

Abstract
Pulmonary artery remodelling it is a major feature of pulmonary hypertension (PH). It is characterised by cellular and structural changes of the pulmonary arteries causing higher pulmonar vascular resistance and right ventricular failure. Abnormal deposition of smooth muscle-like (SM-like) cells in normally non-muscular, small diameter vessels and a deregulated control of endothelial cells are considered pathological features of PH. The origin of the SM-like cells and the mechanisms underlying the development and progression of this remodelling process are not understood. Endothelial cells within the intima may migrate from their organised layer of cells and transition to mesenchymal or SM-like phenotype in a process called endothelial-mesenchymal transition (EnMT). Traditionally, Waddington’s epigenetic landscape illustrates that fates of somatic cells are progressively determined to compulsorily follow a downhill differentiation pathway. EnMT induces the transformation of cells with stem cell traits, therefore contrasting Waddington’s theory and confirming that cell fate seems to be far more flexible than previously thought. The prospect of therapeutic inhibition of EnMT to delay or prevent PH may represent a promising new treatment modality.
Keywords: Endothelial to mesenchymal transition, remodelling, endothelial cells, pulmonary hypertension, cellular reprogramming
Pulmonary hypertension
Pulmonary hypertension (PH) is a complex and progressive disease characterised by increased blood pressure in pulmonary arteries. Hemodynamically, it is defined by a mean pulmonary artery pressure at rest exceeding 25 mmHg [1]. Pulmonary vessel remodelling, which consists of intimal, medial and adventitial hypertrophy, leads to a reduction of the vascular lumen [2]. This causes an increase in pulmonary resistance, severe PH, right ventricular failure, and early death [3]. Currently, there is no cure for PH, but available treatments can lessen symptoms and improve quality of life. PH can occur due to genetic or sporadic causes, but the exact basis of PH remain unknown [4,5].
Pulmonary arterial remodelling
Pulmonary artery remodelling it is a major feature of PH. It is characterised by cellular and structural changes affecting all three layers of the vessel wall of the pulmonary arteries [6,7]. Common pulmonary vascular remodelling changes in PH include increased intimal and/or medial stiffening and thickening, elevated expression of smooth muscle α-actin, collagen synthesis/deposition, and inflammation [8]. Abnormal deposition of smooth muscle-like (SM-like) cells in normally non-muscular, small diameter vessels and a deregulated control of endothelial cells are considered important pathological features of PH [9]. Also, increased production of extracellular matrix proteins, with deposition of collagen and elastin contribute to lumen narrowing and PH [10]. It is this remodelling process inside the pulmonary vessels that is responsible for elevation of pulmonary vascular resistance, progressive PH, right ventricular failure and finally death [11].
The origin of the SM-like cells and the mechanisms underlying the development and progression of this remodelling process are not completely understood. It is been thought that muscularisation of the intimal layer of the vessel wall is caused by proliferation of resident vascular smooth muscle cells of the medial layer which migrate to the intima [12,13] (Figure 1A). Smooth muscle cells of the medial layer retain high cell plasticity and, under specific circumstances, can undergo phenotypic switch towards a synthetic or “de-differentiated” state. De-differentiated SMCs express high levels of extracellular matrix components and reduced expression of SMC contractile proteins. In contrast to differentiated SMCs, de-differentiated SMCs become highly proliferative and migratory [14,15]. Recently, other possible sources of SM-like cells in the intimal layer of pulmonary vessels have been postulated. Circulating progenitor cells have been shown to be recruited to sites of vascular injury and assume a SM-like phenotype [16-19] (Figure 1B). Resident progenitor cells present in the adventitia have also been postulated to be involved in vascular remodelling [20] (Figure 1C). Additionally, resident endothelial cells within the intima may delaminate from their organised layer of cells in the vessel lining, transition to mesenchymal or SM-like phenotype in a process called endothelial-mesenchymal transition (EnMT) and migrate to their underlying tissue [13,21] (Figure 1D). In vitro, these altered cells have an indistinguishable morphology from de-differentiated SMCs and express abundant extracellular matrix proteins. In this review we will go over the evidences for EnMT of vascular endothelial cells and its potential implications in PH.
Figure 1.
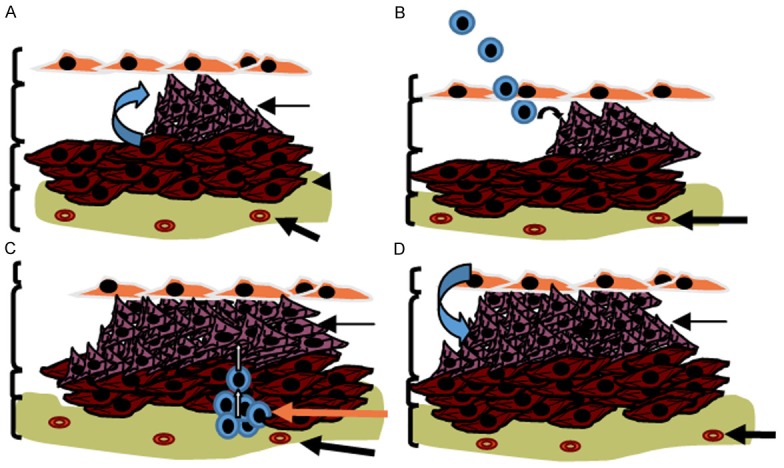
The origin of the SM-like cells in pulmonary arterial remodelling. Resident vascular smooth muscle cells (SMCs) from the medial layer retain high cell plasticity and, under specific circumstances, undergo phenotypic switch towards a synthetic or “de-differentiated” state (A). Differentiated SMCs become highly proliferative and migratory. Circulating progenitor cells could be recruited to sites of vascular injury and assume a SM-like phenotype (B). Resident progenitor cells in the adventitia may also serve as a source of SM-like cells and contribute to the pathophysiological changes in vascular structure (C). Additionally, Endothelial cells within the intima may migrate from their organised layer of cells and transition to mesenchymal or SM-like phenotype in a process called endothelial-mesenchymal transition (EnMT) (D).
