

Abstract
Background
Gastropod-borne parasites may cause debilitating clinical conditions in animals and humans following the consumption of infected intermediate or paratenic hosts. However, the ingestion of fresh vegetables contaminated by snail mucus and/or water has also been proposed as a source of the infection for some zoonotic metastrongyloids (e.g., Angiostrongylus cantonensis). In the meantime, the feline lungworms Aelurostrongylus abstrusus and Troglostrongylus brevior are increasingly spreading among cat populations, along with their gastropod intermediate hosts. The aim of this study was to assess the potential of alternative transmission pathways for A. abstrusus and T. brevior L3 via the mucus of infected Helix aspersa snails and the water where gastropods died. In addition, the histological examination of snail specimens provided information on the larval localization and inflammatory reactions in the intermediate host.
Methodology/Principal Findings
Twenty-four specimens of H. aspersa received ~500 L1 of A. abstrusus and T. brevior, and were assigned to six study groups. Snails were subjected to different mechanical and chemical stimuli throughout 20 days in order to elicit the production of mucus. At the end of the study, gastropods were submerged in tap water and the sediment was observed for lungworm larvae for three consecutive days. Finally, snails were artificially digested and recovered larvae were counted and morphologically and molecularly identified. The anatomical localization of A. abstrusus and T. brevior larvae within snail tissues was investigated by histology. L3 were detected in the snail mucus (i.e., 37 A. abstrusus and 19 T. brevior) and in the sediment of submerged specimens (172 A. abstrusus and 39 T. brevior). Following the artificial digestion of H. aspersa snails, a mean number of 127.8 A. abstrusus and 60.3 T. brevior larvae were recovered. The number of snail sections positive for A. abstrusus was higher than those for T. brevior.
Conclusions
Results of this study indicate that A. abstrusus and T. brevior infective L3 are shed in the mucus of H. aspersa or in water where infected gastropods had died submerged. Both elimination pathways may represent alternative route(s) of environmental contamination and source of the infection for these nematodes under field conditions and may significantly affect the epidemiology of feline lungworms. Considering that snails may act as intermediate hosts for other metastrongyloid species, the environmental contamination by mucus-released larvae is discussed in a broader context.
Author Summary
Gastropod-borne parasites may cause debilitating clinical conditions in animals and humans. The infection occurs by consumption of intermediate hosts (i.e., snails). However, the ingestion of fresh vegetables contaminated by water and/or snail mucus has been proposed as a transmission source for some zoonotic metastrongyloids (e.g., Angiostrongylus cantonensis). Aelurostrongylus abstrusus and Troglostrongylus brevior cause severe clinical conditions in cats and their prevalence is increasing in feline populations. The epidemiology of feline lungworm infections is poorly understood and the role of gastropod molluscs, as vectors of the disease, is questioned. Since metastrongyloid nematodes may use other little-studied infection pathways, the presence of A. abstrusus and T. brevior larvae was herein assessed in the mucus of Helix aspersa snails and in the water where specimens died. The finding of A. abstrusus and T. brevior infective L3 in the mucus shed by H. aspersa or when infected gastropods die submerged in water indicates that alternative pathways for the transmission of feline lungworms may occur. Accordingly, these results should spur the interest of the scientific community towards the delineation of further infection routes for other metastrongyloids of major human concern, such as Angiostrongylus costaricensis and A. cantonenis.
Introduction
Gastropod-borne agents infect approximately 300 million people worldwide, causing major debilitating conditions, as well as adversely affecting quality of life and healthcare. For example, some ailments may contribute to the development of neoplastic tumours, such as cholangiocarcinoma during opisthorchiosis/clonorchiosis [1] or may induce debilitating diseases (e.g., schistosomiosis or fascioliosis) [2,3]. In addition, human beings may act as dead-end hosts for zoonotic metastrongyloids of rats, such as Angiostrongylus cantonensis or Angiostrongylus costaricensis (Strongylida, Angiostrongylidae), which cause eosinophilic meningitis and ileitis, respectively [4,5]. The main pathway for human infection is represented by the consumption of undercooked intermediate hosts (i.e., snails) or raw paratenic hosts (e.g., prawns and freshwater shrimps) [6,7].
In veterinary medicine, the lungworm Aelurostrongylus abstrusus (Strongylida, Angiostrongylidae), along with the less-known Troglostrongylus brevior (Strongylida, Crenosomatidae) are of increasing concern due to their spreading in cat populations [8–10]. Indeed, these parasites share a similar biology and ecological niches, and may infect the same individual [10,11]. Although gastropod molluscs are recognized as the intermediate hosts for feline lungworms [12], the role of snails in the transmission of the infection is debatable because they are not preferred preys for felids [13,14]. As a result, paratenic hosts (i.e., rodents, birds, amphibians and reptiles) may play a crucial role in the epidemiology of A. abstrusus [15,16]. Metastrongyloid nematodes may, however, use additional routes of the infection, as it has been hypothesized for the vertical transmission of T. brevior in cats [17].
