium, and zirconium for PET and SPECT imaging of disease. Chem. Rev. 2010;110:2858–2902. doi: 10.1021/cr900325h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Decristoforo C., Hernandez Gonzalez I., Carlsen J., Rupprich M., Huisman M., Virgolini I., Wester H.J., Haubner R. 68Ga- and 111In-labelled DOTA-RGD peptides for imaging of αvβ3 integrin expression. Eur. J. Nucl. Med. Mol. Imaging. 2008;35:1507–1515. doi: 10.1007/s00259-008-0757-6. [DOI] [PubMed] [Google Scholar]
- 92.Knetsch P.A., Petrik M., Griessinger C.M., Rangger C., Fani M., Kesenheimer C., von Guggenberg E., Pichler B.J., Virgolini I., Decristoforo C., et al. [68Ga]NODAGA-RGD for imaging αvβ3 integrin expression. Eur. J. Nucl. Med. Mol. Imaging. 2011;38:1303–1312. doi: 10.1007/s00259-011-1778-0. [DOI] [PubMed] [Google Scholar]
- 93.Davis R.E., Nicol D.J. Folic acid. Int. J. Biochem. 1988;20:133–139. doi: 10.1016/0020-711X(88)90476-4. [DOI] [PubMed] [Google Scholar]
- 94.Zhao R., Matherly L.H., Goldman I.D. Membrane transporters and folate homeostasis: Intestinal absorption and transport into systemic compartments and tissues. Expert Rev. Mol. Med. 2009;11:e4. doi: 10.1017/S1462399409000969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Westerhof G.R., Rijnboutt S., Schornagel J.H., Pinedo H.M., Peters G.J., Jansen G. Functional activity of the reduced folate carrier in KB, MA104, and IGROV-I cells expressing folate-binding protein. Cancer Res. 1995;55:3795–3802. [PubMed] [Google Scholar]
- 96.Qiu A., Jansen M., Sakaris A., Min S.H., Chattopadhyay S., Tsai E., Sandoval C., Zhao R., Akabas M.H., Goldman I.D. Identification of an intestinal folate transporter and the molecular basis for hereditary folate malabsorption. Cell. 2006;127:917–928. doi: 10.1016/j.cell.2006.09.041. [DOI] [PubMed] [Google Scholar]
- 97.Wang X., Shen F., Freisheim J.H., Gentry L.E., Ratnam M. Differential stereospecificities and affinities of folate receptor isoforms for folate compounds and antifolates. Biochem. Pharmacol. 1992;44:1898–1901. doi: 10.1016/0006-2952(92)90089-2. [DOI] [PubMed] [Google Scholar]
- 98.Rothberg K.G., Ying Y.S., Kolhouse J.F., Kamen B.A., Anderson R.G. The glycophospholipid-linked folate receptor internalizes folate without entering the clathrin-coated pit endocytic pathway. J. Cell Biol. 1990;110:637–649. doi: 10.1083/jcb.110.3.637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Toffoli G., Cernigoi C., Russo A., Gallo A., Bagnoli M., Boiocchi M. Overexpression of folate binding protein in ovarian cancers. Int. J. Cancer. 1997;74:193–198. doi: 10.1002/(SICI)1097-0215(19970422)74:2<193::AID-IJC10>3.0.CO;2-F. [DOI] [PubMed] [Google Scholar]
- 100.Garin-Chesa P., Campbell I., Saigo P.E., Lewis J.L., Jr., Old L.J., Rettig W.J. Trophoblast and ovarian cancer antigen LK26. Sensitivity and specificity in immunopathology and molecular identification as a folate-binding protein. Am. J. Pathol. 1993;142:557–567. [PMC free article] [PubMed] [Google Scholar]
- 101.Sudimack J., Lee R.J. Targeted drug delivery via the folate receptor. Adv. Drug Deliv. Rev. 2000;41:147–162. doi: 10.1016/S0169-409X(99)00062-9. [DOI] [PubMed] [Google Scholar]
- 102.Gigant B., Wang C., Ravelli R.B., Roussi F., Steinmetz M.O., Curmi P.A., Sobel A., Knossow M. Structural basis for the regulation of tubulin by vinblastine. Nature. 2005;435:519–522. doi: 10.1038/nature03566. [DOI] [PubMed] [Google Scholar]
- 103.Marchetti C., Palaia I., Giorgini M., de Medici C., Iadarola R., Vertechy L., Domenici L., di Donato V., Tomao F., Muzii L., et al. Targeted drug delivery via folate receptors in recurrent ovarian cancer: A review. OncoTargets Ther. 2014;7:1223–1236. doi: 10.2147/OTT.S40947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Maurer A.H., Elsinga P., Fanti S., Nguyen B., Oyen W.J., Weber W.A. Imaging the folate receptor on cancer cells with 99mTc-etarfolatide: Properties, clinical use, and future potential of folate receptor imaging. J. Nucl. Med. 2014;55:701–704. doi: 10.2967/jnumed.113.133074. [DOI] [PubMed] [Google Scholar]
