

Abstract
Molybdenum cofactor (Moco) is required for the activities of Moco-dependant enzymes. Cofactor for nitrate reductase and xanthine dehydrogenase (Cnx1) is known to be involved in the biosynthesis of Moco in plants. In this work, a soybean (Glycine max L.) Cnx1 gene (GmCnx1) was transferred into soybean using Agrobacterium tumefaciens-mediated transformation method. Twenty seven positive transgenic soybean plants were identified by coating leaves with phosphinothricin, bar protein quick dip stick and PCR analysis. Moreover, Southern blot analysis was carried out to confirm the insertion of GmCnx1 gene. Furthermore, expression of GmCnx1 gene in leaf and root of all transgenic lines increased 1.04-2.12 and 1.55-3.89 folds, respectively, as compared to wild type with GmCnx1 gene and in line 10 , 22 showing the highest expression. The activities of Moco-related enzymes viz nitrate reductase (NR) and aldehydeoxidase (AO) of T1 generation plants revealed that the best line among the GmCnx1 transgenic plants accumulated 4.25 μg g-1 h-1 and30 pmol L-1, respectively (approximately 2.6-fold and 3.9-fold higher than non-transgenic control plants).In addition, overexpression ofGmCnx1boosted the resistance to various strains of soybean mosaic virus (SMV). DAS-ELISA analysis further revealed that infection rate of GmCnx1 transgenic plants were generally lower than those of non-transgenic plants among two different virus strains tested. Taken together, this study showed that overexpression of a GmCnx1 gene enhanced NR and AO activities and SMV resistance, suggesting its important role in soybean genetic improvement.
Introduction
Molybdenum (Mo) forms a complex with the molybdopterin compound, which consequently forms a cofactor named molybdenum cofactor (Moco). All Mo-containing enzymes characterized to this end, besides nitrogenase contain a pterin-type cofactor[1]. In plants, six gene products have been identified to be involved in the Moco biosynthesis, which can be divided into four steps. The basic step of Moco biosynthesis begins with the conversion of guanosine triphosphate (GTP) to cyclic pyranopterin monophosphate (cPMP, previously identified as precursor Z) with the catalysis of Cnx2 and Cnx3 in the mitochondria, while all subsequent steps proceed in the cytosol. In the next step sulfur is transferred to cPMP so as propagate the intermediate MPT in a reaction catalyzed by copper (Cu) and the enzyme MPT synthase, a complex consisting of Cnx6 and Cnx7 subunits. The third step involves the insertion of Mo into MPT-AMP with adenylation of molybdopterin. In the final step, a product-substrate channel is built by the transference of MPT-AMP to the N-terminal domain of Cnx1 (Cnx1-E). Cnx1-E then cleaves the adenylate, releasing Cu and inserts Mo, thus producing active Moco [2,3].
Molybdenum containing enzymes play vital role in nitrogen assimilation, synthesis of phytohormones, detoxification and purine metabolism[4]. Although more than fifty Mo-containing enzymes are known, only the functions of five Mo-enzymes have been exploited so far, which include: nitrate reductase (NR;EC 1.7.1.1), aldehyde oxidase (AO; EC 1.2.3.1), xanthine dehydrogenase (XDH; EC 1.17.1.4), sulphite oxidase (SO; EC 1.8.3.1), and mitochondrial amidoxime reductase (mARC) [4]. Among these five Mo-enzymes, NR and AO are considered as vital enzymes involved in various processes of plant growth and regulation.
NR is a 200 kDa cytoplasmic enzyme, and consists of three functional domains: the N-terminal domain associated with Moco, the central haem-binding cytochrome b 5 domain, and the C-terminal FAD-binding domain. This dimerization is Moco dependent [5]. The three domains form three redox centers catalyzing the transfer of electrons from the reduced NAD(P)H via FAD, haem and Moco to nitrate.NR catalyses the first step in nitrate assimilation, a pathway that is of key importance for plant nutrition [6,7].The regulation of NR involves both transcriptional and post-translational mechanisms regulating the amount as well as activity of NR protein.
AO is a cytoplasmic enzyme with an apparent molecular mass of 300 kDa. The FAD, Fe, and Moco acts as aprosthetic groups and bind to the enzyme with the propertion of 4:1:1 [8].AO isoform (AAO3) acts on abscisic aldehyde, which is the native precursor of the plant hormone abscisic acid. Abscisic acid is believed to be involved in many developmental processes as well as for a variety of abiotic and biotic stress responses [9–11]. Due to broad substrate specificity AOs are involved in additional metabolic processes other than phytohormone synthesis. Detoxification reactions and pathogen response may be good candidates for these additional functions [12]. Recently it was shown that plant AOs produce H2O2 in response to drought stress and ABA treatment [13]. Hence, AO enzymes in plants are essential for many physiological processes.
However, recent studies about the physiological and biochemical functions of GmCnx1 Moco biosynthesis genes have been minute. Previous literatures have shown the importance of Cnx1 in forming a stable Moco, and that Cnx1 binds to actin filaments of plants cytoskeleton located on the membrane with N-terminal domains [14]. In addition, expression of a senescence-associated gene Ntdin was related to synthesis of Moco in tobacco, resulting in the increase of the content of Moco. NR and XDH activity was also improved in transgenic Ntdin plants [15]. It was also found that expression of the Mocosulfide enzyme in soybean enhanced drought resistance, which in turn improved its yield [16].Furthermore, overexpression of Cnx1 gene was related to SMV infection based on whole-genome microarray analysis [17].
Although studies regarding the function of Cnx1 in Moco biosynthesis have been widely conducted, its specific functions in enhancing NR, AO activities and SMV resistance have not been explored yet. Therefore, the main aim of this study was to elucidate the potential functions of GmCnx1 in enhancing NR, AO activities and SMV resistance using the Agrobacterium tumefaciens-mediated transformation, and characterization of the GmCnx1-overexpressing transgenic soybean plants.
