

Abstract
During blood stage infection, malaria parasites invade, mature, and replicate within red blood cells (RBCs). This results in a regular growth cycle and an exponential increase in the proportion of malaria infected RBCs, known as parasitemia. We describe a flow cytometry based protocol which utilizes a combination of the DNA dye Hoechst, and the mitochondrial membrane potential dye, JC-1, to identify RBCs which contain parasites and therefore the parasitemia, of in vivo blood samples from Plasmodium chabaudi adami DS infected mice. Using this approach, in combination with fluorescently conjugated antibodies, parasitized RBCs can be distinguished from leukocytes, RBC progenitors, and RBCs containing Howell-Jolly bodies (HJ-RBCs), with a limit of detection of 0.007% parasitemia. Additionally, we outline a method for the comparative assessment of merozoite invasion into two different RBC populations. In this assay RBCs, labeled with two distinct compounds identifiable by flow cytometry, are transfused into infected mice. The relative rate of invasion into the two populations can then be assessed by flow cytometry based on the proportion of parasitized RBCs in each population over time. This combined approach allows the accurate measurement of both parasitemia and merozoite invasion in an in vivo model of malaria infection.
Keywords: Infection, Issue 98, Malaria, Plasmodium, chabaudi, flow cytometry, parasitemia, merozoite, invasion, in vivo, JC-1
Introduction
The clinical symptoms associated with malaria occur during the Plasmodium parasite’s asexual replicative cycle within red blood cells (RBCs). Merozoites, released during the liver stage of infection, quickly attach to and invade RBCs. After gaining entry into the cell, the parasite grows and matures, eventually undergoing schizogony, splitting open the cell, and releasing a cluster of newly formed merozoites which go on to repeat this cycle. As such, an assessment of malaria infection often involves monitoring both parasitemia, which is the percentage of RBCs appropriated by one or more parasites, and the rate of merozoite invasion into uninfected RBCs.
Flow cytometry is a powerful tool which can be used to record the properties of vast numbers of cells in a short period of time. This technique has clear applicability for the measurement of malaria parasitemia and invasion, and offers several advantages over traditional microscopy techniques. These include the accurate measurement of very low parasitemia, which would be prohibitively time consuming by microscopy, the unbiased nature of the measurement, and the ability to measure multiple cell parameters simultaneously. Flow cytometry is widely used to determine both parasitemia and merozoite invasion in in vitro culture1-9, however, techniques for measuring these parameters in vivo are less well developed, and can be complicated by the presence of additional cell types which interfere with analysis. No assays have been described for measurement of in vivo invasion, and while some assays exist for the analysis of in vivo parasitemia, these lack the ability to distinguish between parasitized RBCs (pRBCs) and RBCs containing Howell-Jolly bodies (HJ-RBCs)10-13. The later issue is particularly important as in mice HJ-RBCs may account for up to 0.9% of mature RBCs14-16, thereby preventing the accurate measurement of low parasitemia.
We have previously demonstrated an approach for the measurement of parasitemia and merozoite invasion in a rodent model of malaria infection14. Here, we provide a more detailed protocol and accompanying video. This approach builds on previous methodologies and allows for the accurate identification of parasitized RBCs, as distinct from leukocytes, RBC progenitors, and HJ-RBCs. Additionally, this assay allows the simultaneous measurement of merozoite invasion into two labeled RBC populations, a treated, or target, population, and a control population, thereby providing a robust platform for the assessment of invasion into different cell types.
Protocol
All procedures were conducted in accordance with the policies of Macquarie University and conformed to the National Health and Medical Research Council (NHMRC) Australian code of practice. The work was performed under the agreement Ethics No ARA 2012/017 approved and obtained from the Animal Ethics Committee at Macquarie University. All experiments were performed on SJL/J mice unless otherwise stated.
1. Mice and Experimental Malaria Infection
House mice under controlled temperature (21 °C) with a 12:12 hr light-dark cycle.
Thaw an aliquot of cryopreserved P. chabaudi adami DS with 5% parasitized RBCs (pRBCs) and inject 200 µl into the intraperitoneal cavity of C57BL/6 donor mouse.
