

Abstract
The visual system prioritizes information through a variety of mechanisms, including “attentional control settings” that specify features (e.g., colour) that are relevant to current goals. Recent work shows that these control settings may be more complex than previously thought, such that participants can monitor for independent features at different locations (Adamo, Pun, Pratt, & Ferber, 2008). However, this result leaves unclear whether these control settings affect early attentional selection or later target processing. We dissociated between these possibilities in two ways. In Experiment 1, participants were asked to determine whether a target object, which was preceded by an uninformative cue, matched one of two target templates (e.g., a blue vertical object or a green horizontal object). Participants monitored for independent features in the same location, but in different objects, which should reduce the effectiveness of the control setting if it is due to early attentional selection, but not if it is due to later target processing. In Experiment 2, we removed the ability of the cue to prime the target identity, which makes the opposite prediction. Together, the results suggest that complex attentional control settings primarily affect later target identity processing, and not early attentional selection.
Keywords: Attentional control settings, Working memory, Attentional capture, Priming
The complexity of the visual world requires us to prioritize processing of visual information that is most important for our current tasks and goals. One tool for achieving this prioritization is attentional “control settings” that determine what features, objects, or locations are selected. To study these control settings, one technique is to ask a participant to make a response based on a target object that appears with a certain feature (e.g., the red object). In order to quickly find the target, the observer has an incentive to form a “control setting” that selects that feature. To test the efficacy and precision of this control setting, a distracting cue with a different feature (e.g., green) is briefly shown just before the appearance of the target object, in the same or a different location. If the control setting fails to exclude this feature, then the cue will draw attention, impairing performance when it appears in a different location, and potentially helping performance when it appears in the same location. But if the control setting is precise enough to exclude the distractor, it should not affect performance (Folk, Remington, & Johnston, 1992).
There is substantial evidence that the visual system is capable of adopting such settings (Folk, Leber, & Egeth, 2002; Folk et al., 1992). In one of the original demonstrations, observers were able to ignore abruptly appearing dot cues (which typically robustly capture attention; Yantis & Jonides, 1984), when they were searching for certain colours, and were also able to ignore uniquely coloured objects when they were searching for abruptly appearing objects (Folk et al., 1992). These results have led to debate over the tasks and instructions that determine whether such selection is possible (e.g., Leber & Egeth, 2006; Theeuwes, Reimann, & Mortier, 2006) and conditions where such settings might arise without the intentions of the observer (Franconeri, Simons, & Junge, 2004; Gibson & Kelsey, 1998).
For attentional control settings for features such as colour, one potential underlying mechanism is that observers can bias competition for visual processing toward objects that contain a given feature (Desimone & Duncan, 1995). This feature-based mechanism appears to work globally across the visual field, biasing processing of features like colour and motion across spatially separated locations (Andersen, Hillyard, & Müller, 2008; Saenz, Buracas, & Boynton, 2002).
However, the results of a recent study challenge global-feature-based attention as a mechanism for colour-based attentional control settings, by showing that observers were able to select for different colours in different regions of space (Adamo, Pun, Pratt, & Ferber, 2008). Participants fixated the centre of a display containing one square on each side and monitored for different features at particular locations (e.g., “respond when a green square appears on the left, or a blue square appears on the right”). Before these objects appeared, an irrelevant cue (i.e., a blue or green border around either square) was presented 100 ms prior to the target. The cue could be compatible or incompatible with the target by its location (e.g., a cue on the same or different side as the subsequent target) or its colour (e.g., a blue cue, followed by a blue or green target). These two settings showed an interaction, where response times were lowest when both the location and the colour of the cue were compatible with designation of the target set.
If observers can form complex control settings that specify two different colours at two different locations, it would present a challenge to our understanding of how feature-based attention is applied globally across the entire visual field. The results of Adamo et al. (2008) are particularly surprising in light of recent evidence that it is difficult to select for two colours at once, even without location constraints (Folk & Anderson, 2010). But the results of Adamo et al. might also be rooted in later levels of processing. Specifically, the cues might not have attracted attention in a way that facilitated early visual processing, but instead could have primed a representation of a certain target due to its similar appearance. Given the implications of the Adamo et al. finding for our understanding of feature-based attention, it is important to examine the specific stage at which these more complex control settings operate.