Endothelial-mesenchymal transition (EnMT)
Endothelial cells may contribute to vascular remodelling through EnMT. This is a process of endothelial cell “transformation” into mesenchymal cells, by which endothelial cells lose their endothelial characteristics and gain a spindle shaped mesenchymal-like phenotype [22,23]. EnMT is a central process during embryonic development [24-26]. In heart development, endocardial cells with a clearly endothelial phenotype are able to give rise to mesenchymal heart cushion cells through a process of EnMT [27-29]. Arciniegas et al, have also shown that EnMT is an important event in aortic and pulmonary artery development [30] and in the development of the normal arterial intima [31]. Moreover, morphological studies in human embryos suggest that EnMT also occurs during the maturation of both arteries and veins [32].
Recently, some studies have shown that EnMT could also happen in adult life in a variety of pathologic settings, including PH [33,34], atherosclerosis [35] and wound healing [36]. In 1997, Romero et al showed that capillary endothelial cells could undergo mesenchymal transition in response to chronic inflammatory stimuli [37]. Additionally, a number of in vitro studies have demonstrated that endothelial cells from a variety of vascular beds retain the ability to transition into mesenchymal or SM-like cells under specific culture conditions [38,42].
A loss of cell-to-cell contact seems to be a triggering step in the development of EnMT [43]. Cell-cell adhesion glycoprotein, VE-cadherin, plays an important role in vascular biology by controlling the cohesion and organization of cell-cell junctions. The loss of expression of VE-cadherin, consistently preceded endothelial phenotype downregulation and SM-like transformation [44].
Signalling during EnMT
There are several key signalling pathways contributing to the remodelling process and to date, a number of studies have demonstrated the induction of EnMT in vitro. It has been suggested that the TGF-β1 signalling pathway is involved in EnMT [45]. Both mouse and human endothelial cells cultured in the presence of TGF-β1 have shown SM-like cell morphology and an up-regulation of α-SM actin [46-48]. Recently, studies have shown that both TGF-β1 and Wnt-signalling pathways could synergize in the EnMT process [49,50]. Moreover, in addition to TGF-β1 and Wnt-signalling pathways, the Notch pathway has also been shown to be involved in both vascular development and intimal lesion formation processes [51-53]. Although it is known that all these signalling pathways contribute to EnMT, it is not clear whether Notch, Wnt, TGF-β1, or the combination of all pathways, provide the initiation signal for EnMT in vivo [54]. Furthermore, it is likely that other signalling pathways such as VEGF, NFAT and BMP, which interact with TGF-β1 and Notch signalling, also mediate EnMT [55].
Despite these signalling studies, the transcriptional networks which mediate EnMT remain largely unidentified. Recently, the role of the Snail family of transcriptional repressors has been highlighted in control of different transcriptional programs of mesenchymal stem cell differentiation [56]. Snail proteins including Slug are involved in a broad spectrum of biological functions including epithelial to mesenchymal transition (EMT), cell differentiation, cell motility, cell-cycle regulation and apoptosis. In the context of EnMT, Snail and Slug play a critical role in disrupting cell-cell junctions and down-regulating VE-cadherin gene expression [57-59].
EMT/EnMT
EnMT is related to the more generally recognized mechanism of somatic cell plasticity; epithelial-to-mesenchymal transition (EMT) [60]. EMT involves a phenotypic cellular switch, in this case, from an epithelial to mesenchymal phenotype [61]. EnMT, EMT and its reverse pathway, mesenchymal to epithelial transition (MET) are key biological processes that occur naturally during embryonic life [62,63]. Cellular switching from an epithelial to mesenchymal phenotype (EMT), and conversely from a mesenchymal to epithelial phenotype (MET) are important biologic programs fundamental to the generation of several complex body patterns throughout evolution [64,65]. EMT plays a central role in germ layer specification (endoderm, ectoderm and mesoderm) and a range of different recurrent EMT/MET cycles occur before final organ formation [66]. In lungs for example, EMT is a natural process that exists to allow airway branching during fetal development. During cardiac development, EMT/MET is crucial in valve formation and heart septation [67]. In other organs, such as the kidney, successive phases of EMT/MET are necessary to ultimately give rise to nephrons or nephric ducts [68]. Moreover in the adult, EMT contributes to pathology of tissue fibrosis, tumour progression, and tumour metastasis, which results when cells delaminate from the primary tumour, allows them to migrate [69,70].