- Monitor donor mouse parasitemia every 1 - 2 days using flow cytometry as described in Section 3 - 4 of this protocol. Once donors reaches 5 - 15% parasitemia, collect blood by cardiac puncture as follows:
- Aspirate 100 μl of heparin solution (300 U/ml heparin in mouse Ringer solution (MTR) (154 mM NaCl, 5.6 mM KCl, 1 mM MgCl2, 2.2 mM CaCl2, 20 mM HEPES, 10 mM glucose, 30 U/ml heparin, pH 7.4, 0.22 μM filter sterilized)) into a 1 ml syringe with a 26 G needle.
- Anesthetize the mouse by inhalation with 5% isofluorane, confirming the desired depth of anesthesia by absence of pedal withdrawal response and corneal reflexes.
- Place mouse on its side and perpendicularly insert needle just below the elbow, through the ribs, and into the heart. Pull out syringe plunger slowly and rotate needle until 0.5 - 1 ml of blood is obtained.
- Perform humane euthanasia by cervical dislocation under anesthesia.
Calculate the pRBCs per volume of donor blood assuming a blood count of 9 x 106 RBC/µl (i.e., if the donor was at 10% parasitemia when sacrificed, blood will contain 9 x 105 pRBC/µl). Dilute parasitized blood in Krebs buffered saline (126 mM NaCl, 2.5 mM KCl, 25 mM NaHCO3, 1.2 mM NaH2PO4, 1.2 mM MgCl2, 2.5 mM CaCl2, 0.2% glucose, pH 7.2, 0.22 μM filter sterilized) so that the concentration is 1 x 104 pRBCs per 200 µl and inject 200 µl into the intraperitoneal cavity of mice required for the RBC labeling and transfusion experiment.
Monitor parasitemia every 1 - 2 days using flow cytometry as described in Sections 3 - 4 of this protocol.
2. Labeling of RBCs and Transfusion
Collect heparinized blood from mice by cardiac puncture as described in step 1.3, and place on ice. Keep blood at 4 °C at all times. Collect approximately 200 µl blood per mouse to be injected. If necessary, split blood into two samples and treat one sample as desired. NOTE: In this way, invasion of the treated sample can be compared to a control, untreated sample (see Figure 1 for schematic).
- Prepare a 2x solution of each fluorescent RBC label.
- Dilute a 2 mg/ml stock solution of Atto 633-N-Hydroxysuccinimide (Atto 633-NHS) in MTR so that the concentration is 20 μg/ml.
- Dilute a 25 mg/ml stock solution of Biotin-N-Hydroxysuccinimide (Biotin-NHS) in MTR so that the concentration is 250 μg/ml.
Split blood samples into two tubes and add an equal volume of 2x Atto 633-NHS solution to one tube, and an equal volume of the 2x Biotin-NHS to the other tube. Mix immediately. Incubate blood at 4 °C for 1 hr with constant slow mixing, or alternatively, mix sample every 10 - 15 min.
- Wash labeled blood 3 times with MTRC (MTR + 0.5% bovine serum albumin (BSA), 0.22 μM filter sterilized) as follows:
- Add at least 2 volumes of MTRC, centrifuge at 750 x g for 5 min, remove the supernatant, and resuspend the pellet in at least 2 volumes of MTRC. Repeat 2 more times.
- After the final wash, centrifuge at 750 x g for 5 min and combine the pelleted blood in different combinations (i.e., untreated Atto 633/treated Biotin, untreated Biotin/treated Atto 633). Add MTR to make up to a volume of 200 µl per mouse to be injected.
- Inject labeled blood intravascularly into rodent malaria infected mice or uninfected controls as follows:
- Perform transfusion at 2 - 15% parasitemia at the peak of parasite schizogony. NOTE: this occurs approximately halfway through the dark cycle (i.e., between 12 - 2 am on a normal light cycle, or 12 - 2 pm on a reverse light cycle).
- Warm mice using a heat lamp for 5-10 mins to increase blood flow, but take precautions not to overheat the mice. Place mice in a restraining device and intravascularly inject 200 μl labeled blood (approximately 1 - 2 x 109 RBCs) via the tail vein.
Collect blood samples at different time points after injection as described in Section 3. NOTE: invasion rates can be quantified less than 10 min after injection, depending on parasitemia of the recipient mouse.
3. Collection of Blood Samples and Preparation for Flow Cytometry
- Prepare 50 μl of staining solution per sample, plus extra, on the day of the experiment as follows:
- Defrost an aliquot of 6 mM JC-1 stored at -20 °C in dimethyl sulfoxide (DMSO).