- 105.Morris R.T., Joyrich R.N., Naumann R.W., Shah N.P., Maurer A.H., Strauss H.W., Uszler J.M., Symanowski J.T., Ellis P.R., Harb W.A. Phase II study of treatment of advanced ovarian cancer with folate-receptor-targeted therapeutic (vintafolide) and companion SPECT-based imaging agent (99mTc-etarfolatide) Ann. Oncol. 2014;25:852–858. doi: 10.1093/annonc/mdu024. [DOI] [PubMed] [Google Scholar]
- 106.Naumann R.W., Coleman R.L., Burger R.A., Sausville E.A., Kutarska E., Ghamande S.A., Gabrail N.Y., Depasquale S.E., Nowara E., Gilbert L., et al. PRECEDENT: A randomized phase II trial comparing vintafolide (EC145) and pegylated liposomal doxorubicin (PLD) in combination versus PLD alone in patients with platinum-resistant ovarian cancer. J. Clin. Oncol. 2013;31:4400–4406. doi: 10.1200/JCO.2013.49.7685. [DOI] [PubMed] [Google Scholar]
- 107.Reff M.E., Carner K., Chambers K.S., Chinn P.C., Leonard J.E., Raab R., Newman R.A., Hanna N., Anderson D.R. Depletion of B cells in vivo by a chimeric mouse human monoclonal antibody to CD20. Blood. 1994;83:435–445. [PubMed] [Google Scholar]
- 108.Anderson K.C., Bates M.P., Slaughenhoupt B.L., Pinkus G.S., Schlossman S.F., Nadler L.M. Expression of human B cell-associated antigens on leukemias and lymphomas: A model of human B cell differentiation. Blood. 1984;63:1424–1433. [PubMed] [Google Scholar]
- 109.Tedder T.F., Boyd A.W., Freedman A.S., Nadler L.M., Schlossman S.F. The B cell surface molecule B1 is functionally linked with B cell activation and differentiation. J. Immunol. 1985;135:973–979. [PubMed] [Google Scholar]
- 110.Golay J.T., Clark E.A., Beverley P.C. The CD20 (Bp35) antigen is involved in activation of B cells from the G0 to the G1 phase of the cell cycle. J. Immunol. 1985;135:3795–3801. [PubMed] [Google Scholar]
- 111.Kanzaki M., Shibata H., Mogami H., Kojima I. Expression of calcium-permeable cation channel CD20 accelerates progression through the G1 phase in Balb/c 3T3 cells. J. Biol. Chem. 1995;270:13099–13104. doi: 10.1074/jbc.270.22.13099. [DOI] [PubMed] [Google Scholar]
- 112.Kanzaki M., Lindorfer M.A., Garrison J.C., Kojima I. Activation of the calcium-permeable cation channel CD20 by α subunits of the Gi protein. J. Biol. Chem. 1997;272:14733–14739. doi: 10.1074/jbc.272.23.14733. [DOI] [PubMed] [Google Scholar]
- 113.Golay J., Cusmano G., Introna M. Independent regulation of c-myc, B-myb, and c-myb gene expression by inducers and inhibitors of proliferation in human B lymphocytes. J. Immunol. 1992;149:300–308. [PubMed] [Google Scholar]
- 114.White M.W., McConnell F., Shu G.L., Morris D.R., Clark E.A. Activation of dense human tonsilar B cells. Induction of c-myc gene expression via two distinct signal transduction pathways. J. Immunol. 1991;146:846–853. [PubMed] [Google Scholar]
- 115.Von Schilling C. Immunotherapy with anti-CD20 compounds. Semin. Cancer Biol. 2003;13:211–222. doi: 10.1016/S1044-579X(03)00018-X. [DOI] [PubMed] [Google Scholar]
- 116.Maloney D.G. Anti-CD20 antibody therapy for B-cell lymphomas. N. Engl. J. Med. 2012;366:2008–2016. doi: 10.1056/NEJMct1114348. [DOI] [PubMed] [Google Scholar]
- 117.Gatter K., Warnke R. Diffuse large B cell lymphoma. In: Jaffe E., Harris N., Stein H., Vardiman J., editors. Pathology and Genetics of Tumors of Haematopoietic and Lymphoid Tissues. IARC Press; Lyon, France: 2001. [Google Scholar]
- 118.Goldsmith S.J. Radioimmunotherapy of lymphoma: Bexxar and Zevalin. Semin. Nucl. Med. 2010;40:122–135. doi: 10.1053/j.semnuclmed.2009.11.002. [DOI] [PubMed] [Google Scholar]