Materials and Methods
Plant materials and growth
Soybean cultivar, Tianlong 1, was used as the recipient for genetic modification. The Agrobacterium tumefaciens-mediated transformation in soybean was carried out according to Zhang et al.[18] and Paz et al.[19] to get T0 seeds, and the T1 seeds were harvested from self-pollination of T0 progeny.
The wild type (WT) and transgenicGmCnx1 gene soybean plants were grown in a controlled greenhouse at Zijingang campus experimental farm in Zhejiang University, PR China, in 2013. The growth temperature of soybean was 28/20 ± 1°C (day/night) with a 16 h photoperiod under fluorescent white light. Plants were daily supplied with half-length Hoagland’s nutrient solution [20] during growth.
Isolation of the cDNA sequence and the construct of GmCnx1 binary vector
A gene-specific primer pair (5’-GGGGACAAGTTTGTACAAAAAAGCAGGCTTCACC-3’ and 5’-GGGGACCACTTTGTACAAGAAAGCTGGGTG-3’), was designed according to the predicted sequence of Cnx1 (NM_001255600; NCBI Reference Sequence). About 5 μg total RNA in leaf was reversely transcribed into first-strand cDNA by ThermoScripy Kit (Invitrogen, USA). The PCR product was sequenced and inserted into pDONOR221 (Invitrogen) and then transferred to a pB7FWG2 vector via LR recombination reaction of the Gateway system.
Identification of putative transgenic plants
Non-transgenic soybean plants and GmCnx1 transgenic soybean plants both produced the amplified band of GmCnx1 gene by PCR analysis, since the target gene is endogenous. The bar gene and GmCnx1 gene which were contained between LB and RB on the vector, were transferred together into soybean plants, so we selected bar gene as the identification standard for transgenic plants. The following methods were applied:
Firstly, about 10 d after the plantlets were transferred to soil, the T1 plants were screened for tolerance to the herbicide Basta by application of a 135 mg/L solution with a cotton swab lightening the upper surface of the half euphylls and labeling the other half with pen. Leaf tissue was recorded for herbicide tolerance at 6 or 7 d post lightening.
Secondly, quick detection of the bar protein was carried out by using Quickstix Kit for bar gene according to the manufacturer’s instructions (EnviroLogix Inc., USA). The presence of a test line (second line) on the membrane strip between the control line (common to all, including the non-transformed control) and the protective tape indicated the expression of the foreign bar protein in the transgenic plants.
Thirdly, bar-resistant transgenic plants were first identified by PCR amplification [21] with primers for the bar gene. Primers were designed according to the nucleotide sequence of selective maker gene bar (5’-CAGCTGCCAGAAACCCACGT-3’ and 5’-CTGCACCATCGTCAACCACT-3’). Amplification of gene bar used 2×Taq Master Mix (YEASEN, Shanghai, China). After completion of PCR amplification, the amplified product was detected on 1.5% agarose electrophoresis.
Validation of transgenic soybean plants by Southern blot analysis
Genomic DNA was extracted from leaf of non-transgenic and the putative T1 GmCnx1 transgenic plants (based on herbicide resistance, mentioned above) by the method described by Edwards et al.[22] with minor modifications. Plasmid DNA and genomic DNA were digested with restriction enzymes EcoR I and Hind Ⅲ, respectively, at 37°C overnight. Digested DNA was separated on 0.8% (w/v) agarose gel and blotted onto Hybond N+ nylon membrane. The membrane was hybridized with a DIG-labelled GmCnx1-specific probe (5’-CGAGTATCCCGTCGTTG-3’ and 5’-TTCCAGGTAGCCCAAAA-3’) at 68°C for 16 h. The hybridized membrane was washed and detected according to the protocol of High Prime DNA Labeling and Detection Starter Kit I (Roche, USA).
qRT-PCR analysis of the transgenic soybean plants
Fresh tissues of two-week-old T1transgenic and non-transgenic soybean plants were collected in liquid nitrogen before isolation of RNA. Total RNA was isolated leaf and root using Trizol reagent (TaKaRa, Dalian, China). First strand cDNA was synthesized with 1 μg of purified total RNA using PrimeScript RT reagent Kit with gDNA Eraser (TaKaRa). Real-time PCR reaction was carried out according to SYBR Premix Ex TaqII (TaKaRa). The gene-specific primers of GmCnx1 (5’-CCTGGAAACCCTAATGCTGT-3’ and 5’-CACATCTGCAGGAACTGCTT-3’) were used in the RT-PCR and gene—specific primers of actin11 (5’-ATCTTGACTGAGCGTGGTTATTCC-3’ and 5’-GCTGGTCCTGGCTGTCTCC-3’) were amplified as internal control to quantify the relative amounts of cDNA [23]. The amplifications were performed on CFX96 Real Time System (Bio-Rad, Hercules, CA, USA) and the relative expression was calculated by 2-ΔΔCt method [24]. Significant differences of all data were analyzed using DPS software.
NR activity assay
NR activity was measured according to the method described previously[25]. The NR activity of plant crude extracts was assayed using leaf of non-transgenic and T1 GmCnx1 transgenic plants, which were homogenized in extraction buffer and centrifuged for 15 min at 4,000 rpm. The supernatants were collected and added to the reaction buffer. After incubation at 25°C for 30 min, the reaction was stopped by 1 ml 1% sulphanilamide. The mixture was centrifuged for 5 min at 5,000 rpm and the supernatant was used to detect nitrite production by reading the absorbance at 540 nm after addition of N-(1-naphthyl) ethylenediamine dihydrochloride. NO2 - was quantified to calculate NR activity.