- Warm 50 μl of MTRC per sample, plus extra, to 37 °C.
- Whilst continuously vortexing the pre-warmed MTRC, add JC-1 so that the final concentration is 12 μM. This is done to reduce the formation of JC-1 aggregates; if aggregates do form, do not centrifuge tube as this will prevent staining. NOTE: A small amount of aggregation will not affect results.
- Add anti-CD45 APC eFluor 780 and anti-CD71 PerCP eFluor 710 so that the final concentration is 1 μg/ml. If performing the labeled RBC experiment, add streptavidin PE-Cy7 so that the final concentration is 1 μg/ml.
Perform tail bleeding by amputation of the tip of the tail or laceration of the blood vessel within the tail. Place a drop of tail blood onto a small weigh boat or glass slide. After blood collection, ensure that the bleeding has stopped. Pipette 3 μl of tail blood into 50 μl of staining solution pre-warmed to 37 °C and incubate samples at 37 °C for 20 min.
Defrost an aliquot of 4 mM Hoechst 33342 stored at -20 °C in distilled water (if using a 355 nm laser) or 2 mM Hoechst 34580 stored at -20 °C in distilled water (if using a 405 nm laser). Prepare 500 μl per sample, plus extra, of a 4 μM or 2 μM solution of Hoechst in MTRC, respectively.
Add 500 μl of 4 μM Hoechst 33342 or 2 μM Hoechst 34580 to samples and incubate at RT for 20 min.
Centrifuge cells at 750 x g for 3 min at 4 °C, discard supernatant, add 700 μl MTRC and analyze by flow cytometry.
4. Flow Cytometry
Analyze samples using a 355/488/633 nm laser or 405/488/633 nm laser instrument.
Filter samples through a 35 μM cell strainer immediately before analysis. Rinse cell strainer with distilled water between samples and re-use.
Record enough events so that the smallest population contains at least 500 events (usually 1,000,000 - 10,000,000 total events per sample).
Excite Hoechst 33342 using a 355 nm laser and detect events through a 460/50 filter. Excite Hoechst 34580 using a 405 nm laser and detect events through a 460/50 filter. Excite JC-1, anti-CD71 PerCP eFluor 710 and Streptavidin PE-Cy7 using a 488 nm laser and detect events through a 530/40, 692/40, and 750LP filter respectively. Excite Atto 633 and anti-CD45 APC eFluor 780 using a 633 nm laser and detect events through a 670/30 or 750LP filter respectively.
- Perform analysis and compensation using appropriate flow cytometry analysis software as follows:
- Select whole cells and exclude noise, debris, and platelets based on FSC/SSC properties (Figure 2A). Select single cells based on either trigger pulse width (Figure 2B) or by using the FSC peak area to height ratio (Figure 2C).
- Select mature RBCs, and exclude RBC progenitors and leukocytes, based on negative fluorescence in the PerCP eFluor 710 and APC eFluor 780 channels (Figure 2D).
- If performing the labeled RBC experiment, select each population of labeled RBCs based on PE-Cy7 and Atto 633 fluorescence and analyze these separately (Figure 3A).
- Select pRBCs based on positive fluorescence for both JC-1 and Hoechst (Figure 2E-H).
5. Calculations and Statistics
Calculate parasitemia by dividing the number of pRBCs by the total number of RBCs (i.e., number of events in gate Q2 divided by number of events in gate G4). When conducting the invasion assay parasitemia of the two labeled populations can be calculated by dividing the number of labeled pRBCs by the number of labeled RBCs in that population (i.e., number of events which are in both gates L1 and Q2, divided by number of events in gate L1). NOTE: Labeled parasitemia varies considerably depending on factors such as endogenous parasitemia and number of circulating merozoites. For this reason, it is helpful to report the parasitemia ratio, which is calculated as the parasitemia of the treated labeled population divided by the parasitemia of the control labeled population in each individual mouse. To correct for possible dye effects, report results as an average parasitemia ratio across the two dye combinations.
Determine statistical significance of ratios using the one sample t-test with 1 as the hypothetical mean.
Representative Results
Measurement of parasitemia.