Land snails excrete a thin layer of pedal mucus, consisting of water and mucin-like carbohydrate-protein complexes, which acts as glue and lubricant during the locomotion [18]. Again, the production of the mucous trail facilitates several functions of the snail, such as the homing behaviour, aggregation, and protection against desiccation and predation [18]. Interestingly, gastropods may be able to release mucous secretions of variable quality, depending on the nature of the external stimuli (i.e., mechanical or chemical) [19]. The ingestion of fresh vegetables, contaminated by water and/or snail mucus harbouring A. cantonensis third-stage larvae (L3), has been previously suggested in a recent outbreak of human eosinophilic meningitis in Jamaica [20]. Accordingly, the presence of A. cantonensis and A. costaricensis in the mucus of slugs or snails and in contaminated water has been investigated, but their role in the epidemiology of the infection is yet to be confirmed [7].
This study sought to establish whether A. abstrusus and T. brevior L3 could be found in the mucus of infected Helix aspersa snails and in the water where specimens had died. Larval localization in snails and inflammatory reactions in the intermediate hosts has also been investigated by histology.
Materials and Methods
Snail maintenance and infection
Adult H. aspersa (n = 200) were sourced from a commercial farming centre in Barletta (southern Italy). The absence of natural metastrongyloid infections was confirmed by artificially digesting and examining a representative number (i.e., 10%) of snails. During the maintenance, gastropods were placed in plastic boxes filled with natural soil, covered with a net and kept at 23±1°C. Snails were fed every two days with lettuce ad libitum. First-stage larvae of A. abstrusus and T. brevior used for infection were obtained from cats artificially infected with pure isolates, originating from Hungary and Italy, respectively. Two groups of 50 snails each were prepared, being infected with ~500 L1 of A. abstrusus and T. brevior, respectively, as described in [21]. Snails were then placed into two different boxes, according to lungworm species, and two specimens from each group were artificially digested at +6, +12 and +18 days post infection (dpi) to evaluate the success of the infection.
Elimination of third-stage larvae via mucus
In order to assess whether larvae are released in the mucus of snails, six groups (G1–G6) of 4 snails each were formed 20 dpi for each metastrongyloid species. Snails were placed into a 1 l plastic box for bacteriological use, together with 25 ml of tap water, and subjected to different stimuli. Specimens in G1 and G4 were left with water, those in G2 and G5, and G3 and G6 were fed with lettuce (3 g) or commercial cat food (0.8 g), respectively. Group 1, G2 and G3 were placed on a thermostat shaker, applying approximately 50 oscillations per minute overnight, while G4, G5 and G6 were constantly kept without mechanical stimuli. All snails were housed under controlled room temperature (23±1°C). Once a day in the morning for 20 days, snails were taken out from their plastic boxes, which were rinsed with 20 ml of tap water (30°C), and the solution was filtered through a 180-μm meshed sieve. The filtrate was centrifuged at 600 g for 5 min and the pellet examined under a light microscope. Finally, snails were returned to the same experimental group boxes. Larvae recovered were quantified and identified using appropriate morphological keys [15,21].
Release of third-stage larvae in water
In order to evaluate the emergence of larvae from dead submerged snails, at the end of the experiment described above, gastropods were identified with a letter, placed into a 50 ml tube, submerged in tap water and left at room temperature (23±1°C). Every 24 h for three consecutive days, the sediment was collected and centrifuged at 600 g for 5 min, and the pellet was examined under a light microscope. Nematode larvae were considered motile if they were not damaged and were moving within 10 s [22].
Artificial digestion and larval molecular identification
Each snail specimen was artificially digested in a solution of 1% HCl (100 ml) and 1.2 g powdered pepsin (Sigma-Aldrich, St. Louis, Missouri, United States), as described elsewhere [21]. The solution was collected in plastic tubes and centrifuged at 600 g for 5 min. The whole suspension (5 ml) was examined under a light microscope; larvae were morphologically identified according to species and developmental stage and counted.
Ten representative larval specimens of each of A. abstrusus and T. brevior were isolated from the sediment of digested snail tissues and stored in plastic vials, containing phosphate buffer saline (PBS) at −20°C to be analysed by PCR. DNA was extracted using the DNeasy Blood & Tissue Kit (Qiagen, GmbH, Hilden, Germany), in accordance with the manufacturer instructions, and a duplex-PCR was performed, using species-specific forward primer sets, amplifying ITS-2 region of different size (i.e., A. abstrusus: 220 bp; T. brevior: 370 bp) [11]. Sequences were determined from both strands and compared with those available in the GenBank database by Basic Local Alignment Search Tool (BLAST).