AO activity assay
Detection of AO activity was performed by using Plant Aldehyde Oxidase (AO) Elisa Kit (TSZ Co. Ltd., USA) according to the manufacturer’s instructions. Enzyme linked immunosorbent assay——sandwich technique was employed in the experiments. Briefly, standard wells and testing sample wells were set. All of 50μl standards, 50μl samples and 100μl HRP-conjugate reagents were added into standard wells and sample wells, respectively. Cover with an adhesive strip and incubate for 60 min at 37°C. Aspirate each well and wash, repeating the process five times. Wash by filling each well with Wash Solution (400μl) using a squirt bottle. Add 50μl chromogen solution ‘A’ and 50μl chromogen solution ‘B’ to each well. Gently mix and incubate for 15 min at 37°C. Add 50μl Stop Solution to each well. The color in the wells should change from blue to yellow. Read the OD450 using spectrophotometer within 15 min. The concentration of AO in the samples is determined by comparing the OD450 of the samples to the standard curve.
Resistance identification and DAS-ELISA of SMV
The friction inoculation method was employed[26]. Leaves of SMV-1 and SMV-7 strains were contributed by Professor Bao Liu of Northeast Normal University. Soybean plants were grown in soil pots for 20 d prior to inoculation. Inoculums were prepared by grinding infected soybean leaf tissues with emery in 0.01 mol L-1 phosphate buffer, pH 7.0. The unrolled unifoliate leaves were inoculated by gently rubbing with the inoculums. Five soybean plants were infected with two different SMV strains for each plant line and three biological replicates were used for inoculation and sampling. Symptoms of SMV were observed after three weeks. ELISA assay was performed, which was referred to as Soybean Mosaic Virus (SMV)-DAS ELISA (AC Diagnostics, Inc., USA). The OD405 of T1 GmCnx1 transgenic plants (P) and non-transgenic plants (N) were analyzed, and if the ratio of the sample and negative control (P/N) was higher than 2, the sample plant was rendered as susceptible, while if the ratio was lower than 2 it was considered as resistant.
Results
Phylogenetic analysis and homologous sequence alignment of the GmCnx1 genes
The phylogenetic tree ( Fig 1a) was constructed by Mega software version 5.0 using the neighbor-joining method [27].As expected, Glycine maxCnx1 and Medicago truncatulaCnx1both belonging to Legume, Papillionoideae, had the closest genetic relationship. Two Moco domains in protein coded by Glycine max Cnx1 gene were found by online SMART software of EMBL (http://smart.embl-heidelberg.de/). The locations of the two domains were confirmed by DNAMAN software. A comparison of amino acid sequences ( Fig 1b) revealed that Glycine max Cnx1 shared high sequence similarity with the Cnx1 gene of Medicago truncatula, Cucumis melo, Theobroma cacao and Arabidopsis thaliana. The sequence similarity of the proteins coded by the Cnx1 genes was higher than 60% and amino acid sequences at theCnx1 conserved domains in particular were highly conserved.
Fig 1. Phylogenetic tree of GmCnx1 gene and alignment of amino acid residues from different species.

(A) Phylogenetic tree constructed by multiple sequence alignments of the Glycine max Cnx1 and those of other Cnx1 proteins. The sequences used are: Medicago truncatula Cnx1, Theobroma cacao Cnx1, Morus notabilis Cnx1, Malus domestica Cnx1, Prunus mume Cnx1, Arabidopsis thaliana Cnx1,Cucumis melo Cnx1,Hordeum vulgare Cnx1, Zea mays Cnx1, Triticum urartu Cnx1, Aegilops tauschii Cnx1, Micromonas sp Cnx1 and Micromonas pusilla Cnx1(Accession nos AET00019, EOY33170, EXB53803, XP_008385638, XP_008231326, EEH56197, XP_008451034, AAF73075, ABB30174, EMS55337, EMT03164, ACO61591 and AED92917.1, respectively). (B) Through the analysis of two Moco domains and alignment of amino acid residues, encoding protein sequences of GmCnx1, Medicago truncatula Cnx1, Cucumis melo Cnx1, Theobroma cacao Cnx1 and Arabidopsis thaliana Cnx1 had high sequence similarity.
Genetic transformation and identification of transgenic soybean plants
The pB7FWG2-GmCnx1 vector (Fig 2a) was constructed and introduced into soybean via Agrobacterium tumefaciens-mediated transformation (S1 Fig). A total of twenty seven independent putative soybean transgenic lines were generated. Ten positive lines 1, 2, 3, 4, 7, 10, 17, 18, 22 and 26 which had sufficient seeds were selected for subsequent experiments.T1putative transformants were identified by coating leaves with Phosphinothricin (Fig 2b),bar protein quick dip strip (Fig 2c) and PCR (Fig 2d). Southern blot (Fig 2e) of T1 generation revealed successful integration of the target gene into the soybean genome, which showed stable overexpression of GmCnx1. Importantly, the transgene construct was a single copy in line 2, 3, 4 and 7.
Fig 2. Recombination expression vector of pB7FWG2-GmCnx1 and determination of GmCnx1 transgenic soybean plants.

(A) Vector contained selective maker gene (bar) coded phosphinothricin acetyltransferase (PAT) and showed resistance to the herbicide phosphinothricin, green fluorescent protein gene (Egfp) and GmCnx1 gene. LB, left border; RB, right border; p35s, promoter; T35S, terminator. (B) T1 transgenic lines were confirmed by coating leaves with Phosphinothricin. (C)T1 transgenic lines were confirmed by bar protein quick dip strip. (D) T1 transgenic lines were confirmed by PCR. (E) Southern blot analysis of transgene copy number in T1 transgenic soybean and WT. Genomic DNA and plasmid DNA was digested with restriction enzyme EcoR Iand HindⅢ. The probe was used for GmCnx1. M, DL2000 marker; +, positive control (plasmid DNA); WT, negative control, non-transgenic plants; 0, blank. L1, L2, L3, L4, L7, L10, L17, L18, L22 and L26 represent the GmCnx1 transgenic line numbers.