To summarize, complex attentional control settings might influence processing on many levels, including early selection and target representation priming. The present study dissociates among these possibilities for the types of control settings used by Adamo et. al. (2008). Experiment 1 replicates the facilitation effect for a complex attentional setting involving different colours and locations but then shows that this effect is equally strong in a situation where the settings are unlikely to reflect tuning of early selection. Experiment 2 supports the idea that the cues prime the target representation, by showing that the cueing effect disappears when the identity of the cue is made nonpredictive of the target identity, despite sharing the same location as the target.
EXPERIMENT 1
Experiment 1 presents participants with two tasks involving complex attentional control settings. We define complex control settings as those that allow observers to simultaneously monitor for two independent targets in different locations, though broader definitions are also possible (e.g., any settings that require “conjunctive” combinations of multiple features, locations, or objects). The first task replicates the effect found in Adamo et al. (2008) of response time benefits for cues that match both the colour and the location of the future target object. One likely way to implement these separate attentional control settings would be to bias processing toward different colours within local spatial regions. If this were true, then placing the objects in the same local spatial region should make complex control settings more difficult to implement. Therefore, the second task introduces a new condition where the two potential target objects overlap in location. While the results of many cueing studies show that selection can act across whole visual objects (but see Shomstein & Yantis, 2002) there are also strong effects of location difference between cues and targets, even within the same object (Egly, Driver, & Rafal, 1994). If complex attentional control settings influence how regions of the display are selected, then using overlapping objects should reduce their effects.
Method
Participants
Twelve members of the Northwestern community (18–35 years old) completed Experiment 1 and were paid $10 or received course credit. Two additional participants showed less than 75% accuracy and were omitted from the analysis.
Stimuli
Stimuli were presented on a 17″ Viewsonic monitor driven by a MacMini computer running SR-Research Experiment Builder. The display subtended 32.6° × 24.4° at an approximate viewing distance of 56 cm (1,024 × 768 pixels, 3.6 pixels/degree). Two pairs of interwoven objects (centred 153 and 371 pixels from fixation) were placed on one side (blocked across participants) of a grey central fixation cross on a light grey background (see Figure 1). Object pairs were placed on the same side of fixation in order to control hemifield effects across conditions. Each pair consisted of two interwoven outline rectangles, each measuring 167 pixels in length and 180 pixels in width, one oriented vertically and one horizontally. Each rectangle has inner and outer borders that were dark grey and 5 pixels thick, separated by a 10-pixel thick internal fill area filled with the screen’s background colour. During cue periods the borders of one object would turn green or blue, and during target periods the internal fill area (between the borders) of one object would fill with green or blue. For the separated location condition, only one object from each pair was used (the horizontal object of the outer pair and the vertical object of the inner pair), allowing participants to select objects by their location. In the overlapping location condition, only the pair of objects closest to fixation was used.
Figure 1.
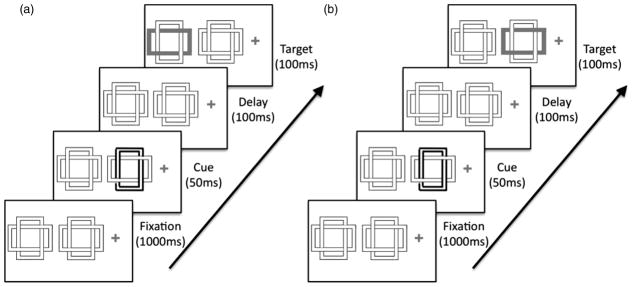
Sequence of events in (a) a separate-location trial and (b) an overlapping-location trial of Experiment 1. The compatibility between the colour and object of the cue varied relative to the target. Displays not drawn to scale.
At the start of each trial, the two pairs of objects appeared for 1,000 ms. Then, on 80% of trials one object outline (either a horizontal or a vertical object within a pair) was cued for 50 ms, followed by the removal of colour for 100 ms. On uncued trials, there was no colour for the entire 150 ms. The no-cue trials were included to emulate the design of Adamo et al. (2008) but because they did not contribute to the conclusions of Adamo et al. or the present experiments, the response times from these trials are not reported. Targets (i.e., a filled colour region) were then presented for 100 ms followed by the removal of colour. Object outlines remained onscreen until a participant indicated a response, or until 750 ms had elapsed from the presentation of the target. An intertrial interval (1,000 ms) followed each trial.
Procedure
Participants were instructed to press the space bar when one of two target colour/object combinations was presented (e.g., green-vertical or blue-horizontal object), and these combinations were counterbalanced across participants. The cue’s colour and object were unpredictive of the target. The combination of 2 cue colours, 2 target colours, 2 cue objects, and 2 target objects led to 16 possible trial types, plus 4 no-cue trials. This set of 20 trials was replicated 15 times to create 300 trials per condition (randomly ordered), with condition order counterbalanced across participants.