Statistical analysis
The arithmetic mean of L3 counted in the mucus, in water and in sediment following artificial digestion was calculated for each group. Larval counts obtained from these three experiments were totalled for each snail and the infection rate was calculated using the following formula: infection percentage = (500—T)/500, where T was the total number of L3 recovered. Accordingly, the values were compared between A. abstrusus and T. brevior infected snails using the Mann-Whitney U-Test. The statistical analysis was two-sided, at the significance level p = 0.05.
Histological examination of snails
To assess the anatomical localization of metastrongyloid larvae, two infected snails were examined every three days, from +1 to +30 dpi (i.e., 11 time points). Snails were anesthetized with menthol steam in a plastic box for 3–5 hours and, as soon as the foot was completely extended and insensitive to touch, they were deprived of their shells and fixed in a 50 ml vial with 10% neutral buffered formalin solution, to be histologically examined. Longitudinal sections across the middle of the body and parasagittal sections through the coiled part of the snail were routinely processed, embedded in paraffin and 5 μm slices were stained with haematoxylin and eosin (H&E). The presence of a tissue inflammatory response around larvae was also recorded.
Results
No larval nematodes were recovered from H. aspersa specimens digested before the infection. Conversely, larval nematodes were recovered from all snails experimentally infected with either A. abstrusus or T. brevior (n = 6 for each parasite) at each sampling point.
Third-stage larvae of both feline lungworm species were detected in the mucus of snails in all groups, except G1; only a single L2 of T. brevior was recovered from G4 at 20 dpi (Table 1). In particular, out of 37 A. abstrusus (mean 6.2±12.4) and 19 T. brevior (mean 3.2±7.3) L3 recovered, the highest number of larvae was observed in G2 and G5 for both lungworm species. Overall, 7.8% and 1.8% of A. abstrusus and T. brevior L3, respectively, were recovered in the mucus without significant difference recorded in the total number of L3 between A. abstrusus and T. brevior groups (p>0.05).
Table 1. Group and total number of Aelurostrongylus abstrusus and Troglostrongylus brevior larvae detected in the mucus of Helix aspersa snails or in the water solution where gastropods had died.
| Larvae in the mucus | Larvae in water | |||
|---|---|---|---|---|
| Groups (Stimulus) | A. abstrusus | T. brevior | A. abstrusus | T. brevior |
| G1 (Water and stirring) | - | - | 18 | 4 |
| G2 (Lettuce and stirring) | 9 | 12 | 12 | 4 |
| G3 (Cat food and stirring) | 2 | 1 | 27 | 14 |
| G4 (Water) | 7 | 1* | 18 | 6 |
| G5 (Lettuce) | 13 | 4 | 34 | 0 |
| G6 (Cat food) | 6 | 2 | 63 | 11 |
| Total | 37 | 20 | 172 | 39 |
*Second-stage larva
In snails that had died through submersion, larvae of A. abstrusus (n = 172; mean 7.2±12.2) and T. brevior (n = 39; mean 1.6±2.6) were detected in the sediment of 21/24 (87.5%) and 10/24 (41.7%) specimens, respectively (Tables 1 and 2); the mean number of A. abstrusus L3 was significantly higher (p = 0.00076). In total, 5.7% A. abstrusus and 2.7% T. brevior L3 were found in the water (Table 1).
Table 2. Total number and percentage of Aelurostrongylus abstrusus and Troglostrongylus brevior L3 detected in each snail specimens from different groups following water flooding and digestion examinations.