Expression of GmCnx1in transgenic soybean lines
The expression of GmCnx1 was investigated by qRT-PCR in leaf and root under normal growth conditions. The results (Fig 3) showed that the expression of GmCnx1 gene increased from 1.04 to 2.12 fold in leaf and 1.55 to 3.89 fold in root of all transgenic positive plants in comparison to the WT, with line 10 and 22 exhibiting the highest expression levels among the different transgenic lines. These results indicated that theGmCnx1 gene expression was more significantly upregulated in root than in leaf.
Fig 3. Expression of target gene GmCnx1 in all two-week-old leaf (upper) and root (bottom) of WT and T1GmCnx1transgenic soybean plants.
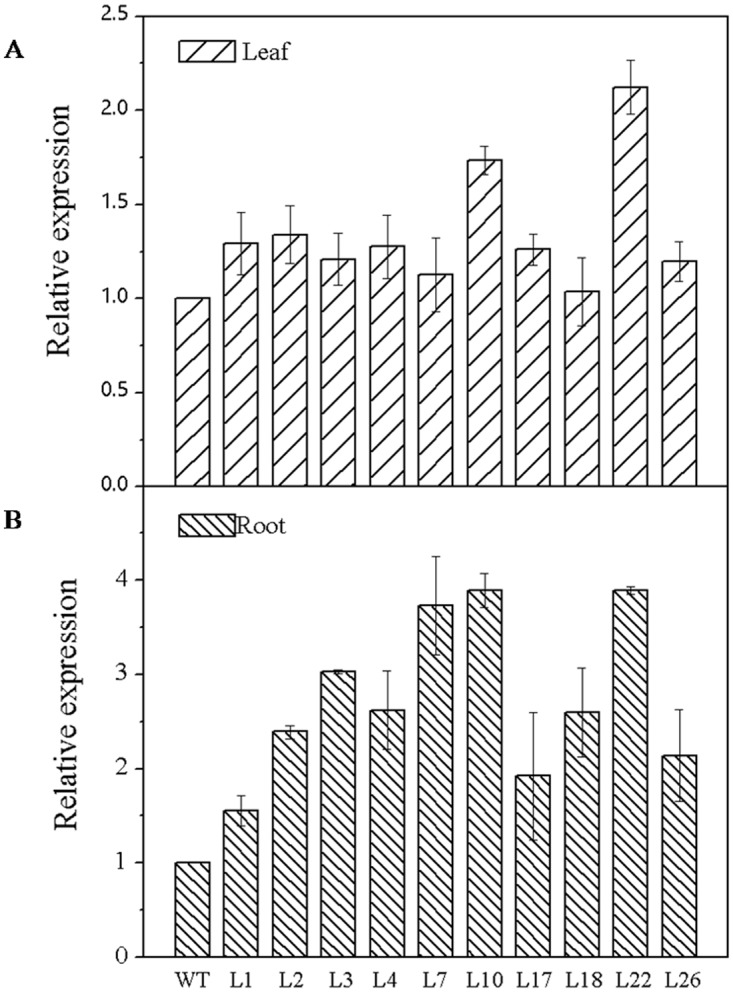
WT, negative control, non-transgenic plants. L1, L2, L3, L4, L7, L10, L17, L18, L22 and L26 represent the GmCnx1 transgenic line numbers. Values are the mean ± SD (n = 3). Error bars represent standard errors of means from three replicates.
NR and AO activity in the GmCnx1-overexpressing transgenic soybean lines
As NR and AO activity was causally linked with the biosynthesis of Moco, they were measured in leaves of WT and T1 transgenic plants. The results showed that, NR and AO activities increased in GmCnx1 transgenic plants compared with WT (Fig 4). In particular, for NR activity, the GmCnx1 transgenic soybean plants produced high level (4.25 μg g-1 h-1) in line 10, which were 2.6-fold higher than the non-transgenic plants (1.62 μg g-1 h-1), and for AO activity, the level in the leaf was 3.9-fold higher in GmCnx1 transgenic soybean plants (30.00 pmol L-1) in line 10 compared with the non-transgenic plants (7.71 pmol L-1). That was consistent with the expression of GmCnx1 gene in leaf and root. Taken together, it can be concluded that the differentGmCnx1 levels among the different transgenic lines correspond to similar differences at NR and AO activities in leaf.
Fig 4. NR (upper) and AO (bottom) activities in leaf of both WT and T1 GmCnx1transgenic soybean plants.