For the measurement of parasitemia, blood cells should first be selected, and noise, debris and platelets excluded, based on FSC/SSC properties (Figure 2A). Depending on the cytometer used, single cells should then be selected based on either trigger pulse width (Figure 2B), or FSC peak height to area ratio (Figure 2C). Remaining events should consist of leukocytes, stained positive for APC eFluor 780, RBC progenitors (including reticulocytes), stained positive for PerCP eFluor 710, and mature RBCs, negative for both these stains (Figure 2D). After selecting the mature RBC population, events should separate into four populations when JC-1 fluorescence is plotted against Hoechst 33342 (Figure 2E,F) or Hoechst 34580 (Figure 2G,H) fluorescence. The dual positive population represents parasitized RBCs, while the remaining populations are either uninfected RBCs (dual negative), HJ-RBCs (Hoechst positive, JC-1 negative), or unknown cells (JC-1 positive, Hoechst negative). Notably, JC-1 fluorescence may shift from a maximum at 529 nm to a broad emission peaking at 590 nm, depending on the mitochondrial membrane potential of the cell. Regardless of this shift JC-1 will fluoresce strongly in the 530/40 nm channel, making this channel most suitable for analysis. However, in some instances it may be worthwhile to additionally record fluorescence in the 585/42 nm channel, which may provide an indication of the mitochondrial membrane potential of cells. In uninfected samples less than 0.007% of events should be dual positive (Figure 2E,G). This result will depend on the cleanliness of the flow cytometer and may vary considerably, extensive cleaning before analysis may improve results. HJ-RBCs account for 0.3 - 0.9% of mature RBCs, making the addition of the JC-1 dye critical for the accurate measurement of low parasitemia.
Assessment of merozoite invasion.
After injecting labeled RBCs into infected mice the relative invasion rates into the two labeled populations can be determined. Blood samples should be taken and prepared as described for the parasitemia measurement with the addition of Streptavidin PE Cy7. The time at which the sample is taken depends on the experimental conditions and desired outcome of the analysis. In general, an earlier time point will best reflect an invasion phenotype, although it may be beneficial to collect multiple time points to increase accuracy. For the analysis of flow cytometry data, mature RBCs should be selected as in Figure 2. From these cells, Atto 633 and biotin labeled RBCs can be identified based on fluorescence in the 670/30 and 750LP channels respectively (Figure 3A). From here on the three RBC populations (Atto 633, biotin, and unlabeled) should be analyzed separately. In each of these populations parasitemia can be determined based on staining with Hoechst and JC-1 (Figure 3B-D).
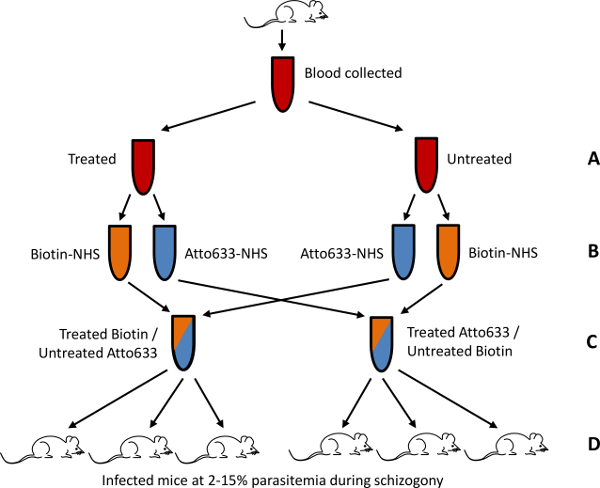 Figure 1. Schematic of the in vivo parasite invasion assay. An example of how this assay can be used to determine relative invasion rates in RBCs treated according to the user’s biological question is shown. Blood collected from uninfected mice is divided into two tubes. One tube is treated as desired while the other sample is left untreated (A). Tubes are again divided with one labeled with NHS-Biotin and the other with NHS-Atto 633 (B). Samples are then combined in two combinations; Biotin labeled treated RBCs with Atto 633 labeled untreated RBCs and Atto 633 labeled treated RBCs with Biotin labeled untreated RBCs (C). These two combinations are injected separately into two lots of infected mice during schizogony at 2 - 15% parasitemia (D). A total of 6 recipient mice is recommended to gain statistical significance in the result, although more or less may be used if needed. Adapted from Lelliott et al.14, originally published by BioMed Central.