Cellular reprogramming
Traditionally, Waddington’s epigenetic landscape illustrates that fates of somatic cells are progressively determined to compulsorily follow a downhill differentiation pathway [71] (Figure 2A). As such any reversal of cell differentiation status would require external intervention in nuclear function such as nuclear transfer [72,73] or the introduction of several transcription factors [74]. The plasticity of somatic cell fate is a complex concept that has evolved through decades of research. It is currently known that some somatic cells seem to possess a greater plasticity when exposed to certain stimuli. Smith et al, showed in 1998 that Oct4 transcription factor is required for maintaining mouse embryonic stem cell pluripotency [75], and in 2006 Takahashi and Yamanaka et al, were able to reprogram mouse fibroblasts to induced pluripotent stem cells (iPS) using 4 transcription factors (Sox2, Klf4, Oct4 and c-Myc) [76]. EMT and EnMT are examples of remarkable somatic cell plasticity occurring naturally though organogenesis. There is current evidence that cells undergoing EMT/EnMT acquire stem cell properties [77,78] and it has recently been postulated that MET is a key cellular mechanism required for transforming somatic cells toward the generation of induced pluripotent stem cells [79]. Li et al, showed that while Sox2/Oct4 upregulation suppresses the EMT mediator Snail, c-Myc downregulates TGF-β1 and TGF-β receptor 2, whilst, Klf4 induces the expression of epithelial genes including E-cadherin [80]. These recent findings are important and raise many questions. They oppose Waddington’s epigenetic view in which a fully differentiated cell had completed a downhill journey into a valley from which it could not then escape. Cell fate seems now to be far more flexible than previously thought [81].
Figure 2.
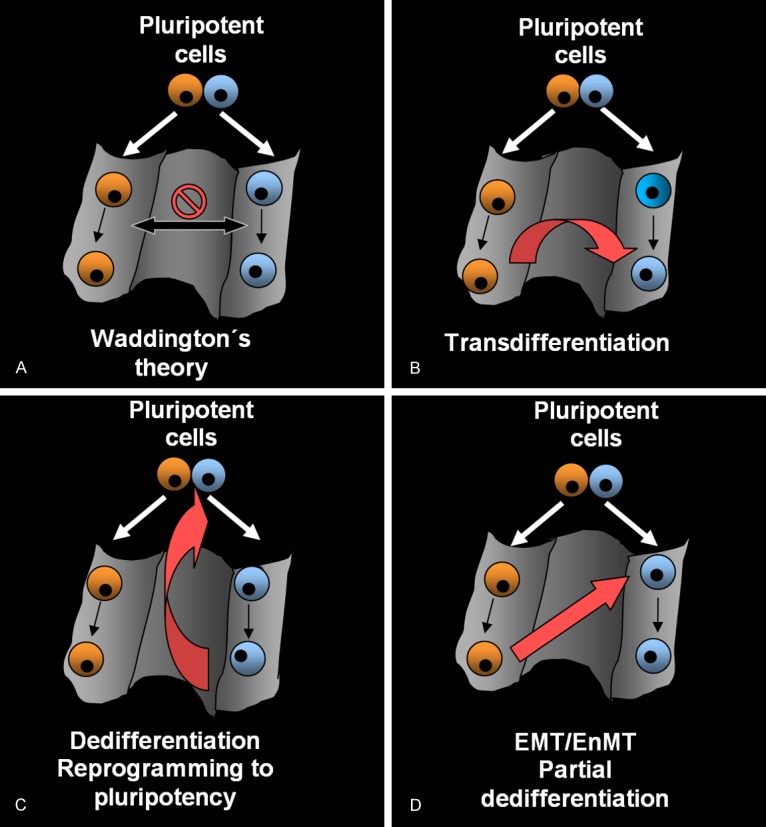
Cellular differentiation and reprogramming patterns. Waddington’s epigenetic landscape. Somatic cells take on a specific fate by compulsorily progressing from the pluripotent state to a terminal differentiated state (A). Transdifferentiation. A mature cell switches its phenotype and function to that of another mature differentiated cell type without undergoing an intermediate pluripotent state or becoming a progenitor cell (B). Dedifferentiation of reprogramming to pluripotency. Differentiated cells return to an immature state and regain pluripotency (C). EMT/EnMT; partial dedifferentiation. Cells change their specific fate, dedifferentiate and acquire a more immature, proliferative phenotype from another somatic lineage without converting to a pluripotent cell (D).
Transdifferentiation refers to a process where one mature cell switches its phenotype and function to that of another mature differentiated cell type without undergoing an intermediate pluripotent state or becoming a progenitor cell [82] (Figure 2B). Davis et al, showed in 1987, that mouse embryonic fibroblasts could transdifferentiate directly into mature myoblasts [83]. EMT/EnMT induces the generation of cells with stem cell traits, thus it does not fulfil Takahashi’s definition of transdifferentiation. Dedifferentiation is a process which induces cell rejuvenation. It refers to a process where cells travel back up their differentiation path, to become more immature and finally convert into a pluripotent cell [84] (Figure 2C). EMT/EnMT could be considered an initial stage of cellular dedifferentiation or reprogramming processes where cells dedifferentiate and acquire a more immature, proliferative phenotype but do not convert to a pluripotent cell. Cellular reprogramming and change of cell fate decisions through EMT/EnMT provides new ways to traverse across Waddington’s epigenetic landscape (Figure 2D). As EMT/EnMT are essential and occurs frequently during embryonic development one could postulate that cellular reprogramming occurs spontaneously in the embryo and in certain adult pathological conditions.
Reversibility of EMT/EnMT. Future perspectives
During embryogenesis, EMT and its reverse process, MET, occur spontaneously during organ morphogenesis [85]. However, less is known about the reversibility of EnMT [86]. It has been suggested that endothelial cells transformed to a more SM-like cell phenotype could be restored by administration of FGF [87]. Recently our group have shown that in vitro, following withdrawal of TNF-α, SM-like cells acquired a normal phenotype (manuscript submitted). However, more studies are necessary to determine the reversibility of this EnMT process in vitro and in vivo.