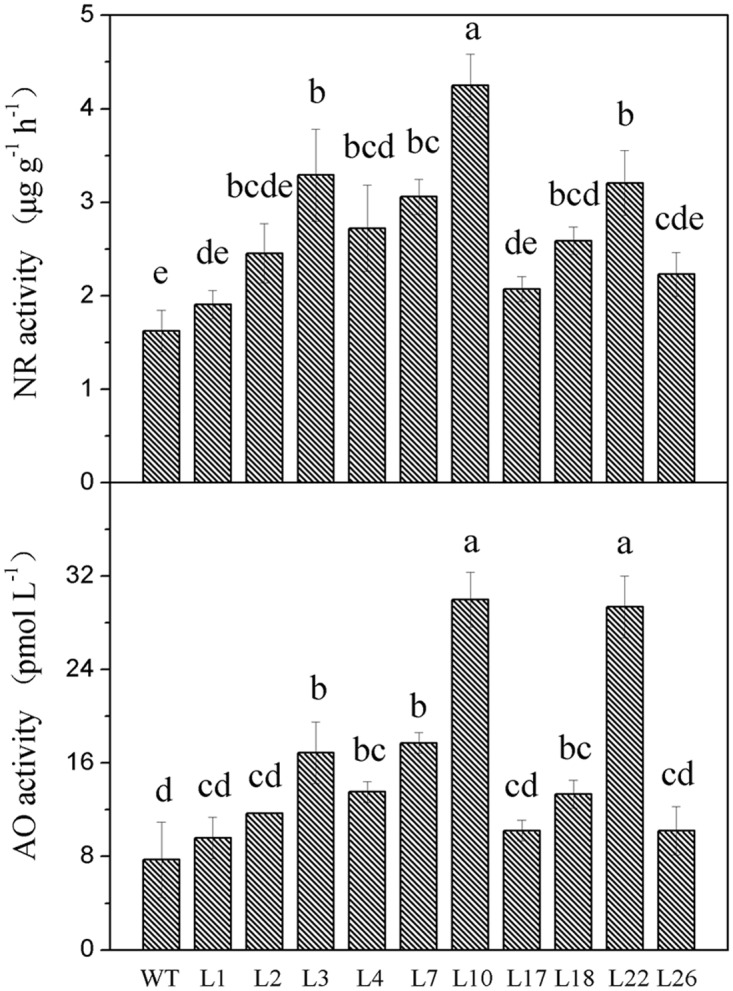
Data were calculated by significance analysis method, and the significant differences were compared with the control. WT, negative control, non-transgenic plants. L1, L2, L3, L4, L7, L10, L17, L18, L22 and L26 represent the GmCnx1 transgenic line numbers. Values are the mean ± SD (n = 3). Error bars represent standard errors of means from three replicates. The same letter at each bar means no significant difference at P< 0.05.
Resistance to SMV was increased in GmCnx1 transgenic soybeanlines
After being infected by SMV-1 and SMV-7 mosaic virus strains, non-transgenic soybean plants showed dwarfing (Fig 5a), mosaic leaves, browning veins and crimple leaves (Fig 5b). In contrast, leaves of T1 transgenic soybean plants remained green, showing no mosaic, shrinkage leaves and other symptoms. Resistance analysis by DAS-ELISA (Fig 6) showed that infection rate of GmCnx1 transgenic plants were generally lower than those of non-transgenic plants. Specifically, for SMV-1, the infection rates of non-transgenic and10 lines of GmCnx1 transgenic plants were 87%, 47%, 40%, 33%, 53%, 33%, 33%, 47%, 40%, 33% and 47%, respectively. For SMV-7, the infection rates of non-transgenic and 10 lines of GmCnx1 transgenic plants were 93%, 40%, 53%, 47%, 40%, 33%, 40%, 47%, 33%, 33% and 40%, respectively. The results above indicated that the GmCnx1 transgenic soybean lines we generated showed higher degrees of resistance to two SMV strains tested.
Fig 5. Comparison of SMV resistance in soybean three weeks after friction inoculation.

(A) Growth performance of WT and T1 GmCnx1 transgenic plants before and after infection. (B) Symptoms on tender leaves (top second leaf) of WT and T1 GmCnx1 transgenic plants before and after infection. WT, negative control, non-transgenic plants.
Fig 6. The infection rate of SMV-1 (upper) and SMV-7 (bottom) in both non-transgenic and GmCnx1 transgenic soybean plants by DAS-ELISA.
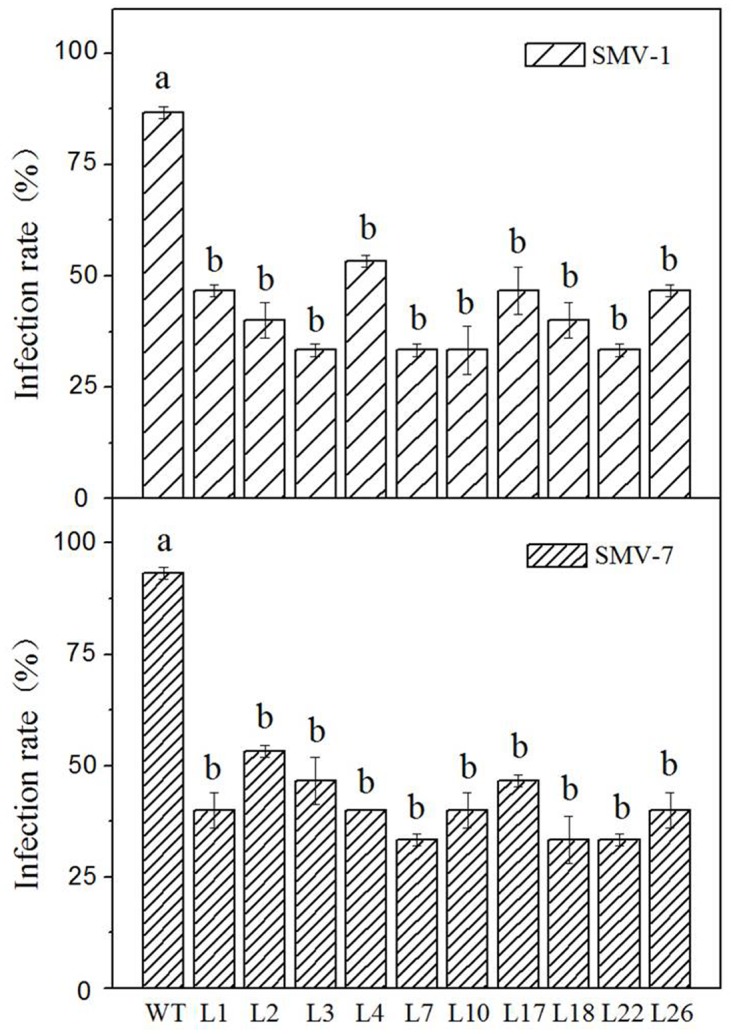
WT, negative control, non-transgenic plants. L1, L2, L3, L4, L7, L10, L17, L18, L22 and L26 represent the GmCnx1 transgenic line numbers. Values are the mean ± SD (n = 3). Error bars represent standard errors of means from three replicates. The same letter at each bar means no significant difference at P< 0.05.