Participants were instructed to maintain fixation throughout the experiment, and the instructions emphasized accuracy. Space bar responses were made following half of the trials. Correct responses included a space bar press to target colour and object combinations consistent with the indicated instructions and the absence of a response for 750 ms to combinations inconsistent with those instructions. Incorrect responses resulted in an “incorrect” message for 8 s.
Results and discussion
Response time data are depicted in Figure 2. We conducted a 2 (object location: separated or overlapping) × 2 (object compatibility: match or mismatch) × 2 (colour compatibility: match or mismatch) repeated measures analysis of variance (ANOVA) with instruction type (target is blue-horizontal or green-vertical, vs. green-horizontal, blue-vertical) as a between-subjects factor. There were no main effects or interactions involving object location, suggesting no difference in performance when objects were spatially separated compared to when they overlapped in location. There were also no effects involving instruction type. Response times were faster when cue and target appeared on the same object (M = 559 ms) than when they appeared on different objects (M = 592 ms), F(1, 11) = 79.1, p < .001. Response times were also faster when the cue and target matched in colour (M = 554 ms) than when they did not match (M = 597 ms), F(1, 11) = 47.0, p < .001. Critically, there was a significant interaction between object and colour compatibility, F(1, 11) = 11.1, p = .007, driven by faster response times when the cue matched the target in both location and colour. A similar ANOVA using accuracy data demonstrated congruent results (see Table 1).
Figure 2.
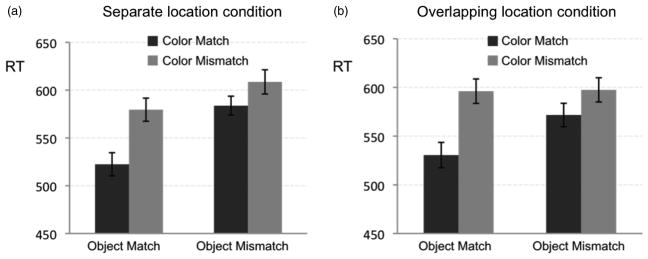
Mean response time in each of the conditions of Experiment 1, with error bars that represent the standard error of the mean for the (a) separate-location and (b) overlapping-location condition, respectively.
Table 1.
A summary of an ANOVA on accuracy in Experiment 1
| Object location | Compatibility | Accuracy (%)
|
F(1, 11) | p | |
|---|---|---|---|---|---|
| Match | Mismatch | ||||
| Separated | Object | 95.0 | 94.5 | 0.10 | .7 |
| Colour | 95.4 | 92.6 | 7.00 | .02 | |
| Object × Coloura | 3.90 | .07 | |||
| Overlapping | Object | 92.5 | 89.5 | 9.70 | .01 |
| Colour | 92.6 | 89.4 | 3.90 | .07 | |
| Object × Colourb | 0.04 | .85 | |||
Note: ANOVA = analysis of variance.
Lower accuracy for dual object/colour mismatch.
No interaction.
While there were main effects of colour and object compatibility on response time, there was also an interaction between the two factors, suggesting an additional advantage when the cue was fully consistent with the subsequent target. This effect was equally strong when the objects overlapped in location, making it less likely that the effect reflects tuning of early selection. In contrast, the strong effect for overlapping objects is consistent with postselection accounts such as target-representation priming. Experiment 2 tests this possibility directly by severing the relationship between the cue and the target identity, while requiring that participants maintain a complex attentional control setting.
EXPERIMENT 2
Experiment 2 maintains the same type of complex attentional control settings for targets and their related cues, but removes the perceptual similarity between the cues and targets so that the cue can no longer prime the target representation (see Folk et al., 1992, for a similar target–cue relationship). Cues were still briefly presented colour outlines, but targets were changed to a letter identification task for letters that match the target designation. Although participants now perform a discrimination task instead of a detection task, discrimination tasks have previously shown robust effects of simple attentional control settings (Folk et al., 1992).
Method
Participants
Seventeen members of the Northwestern community (18–35 years old) completed Experiment 2 and received course credit.
Stimuli
Displays were similar to the original Adamo et al. (2008) stimuli (see Figure 3). A fixation point was flanked by two light grey rectangular boxes (114 × 167 pixels, border stroke width of 5 pixels) on a dark grey background, centred 238 pixels to either side of a grey central fixation cross. To increase the participant’s incentive to select only the relevant target object and not all objects, each potential target position was surrounded by an arrangement of grey letters (Ts and Ls, font size 35); see Figure 3 for sample arrangement. As in Experiment 1, cues consisted of border colour changes. Targets consisted of a T or L presented in either blue or green, presented within one of the two boxes. All other aspects were similar to those in Experiment 1.