- 119.Witzig T.E., Gordon L.I., Cabanillas F., Czuczman M.S., Emmanouilides C., Joyce R., Pohlman B.L., Bartlett N.L., Wiseman G.A., Padre N., et al. Randomized controlled trial of yttrium-90-labeled ibritumomab tiuxetan radioimmunotherapy versus rituximab immunotherapy for patients with relapsed or refractory low-grade, follicular, or transformed B-cell non-Hodgkin’s lymphoma. J. Clin. Oncol. 2002;20:2453–2463. doi: 10.1200/JCO.2002.11.076. [DOI] [PubMed] [Google Scholar]
- 120.Conti P.S., White C., Pieslor P., Molina A., Aussie J., Foster P. The role of imaging with 111In-ibritumomab tiuxetan in the ibritumomab tiuxetan (Zevalin) regimen: Results from a Zevalin imaging registry. J. Nucl. Med. 2005;46:1812–1818. [PubMed] [Google Scholar]
- 121.Citri A., Yarden Y. EGF-ERBB signalling: Towards the systems level. Nat. Rev. Mol. Cell. Biol. 2006;7:505–516. doi: 10.1038/nrm1962. [DOI] [PubMed] [Google Scholar]
- 122.Hynes N.E., Lane H.A. ERBB receptors and cancer: The complexity of targeted inhibitors. Nat. Rev. Cancer. 2005;5:341–354. doi: 10.1038/nrc1609. [DOI] [PubMed] [Google Scholar]
- 123.Scholl S., Beuzeboc P., Pouillart P. Targeting HER2 in other tumor types. Ann. Oncol. 2001;12:S81–S87. doi: 10.1093/annonc/12.suppl_1.S81. [DOI] [PubMed] [Google Scholar]
- 124.Ignatiadis M., Desmedt C., Sotiriou C., de Azambuja E., Piccart M. HER-2 as a target for breast cancer therapy. Clin. Cancer Res. 2009;15:1848–1852. doi: 10.1158/1078-0432.CCR-08-1844. [DOI] [PubMed] [Google Scholar]
- 125.Capala J., Bouchelouche K. Molecular imaging of HER2-positive breast cancer: A step toward an individualized “image and treat” strategy. Curr. Opin. Oncol. 2010;22:559–566. doi: 10.1097/CCO.0b013e32833f8c3a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Chari R.V., Miller M.L., Widdison W.C. Antibody-drug conjugates: An emerging concept in cancer therapy. Angew. Chem. Int. Ed. Engl. 2014;53:3796–3827. doi: 10.1002/anie.201307628. [DOI] [PubMed] [Google Scholar]
- 127.Zhang Y., Hong H., Cai W. PET tracers based on Zirconium-89. Curr. Radiopharm. 2011;4:131–139. doi: 10.2174/1874471011104020131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Dijkers E.C., Kosterink J.G., Rademaker A.P., Perk L.R., van Dongen G.A., Bart J., de Jong J.R., de Vries E.G., Lub-de Hooge M.N. Development and characterization of clinical-grade 89Zr-trastuzumab for HER2/neu immunoPET imaging. J. Nucl. Med. 2009;50:974–981. doi: 10.2967/jnumed.108.060392. [DOI] [PubMed] [Google Scholar]
- 129.Dijkers E.C., Oude Munnink T.H., Kosterink J.G., Brouwers A.H., Jager P.L., de Jong J.R., van Dongen G.A., Schroder C.P., Lub-de Hooge M.N., de Vries E.G. Biodistribution of 89Zr-trastuzumab and PET imaging of HER2-positive lesions in patients with metastatic breast cancer. Clin. Pharmacol. Ther. 2010;87:586–592. doi: 10.1038/clpt.2010.12. [DOI] [PubMed] [Google Scholar]
- 130.Tamura K., Kurihara H., Yonemori K., Tsuda H., Suzuki J., Kono Y., Honda N., Kodaira M., Yamamoto H., Yunokawa M., et al. 64Cu-DOTA-trastuzumab PET imaging in patients with HER2-positive breast cancer. J. Nucl. Med. 2013;54:1869–1875. doi: 10.2967/jnumed.112.118612. [DOI] [PubMed] [Google Scholar]
- 131.Lofblom J., Feldwisch J., Tolmachev V., Carlsson J., Stahl S., Frejd F.Y. Affibody molecules: Engineered proteins for therapeutic, diagnostic and biotechnological applications. FEBS Lett. 2010;584:2670–2680. doi: 10.1016/j.febslet.2010.04.014. [DOI] [PubMed] [Google Scholar]
- 132.Uhlen M., Guss B., Nilsson B., Gatenbeck S., Philipson L., Lindberg M. Complete sequence of the staphylococcal gene encoding protein A. A gene evolved through multiple duplications. J. Biol. Chem. 1984;259:1695–1702. [PubMed] [Google Scholar]