Figure 1. Schematic of the in vivo parasite invasion assay. An example of how this assay can be used to determine relative invasion rates in RBCs treated according to the user’s biological question is shown. Blood collected from uninfected mice is divided into two tubes. One tube is treated as desired while the other sample is left untreated (A). Tubes are again divided with one labeled with NHS-Biotin and the other with NHS-Atto 633 (B). Samples are then combined in two combinations; Biotin labeled treated RBCs with Atto 633 labeled untreated RBCs and Atto 633 labeled treated RBCs with Biotin labeled untreated RBCs (C). These two combinations are injected separately into two lots of infected mice during schizogony at 2 - 15% parasitemia (D). A total of 6 recipient mice is recommended to gain statistical significance in the result, although more or less may be used if needed. Adapted from Lelliott et al.14, originally published by BioMed Central.
 Figure 2. Measurement of parasitemia by flow cytometry. Debris, noise, and platelets are removed from analysis based on FSC/SSC properties (A). Single cells are then selected from the remaining cells based on either trigger pulse width (B), or FSC peak to area ratio (C). Cell types can then be distinguished based on positive staining with CD45 APC eFluor 780 (leukocytes), CD71 PerCP eFluor 710 (RBC progenitors), or negative staining (mature RBCs) (D). After selecting mature RBCs, cells are either: Hoechst and JC-1 positive (parasitized RBCs), Hoechst positive and JC-1 negative (RBCs containing Howell Jolly bodies), JC-1 positive and Hoechst negative (unknown cells), or dual negative (uninfected RBCs). Uninfected and P. chabaudi infected samples stained with Hoechst 33342 (E,F), or Hoechst 34580 (G,H), are shown. Please click here to view a larger version of this figure.
Figure 2. Measurement of parasitemia by flow cytometry. Debris, noise, and platelets are removed from analysis based on FSC/SSC properties (A). Single cells are then selected from the remaining cells based on either trigger pulse width (B), or FSC peak to area ratio (C). Cell types can then be distinguished based on positive staining with CD45 APC eFluor 780 (leukocytes), CD71 PerCP eFluor 710 (RBC progenitors), or negative staining (mature RBCs) (D). After selecting mature RBCs, cells are either: Hoechst and JC-1 positive (parasitized RBCs), Hoechst positive and JC-1 negative (RBCs containing Howell Jolly bodies), JC-1 positive and Hoechst negative (unknown cells), or dual negative (uninfected RBCs). Uninfected and P. chabaudi infected samples stained with Hoechst 33342 (E,F), or Hoechst 34580 (G,H), are shown. Please click here to view a larger version of this figure.
 Figure 3. Assessment of merozoite invasion into two labeled RBC populationsin vivo. Plots are of mature RBCs, gated as in Figure 2. The two labeled RBC populations can be selected based on their label (A). Atto 633 labeled cells fluoresce in the 670/30 channel (L1), biotin labeled cells bind to streptavidin and fluoresce in the PE-Cy7 channel (L3), and unlabeled cells are negative for these stains (L4). In each of these populations parasitized RBCs can be identified based on Hoechst and JC-1 positive staining (Q2) (B-D). Please click here to view a larger version of this figure.
Figure 3. Assessment of merozoite invasion into two labeled RBC populationsin vivo. Plots are of mature RBCs, gated as in Figure 2. The two labeled RBC populations can be selected based on their label (A). Atto 633 labeled cells fluoresce in the 670/30 channel (L1), biotin labeled cells bind to streptavidin and fluoresce in the PE-Cy7 channel (L3), and unlabeled cells are negative for these stains (L4). In each of these populations parasitized RBCs can be identified based on Hoechst and JC-1 positive staining (Q2) (B-D). Please click here to view a larger version of this figure.
Discussion
We have described a method for the measurement of both parasitemia and merozoite invasion of in vivo samples. In terms of parasitemia measurement, this method offers an advantage over previous methods10-13 in that HJ-RBCs can be distinguished from pRBCs, thereby reducing the number of false positive events. While HJ-RBCs are usually rare in humans, some studies report high levels in mice15,16 making the distinction between these cells and pRBCs important for the accurate measurement of rodent parasitemia. Using this method the limit of detection for parasitemia is approximately 0.007%14, although it is possible to reduce this as low as 0.0005% under the right conditions. Notably, this limit is dictated by the cleanliness of the flow cytometer used and the ability to obtain large numbers of events in a reasonable amount of time. The most common problem associated with the use of this technique is insufficient staining with JC-1. The fluorescence of JC-1 depends on the mitochondrial membrane potential of the parasite, and parasites must be healthy in order to obtain optimal staining. Prolonged storage of parasites or fixation should therefore be avoided before staining.