Discussion
Although soybean is one of the most important food and feed crops worldwide, its transformation efficiency is relatively low. In comparison with other crops, there are still several problems that need to be tackled such as difficulties in the regeneration potential of the tissues, low reproducibility, and greater genotypic differences[28].In the light of these issues, we optimized several physical and chemical parameters of the standard Agrobacterium tumefaciens-mediated transformation protocol for soybean, and obtained significantly improved results [29].Here, in soybean, we successfully generated transgenic soybean plants for the GmCnx1 gene, validated the previous function of the gene in enhancing NR activity and unveiled the novel function of the gene in enhancing AO activity and SMV resistance.
Cnx1 is a functional protein with two domains, which are essential for catalyzing the insertion of Mo into MPT in plants [30]. E and G domains that were detected in Arabidopsis were involved in the activity of Cnx1 [14]. In the present study, the comparison of amino acid sequences of Cnx1 between Arabidopsis and soybean shows two highly conserved amino acid motifs involved in biosynthesis of molybdopterin cofactor present in soybean (Fig 1b). Moreover, these two domains were suggested in binding with molybdopterin and form an alpha/beta structure. So it was predicted that these two domains would interact with each other for the function of GmCnx1, thus catalyzing the incorporation of Mo into MPT in soybean.
According to Mendel et al. [31], the third step in the biosynthesis of Moco is catalyzed by the two domain-containing proteins Cnx1 (Cnx1-G, Cnx1-E). Mature Moco can bind to NR and AO. Moco-deficient plant mutants showed a pleiotropic loss of all four Mo-enzymes activities NR, AO, XDH and SO. Moco-mutants have been described in numerous higher plants, for example, in tobacco [32, 33], Nicotiana plumbaginigolia [34] and barley [35].Cnx1 expression was found in all organs of Arabidopsis plants, and it was noted that transgenic Cnx1gene could increase activity of NR and content of MPT [14]. Moreover, blockage of Moco biosynthesis due to mutations led to the impairment of essential metabolic functions because of the loss of functions of all Mo-dependent enzymes, which ultimately led to death of the organism [36]. On the other hand, there is paucity of literature regarding AO activities in plants. Early reports showed in potato the presence of two isoenzymes for AO that are able to use diverse aldehydes but not xanthine as substrates [37]. The reduction of ABA levels due to the lack of AO activity had a direct impact on Moco-sulfurase deficiency [38,39]. A barley cnx mutant showed a thermo sensitive, wilty phenotype, which was attributed to an ABA deficiency due to the absence of activity of a Moco-dependent AO[40].We found that overexpression of GmCnx1 gene indeed enhanced activities of NR and AO in the transgenic soybean plants (Figs 3 and 4), consistent with previous studies in other plants. In addition, for evaluation of agronomic traits, three transgenic and three non-transgenic soybean plants were randomly sampled before flowering stage from the experimental greenhouse. Plant height, average leaf length, average leaf width, nodules number, fresh weight of nodules, root and shoot fresh weight, root and shoot dry weight and total biomass were measured (S1 Table).
SMV seriously affects yield and quality of soybean globally. Due to its wide distribution, great harmfulness, and difficulties in chemical control, cultivating and planting resistant varieties is an economical, safe and effective way to prevent SMV. In the present study we aimed at checking the relationship between GmCnx1 gene and its function in plants with regards to SMV resistance. Based on a microarray analysis of gene expression differences between two contrasting genotypes (SMV resistant and SMV susceptible) upon SMV inoculation, GmCnx1 was selected as a candidate SMV resistant gene [17]. We therefore deduced that the GmCnx1 gene expression in soybean is probably related to the positive regulation on the resistance to SMV. In this study, SMV-1 and SMV-7 were used to test for plant resistance to SMV. Consistent with the results from the microarray experiments, our results showed that transgenic plants indeed have higher SMV resistance compared with non-transgenic plants after the virus infection (Fig 6). We suspect that resistance to SMV might be due to the increase in NR and AO activities which are enhanced by the overexpression of GmCnx1 gene in soybean (Fig 7). In any case, this is the first report about the novel function of GmCnx1 against SMV, which has apparent breeding implications in soybean.
Fig 7. Showing the relationship among GmCnx1 gene, molybdenum enzymes and SMV.

Cnx1, Cnx2, Cnx3, Cnx6 and Cnx7 are involved in Moco biosynthesis. Solid arrows represent functions of GmCnx1 observed in this present work while dotted arrows represent future research prospective and open arrows show Moco-dependent enzymes that have been published in other research work [2,3].
Follow-up studies are required to elucidate the underlying mechanism of GmCnx1 function in response to NR, AO activities and SMV resistance, and in particular, the exact role of AO activity in post-invasive cell wall defense to SMV. Here, we observed that GmCnx1 gene enhanced SMV resistance of plants, but whether Cnx2, Cnx3, Cnx6 and Cnx7, which function in the initial steps of Moco biosynthesis (Fig 7), had similar effects on SMV resistance remain unknown.
In conclusion, we show that overexpression of GmCnx1 gene enhanced NR, AO activities and SMV resistance in soybean. This report provided valuable information on molecular breeding and improved our understanding of the biological and molecular functions of GmCnx1. These data provide a novel route for initiating molecular breeding to improve NR, AO activities and SMV resistance in soybean, a significant crop worldwide.