It is accepted that EMT/EnMT occurs not only during embryonic development but also in the pathogenesis of various cardiovascular diseases such as heart failure and PH. The prospect of therapeutic manipulation of EnMT/EMT in the treatment of these conditions is particularly attractive [88,89]. In particular, therapies directed at inhibiting EnMT to delay or prevent PH, may represent a promising new treatment modality. Nevertheless, additional studies are needed to identify the precise molecular mechanism of EnMT in disease, to provide novel insights into the mechanisms of such diseases and to determine which signalling components might be viable therapeutic targets.
Acknowledgements
The authors would like to thank K. Samuel for her always faithful advice.
References
- 1.McLaughlin VV, McGoon MD. Pulmonary arterial hypertension. Circulation. 2006;114:1417–31. doi: 10.1161/CIRCULATIONAHA.104.503540. [DOI] [PubMed] [Google Scholar]
- 2.Hopkins N, Mcloughlin P. The structural basis of pulmonary hypertension in chronic lung disease: remodelling, rarefaction or angiogenesis? J Anat. 2002;201:335–48. doi: 10.1046/j.1469-7580.2002.00096.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Eddahibi S, Morrell N, d’Ortho MP, Naeije R, Adnot S. Pathobiology of pulmonary arterial hypertension. Eur Respir J. 2002;20:1559–72. doi: 10.1183/09031936.02.00081302. [DOI] [PubMed] [Google Scholar]
- 4.Rubin LJ. Primary pulmonary hypertension. N Engl J Med. 1997;336:111–7. doi: 10.1056/NEJM199701093360207. [DOI] [PubMed] [Google Scholar]
- 5.Shimoda LA, Laurie SS. Vascular remodeling in pulmonary hypertension. J Mol Med (Berl) 2013;91:297–309. doi: 10.1007/s00109-013-0998-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Arciniegas E, Frid MG, Douglas IS, Stenmark KR. Perspectives on endothelial-to-mesenchymal transition: potential contribution to vascular remodeling in chronic pulmonary hypertension. Am J Physiol Lung Cell Mol Physiol. 2007;293:L1–L8. doi: 10.1152/ajplung.00378.2006. [DOI] [PubMed] [Google Scholar]
- 7.Hassoun PM. Deciphering the “matrix” in pulmonary vascular remodelling. Eur Respir J. 2005;25:778–9. doi: 10.1183/09031936.05.00027305. [DOI] [PubMed] [Google Scholar]
- 8.Arciniegas E, Frid MG, Douglas IS, Stenmark KR. Perspectives on endothelial-to-mesenchymal transition: potential contribution to vascular remodeling in chronic pulmonary hypertension. Am J Physiol Lung Cell Mol Physiol. 2007;293:L1–L8. doi: 10.1152/ajplung.00378.2006. [DOI] [PubMed] [Google Scholar]
- 9.Yi ES, Kim H, Ahn H, Strother J, Morris T, Masliah E, Hansen LA, Park K, Friedman PJ. Distribution of obstructive intimal lesions and their cellular phenotypes in chronic pulmonary hypertension. A morphometric and immunohistochemical study. Am J Respir Crit Care Med. 2000;162:1577–86. doi: 10.1164/ajrccm.162.4.9912131. [DOI] [PubMed] [Google Scholar]
- 10.Hassoun PM. Deciphering the “matrix” in pulmonary vascular remodelling. Eur Respir J. 2005;25:778–9. doi: 10.1183/09031936.05.00027305. [DOI] [PubMed] [Google Scholar]
- 11.Jungebluth P, Macchiarini P. Stem cell-based therapy and regenerative approaches to diseases of the respiratory system. Br Med Bull. 2011;99:169–87. doi: 10.1093/bmb/ldr028. [DOI] [PubMed] [Google Scholar]
- 12.Arciniegas E, Frid MG, Douglas IS, Stenmark KR. Perspectives on endothelial-to-mesenchymal transition: potential contribution to vascular remodeling in chronic pulmonary hypertension. Am J Physiol Lung Cell Mol Physiol. 2007;293:L1–L8. doi: 10.1152/ajplung.00378.2006. [DOI] [PubMed] [Google Scholar]
- 13.Fleenor BS, Marshall KD, Rippe C, Seals DR. Replicative aging induces endothelial to mesenchymal transition in human aortic endothelial cells: potential role of inflammation. J Vasc Res. 2012;49:59–64. doi: 10.1159/000329681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Owens GK. Regulation of differentiation of vascular smooth muscle cells. Physiol Rev. 1995;75:487–517. doi: 10.1152/physrev.1995.75.3.487. [DOI] [PubMed] [Google Scholar]
- 15.Owens GK, Kumar MS, Wamhoff BR. Molecular regulation of vascular smooth muscle cell differentiation in development and disease. Physiol Rev. 2004;84:767–801. doi: 10.1152/physrev.00041.2003. [DOI] [PubMed] [Google Scholar]
- 16.Diez M, Musri MM, Ferrer E, Barbera JA, Peinado VI. Endothelial progenitor cells undergo an endothelial-to-mesenchymal transition-like process mediated by TGFbetaRI. Cardiovasc Res. 2010;88:502–11. doi: 10.1093/cvr/cvq236. [DOI] [PubMed] [Google Scholar]