| A. abstrusus | T. brevior | ||||||
|---|---|---|---|---|---|---|---|
| Groups (Stimulus) | Specimen | Flooding | Digestion | Total L3 | Flooding | Digestion | Total L3 |
| G1 (Water andstirring) | A | - | 271 | 271 | 4 | 17 | 21 |
| B | 6 | 72 | 78 | - | 30 | 30 | |
| C | 8 | 98 | 106 | - | 170 | 170 | |
| D | 4 | 78 | 82 | - | 47 | 47 | |
| G2 (Lettuce andstirring) | A | 1 | 66 | 67 | - | 98 | 98 |
| B | 7 | 216 | 223 | - | 56 | 56 | |
| C | 3 | 184 | 187 | - | 158 | 158 | |
| D | 1 | 14 | 14 | 4 | 50 | 54 | |
| G3 (Cat foodand stirring) | A | 7 | 156 | 163 | 8 | 68 | 76 |
| B | 12 | 91 | 103 | - | 39 | 39 | |
| C | 3 | 23 | 26 | 6 | 105 | 111 | |
| D | 5 | 98 | 103 | - | 43 | 43 | |
| G4 (Water) | A | 5 | 386 | 391 | - | 50 | 50 |
| B | 10 | 98 | 108 | 3 | 71 | 74 | |
| C | 2 | 73 | 75 | 3 | 55 | 58 | |
| D | 1 | 19 | 20 | - | 41 | 41 | |
| G5 (Lettuce) | A | 1 | 67 | 68 | - | 55 | 56 |
| B | 20 | 95 | 115 | - | 64 | 64 | |
| C | 5 | 77 | 82 | - | 10 | 10 | |
| D | 8 | 145 | 151 | - | 30 | 30 | |
| G6 (Cat food) | A | 2 | 323 | 325 | 1 | 19 | 20 |
| B | 60 | 49 | 109 | 2 | 39 | 41 | |
| C | - | 86 | 86 | - | 57 | 57 | |
| D | 1 | 73 | 74 | 8 | 18 | 26 | |
| Total | 172 (5.7%) | 2858 (94.3%) | 3030 | 39 (2.7%) | 1390 (97.3%) | 1429 | |
Following the artificial digestion, all H. aspersa snails scored positive for metastrongyloid larvae. A total of 2858 A. abstrusus and 1390 T. brevior L3 were recovered and individual larval counts are reported in Table 2. The mean number of L3 was 127.8 (min 14–max 386) for A. abstrusus and 60.3 (min 10–max 170 L3) for T. brevior. Taking into account the original 500 L1 administered to each snail and the total number of larvae recovered, the proportion of L1 that had moulted to L3 was 25.6% for A. abstrusus and 12.1% for T. brevior (see statistical analysis paragraph). All nematodes detected were actively motile, and all specimens were morphologically identified as infective L3, since they had lost their outer sheaths and measured 442.7±17.8 μm (i.e., T. brevior and Fig 1A) and 548.6±30.3 μm (i.e., A. abstrusus and Fig 1B). This identification was confirmed by molecular amplification and sequencing of partial ITS-2 gene. Nucleotide sequences, examined by BLAST, displayed a 100% homology with those of A. abstrusus and T. brevior in GenBank (accession numbers KF751656 and KF751655).
Fig 1. Infective L3 of T. brevior.

(A) and A. abstrusus (B) detected in the mucus of H. aspersa snails at 25 days post-infection (scale bar = 50μm).
During the histological examination, larvae of metastrongyloids were found at different time points, starting from 1 dpi and 9 dpi for A. abstrusus and T. brevior, respectively. Larval transverse sections were recognized as belonging to A. abstrusus or T. brevior according to their mean diameter, detected on round bodies, measuring ~25 and 20 μm, respectively. The number of snail sections positive for A. abstrusus (n = 103) was higher than that for T. brevior (n = 16), with a mean number of 6.4 for the former and 1.0 for the latter species. For both metastrongyloids, larvae were mainly observed in the anterior and posterior parts of fibro-muscular tissue (Fig 2A) of the foot and in the skirt, close to the pedal and oesophageal glands. Larvae were randomly detected in other organs, such as the kidney parenchyma, the wall of the pallial cavity and the connective sub-epithelial layer of the intestine (Fig 2B). Nonetheless, free larvae were rarely found in infected snails at different time points (i.e., at 1, 12 and 21 dpi for A. abstrusus and at 9 dpi for T. brevior). Specimens were localized in the fibro-muscular tissue of the foot, with some of them near vessels, and one specimen in the coelom. All individuals were separated from the surrounding tissue by a thin optically empty space.
Fig 2. Histopathology: Free larvae of A. abstrusus (21 dpi) in snail foot observed in transverse (hash) and oblique sections (star) in the fibro-muscular tissues.
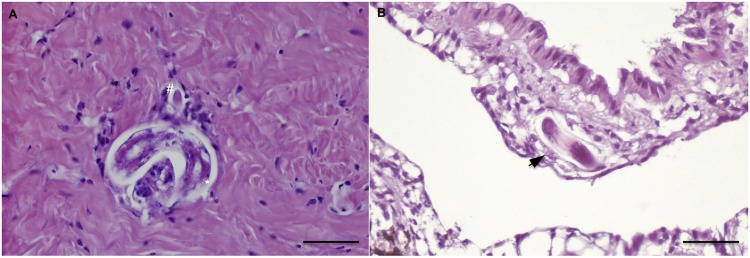
(A); oblique larval section of T. brevior (9 dpi) the subpallial tissue (arrow) (B) (scale bar = 50μm; H&E).
The tissue response to nematode larvae included: i) cell-poor (3 dpi; Fig 3A) and cell-rich (9 dpi; Fig 3B) granuloma-like formations, composed of non-vacuolated or vacuolated epithelioid amebocytes; ii) small necrotic granulomas (15 dpi; Fig 3C); iii) fibroblast-like encapsulations (27 dpi; Fig 3D). The severity of the inflammatory pattern ranged from mild reactions, featured by vessel dilatation, mild increase of the cellularity and small granulomas (Fig 4A), to strong focal reactivity. In the latter case, large necrotic granulomas were characterized by nodular aggregates of amebocytes in the periphery and their debrided remnants in the centre (Fig 4B). This last response was mainly observed in T. brevior infected snails, which were often featured by enlarged ventral surfaces of the foot and prominent vessel dilatation, along with few larval granulomas (Fig 4C). The occurrence of large amount of amebocytes was also seen in the kidney of T. brevior snails at 3 dpi (Fig 4D).