Supporting Information
(A) Seed sterilization by chlorine gas method. (B) Seed germination on GM for 1 d. (C) Agrobacterium tumefaciens infection in LCCM for 30 min. (D) Co-cultivation explants with Agrobacterium for 4 d. (E) Multiple shoot induction on SI after two weeks. (F) Elongation of multiple shoot on SE after at least two weeks. (G) Elongation of multiple shoots. (H) Rooting on RM. (I) Plantlet domestication in greenhouse. (J) Plantlet transplanting and seed pod in greenhouse.
(TIFF)
(XLSX)
Acknowledgments
The authors thank Prof. Xinan Zhou (Oilcrops Research Institute, Chinese Academy of Agricultural Sciences) for providing soybean cultivar, Tianlong 1.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
Support was provided by the National Natural Science Foundation of China (no. 31071443), [http://www.nsfc.gov.cn/] and the National Major Special Project for Transgenic Organisms, Ministry of Agriculture in China (nos. 2013ZX08004-004-008 and 2014ZX08004-004-008), [http://www.ny3721.com/url/2805/]. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Hille R (1996) The mononuclear molybdenum enzymes. Chem Rev 96(7):2757–2816. [DOI] [PubMed] [Google Scholar]
- 2. Mendel RR, Schwarz G (2002) Biosynthesis and molecular biology of the molybdenum cofactor (Moco). Met Ions in Biol Syst 39: 317–368. [PubMed] [Google Scholar]
- 3. Mendel RR, Schwarz G (2011) Molybdenum cofactor biosynthesis in plants and humans. Coordin Chem Rev 255: 1145–1158. [Google Scholar]
- 4. Bittner F, Mendel RR (2010) Cell biology of molybdenum. Cell Biology of Metals and Nutrients, Plant Cell Monographs 17: 119–143. [Google Scholar]
- 5. Campbell WH (2001) Structure and function of eukaryotic NAD(P)H: nitrate reductase. Cell. Mol. Life Sci 58(2): 194–204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Crawford NM, Forde BG (2002) Molecular and developmental biology of inorganic nitrogen nutrition. The Arabidopsis book 1: e0011 10.1199/tab.0011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Campbell WH (1999) Nitrate reductase structure, function, and regulation. Bridging the gap between biochemistry and physiology. In: Jones RL, Bohnert HJ, Walbot V, editors. Annual Review of Plant Physiology and Plant Molecular Biology 50(1): 277–303. [DOI] [PubMed] [Google Scholar]
- 8. Koshiba T, Saito E, Ono N, Yamamoto N, Sato M (1996) Purification and properties of flavin-and molybdenum containing aldehyde oxidase from coleoptiles of maize. Plant Physiol 110(3): 781–789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Seo M, Koshiba T (2002) Complex regulation of ABA biosynthesis in plants. Trends in Plant Science 7(1): 41–48. [DOI] [PubMed] [Google Scholar]
- 10. Mauch-Mani B, Mauch F (2005) The role of abscisic acid in plant-pathogen interactions. Current Opinion in Plant Biology 8(4): 409–414. [DOI] [PubMed] [Google Scholar]
- 11. Verslues PE, Zhu JK (2005) Before and beyond ABA: upstream sensing and internal signals that determine ABA accumulation and response under abiotic stress. Biochemical Society Transactions 33(Pt 2): 375–379. [DOI] [PubMed] [Google Scholar]
- 12. Mendel RR, Hänsch R (2002) Molybdoenzymes and molybdenum cofactor in plants. J Exp Bot 53(375): 1689–1698. [DOI] [PubMed] [Google Scholar]
- 13. Yesbergenova Z, Yang G, Oron E, Soffer D, Fluhr R, Sagi M, et al. (2005) The plant Mo-hydroxylases aldehyde oxidase and xanthine dehydrogenase have distinct reactive oxygen species signatures and are induced by drought and abscisic acid. Plant J 42(6): 862–876. [DOI] [PubMed] [Google Scholar]
- 14. Schwarz G, Schulze J, Bitter F, Eilers T, Kuper J, Bollmann G, et al. (2000) The molybdenum cofactor biosynthetic protein Cnx1 complements molybdate-repairable mutants, transfers molybdenum to the metal binding pterin, and is associated with the cytoskeleton. Plant Cell 12(12): 2455–2471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Yang SH, Berberich T, Miyazaki A, Sano H, Kusano T (2003) Ntdin, a tobacco senescence-associated gene, is involved in molybdenum cofactor biosynthesis. Plant Cell Physiol 44(10): 1037–1044. [DOI] [PubMed] [Google Scholar]
- 16. Li YJ, Zhang JC, Zhang J, Hao L, Hua JP, Duan LS, et al. (2013) Expression of an Arabidopsis molybdenum cofactor sulphurase gene in soybean enhances drought tolerance and increases yield under field conditions. Plant Biotechnol J 11(6): 747–758. 10.1111/pbi.12066 [DOI] [PubMed] [Google Scholar]
- 17. He HL (2011) Frog leaf spot and soybean mosaic virus disease resistance gene screening, cloning and preliminary analysis of the genesMaster Dissertation, Northeast Normal University, Changchun, China. [Google Scholar]
- 18. Zhang ZY, Xing AQ, Staswick P, Clemente TE (1999) The use of glufosinate as a selective agent in Agrobacterium-mediated transformation of soybean. Plant Cell Tiss Org 56(1): 37–46. [Google Scholar]
- 19. Paz MM, Martinez JC, Kalvig AB, Fonger TM, Wang K (2006) Improved cotyledonary node method using an alternative explant derived from mature seed for efficient Agrobacterium-mediated soybean transformation. Plant Cell Rep 25(3): 248. [DOI] [PubMed] [Google Scholar]