- 17.Frid MG, Brunetti JA, Burke DL, Carpenter TC, Davie NJ, Reeves JT, Roedersheimer MT, van RN, Stenmark KR. Hypoxia-induced pulmonary vascular remodeling requires recruitment of circulating mesenchymal precursors of a monocyte/macrophage lineage. Am J Pathol. 2006;168:659–69. doi: 10.2353/ajpath.2006.050599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Liu Y, Sinha S, McDonald OG, Shang Y, Hoofnagle MH, Owens GK. Kruppel-like factor 4 abrogates myocardin-induced activation of smooth muscle gene expression. J Biol Chem. 2005;280:9719–27. doi: 10.1074/jbc.M412862200. [DOI] [PubMed] [Google Scholar]
- 19.Sata M. Role of circulating vascular progenitors in angiogenesis, vascular healing, and pulmonary hypertension: lessons from animal models. Arterioscler Thromb Vasc Biol. 2006;26:1008–14. doi: 10.1161/01.ATV.0000206123.94140.f3. [DOI] [PubMed] [Google Scholar]
- 20.Stenmark KR, Davie N, Frid M, Gerasimovskaya E, Das M. Role of the adventitia in pulmonary vascular remodeling. Physiology (Bethesda) 2006;21:134–45. doi: 10.1152/physiol.00053.2005. [DOI] [PubMed] [Google Scholar]
- 21.Arciniegas E, Frid MG, Douglas IS, Stenmark KR. Perspectives on endothelial-to-mesenchymal transition: potential contribution to vascular remodeling in chronic pulmonary hypertension. Am J Physiol Lung Cell Mol Physiol. 2007;293:L1–L8. doi: 10.1152/ajplung.00378.2006. [DOI] [PubMed] [Google Scholar]
- 22.Arciniegas E, Frid MG, Douglas IS, Stenmark KR. Perspectives on endothelial-to-mesenchymal transition: potential contribution to vascular remodeling in chronic pulmonary hypertension. Am J Physiol Lung Cell Mol Physiol. 2007;293:L1–L8. doi: 10.1152/ajplung.00378.2006. [DOI] [PubMed] [Google Scholar]
- 23.Piera-Velazquez S, Jimenez SA. Molecular mechanisms of endothelial to mesenchymal cell transition (EndoMT) in experimentally induced fibrotic diseases. Fibrogenesis Tissue Repair. 2012;5(Suppl 1 Proceedings of Fibroproliferative disorders: from biochemical analysis to targeted therapiesPetro E Petrides and David Brenner):S7. doi: 10.1186/1755-1536-5-S1-S7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Arciniegas E, Frid MG, Douglas IS, Stenmark KR. Perspectives on endothelial-to-mesenchymal transition: potential contribution to vascular remodeling in chronic pulmonary hypertension. Am J Physiol Lung Cell Mol Physiol. 2007;293:L1–L8. doi: 10.1152/ajplung.00378.2006. [DOI] [PubMed] [Google Scholar]
- 25.Kovacic JC, Mercader N, Torres M, Boehm M, Fuster V. Epithelial-to-mesenchymal and endothelial- to-mesenchymal transition: from cardiovascular development to disease. Circulation. 2012;125:1795–808. doi: 10.1161/CIRCULATIONAHA.111.040352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Markwald RR, Fitzharris TP, Smith WN. Sturctural analysis of endocardial cytodifferentiation. Dev Biol. 1975;42:160–80. doi: 10.1016/0012-1606(75)90321-8. [DOI] [PubMed] [Google Scholar]
- 27.Armstrong EJ, Bischoff J. Heart valve development: endothelial cell signaling and differentiation. Circ Res. 2004;95:459–70. doi: 10.1161/01.RES.0000141146.95728.da. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Eisenberg LM, Markwald RR. Molecular regulation of atrioventricular valvuloseptal morphogenesis. Circ Res. 1995;77:1–6. doi: 10.1161/01.res.77.1.1. [DOI] [PubMed] [Google Scholar]
- 29.Liebner S, Cattelino A, Gallini R, Rudini N, Iurlaro M, Piccolo S, Dejana E. Beta-catenin is required for endothelial-mesenchymal transformation during heart cushion development in the mouse. J Cell Biol. 2004;166:359–67. doi: 10.1083/jcb.200403050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Arciniegas E, Neves CY, Carrillo LM, Zambrano EA, Ramirez R. Endothelial-mesenchymal transition occurs during embryonic pulmonary artery development. Endothelium. 2005;12:193–200. doi: 10.1080/10623320500227283. [DOI] [PubMed] [Google Scholar]
- 31.Arciniegas E, Neves CY, Carrillo LM, Zambrano EA, Ramirez R. Endothelial-mesenchymal transition occurs during embryonic pulmonary artery development. Endothelium. 2005;12:193–200. doi: 10.1080/10623320500227283. [DOI] [PubMed] [Google Scholar]
- 32.Hall SM, Hislop AA, Haworth SG. Origin, differentiation, and maturation of human pulmonary veins. Am J Respir Cell Mol Biol. 2002;26:333–40. doi: 10.1165/ajrcmb.26.3.4698. [DOI] [PubMed] [Google Scholar]
- 33.Arciniegas E, Frid MG, Douglas IS, Stenmark KR. Perspectives on endothelial-to-mesenchymal transition: potential contribution to vascular remodeling in chronic pulmonary hypertension. Am J Physiol Lung Cell Mol Physiol. 2007;293:L1–L8. doi: 10.1152/ajplung.00378.2006. [DOI] [PubMed] [Google Scholar]