Fig 3. Histopathology: Inflammatory response to A. abstrusus in the H. aspersa.

Cell-poor granuloma formation with vacuolated amebocytes at 3dpi. (A); cell-rich granuloma formation at 9 dpi (B); small necrotic granuloma at 15 dpi (C); fibroblast-like reaction at 27 dpi (D) (scale bar = 50μm; H&E).
Fig 4. Histopathology: Larvae of A. abstrusus (asterisks) in the snail foot at 9 dpi, within granuloma formations.

(A); large necrotic granuloma with peripheral well-preserved amebocytes and their debris in the centre in snail infected by T. brevior at 21 dpi (B); polipoid enlargement of the ventral surface of the foot with dilated vessels and an abscess-like formation in T. brevior-infected snail (21 dpi) (C); multifocally-distributed amebocytes aggregates in the kidney parenchyma in T. brevior-infected snail (3 dpi) (D) (scale bar = 50μm; H&E).
Discussion
Results of this study indicate that L3 of A. abstrusus and T. brevior are either shed in the mucus by live terrestrial gastropods or released into the environment when snails had died in water. Both of these pathways of L3 elimination may represent alternative route(s) of transmission for feline lungworms under field conditions.
The emergence of A. abstrusus and T. brevior larvae from living snails, without either mechanical stressors or lack of food, indicates that this can occur spontaneously. This phenomenon has also been shown for other metastrongyloids [23]. Indeed, A. costaricensis L3 may leave Biomphalaria glabrata freshwater snails, as well as Phyllocaulis soleiformis and Sarasinula marginata slugs [24–26]. Similarly, A. cantonensis has been detected in the mucus of Parmarion martensi semi-slugs [7] and larvae of the muskoxen lungworm Umingmakstrongylus pallikuukensis (Strongylida, Protostrongylidae) emerged from Deroceras slugs and Catinella snails, being both found in the sediment of uneaten gastropod food or in water, coated with snail mucus [27]. In addition, Angiostrongylus vasorum (Strongylida, Angiostrongylidae) L3 may abandon B. glabrata gastropods if exposed at different temperatures [28]. Interestingly, when infected snails are given food, the elimination of L3 increases, probably as the result of enhanced mucus secretion [19]. Accordingly, the contamination of cat food left outdoors by snails shedding L3 might be a significant route of infection. This is, however, less likely to be the case when cats ingest grass or vegetables (e.g., the catnip Nepeta cataria) to induce vomiting as a remedy of hairballs.
Again, the elimination of A. abstrusus and T. brevior larvae by dead snails into water represents a further route of transmission for both lungworms, as corroborated by previous findings on A. cantonensis and A. costaricensis outbreaks, which most likely had occurred following the consumption of contaminated vegetables [20,29,30]. Indeed, considering that cats may also drink mouldy water, the swallowing of L3 cannot be ruled out. Although the longevity of L3 that have emerged from snails has not been assessed for both feline lungworms, it has been estimated to be up to 72 h for A. cantonensis from land snail [30] and seven days from water snails [24]. Although in vivo models documenting the above mentioned pathways are not available for feline lungworms, the transmission of free-living metastrongyloid larvae from freshwater snails to the final host has been demonstrated for A. cantonensis [7,29] and A. vasorum [28].
Whether the shedding/elimination of L3 represents an active or a passive process (i.e., triggered by the metastrongyloid itself or primed by gastropod hosts) is yet to be determined. However, considering that all A. abstrusus and T. brevior larvae seen here were L3 without any larval sheath, they are likely to have actively left their hosts, once reached the infective stage. Indeed, L3 of metastrongyloids are characterised by being motile, which enables them to move through the host (i.e., from the gut to the small intestine and blood stream) [12]. This hypothesis is supported by data on U. pallikuukensis, whose larvae do not emerge from the intermediate gastropod hosts, until they have reached the infective stage around three weeks post infection [25].
Results of the histology confirm that lungworm larvae can be found either in the foot and the viscera of infected specimens [31]. The localization of L3, mainly in the fibro-muscular layer of the foot, was probably due to the massive vascular supply in this organ, which provides the best conditions for the homing of the parasites, as already suggested for A. costaricensis [32]. Based on the data herein recorded, the intrinsic mechanical action of foot fibro-muscular layer might also account for the release of the L3 into the external habitat, along with the mucus secretions. Whether the detection of L3 in the foot represents a primary site of entrance for L1 throughout the skin (see [32,33]), or it occurs following the ingestion and vascular dissemination from other organs (e.g., kidney or skin), could not be assessed.