Figure 3.

Sequence of events in a trial of Experiment 2. The compatibility between the colour and object of the cue varied relative to the target, but the target identity was unrelated to the cue identity. Displays not drawn to scale.
Procedure
The timing of cues was identical to that in Experiment 1. Following the initial display and cue, the target (a coloured “T” or “L”) appeared inside one of the two objects until response, with a simultaneous presentation of an uncoloured letter inside the other object. Trials that did not require a response remained on the screen until a response or 2,000 ms had elapsed. Participants pressed the “T” or “L” key identifying, for example, the blue letter on the left or the green letter on the right. Other letter/colour combinations should lead to no response (50% of trials). There were a total of 600 trials.
Results and discussion
Response time data (Figure 4) were submitted to a 2 × 2 ANOVA with object and colour compatibility as factors. There was a main effect of object compatibility, F(1, 16) = 31.0, p < .001, but no effect of colour compatibility, F(1, 16) = 1.4, p = .25, and no interaction, F(1, 16) = 2.4, p = .14. Accuracy was between 96.3–97.7% in all conditions, and a similar ANOVA failed to reveal evidence of a speed–accuracy trade-off. When the ability of the cue to prime the target identity was eliminated, there was no longer an interaction that would suggest the existence of a complex attentional control setting for different colours on different objects or locations.
Figure 4.

Mean response times in each of the conditions of Experiment 2, with error bars that represent the standard error of the mean for each condition.
CONCLUSIONS
Recent findings show that attentional control settings can be surprisingly complex, allowing the selection of different colours at different locations (Adamo et al., 2008). These findings could be explained by facilitation at multiple levels of visual processing. These levels include early attentional selection for locations or objects, or target representation priming. Simple control settings for individual features or locations may affect all of these levels (Folk et al., 1992). But for more complex control settings, such as two different colours in different locations (Adamo et al., 2008), the present results suggest that this facilitation is limited to the level of the target representation or beyond, driven by the perceptual similarity of the cue and target. Experiment 1 demonstrated that the complex attentional set was equally powerful when participants monitored two objects in the same spatial location, which should have impaired a process that relied upon a location-based mechanism. Experiment 2 showed that when the ability of the cue to prime a target identity or associated response was severed, the effect of the complex control setting disappeared. Whereas the results of Experiment 1 ruled out a specific form of early attentional selection, the results of Experiment 2 suggest that, when applying complex control settings, cues provide a benefit by facilitating later processing of perceptually similar targets.
There are several possible postselection processes that might be influenced by cues that match complex attentional control settings for later targets. The cue might facilitate encoding or processing of the target information in visual working memory (Duncan, 1985). In Experiment 1, participants may have matched the colour and orientation of the cue (e.g., green horizontal) to the same characteristics of one of their potential target representations. Facilitation might also occur if there is a link between the target identity and the appropriate response (Eriksen & Schultz, 1979). While less likely in the present designs, there is also evidence that cueing effects can be related to decision processes at even later stages, such as an alteration in the criterion for responding to information at cued locations (Shiu & Pashler, 1994). Although the specific postselection process responsible for the effect observed in Experiment 1 cannot be isolated from the present results, together our experiments suggest that the role of an early-selection process is minimal.
A recent study used electrophysiology to begin to determine the processing stage influenced by the type of complex attentional set used here (Adamo, Pun, & Ferber, 2010). Using a paradigm similar to that of Adamo et al. (2008), participants saw a cue that was compatible with the colour and/or location of a subsequent target defined by a complex control setting. There was no selectivity for the complex control setting in early components thought to reflect early selection (n2pc), but there was selectivity in later components that should reflect later attentional processes and processing of targets within visual working memory. Converging evidence from a different paradigm also suggests that it is difficult for observers to simultaneously extract the shape of differently coloured regions from different locations in a display (Huang & Pashler, 2002). Our study provides converging behavioural evidence with both of these studies. While it is possible to maintain complex attentional control settings, they appear to influence processing stages beyond early attentional selection.