- 34.Zhu P, Huang L, Ge X, Yan F, Wu R, Ao Q. Transdifferentiation of pulmonary arteriolar endothelial cells into smooth muscle-like cells regulated by myocardin involved in hypoxia-induced pulmonary vascular remodelling. Int J Exp Pathol. 2006;87:463–74. doi: 10.1111/j.1365-2613.2006.00503.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Mironov AA, Rekhter MD, Kolpakov VA, Andreeva ER, Polishchuk RS, Bannykh SI, Filippov SV, Peretjatko LP, Kulida LV, Orekhov AN. Heterogeneity of smooth muscle cells in embryonic human aorta. Tissue Cell. 1995;27:31–8. doi: 10.1016/s0040-8166(95)80006-9. [DOI] [PubMed] [Google Scholar]
- 36.Lee JG, Kay EP. FGF-2-induced wound healing in corneal endothelial cells requires Cdc42 activation and Rho inactivation through the phosphatidylinositol 3-kinase pathway. Invest Ophthalmol Vis Sci. 2006;47:1376–86. doi: 10.1167/iovs.05-1223. [DOI] [PubMed] [Google Scholar]
- 37.Romero LI, Zhang DN, Herron GS, Karasek MA. Interleukin-1 induces major phenotypic changes in human skin microvascular endothelial cells. J Cell Physiol. 1997;173:84–92. doi: 10.1002/(SICI)1097-4652(199710)173:1<84::AID-JCP10>3.0.CO;2-N. [DOI] [PubMed] [Google Scholar]
- 38.Arciniegas E, Sutton AB, Allen TD, Schor AM. Transforming growth factor beta 1 promotes the differentiation of endothelial cells into smooth muscle-like cells in vitro. J Cell Sci. 1992;103:521–9. doi: 10.1242/jcs.103.2.521. [DOI] [PubMed] [Google Scholar]
- 39.Cooley BC, Nevado J, Mellad J, Yang D, St HC, Negro A, Fang F, Chen G, San H, Walts AD, Schwartzbeck RL, Taylor B, Lanzer JD, Wragg A, Elagha A, Beltran LE, Berry C, Feil R, Virmani R, Ladich E, Kovacic JC, Boehm M. TGF-beta signaling mediates endothelial-to-mesenchymal transition (EndMT) during vein graft remodeling. Sci Transl Med. 2014;6:227ra34. doi: 10.1126/scitranslmed.3006927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Frid MG, Kale VA, Stenmark KR. Mature vascular endothelium can give rise to smooth muscle cells via endothelial-mesenchymal transdifferentiation: in vitro analysis. Circ Res. 2002;90:1189–96. doi: 10.1161/01.res.0000021432.70309.28. [DOI] [PubMed] [Google Scholar]
- 41.Ishisaki A, Hayashi H, Li AJ, Imamura T. Human umbilical vein endothelium-derived cells retain potential to differentiate into smooth muscle-like cells. J Biol Chem. 2003;278:1303–9. doi: 10.1074/jbc.M207329200. [DOI] [PubMed] [Google Scholar]
- 42.Qiao L, Nishimura T, Shi L, Sessions D, Thrasher A, Trudell JR, Berry GJ, Pearl RG, Kao PN. Endothelial fate mapping in mice with pulmonary hypertension. Circulation. 2014;129:692–703. doi: 10.1161/CIRCULATIONAHA.113.003734. [DOI] [PubMed] [Google Scholar]
- 43.Cavarretta E, Latronico MV, Condorelli G. Endothelial-to-mesenchymal transition and microRNA-21: the game is on again. Arterioscler Thromb Vasc Biol. 2012;32:165–6. doi: 10.1161/ATVBAHA.111.242008. [DOI] [PubMed] [Google Scholar]
- 44.Cavarretta E, Latronico MV, Condorelli G. Endothelial-to-mesenchymal transition and microRNA-21: the game is on again. Arterioscler Thromb Vasc Biol. 2012;32:165–6. doi: 10.1161/ATVBAHA.111.242008. [DOI] [PubMed] [Google Scholar]
- 45.Arciniegas E, Sutton AB, Allen TD, Schor AM. Transforming growth factor beta 1 promotes the differentiation of endothelial cells into smooth muscle-like cells in vitro. J Cell Sci. 1992;103:521–9. doi: 10.1242/jcs.103.2.521. [DOI] [PubMed] [Google Scholar]
- 46.Diez M, Musri MM, Ferrer E, Barbera JA, Peinado VI. Endothelial progenitor cells undergo an endothelial-to-mesenchymal transition-like process mediated by TGFbetaRI. Cardiovasc Res. 2010;88:502–11. doi: 10.1093/cvr/cvq236. [DOI] [PubMed] [Google Scholar]
- 47.van Meeteren LA, ten DP. Regulation of endothelial cell plasticity by TGF-beta. Cell Tissue Res. 2012;347:177–86. doi: 10.1007/s00441-011-1222-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Wang H, Radjendirane V, Wary KK, Chakrabarty S. Transforming growth factor beta regulates cell-cell adhesion through extracellular matrix remodeling and activation of focal adhesion kinase in human colon carcinoma Moser cells. Oncogene. 2004;23:5558–61. doi: 10.1038/sj.onc.1207701. [DOI] [PubMed] [Google Scholar]
- 49.Aisagbonhi O, Rai M, Ryzhov S, Atria N, Feoktistov I, Hatzopoulos AK. Experimental myocardial infarction triggers canonical Wnt signaling and endothelial-to-mesenchymal transition. Dis Model Mech. 2011;4:469–83. doi: 10.1242/dmm.006510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Liebner S, Cattelino A, Gallini R, Rudini N, Iurlaro M, Piccolo S, Dejana E. Beta-catenin is required for endothelial-mesenchymal transformation during heart cushion development in the mouse. J Cell Biol. 2004;166:359–67. doi: 10.1083/jcb.200403050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Iso T, Hamamori Y, Kedes L. Notch signaling in vascular development. Arterioscler Thromb Vasc Biol. 2003;23:543–53. doi: 10.1161/01.ATV.0000060892.81529.8F. [DOI] [PubMed] [Google Scholar]