Even if the histology cannot be considered the best method to quantify the number of larvae in snails, L3 of A. abstrusus were more frequently detected in all experiments herein carried out than those of T. brevior. Therefore, the small number of T. brevior larvae might be accounted for by the presence of numerous large necrotic granulomas in snails, which most likely represented a first defence of the gastropod against the larval invasion. This pattern may indicate that T. brevior is more harmful for H. aspersa, and therefore less adapted to this intermediate host than A. abstrusus. This may be also inferred by the higher moulting percentage of A. abstrusus L3 (i.e., 25.6%) than T. brevior (i.e., 12.1%) under the same laboratory conditions. This hypothesis requires to be verified for other snail species, but it may explain the wider geographical distribution and higher prevalence of A. abstrusus infection in feline populations [9,34]. Further studies are required to assess the interactions between the nematode and the gastropod, in order to provide a more complete picture of the influence of host species on the epidemiology of these lungworms, as already carried out for schistosomosis [2]. Although several studies have reported the host range for metastrongyloid hosts [15], few have assessed the relative susceptibility of gastropods for those pathogens.
Nonetheless, based on the findings documented here, the existence of competitive exclusion between the two lungworms in the same intermediate host cannot be ruled out. Indeed, the competitive exclusions principle, also known as Gause’s law, says that if one of two species has even the slightest advantage over the other, that species will dominate [35]. Gause’s law has already been demonstrated for several parasite—host systems (e.g., human filarioids, livestock haemoparasites, Taenia cestodes) [36]. Accordingly, it has been demonstrated that Schistosoma mansoni and Schistosoma rodhaini trematodes display a species-specific mate preference in Mus musculus mice, with males of the latter species being dominant over those of the former [37]. Importantly, the inter-specific competition has been implicated with an increase in parasite virulence [37]. However, since none of these models has ever been applied to feline metastrongyloids, studies on snail immunology could provide useful data to help demonstrate whether gastropods are more susceptible to particular metastrongyloid species.
Concluding remarks
Further large-scale epidemiological studies that will help identify risk factors associated with the spread of A. abstrusus and T. brevior are needed. Based on data herein reported, owners should pay attention to their cats living outdoor and clean the water and food bowls if left outside, considering that snails and slugs may shed L3. In addition, since H. aspersa snails may act as intermediate hosts for other metastrongyloid nematodes, such as Oslerus rostratus (Strongylida, Filaridae) [38] or A. vasorum [12], the potential shedding of both lungworms within gastropod mucus should be investigated. Therefore, the identification of alternative pathways for parasite transmission is important particularly since the geographic range of gastropod-borne diseases is expanding due to inter alia introduction of allochthonous infected snails [39,40].
Acknowledgments
Authors would like to thank Alfonsa Cavalera and Francesca Fazio (University of Bari) for their technical support during the laboratory activities, as well as the two unknown reviewers for their valuable suggestions.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
The author(s) received no specific funding for this work.
References
- 1. Petney TN, Andrews RH, Saijuntha W, Wenz-Mücke A, Sithithaworn P (2013) The zoonotic, fish-borne liver flukes Clonorchis sinensis, Opisthorchis felineus and Opisthorchis viverrini . Int J Parasitol 43:1031–1046. 10.1016/j.ijpara.2013.07.007 [DOI] [PubMed] [Google Scholar]
- 2. Adema CM, Bayne CJ, Bridger JM, Knight M, Loker ES, et al. (2012) Will all scientists working on snails and the diseases they transmit please stand up? PloS Negl Trop Dis 6:e1835 10.1371/journal.pntd.0001835 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Olveda DU, Olveda RM, McManus DP, Cai P, Chau TN, et al. (2014) The chronic enteropathogenic disease schistosomiasis. Int J Infect Dis 28:193–203. 10.1016/j.ijid.2014.07.009 [DOI] [PubMed] [Google Scholar]