- 133.Nilsson B., Moks T., Jansson B., Abrahmsen L., Elmblad A., Holmgren E., Henrichson C., Jones T.A., Uhlen M. A synthetic IgG-binding domain based on staphylococcal protein A. Protein Eng. 1987;1:107–113. doi: 10.1093/protein/1.2.107. [DOI] [PubMed] [Google Scholar]
- 134.Feldwisch J., Tolmachev V., Lendel C., Herne N., Sjoberg A., Larsson B., Rosik D., Lindqvist E., Fant G., Hoiden-Guthenberg I., et al. Design of an optimized scaffold for affibody molecules. J. Mol. Biol. 2010;398:232–247. doi: 10.1016/j.jmb.2010.03.002. [DOI] [PubMed] [Google Scholar]
- 135.Lundberg E., Hoiden-Guthenberg I., Larsson B., Uhlen M., Graslund T. Site-specifically conjugated anti-HER2 Affibody molecules as one-step reagents for target expression analyses on cells and xenograft samples. J. Immunol. Methods. 2007;319:53–63. doi: 10.1016/j.jim.2006.10.013. [DOI] [PubMed] [Google Scholar]
- 136.Baum R.P., Prasad V., Muller D., Schuchardt C., Orlova A., Wennborg A., Tolmachev V., Feldwisch J. Molecular imaging of HER2-expressing malignant tumors in breast cancer patients using synthetic 111In- or 68Ga-labeled affibody molecules. J. Nucl. Med. 2010;51:892–897. doi: 10.2967/jnumed.109.073239. [DOI] [PubMed] [Google Scholar]
- 137.Tolmachev V., Velikyan I., Sandstrom M., Orlova A. A HER2-binding Affibody molecule labelled with 68Ga for PET imaging: Direct in vivo comparison with the 111In-labelled analogue. Eur. J. Nucl. Med. Mol. Imaging. 2010;37:1356–1367. doi: 10.1007/s00259-009-1367-7. [DOI] [PubMed] [Google Scholar]
- 138.Ahlgren S., Orlova A., Wallberg H., Hansson M., Sandstrom M., Lewsley R., Wennborg A., Abrahmsen L., Tolmachev V., Feldwisch J. Targeting of HER2-expressing tumors using 111In-ABY-025, a second-generation affibody molecule with a fundamentally reengineered scaffold. J. Nucl. Med. 2010;51:1131–1138. doi: 10.2967/jnumed.109.073346. [DOI] [PubMed] [Google Scholar]
- 139.Sorensen J., Sandberg D., Sandstrom M., Wennborg A., Feldwisch J., Tolmachev V., Astrom G., Lubberink M., Garske-Roman U., Carlsson J., et al. First-in-human molecular imaging of HER2 expression in breast cancer metastases using the 111In-ABY-025 affibody molecule. J. Nucl. Med. 2014;55:730–735. doi: 10.2967/jnumed.113.131243. [DOI] [PubMed] [Google Scholar]
- 140.Rodriguez A.M., Perron B., Lacroix L., Caillou B., Leblanc G., Schlumberger M., Bidart J.M., Pourcher T. Identification and characterization of a putative human iodide transporter located at the apical membrane of thyrocytes. J. Clin. Endocrinol. Metab. 2002;87:3500–3503. doi: 10.1210/jcem.87.7.8797. [DOI] [PubMed] [Google Scholar]
- 141.Jung H. The sodium/substrate symporter family: Structural and functional features. FEBS Lett. 2002;529:73–77. doi: 10.1016/S0014-5793(02)03184-8. [DOI] [PubMed] [Google Scholar]
- 142.De La Vieja A., Dohan O., Levy O., Carrasco N. Molecular analysis of the sodium/iodide symporter: Impact on thyroid and extrathyroid pathophysiology. Physiol. Rev. 2000;80:1083–1105. doi: 10.1152/physrev.2000.80.3.1083. [DOI] [PubMed] [Google Scholar]
- 143.Dohan O., Carrasco N. Advances in Na+/I− symporter (NIS) research in the thyroid and beyond. Mol. Cell. Endocrinol. 2003;213:59–70. doi: 10.1016/j.mce.2003.10.059. [DOI] [PubMed] [Google Scholar]
- 144.Dadachova E., Carrasco N. The Na/I symporter (NIS): Imaging and therapeutic applications. Semin. Nucl. Med. 2004;34:23–31. doi: 10.1053/j.semnuclmed.2003.09.004. [DOI] [PubMed] [Google Scholar]
- 145.Mitchell A.M., Manley S.W., Morris J.C., Powell K.A., Bergert E.R., Mortimer R.H. Sodium iodide symporter (NIS) gene expression in human placenta. Placenta. 2001;22:256–258. doi: 10.1053/plac.2000.0609. [DOI] [PubMed] [Google Scholar]