- 52.Liebner S, Cattelino A, Gallini R, Rudini N, Iurlaro M, Piccolo S, Dejana E. Beta-catenin is required for endothelial-mesenchymal transformation during heart cushion development in the mouse. J Cell Biol. 2004;166:359–67. doi: 10.1083/jcb.200403050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Noseda M, McLean G, Niessen K, Chang L, Pollet I, Montpetit R, Shahidi R, Dorovini-Zis K, Li L, Beckstead B, Durand RE, Hoodless PA, Karsan A. Notch activation results in phenotypic and functional changes consistent with endothelial-to-mesenchymal transformation. Circ Res. 2004;94:910–7. doi: 10.1161/01.RES.0000124300.76171.C9. [DOI] [PubMed] [Google Scholar]
- 54.Potenta S, Zeisberg E, Kalluri R. The role of endothelial-to-mesenchymal transition in cancer progression. Br J Cancer. 2008;99:1375–9. doi: 10.1038/sj.bjc.6604662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Potenta S, Zeisberg E, Kalluri R. The role of endothelial-to-mesenchymal transition in cancer progression. Br J Cancer. 2008;99:1375–9. doi: 10.1038/sj.bjc.6604662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Torreggiani E, Lisignoli G, Manferdini C, Lambertini E, Penolazzi L, Vecchiatini R, Gabusi E, Chieco P, Facchini A, Gambari R, Piva R. Role of Slug transcription factor in human mesenchymal stem cells. J Cell Mol Med. 2012;16:740–51. doi: 10.1111/j.1582-4934.2011.01352.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Barrallo-Gimeno A, Nieto MA. The Snail genes as inducers of cell movement and survival: implications in development and cancer. Development. 2005;132:3151–61. doi: 10.1242/dev.01907. [DOI] [PubMed] [Google Scholar]
- 58.Conacci-Sorrell M, Simcha I, Ben-Yedidia T, Blechman J, Savagner P, Ben-Ze’ev A. Autoregulation of E-cadherin expression by cadherin-cadherin interactions: the roles of beta-catenin signaling, Slug, and MAPK. J Cell Biol. 2003;163:847–57. doi: 10.1083/jcb.200308162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Nieto MA. The snail superfamily of zinc-finger transcription factors. Nat Rev Mol Cell Biol. 2002;3:155–66. doi: 10.1038/nrm757. [DOI] [PubMed] [Google Scholar]
- 60.van Meeteren LA, ten DP. Regulation of endothelial cell plasticity by TGF-beta. Cell Tissue Res. 2012;347:177–86. doi: 10.1007/s00441-011-1222-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Arciniegas E, Frid MG, Douglas IS, Stenmark KR. Perspectives on endothelial-to-mesenchymal transition: potential contribution to vascular remodeling in chronic pulmonary hypertension. Am J Physiol Lung Cell Mol Physiol. 2007;293:L1–L8. doi: 10.1152/ajplung.00378.2006. [DOI] [PubMed] [Google Scholar]
- 62.Arciniegas E, Frid MG, Douglas IS, Stenmark KR. Perspectives on endothelial-to-mesenchymal transition: potential contribution to vascular remodeling in chronic pulmonary hypertension. Am J Physiol Lung Cell Mol Physiol. 2007;293:L1–L8. doi: 10.1152/ajplung.00378.2006. [DOI] [PubMed] [Google Scholar]
- 63.Thiery JP, Acloque H, Huang RY, Nieto MA. Epithelial-mesenchymal transitions in development and disease. Cell. 2009;139:871–90. doi: 10.1016/j.cell.2009.11.007. [DOI] [PubMed] [Google Scholar]
- 64.Arciniegas E, Frid MG, Douglas IS, Stenmark KR. Perspectives on endothelial-to-mesenchymal transition: potential contribution to vascular remodeling in chronic pulmonary hypertension. Am J Physiol Lung Cell Mol Physiol. 2007;293:L1–L8. doi: 10.1152/ajplung.00378.2006. [DOI] [PubMed] [Google Scholar]
- 65.Kovacic JC, Mercader N, Torres M, Boehm M, Fuster V. Epithelial-to-mesenchymal and endothelial-to-mesenchymal transition: from cardiovascular development to disease. Circulation. 2012;125:1795–808. doi: 10.1161/CIRCULATIONAHA.111.040352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Kovacic JC, Mercader N, Torres M, Boehm M, Fuster V. Epithelial-to-mesenchymal and endothelial-to-mesenchymal transition: from cardiovascular development to disease. Circulation. 2012;125:1795–808. doi: 10.1161/CIRCULATIONAHA.111.040352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Kovacic JC, Mercader N, Torres M, Boehm M, Fuster V. Epithelial-to-mesenchymal and endothelial-to-mesenchymal transition: from cardiovascular development to disease. Circulation. 2012;125:1795–808. doi: 10.1161/CIRCULATIONAHA.111.040352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Yoshino T, Saito D, Tadokoro R, Takahashi Y. In vivo gene manipulations of epithelial cell sheets: a novel model to study epithelial-to-mesenchymal transition. Dev Growth Differ. 2011;53:378–88. doi: 10.1111/j.1440-169X.2011.01252.x. [DOI] [PubMed] [Google Scholar]