- 4. Wu SS, French SW, Turner JA (1997) Eosinophilic ileitis with perforation caused by Angiostrongylus (Parastrongylus) costaricensis. A case study and review. Arch Pathol Lab Med 121:989–991. [PubMed] [Google Scholar]
- 5. Wang QP, Lai DH, Zhu XQ, Chen XG, Lun ZR (2008) Human angiostrongyliasis. Lancet Infect Dis 8:621–630. 10.1016/S1473-3099(08)70229-9 [DOI] [PubMed] [Google Scholar]
- 6. Yii CY (1976) Clinical observations on eosinophilic meningitis and meningoencephalitis caused by Angiostrongylus cantonensis on Taiwan. Am J Trop Med Hyg 25:233–249. [DOI] [PubMed] [Google Scholar]
- 7. Cowie RH (2013) Pathways for transmission of angiostrongyliasis and the risk of disease associated with them. Hawaii J Med Public Health 72:70–74. [PMC free article] [PubMed] [Google Scholar]
- 8. Otranto D, Brianti E, Dantas-Torres F (2013) Troglostrongylus brevior and a non existent 'dilemma'. Trends Parasitol 29:517–518. 10.1016/j.pt.2013.09.001 [DOI] [PubMed] [Google Scholar]
- 9. Brianti E, Giannetto S, Dantas-Torres F, Otranto D (2014) Lungworms of the genus Troglostrongylus (Strongylida: Crenosomatidae): neglected parasites for domestic cats. Vet Parasitol 202:104–112. 10.1016/j.vetpar.2014.01.019 [DOI] [PubMed] [Google Scholar]
- 10. Giannelli A, Passantino G, Ramos RA, Lo Presti G, Lia RP, et al. (2014) Pathological and histological findings associated with the feline lungworm Troglostrongylus brevior . Vet Parasitol 204:416–419. 10.1016/j.vetpar.2014.05.020 [DOI] [PubMed] [Google Scholar]
- 11. Annoscia G, Latrofa MS, Campbell BE, Giannelli A, Ramos RA, et al. (2014) Simultaneous detection of the feline lungworms Troglostrongylus brevior and Aelurostrongylus abstrusus by a newly developed duplex-PCR. Vet Parasitol 199:172–178. 10.1016/j.vetpar.2013.10.015 [DOI] [PubMed] [Google Scholar]
- 12. Anderson RC (2000) The superfamily Metastrongyloidea. In: Nematode Parasites of Vertebrates Their Development and Transmission. Ed CABI, Wallingford, UK, pp. 163–164. [Google Scholar]
- 13. Caro TM (1980) Predatory behaviour in domestic cat mothers. Behaviour 74:128–148. [Google Scholar]
- 14. Caro TM (1981) Predatory behaviour and social play in kittens. Behaviour 76:1–24. [Google Scholar]
- 15. Gerichter CB (1949) Studies on the nematodes parasitic in the lungs of Felidae in Palestine. Parasitology 39:251–262. [DOI] [PubMed] [Google Scholar]
- 16. Jeżewski W, Buńkowska-Gawlik K, Hildebrand J, Perec-Matysiak A, Laskowski Z (2013) Intermediate and paratenic hosts in the life cycle of Aelurostrongylus abstrusus in natural environment. Vet Parasitol 198:401–405. 10.1016/j.vetpar.2013.09.003 [DOI] [PubMed] [Google Scholar]
- 17. Brianti E, Gaglio G, Napoli E, Falsone L, Giannetto S, et al. (2013) Evidence for direct transmission of the cat lungworm Troglostrongylus brevior (Strongylida: Crenosomatidae). Parasitology 140:821–824. 10.1017/S0031182013000188 [DOI] [PubMed] [Google Scholar]
- 18. Ng TP, Saltin SH, Davies MS, Johannesson K, Stafford R, et al. (2013) Snails and their trails: the multiple functions of trail-following in gastropods. Biol Rev Camb Philos Soc 88:683–700. 10.1111/brv.12023 [DOI] [PubMed] [Google Scholar]
- 19. Campion M (1961) The structure and function of the cutaneous glands in Helix aspersa . J Microsc Science 102:195–216. [Google Scholar]
- 20. Waugh CA, Shafir S, Wise M, Robinson RD, Eberhard ML, et al. (2005) Human Angiostrongylus cantonensis, Jamaica. Emerg Infect Dis 11:1977–1978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Giannelli A, Ramos RA, Annoscia G, Di Cesare A, Colella V, et al. (2014) Development of the feline lungworms Aelurostrongylus abstrusus and Troglostrongylus brevior in Helix aspersa snails. Parasitology 141:563–569. 10.1017/S003118201300187X [DOI] [PubMed] [Google Scholar]
- 22. Ramos RA, Giannelli A, Dantas-Torres F, Brianti E, Otranto D (2013) Survival of first-stage larvae of the cat lungworm Troglostrongylus brevior (Strongylida: Crenosomatidae) under different conditions. Exp Parasitol 135:570–572. 10.1016/j.exppara.2013.09.009 [DOI] [PubMed] [Google Scholar]