- 146.Jhiang S.M., Cho J.Y., Ryu K.Y., de Young B.R., Smanik P.A., McGaughy V.R., Fischer A.H., Mazzaferri E.L. An immunohistochemical study of Na+/I− symporter in human thyroid tissues and salivary gland tissues. Endocrinology. 1998;139:4416–4419. doi: 10.1210/endo.139.10.6329. [DOI] [PubMed] [Google Scholar]
- 147.La Perle K.M., Kim D.C., Hall N.C., Bobbey A., Shen D.H., Nagy R.S., Wakely P.E., Jr., Lehman A., Jarjoura D., Jhiang S.M. Modulation of sodium/iodide symporter expression in the salivary gland. Thyroid. 2013;23:1029–1036. doi: 10.1089/thy.2012.0571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Russo D., Scipioni A., Durante C., Ferretti E., Gandini L., Maggisano V., Paoli D., Verrienti A., Costante G., Lenzi A., et al. Expression and localization of the sodium/iodide symporter (NIS) in testicular cells. Endocrine. 2011;40:35–40. doi: 10.1007/s12020-011-9469-y. [DOI] [PubMed] [Google Scholar]
- 149.Nicola J.P., Basquin C., Portulano C., Reyna-Neyra A., Paroder M., Carrasco N. The Na+/I− symporter mediates active iodide uptake in the intestine. Am. J. Physiol. Cell. Physiol. 2009;296:C654–C662. doi: 10.1152/ajpcell.00509.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Saito T., Endo T., Kawaguchi A., Ikeda M., Katoh R., Kawaoi A., Muramatsu A., Onaya T. Increased expression of the sodium/iodide symporter in papillary thyroid carcinomas. J. Clin. Investig. 1998;101:1296–1300. doi: 10.1172/JCI1259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Chatterjee S., Malhotra R., Varghese F., Bukhari A.B., Patil A., Budrukkar A., Parmar V., Gupta S., De A. Quantitative immunohistochemical analysis reveals association between sodium iodide symporter and estrogen receptor expression in breast cancer. PLoS One. 2013;8:e54055. doi: 10.1371/journal.pone.0054055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Dohan O., Baloch Z., Banrevi Z., Livolsi V., Carrasco N. Rapid communication: Predominant intracellular overexpression of the Na+/I− symporter (NIS) in a large sampling of thyroid cancer cases. J. Clin. Endocrinol. Metab. 2001;86:2697–2700. doi: 10.1210/jcem.86.6.7746. [DOI] [PubMed] [Google Scholar]
- 153.Luster M., Clarke S.E., Dietlein M., Lassmann M., Lind P., Oyen W.J., Tennvall J., Bombardieri E. Guidelines for radioiodine therapy of differentiated thyroid cancer. Eur. J. Nucl. Med. Mol. Imaging. 2008;35:1941–1959. doi: 10.1007/s00259-008-0883-1. [DOI] [PubMed] [Google Scholar]
- 154.Wapnir I.L., van de Rijn M., Nowels K., Amenta P.S., Walton K., Montgomery K., Greco R.S., Dohan O., Carrasco N. Immunohistochemical profile of the sodium/iodide symporter in thyroid, breast, and other carcinomas using high density tissue microarrays and conventional sections. J. Clin. Endocrinol. Metab. 2003;88:1880–1888. doi: 10.1210/jc.2002-021544. [DOI] [PubMed] [Google Scholar]
- 155.Chung J.K. Sodium iodide symporter: Its role in nuclear medicine. J. Nucl. Med. 2002;43:1188–1200. [PubMed] [Google Scholar]
- 156.Klein H.A., DiSibio K.J., Sims D., Singleton H.C., Worthy L.J. I-123 whole body scanning: Case report and discussion. Clin. Nucl. Med. 2005;30:312–316. doi: 10.1097/01.rlu.0000159526.99677.ed. [DOI] [PubMed] [Google Scholar]
- 157.Gerard S.K., Cavalieri R.R. I-123 diagnostic thyroid tumor whole-body scanning with imaging at 6, 24, and 48 hours. Clin. Nucl. Med. 2002;27:1–8. doi: 10.1097/00003072-200201000-00001. [DOI] [PubMed] [Google Scholar]
- 158.Mandel S.J., Shankar L.K., Benard F., Yamamoto A., Alavi A. Superiority of iodine-123 compared with iodine-131 scanning for thyroid remnants in patients with differentiated thyroid cancer. Clin. Nucl. Med. 2001;26:6–9. doi: 10.1097/00003072-200101000-00002. [DOI] [PubMed] [Google Scholar]
- 159.Cancroft E.T., Goldsmith S.J. 99mTc-pertechnetate scintigraphy as an aid in the diagnosis of breast masses. Radiology. 1973;106:441–444. doi: 10.1148/106.2.441. [DOI] [PubMed] [Google Scholar]