- 69.Lamouille S, Xu J, Derynck R. Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol. 2014;15:178–96. doi: 10.1038/nrm3758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Ocana OH, Nieto MA. Epithelial plasticity, stemness and pluripotency. Cell Res. 2010;20:1086–8. doi: 10.1038/cr.2010.127. [DOI] [PubMed] [Google Scholar]
- 71.Obokata H, Wakayama T, Sasai Y, Kojima K, Vacanti MP, Niwa H, Yamato M, Vacanti CA. Stimulus-triggered fate conversion of somatic cells into pluripotency. Nature. 2014;505:641–7. doi: 10.1038/nature12968. [DOI] [PubMed] [Google Scholar]
- 72.Gurdon JB. Adult frogs derived from the nuclei of single somatic cells. Dev Biol. 1962;4:256–73. doi: 10.1016/0012-1606(62)90043-x. [DOI] [PubMed] [Google Scholar]
- 73.Wakayama T, Perry AC, Zuccotti M, Johnson KR, Yanagimachi R. Full-term development of mice from enucleated oocytes injected with cumulus cell nuclei. Nature. 1998;394:369–74. doi: 10.1038/28615. [DOI] [PubMed] [Google Scholar]
- 74.Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126:663–76. doi: 10.1016/j.cell.2006.07.024. [DOI] [PubMed] [Google Scholar]
- 75.Smith A. Cell therapy: in search of pluripotency. Curr Biol. 1998;8:R802–R804. doi: 10.1016/s0960-9822(07)00504-0. [DOI] [PubMed] [Google Scholar]
- 76.Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126:663–76. doi: 10.1016/j.cell.2006.07.024. [DOI] [PubMed] [Google Scholar]
- 77.Kovacic JC, Mercader N, Torres M, Boehm M, Fuster V. Epithelial-to-mesenchymal and endothelial-to-mesenchymal transition: from cardiovascular development to disease. Circulation. 2012;125:1795–808. doi: 10.1161/CIRCULATIONAHA.111.040352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.McDonald OG, Wu H, Timp W, Doi A, Feinberg AP. Genome-scale epigenetic reprogramming during epithelial-to-mesenchymal transition. Nat Struct Mol Biol. 2011;18:867–74. doi: 10.1038/nsmb.2084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Esteban MA, Bao X, Zhuang Q, Zhou T, Qin B, Pei D. The mesenchymal-to-epithelial transition in somatic cell reprogramming. Curr Opin Genet Dev. 2012;22:423–8. doi: 10.1016/j.gde.2012.09.004. [DOI] [PubMed] [Google Scholar]
- 80.Li R, Liang J, Ni S, Zhou T, Qing X, Li H, He W, Chen J, Li F, Zhuang Q, Qin B, Xu J, Li W, Yang J, Gan Y, Qin D, Feng S, Song H, Yang D, Zhang B, Zeng L, Lai L, Esteban MA, Pei D. A mesenchymal-to-epithelial transition initiates and is required for the nuclear reprogramming of mouse fibroblasts. Cell Stem Cell. 2010;7:51–63. doi: 10.1016/j.stem.2010.04.014. [DOI] [PubMed] [Google Scholar]
- 81.Takahashi K. Cellular reprogramming--lowering gravity on Waddington’s epigenetic landscape. J Cell Sci. 2012;125:2553–60. doi: 10.1242/jcs.084822. [DOI] [PubMed] [Google Scholar]
- 82.Takahashi K. Cellular reprogramming--lowering gravity on Waddington’s epigenetic landscape. J Cell Sci. 2012;125:2553–60. doi: 10.1242/jcs.084822. [DOI] [PubMed] [Google Scholar]
- 83.Davis RL, Weintraub H, Lassar AB. Expression of a single transfected cDNA converts fibroblasts to myoblasts. Cell. 1987;51:987–1000. doi: 10.1016/0092-8674(87)90585-x. [DOI] [PubMed] [Google Scholar]
- 84.Takahashi K. Cellular reprogramming--lowering gravity on Waddington’s epigenetic landscape. J Cell Sci. 2012;125:2553–60. doi: 10.1242/jcs.084822. [DOI] [PubMed] [Google Scholar]
- 85.Hay ED. The mesenchymal cell, its role in the embryo, and the remarkable signaling mechanisms that create it. Dev Dyn. 2005;233:706–20. doi: 10.1002/dvdy.20345. [DOI] [PubMed] [Google Scholar]
- 86.Cavarretta E, Latronico MV, Condorelli G. Endothelial-to-mesenchymal transition and microRNA-21: the game is on again. Arterioscler Thromb Vasc Biol. 2012;32:165–6. doi: 10.1161/ATVBAHA.111.242008. [DOI] [PubMed] [Google Scholar]
- 87.Potenta S, Zeisberg E, Kalluri R. The role of endothelial-to-mesenchymal transition in cancer progression. Br J Cancer. 2008;99:1375–9. doi: 10.1038/sj.bjc.6604662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Kovacic JC, Mercader N, Torres M, Boehm M, Fuster V. Epithelial-to-mesenchymal and endothelial-to-mesenchymal transition: from cardiovascular development to disease. Circulation. 2012;125:1795–808. doi: 10.1161/CIRCULATIONAHA.111.040352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Potenta S, Zeisberg E, Kalluri R. The role of endothelial-to-mesenchymal transition in cancer progression. Br J Cancer. 2008;99:1375–9. doi: 10.1038/sj.bjc.6604662. [DOI] [PMC free article] [PubMed] [Google Scholar]