- 23. Boev SN (1975) Protostrongylids Fundamentals of nematology, Vol. 25 Helminthological Laboratory, Academy of Sciences of the USSR, Moscow: [English Translation, 1984, U.S. Department of Agriculture, Washington, D.C., and Amerind Publishing Co., New Delhi, India, 337 p]. [Google Scholar]
- 24. Ubelaker JE, Bullick GR, Caruso J (1980) Emergence of third-stage larvae of Angiostrongylus costaricensis Morera and Cespedes 1971 from Biomphalaria glabrata (Say). J Parasitol 66:856–857. [PubMed] [Google Scholar]
- 25. Bonetti VC, Graeff-Teixeira C (1998) Angiostrongylus costaricensis and the intermediate hosts: observations on elimination of L3 in the mucus and inoculation of L1 through the tegument of mollusc. Rev Soc Bras Med Trop 31:289–294. [DOI] [PubMed] [Google Scholar]
- 26. Mendonça CL, Carvalho OS, Mota EM, Pelajo-Machado M, Caputo LF, et al. (2003) Angiostrongylus costaricensis and experimental infection of Sarasinula marginata II: elimination routes. Mem Inst Oswaldo Cruz 98:893–898. [DOI] [PubMed] [Google Scholar]
- 27. Kutz SJ, Hoberg EP, Polley L (2000) Emergence of third-stage larvae of Umingmakstrongylus pallikuukensis from three gastropod intermediate host species. J Parasitol 86:743–749. [DOI] [PubMed] [Google Scholar]
- 28. Barçante TA, Barçante JM, Dias SR, Lima Wdos S (2003) Angiostrongylus vasorum (Baillet, 1866) Kamensky, 1905: emergence of third-stage larvae from infected Biomphalaria glabrata snails. Parasitol Res 91:471–475. [DOI] [PubMed] [Google Scholar]
- 29. Heyneman D, Lim BL (1967) Angiostrongylus cantonensis: proof of direct transmission with its epidemiological implications. Science 158:1057–1058. [DOI] [PubMed] [Google Scholar]
- 30. Cheng TC, Alicata JE (1964) Possible role of water in the transmission of Angiostrongylus cantonensis (Nematoda: Metastrongylidae). J Parasitol 2:39–40. [Google Scholar]
- 31. Di Cesare A, Crisi PE, Di Giulio E, Veronesi F, Frangipane di Regalbono A, Talone T, Traversa D (2013) Larval development of the feline lungworm Aelurostrongylus abstrusus in Helix aspersa . Parasitol Res 112:3101–3108. 10.1007/s00436-013-3484-2 [DOI] [PubMed] [Google Scholar]
- 32. Mendonça CLGF, Carvalho OS, Mota EM, Pelajo-Machado M, Caputo LFG, Lenzi HL (1999) Penetration Sites and Migratory Routes of Angiostrongylus costaricensis in the Experimental Intermediate Host (Sarasinula marginata). Mem Inst Oswaldo Cruz Rio de Janeiro 94:549–556. [DOI] [PubMed] [Google Scholar]
- 33. Thiengo SC (1996) Mode of infection of Sarasinula marginata (Mollusca) with larvae of Angiostrongylus costaricensis (Nematode) Mem Inst Oswaldo Cruz Rio de Janeiro 91:277–278. [Google Scholar]
- 34. Tamponi C, Varcasia A, Brianti E, Pipia AP, Frau V, et al. (2014) New insights on metastrongyloid lungworms infecting cats of Sardinia, Italy. Vet Parasitol 203:222–226. 10.1016/j.vetpar.2014.04.001 [DOI] [PubMed] [Google Scholar]
- 35. Paperna I (1964) Competitive exclusion of the Dactylogyrus extensus by Dactylogyrus vastator (Trematoda, Monogenea) on the gills of the reared carp. J Parasitol 50:94–98. [Google Scholar]
- 36. Molyneux DH, Mitre E, Bockarie MJ, Kelly-Hope LA (2014) Filaria zoogeography in Africa: ecology, competitive exclusion, and public health relevance. Trends Parasitol 30:163–169. 10.1016/j.pt.2014.02.002 [DOI] [PubMed] [Google Scholar]
- 37. Norton AJ, Webster JP, Kane RA, Rollinson D (2008) Inter-specific parasite competition: mixed infections of Schistosoma mansoni and S. rodhaini in the definitive host. Parasitology 135:473–484. 10.1017/S0031182007004118 [DOI] [PubMed] [Google Scholar]
- 38. Seneviratna P (1959) Studies on Anafilaroides rostratus Gerichter, 1949 in cats. II. The life cycle. J Helminthol 33:109–122. [DOI] [PubMed] [Google Scholar]
- 39. Morgan ER, Jefferies R, Krajewski M, Ward P, Shaw SE (2009) Canine pulmonary angiostrongylosis: the influence of climate on parasite distribution. Parasitol Int 58:406–410. 10.1016/j.parint.2009.08.003 [DOI] [PubMed] [Google Scholar]
- 40. Teem JL, Qvarnstrom Y, Bishop HS, da Silva AJ, Carter J, et al. (2013) The occurrence of the rat lungworm, Angiostrongylus cantonensis, in non indigenous snails in the Gulf of Mexico region of the United States. Hawaii J Med Public Health 72:11–14. [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.