- 160.Joseph J.K., Patel R.B., Damle A.A., Nair N., Badwe R.A., Basu S. Functional radionuclide imaging, in-vitro radioiodine uptake estimation and RT-PCR in the evaluation of sodium iodide symporter (NIS) expression and functionality in breast cancer: A pilot study. Indian J. Surg. Oncol. 2013;4:80–91. doi: 10.1007/s13193-012-0205-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Pinto J.T., Suffoletto B.P., Berzin T.M., Qiao C.H., Lin S., Tong W.P., May F., Mukherjee B., Heston W.D. Prostate-specific membrane antigen: A novel folate hydrolase in human prostatic carcinoma cells. Clin. Cancer Res. 1996;2:1445–1451. [PubMed] [Google Scholar]
- 162.Silver D.A., Pellicer I., Fair W.R., Heston W.D., Cordon-Cardo C. Prostate-specific membrane antigen expression in normal and malignant human tissues. Clin. Cancer Res. 1997;3:81–85. [PubMed] [Google Scholar]
- 163.Wright G.L., Jr., Haley C., Beckett M.L., Schellhammer P.F. Expression of prostate-specific membrane antigen in normal, benign, and malignant prostate tissues. Urol. Oncol. 1995;1:18–28. doi: 10.1016/1078-1439(95)00002-Y. [DOI] [PubMed] [Google Scholar]
- 164.Mhawech-Fauceglia P., Zhang S., Terracciano L., Sauter G., Chadhuri A., Herrmann F.R., Penetrante R. Prostate-specific membrane antigen (PSMA) protein expression in normal and neoplastic tissues and its sensitivity and specificity in prostate adenocarcinoma: An immunohistochemical study using mutiple tumour tissue microarray technique. Histopathology. 2007;50:472–483. doi: 10.1111/j.1365-2559.2007.02635.x. [DOI] [PubMed] [Google Scholar]
- 165.Israeli R.S., Powell C.T., Corr J.G., Fair W.R., Heston W.D. Expression of the prostate-specific membrane antigen. Cancer Res. 1994;54:1807–1811. [PubMed] [Google Scholar]
- 166.Chang S.S., Reuter V.E., Heston W.D., Hutchinson B., Grauer L.S., Gaudin P.B. Short term neoadjuvant androgen deprivation therapy does not affect prostate specific membrane antigen expression in prostate tissues. Cancer. 2000;88:407–415. doi: 10.1002/(SICI)1097-0142(20000115)88:2<407::AID-CNCR23>3.0.CO;2-0. [DOI] [PubMed] [Google Scholar]
- 167.Wernicke A.G., Edgar M.A., Lavi E., Liu H., Salerno P., Bander N.H., Gutin P.H. Prostate-specific membrane antigen as a potential novel vascular target for treatment of glioblastoma multiforme. Arch. Pathol. Lab. Med. 2011;135:1486–1489. doi: 10.5858/arpa.2010-0740-OA. [DOI] [PubMed] [Google Scholar]
- 168.Haffner M.C., Kronberger I.E., Ross J.S., Sheehan C.E., Zitt M., Muhlmann G., Ofner D., Zelger B., Ensinger C., Yang X.J., et al. Prostate-specific membrane antigen expression in the neovasculature of gastric and colorectal cancers. Hum. Pathol. 2009;40:1754–1761. doi: 10.1016/j.humpath.2009.06.003. [DOI] [PubMed] [Google Scholar]
- 169.Rajasekaran A.K., Anilkumar G., Christiansen J.J. Is prostate-specific membrane antigen a multifunctional protein? Am. J. Physiol. Cell. Physiol. 2005;288:C975–C981. doi: 10.1152/ajpcell.00506.2004. [DOI] [PubMed] [Google Scholar]
- 170.Rajasekaran S.A., Anilkumar G., Oshima E., Bowie J.U., Liu H., Heston W., Bander N.H., Rajasekaran A.K. A novel cytoplasmic tail MXXXL motif mediates the internalization of prostate-specific membrane antigen. Mol. Biol. Cell. 2003;14:4835–4845. doi: 10.1091/mbc.E02-11-0731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Eder M., Eisenhut M., Babich J., Haberkorn U. PSMA as a target for radiolabelled small molecules. Eur. J. Nucl. Med. Mol. Imaging. 2013;40:819–823. doi: 10.1007/s00259-013-2374-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Murphy G.P., Elgamal A.A., Su S.L., Bostwick D.G., Holmes E.H. Current evaluation of the tissue localization and diagnostic utility of prostate specific membrane antigen. Cancer. 1998;83:2259–2269. doi: 10.1002/(SICI)1097-0142(19981201)83:11<2259::AID-CNCR5>3.0.CO;2-T. [DOI] [PubMed] [Google Scholar]