Care should also be exercised when analyzing parasites which have been subject to drug interventions, as this may also affect the extent of staining with JC-1. In these cases it may be helpful to analyze parasitemia based on Hoechst fluorescence alone, as compared to parasitemia with the addition of the JC-1 dye. Indeed, this may provide a measure of parasite count, as well as an overall indication of parasite health. Finally, the JC-1 dye may shift its fluorescence from a peak at 529 nm to a peak at 590 nm by forming J-aggregates. The formation of these aggregates depends on dye concentration, which in turn depends on the mitochondrial membrane potential of the cell. The loss of mitochondrial membrane potential causes a shift in fluorescence towards 529 nm, which may indicate cells are apoptotic. In our experience pRBCs stained with JC-1 always exhibited a similar fluorescence pattern which was of approximately equal intensity in both the 530/40 nm channel and the 585/42 nm channel. However, in some studies, such as those employing drug interventions, it may be worthwhile to monitor changes in the fluorescence ratio between the 530/40 nm channel and the 585/42 nm channel for any indications of the loss of mitochondrial membrane potential. It is also important to note that JC-1 is particularly susceptible to degradation and should be freshly prepared from a frozen aliquot as described, while freeze/thaw cycles should be avoided. After staining, JC-1 fluorescence is relatively stable with little or no reduction in fluorescence, at least over several hours.
For the in vivo invasion assay, the two most critical factors are consistency in the RBC labeling process and the time of injection of labeled blood. For RBC labeling, the minimum amount of dye should be used to prevent any interference with the ability of the parasite to invade the RBC, which has been raised as a possibility when using surface labels7. Under the labeling conditions described here invasion is not inhibited14, however careful attention should be given to maintaining consistent label concentrations, and dyes should be switched, in order to prevent any inaccuracies due to invasion inhibition by the RBC label. As parasites undergo a synchronized invasion cycle, it is important that labeled cells are injected at the right time in the parasites lifecycle. This should be done in order to maximize the number of invasion events, and therefore the accuracy of the assay. In our experience the peak of invasion occurs approximately halfway through the dark cycle, making a reverse light cycle room advantageous for this assay.
The two label invasion assay offers an accurate method to determine merozoite invasion efficiency into different types of RBC in vivo. This approach therefore allows a comparison between treated RBCs, i.e., trypsinized, and control RBCs. Alternatively, RBCs from genetically modified mice could be compared to those of control mice, or RBCs of different physiological parameters or different age could be compared. While a similar assay has been described for the measurement of invasion in vitro9, the assay here provides the potential to account for effects only present in vivo, including the influence of the immune system, which can potentially alter parasite invasion mechanisms17-19. Finally, this assay also has the potential to provide insights into pathologies such as the immune clearance of pRBCs, and parasite growth, if the pRBCs within the labeled populations are monitored over an extended period.
Overall, a detailed protocol for the accurate measurement of parasitemia in in vivo samples is described, which has an advantage over previous assays in that HJ-RBCs can be distinguished from pRBCs. Furthermore, an assay is described for quantification of invasion efficiency into different RBC types in vivo.
Disclosures
The authors have nothing to disclose.
Acknowledgments
We acknowledge funding support from the National Health and Medical Research Council (grant APP605524, 490037 and 1047082), the Australian Research Council (grant DP12010061), the National Collaborative Research Infrastructure Strategy of Australia and the Education investment fund from the Department of Innovation, Industry, Science and Research. PML is a recipient of an Australian Postgraduate award.