References
- Adamo M, Pun C, Ferber S. Multiple attentional control settings influence late attentional selection but do not provide an early attentional filter. Cognitive Neuroscience. 2010;1(2):102–110. doi: 10.1080/17588921003646149. [DOI] [PubMed] [Google Scholar]
- Adamo M, Pun C, Pratt J, Ferber S. Your divided attention, please! The maintenance of multiple attentional control sets over distinct regions in space. Cognition. 2008;107:295–303. doi: 10.1016/j.cognition.2007.07.003. [DOI] [PubMed] [Google Scholar]
- Andersen SK, Hillyard SA, Müller MM. Attention facilitates multiple stimulus features in parallel in human visual cortex. Current Biology. 2008;18:1006–1009. doi: 10.1016/j.cub.2008.06.030. [DOI] [PubMed] [Google Scholar]
- Desimone R, Duncan J. Neural mechanisms of selective visual attention. Annual Review of Neuroscience. 1995;18:193–222. doi: 10.1146/annurev.ne.18.030195.001205. [DOI] [PubMed] [Google Scholar]
- Duncan J. Visual search and visual attention. In: Posner MI, Marin OS, editors. Attention and Performance XI. Hillsdale, NJ: Lawrence Erlbaum Associates; 1985. pp. 85–106. [Google Scholar]
- Egly R, Driver J, Rafal RD. Shifting visual attention between objects and locations: Evidence from normal and parietal lesion subjects. Journal of Experimental Psychology: General. 1994;123:161–177. doi: 10.1037//0096-3445.123.2.161. [DOI] [PubMed] [Google Scholar]
- Eriksen CW, Schultz DW. Information processing in visual search: A continuous flow conception and experimental results. Perception and Psychophysics. 1979;25:249–263. doi: 10.3758/bf03198804. [DOI] [PubMed] [Google Scholar]
- Folk CL, Anderson BA. Target-uncertainty effects in attentional capture: Color singleton set or multiple attentional control settings? Psychonomic Bulletin and Review. 2010;17(3):421–426. doi: 10.3758/PBR.17.3.421. [DOI] [PubMed] [Google Scholar]
- Folk CL, Leber AB, Egeth HE. Made you blink! Contingent attentional capture produces a spatial blink. Perception and Psychophysics. 2002;64(5):741–753. doi: 10.3758/bf03194741. [DOI] [PubMed] [Google Scholar]
- Folk CL, Remington RW, Johnston JC. Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception and Performance. 1992;18:1030–1044. [PubMed] [Google Scholar]
- Franconeri SL, Simons DJ, Junge JA. Searching for stimulus-driven shifts of attention. Psychonomic Bulletin and Review. 2004;11(5):876–881. doi: 10.3758/bf03196715. [DOI] [PubMed] [Google Scholar]
- Gibson B, Kelsey E. Stimulus-driven attentional capture is contingent on attentional set for display-wide visual features. Journal of Experimental Psychology: Human Perception and Performance. 1998;24:699–706. doi: 10.1037//0096-1523.24.3.699. [DOI] [PubMed] [Google Scholar]
- Huang L, Pashler H. Symmetry detection and visual attention: A “binary-map” hypothesis. Vision Research. 2002;42:1421–1430. doi: 10.1016/s0042-6989(02)00059-7. [DOI] [PubMed] [Google Scholar]
- Leber AB, Egeth HE. It’s under control: Top-down search strategies can override attentional capture. Psychonomic Bulletin & Review. 2006;13:132–138. doi: 10.3758/bf03193824. [DOI] [PubMed] [Google Scholar]
- Saenz M, Buracas GT, Boynton GM. Global effects of feature-based attention in human visual cortex. Nature Neuroscience. 2002;5:631–632. doi: 10.1038/nn876. [DOI] [PubMed] [Google Scholar]
- Shiu L, Pashler H. Negligible effect of spatial precuing on identification of single digits. Journal of Experimental Psychology: Human Perception and Performance. 1994;20:1037–1054. [Google Scholar]
- Shomstein S, Yantis S. Object-based attention: Sensory modulation or priority setting? Perception & Psychophysics. 2002;64:41–51. doi: 10.3758/bf03194556. [DOI] [PubMed] [Google Scholar]
- Theeuwes J, Reimann B, Mortier K. Visual search for featural singletons: No top-down modulation, only bottom-up priming. Visual Cognition. 2006;14:466–489. [Google Scholar]
- Yantis S, Jonides J. Abrupt visual onsets and selective attention: Evidence from visual search. Journal of Experimental Psychology: Human Perception and Performance. 1984;10:601–621. doi: 10.1037//0096-1523.10.5.601. [DOI] [PubMed] [Google Scholar]