- 173.Troyer J.K., Beckett M.L., Wright G.L., Jr. Detection and characterization of the prostate-specific membrane antigen (PSMA) in tissue extracts and body fluids. Int. J. Cancer. 1995;62:552–558. doi: 10.1002/ijc.2910620511. [DOI] [PubMed] [Google Scholar]
- 174.Luthi-Carter R., Barczak A.K., Speno H., Coyle J.T. Molecular characterization of human brain N-acetylated α-linked acidic dipeptidase (NAALADase) J. Pharmacol. Exp. Ther. 1998;286:1020–1025. [PubMed] [Google Scholar]
- 175.Tiffany C.W., Lapidus R.G., Merion A., Calvin D.C., Slusher B.S. Characterization of the enzymatic activity of PSM: Comparison with brain NAALADase. Prostate. 1999;39:28–35. doi: 10.1002/(SICI)1097-0045(19990401)39:1<28::AID-PROS5>3.0.CO;2-A. [DOI] [PubMed] [Google Scholar]
- 176.Tang H., Brown M., Ye Y., Huang G., Zhang Y., Wang Y., Zhai H., Chen X., Shen T.Y., Tenniswood M. Prostate targeting ligands based on N-acetylated α-linked acidic dipeptidase. Biochem. Biophys. Res. Commun. 2003;307:8–14. doi: 10.1016/S0006-291X(03)01119-7. [DOI] [PubMed] [Google Scholar]
- 177.Oliver A.J., Wiest O., Helquist P., Miller M.J., Tenniswood M. Conformational and SAR analysis of NAALADase and PSMA inhibitors. Bioorg. Med. Chem. 2003;11:4455–4461. doi: 10.1016/S0968-0896(03)00427-9. [DOI] [PubMed] [Google Scholar]
- 178.Eder M., Neels O., Muller M., Bauder-Wust U., Remde Y., Schafer M., Hennrich U., Eisenhut M., Afshar-Oromieh A., Haberkorn U., et al. Novel preclinical and radiopharmaceutical aspects of [68Ga]Ga-PSMA-HBED-CC: A new PET tracer for imaging of prostate cancer. Pharmaceuticals (Basel) 2014;7:779–796. doi: 10.3390/ph7070779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Eder M., Wangler B., Knackmuss S., LeGall F., Little M., Haberkorn U., Mier W., Eisenhut M. Tetrafluorophenolate of HBED-CC: A versatile conjugation agent for 68Ga-labeled small recombinant antibodies. Eur. J. Nucl. Med. Mol. Imaging. 2008;35:1878–1886. doi: 10.1007/s00259-008-0816-z. [DOI] [PubMed] [Google Scholar]
- 180.Eder M., Schafer M., Bauder-Wust U., Hull W.E., Wangler C., Mier W., Haberkorn U., Eisenhut M. 68Ga-complex lipophilicity and the targeting property of a urea-based PSMA inhibitor for PET imaging. Bioconjug. Chem. 2012;23:688–697. doi: 10.1021/bc200279b. [DOI] [PubMed] [Google Scholar]
- 181.Flohr T. Messprinzip, Bildrekonstruktion, Gerätetyp und Aufnahmetechniken. In: Alkadhi H., editor. Wie funktioniert CT? Springer-Verlag Berlin Heidelberg; New York, NY, USA: 2011. (In German) [Google Scholar]
- 182.Zhang B.C., Hou L., Lv B., Xu Y.W. Post-marketing surveillance study with iodixanol in 20,185 Chinese patients from routine clinical practices. Br. J. Radiol. 2014;87:20130325. doi: 10.1259/bjr.20130325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Hasebroock K.M., Serkova N.J. Toxicity of MRI and CT contrast agents. Expert Opin. Drug Metab. Toxicol. 2009;5:403–416. doi: 10.1517/17425250902873796. [DOI] [PubMed] [Google Scholar]
- 184.Kuchmiy A.A., Efimov G.A., Nedospasov S.A. Methods for in vivo molecular imaging. Biochemistry (Moscow) 2012;77:1339–1353. doi: 10.1134/S0006297912120012. [DOI] [PubMed] [Google Scholar]
- 185.Histed S.N., Lindenberg M.L., Mena E., Turkbey B., Choyke P.L., Kurdziel K.A. Review of functional/anatomical imaging in oncology. Nucl. Med. Commun. 2012;33:349–361. doi: 10.1097/MNM.0b013e32834ec8a5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Lukaski H.C. Methods for the assessment of human body composition: Traditional and new. Am. J. Clin. Nutr. 1987;46:537–556. doi: 10.1093/ajcn/46.4.537. [DOI] [PubMed] [Google Scholar]