References
- Jacobberger JW, Horan PK, Hare JD. Analysis of malaria parasite-infected blood by flow cytometry. Cytometry. 1983;4(3):228–237. doi: 10.1002/cyto.990040307. [DOI] [PubMed] [Google Scholar]
- Bianco AE, Battye FL, Brown GV. Plasmodium falciparum: rapid quantification of parasitemia in fixed malaria cultures by flow cytometry. Exp Parasitol. 1986;62(2):275–282. doi: 10.1016/0014-4894(86)90032-9. [DOI] [PubMed] [Google Scholar]
- Makler MT, Lee LG, Recktenwald D. Thiazole orange: a new dye for Plasmodium species analysis. Cytometry. 1987;8(6):568–570. doi: 10.1002/cyto.990080606. [DOI] [PubMed] [Google Scholar]
- Heyde HC, Elloso MM, van de Waa J, Schell K, Weidanz WP. Use of hydroethidine and flow cytometry to assess the effects of leukocytes on the malarial parasite Plasmodium falciparum. Clin Diagn Lab Immunol. 1995;2(4):417–425. doi: 10.1128/cdli.2.4.417-425.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pattanapanyasat K, et al. Culture of malaria parasites in two different red blood cell populations using biotin and flow cytometry. Cytometry. 1996;25(3):287–294. doi: 10.1002/(SICI)1097-0320(19961101)25:3<287::AID-CYTO10>3.0.CO;2-S. [DOI] [PubMed] [Google Scholar]
- Grimberg BT, Erickson JJ, Sramkoski RM, Jacobberger JW, Zimmerman PA. Monitoring Plasmodium falciparum growth and development by UV flow cytometry using an optimized Hoechst-thiazole orange staining strategy. Cytometry A. 2008;73(6):546–554. doi: 10.1002/cyto.a.20541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Theron M, Hesketh RL, Subramanian S, Rayner JC. An adaptable two-color flow cytometric assay to quantitate the invasion of erythrocytes by Plasmodium falciparum parasites. Cytometry A. 2010;77(11):1067–1074. doi: 10.1002/cyto.a.20972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bei AK, et al. A flow cytometry-based assay for measuring invasion of red blood cells by Plasmodium falciparum. Am J Hematol. 2010;85(4):234–237. doi: 10.1002/ajh.21642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark MA, et al. RBC barcoding allows for the study of erythrocyte population dynamics and P. falciparum merozoite invasion. PLoS One. 2014;9(7):e101041. doi: 10.1371/journal.pone.0101041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malleret B, et al. A rapid and robust tri-color flow cytometry assay for monitoring malaria parasite development. Sci Rep. 2011;1(118) doi: 10.1038/srep00118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jimenez-Diaz MB, et al. Quantitative measurement of Plasmodium-infected erythrocytes in murine models of malaria by flow cytometry using bidimensional assessment of SYTO-16 fluorescence. Cytometry A. 2009;75(3):225–235. doi: 10.1002/cyto.a.20647. [DOI] [PubMed] [Google Scholar]
- Jimenez-Diaz MB, et al. Improvement of detection specificity of Plasmodium-infected murine erythrocytes by flow cytometry using autofluorescence and YOYO-1. Cytometry A. 2005;67(1):27–36. doi: 10.1002/cyto.a.20169. [DOI] [PubMed] [Google Scholar]
- Jun G, Lee JS, Jung YJ, Park JW. Quantitative determination of Plasmodium parasitemia by flow cytometry and microscopy. J Korean Med Sci. 2012;27(10):1137–1142. doi: 10.3346/jkms.2012.27.10.1137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lelliott PM, Lampkin S, McMorran BJ, Foote SJ, Burgio G. A flow cytometric assay to quantify invasion of red blood cells by rodent Plasmodium parasites in vivo. Malar J. 2014;13(1):100. doi: 10.1186/1475-2875-13-100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morohashi K, et al. Structural and functional abnormalities in the spleen of an mFtz-F1 gene-disrupted mouse. Blood. 1999;93(5):1586–1594. [PubMed] [Google Scholar]
- Shet AS, et al. Morphological and functional platelet abnormalities in Berkeley sickle cell mice. Blood Cells Mol Dis. 2008;41(1):109–118. doi: 10.1016/j.bcmd.2008.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duraisingh MT, et al. Phenotypic variation of Plasmodium falciparum merozoite proteins directs receptor targeting for invasion of human erythrocytes. EMBO J. 2003;22(5):1047–1057. doi: 10.1093/emboj/cdg096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okoyeh JN, Pillai CR, Chitnis CE. Plasmodium falciparum field isolates commonly use erythrocyte invasion pathways that are independent of sialic acid residues of glycophorin A. Infect Immun. 1999;67(11):5784–5791. doi: 10.1128/iai.67.11.5784-5791.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dolan SA, Miller LH, Wellems TE. Evidence for a switching mechanism in the invasion of erythrocytes by Plasmodium falciparum. J Clin Invest. 1990;86(2):618–624. doi: 10.1172/JCI114753. [DOI] [PMC free article] [PubMed] [Google Scholar]