- 20. Hoagland DR, Arnon DI (1950) The water-culture method for growing plants without soil. Calif AES 347(2): 1–32. [Google Scholar]
- 21. Wang XD, Lu HY, Zhang J, Zhu BG (2004) Comparative study on methods of extracting DNA from soybean leaf for PCR. Molecular Plant Breeding 2(6): 891–894. [Google Scholar]
- 22. Edwards K, Johnstone C, Thompson C (1991) A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucleic Acids Res 19(6): 1349 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Xue ZG, Zhang XM, Lei CF, Chen XJ, Fu YF (2012) Molecular cloning and functional analysis of one ZEITLUPE homolog GmZTL3 in soybean. Mol Biol Rep 39(2): 1411–1418. 10.1007/s11033-011-0875-2 [DOI] [PubMed] [Google Scholar]
- 24. Schmittgen TD, Livak KJ (2008) Analyzing real-time PCR data by the comparative C T method. Nat Protoc 3(6): 1101–1108. [DOI] [PubMed] [Google Scholar]
- 25. Sagi M, Lips HS (1998)The levels of nitrate reductase and MoCo annual ryegrass as affected by nitrate and ammoinium nutrition. Plant Sci 135: 17–24. [Google Scholar]
- 26. Yang XF, Yang YQ, Zheng GJ (2013)Fine mapping of resistance genes to SMV strains SC6 and SC17 in soybean. Acta Agronomica Sinica 39(2): 216–221. [Google Scholar]
- 27. Tamura K, Peterson D, Peterson N, Stecher G. Nei M, Kumar S (2011) MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28(10): 2731–2739. 10.1093/molbev/msr121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Hinchee MAW, Connor-Ward DV, Newell CA, McDonnell RE, Sato SJ, Gasser CS, et al. (1988) Production of transgenic soybean plants using Agrobacterium-mediated DNA transfer. Nat Biotechnol 6(8): 915–922. [Google Scholar]
- 29. Yang XF (2011) Optimization of Agrobacterium tumefacien-mediated soybean transformation system using cotyledoonary explants and developing of transgenic soybean with improved insect-resistant the Sporamin and Chitinase KDEL gene transformation study Master Dissertation, Zhejiang University, Hangzhou, China. [Google Scholar]
- 30. Stallmeyer B, Nerlich A, Schiemann J, Brinkmann H, Mendel RR (1995) Molybdenum co-factor biosynthesis: The Arabidopsis thaliana cDNA cnx1 encodes a multifunctional two-domain protein homologous to a mammalian neuroprotein, the insect protein Cinnamon and three Escherichin coli proteins. Plant J 8(5):751–762. [DOI] [PubMed] [Google Scholar]
- 31. Mendel RR (2007) Biology of the molybdenum cofactor. J Exp Bot 58(9): 2289–2296. [DOI] [PubMed] [Google Scholar]
- 32. Mendel RR, Müller AJ (1976) A common genetic determinant of xanthine dehydrogenase and nitrate reductase in Nicotiana tabacum . Biochemie and Physiologie der Pflanzen 170: 538–541. [Google Scholar]
- 33. Mendel RR, Müller AJ (1978) Reconstitution of NADH-nitrate reductase in vitro from nitrate reductase-deficient Nicotiana tabacum mutants. Molecular and General Genetics 161(1): 77–80. [Google Scholar]
- 34. Müller AJ, Mendel RR (1989) Biochemical and somatic cell genetics of nitrate reductase in Nicotiana In: Wray JL, Kinghorn JR, editors. Molecular and genetic aspects of nitrate assimilation. Oxford: Oxford University Press; pp. 166–185. [Google Scholar]
- 35. Kleinhofs A, Warner RL, Lawrence JM, Jeter JM, Kudrna DA (1989) Molecular genetics of nitrate reductase in barley In: Wray JL, Kinghorn JR, editors. Molecular and genetic aspects of nitrate assimilation. Oxford: Oxford University Press; pp.197–211. [Google Scholar]
- 36. Schwarz G, Mendel RR (2006) Molybdenum cofactor biosynthesis and molybdenum enzymes. Annu Rev Plant Biol 57: 623–647. [DOI] [PubMed] [Google Scholar]
- 37. Rothe GM (1974) Aldehyde oxidase isoenzymes (E.C. 1.2.3.1) in potato tubers (Solanum tuberosum). Plant Cell Physiol 15(3): 493–499. [Google Scholar]
- 38. Léon-Kloosterziel KM, Gil MA, Ruijs GJ, Jacobsen SE, Olszewski NE, et al. (1996) Isolation and characterization of abscisic acid-deficient Arabidopsis mutants at two new loci. Plant J 10(4): 655–661. [DOI] [PubMed] [Google Scholar]
- 39. Xiong L, Ishitani M, Lee H, Zhu JK (2001) The Arabidopsis LOS5/ABA3 locus encodes a molybdenum cofactor sulfurase and modulates cold stress—and osmotic stress—responsive gene expression. Plant Cell 13(9): 2063–2083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Walker-Simmons M, Kudrna DA, Warner RL (1989) Reduced accumulation of ABA during water stress in a molybdenum cofactor mutant of barley. Plant Physiol 90(2): 728–733. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(A) Seed sterilization by chlorine gas method. (B) Seed germination on GM for 1 d. (C) Agrobacterium tumefaciens infection in LCCM for 30 min. (D) Co-cultivation explants with Agrobacterium for 4 d. (E) Multiple shoot induction on SI after two weeks. (F) Elongation of multiple shoot on SE after at least two weeks. (G) Elongation of multiple shoots. (H) Rooting on RM. (I) Plantlet domestication in greenhouse. (J) Plantlet transplanting and seed pod in greenhouse.
(TIFF)
(XLSX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.