- 187.Hansen G., Crooks L.E., Davis P., de Groot J., Herfkens R., Margulis A.R., Gooding C., Kaufman L., Hoenninger J., Arakawa M., et al. In vivo imaging of the rat anatomy with nuclear magnetic resonance. Radiology. 1980;136:695–700. doi: 10.1148/radiology.136.3.7403549. [DOI] [PubMed] [Google Scholar]
- 188.Caravan P., Ellison J.J., McMurry T.J., Lauffer R.B. Gadolinium(III) chelates as MRI contrast agents: Structure, dynamics, and applications. Chem. Rev. 1999;99:2293–2352. doi: 10.1021/cr980440x. [DOI] [PubMed] [Google Scholar]
- 189.Lee S.H., Kim B.H., Na H.B., Hyeon T. Paramagnetic inorganic nanoparticles as T1 MRI contrast agents. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2014;6:196–209. doi: 10.1002/wnan.1243. [DOI] [PubMed] [Google Scholar]
- 190.Na H.B., Song I.C., Hyeon T. Inorganic nanoparticles for MRI contrast agents. Adv. Mate. 2009;21:2133–2148. doi: 10.1002/adma.200802366. [DOI] [Google Scholar]
- 191.Bar-Shalom R., Valdivia A.Y., Blaufox M.D. PET imaging in oncology. Semin. Nucl. Med. 2000;30:150–185. doi: 10.1053/snuc.2000.7439. [DOI] [PubMed] [Google Scholar]
- 192.Moses W.W. Recent advances and future advances in time-of-flight PET. Nucl. Instrum. Methods Phys. Res. A. 2007;580:919–924. doi: 10.1016/j.nima.2007.06.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Yankeelov T.E., Abramson R.G., Quarles C.C. Quantitative multimodality imaging in cancer research and therapy. Nat. Rev. Clin. Oncol. 2014;11:670–680. doi: 10.1038/nrclinonc.2014.134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Kauhanen S.P., Komar G., Seppanen M.P., Dean K.I., Minn H.R., Kajander S.A., Rinta-Kiikka I., Alanen K., Borra R.J., Puolakkainen P.A., et al. A prospective diagnostic accuracy study of 18F-fluorodeoxyglucose positron emission tomography/computed tomography, multidetector row computed tomography, and magnetic resonance imaging in primary diagnosis and staging of pancreatic cancer. Ann. Surg. 2009;250:957–963. doi: 10.1097/SLA.0b013e3181b2fafa. [DOI] [PubMed] [Google Scholar]
- 195.Rosenthal M.S., Cullom J., Hawkins W., Moore S.C., Tsui B.M., Yester M. Quantitative SPECT imaging: A review and recommendations by the Focus Committee of the Society of Nuclear Medicine Computer and Instrumentation Council. J. Nucl. Med. 1995;36:1489–1513. [PubMed] [Google Scholar]
- 196.Alirezapour B., Jalilian A.R., Bolourinovin F., Moradkhani S. Production and quality control of [(67)Ga]-DOTA-trastuzumab for radioimmunoscintigraphy. Iran. J. Pharm. Res. 2013;12:355–366. [PMC free article] [PubMed] [Google Scholar]
- 197.Pentlow K.S., Finn R.D., Larson S.M., Erdi Y.E., Beattie B.J., Humm J.L. Quantitative imaging of yttrium-86 with PET. The occurrence and correction of anomalous apparent activity in high density regions. Clin. Positron Imaging. 2000;3:85–90. doi: 10.1016/S1095-0397(00)00046-7. [DOI] [PubMed] [Google Scholar]
- 198.Forrer F., Valkema R., Kwekkeboom D.J., de Jong M., Krenning E.P. Neuroendocrine tumors. Peptide receptor radionuclide therapy. Best Pract. Res. Clin. Endocrinol. Metab. 2007;21:111–129. doi: 10.1016/j.beem.2007.01.007. [DOI] [PubMed] [Google Scholar]
- 199.Khalil M.M., Tremoleda J.L., Bayomy T.B., Gsell W. Molecular SPECT imaging: An overview. Int. J. Mol. Imaging. 2011;2011:796025. doi: 10.1155/2011/796025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Ohya S., Tamura T. Nuclear Data: I-123. [(accessed on 26 February 2015)]. Available online: http://crete.homeip.net/show_nuclide/530123/
- 201.NCHPS Nuclide Safety Data Sheet Iodine-131. [(accessed on 23 November 2014)]. Available online: http://hpschapters.org/northcarolina/NSDS/131IPDF.pdf.
- 202.McKillop J.H. Thallium 201 scintigraphy. West. J. Med. 1980;133:26–43. [PMC free article] [PubMed] [Google Scholar]