

ABSTRACT
Understanding the mechanisms of lipid accumulation in microorganisms is important for several reasons. In addition to providing insight into assembly of biological membranes, lipid accumulation has important applications in the production of renewable fuels and chemicals. The photosynthetic bacterium Rhodobacter sphaeroides is an attractive organism to study lipid accumulation, as it has the ability to increase membrane production at low O2 tensions. Under these conditions, R. sphaeroides develops invaginations of the cytoplasmic membrane to increase its membrane surface area for housing of the membrane-bound components of its photosynthetic apparatus. Here we use fatty acid levels as a reporter of membrane lipid content. We show that, under low-O2 and anaerobic conditions, the total fatty acid content per cell increases 3-fold. We also find that the increases in the amount of fatty acid and photosynthetic pigment per cell are correlated as O2 tensions or light intensity are changed. To ask if lipid and pigment accumulation were genetically separable, we analyzed strains with mutations in known photosynthetic regulatory pathways. While a strain lacking AppA failed to induce photosynthetic pigment-protein complex accumulation, it increased fatty acid content under low-O2 conditions. We also found that an intact PrrBA pathway is required for low-O2-induced fatty acid accumulation. Our findings suggest a previously unknown role of R. sphaeroides transcriptional regulators in increasing fatty acid and phospholipid accumulation in response to decreased O2 tension.
IMPORTANCE Lipids serve important functions in living systems, either as structural components of membranes or as a form of carbon storage. Understanding the mechanisms of lipid accumulation in microorganisms is important for providing insight into the assembly of biological membranes and additionally has important applications in the production of renewable fuels and chemicals. In this study, we investigate the ability of Rhodobacter sphaeroides to increase membrane production at low O2 tensions in order to house its photosynthetic apparatus. We demonstrate that this bacterium has a mechanism to increase lipid content in response to decreased O2 tension and identify a transcription factor necessary for this response. This is significant because it identifies a transcriptional regulatory pathway that can increase microbial lipid content.
INTRODUCTION
Lipids serve important functions in living systems, either as structural components of membranes or as a form of carbon storage. Lipids derived from oil-rich microorganisms (including bacteria, yeasts, and microalgae) also offer a promising source of renewable fuels and chemicals (1, 2). However, the genetic and biochemical mechanisms regulating lipid accumulation in microorganisms are poorly understood, have been difficult to study, and are typically linked to stress conditions that hinder growth (3–5). Much effort has been directed at increasing lipid accumulation through alteration of enzymes involved in biosynthesis or catabolism of fatty acids, but little is known about the endogenous systems that regulate lipid accumulation (6, 7).
With an interest in understanding the molecular mechanisms regulating microbial lipid accumulation, we are analyzing the control of lipid accumulation in purple bacteria, which have the ability to increase their membrane content under low-O2 conditions. In this study, we investigate this process in Rhodobacter sphaeroides, a facultative purple bacterium that can grow via aerobic or anaerobic respiration or photosynthesis (8). Unlike many well-studied facultative bacteria, changes in O2 tension cause significant morphological changes in R. sphaeroides. At high O2 tensions, its cell envelope resembles that of other Gram-negative bacteria. In response to low O2 tension, R. sphaeroides increases its membrane surface area for assembly of a photosynthetic apparatus by developing specialized intracytoplasmic membrane (ICM) invaginations (9–11). Three major types of integral membrane pigment-protein complexes populate the ICM: the photochemical reaction center (RC) and two distinct light-harvesting complexes (LH1 and LH2) that together collect and convert the energy of light into biological energy (10, 12).
The pathways that control transcription of many genes required for the synthesis and function of the photosynthetic apparatus have been extensively studied at the genetic and genomic levels (12–15). The photosynthesis response regulator (Prr) proteins comprise a two-component signal transduction system activated by low O2 tension, in which PrrB is a membrane-spanning sensor histidine kinase and PrrA is a DNA-binding response regulator (16, 17). The AppA-PpsR regulatory system is composed of a DNA-binding repressor, PpsR, and the light- and redox-sensitive antirepressor AppA (18). FnrL, a member of the FNR (fumarate and nitrate reductase) family of transcription factors, is a global regulator of anaerobic gene expression that is directly inhibited by the presence of O2 (19, 20). These three pathways each contribute to increased expression of photosynthesis genes at low O2 tension, including those in the puf and puc operons (encoding polypeptides of the RC, LH1, and LH2 complexes), biosynthetic enzymes for bacteriochlorophyll (BChl) and carotenoid (Crt) pigments, and electron transport chain components (12, 13, 16, 21–23).
Despite the extensive knowledge of the systems regulating synthesis of the pigment and protein components of photosynthetic membranes, little is known about synthesis of phospholipids that form the ICM bilayer. Cells that make ICM have a higher rate of phospholipid synthesis (24, 25) and a greater proportion of lipid in cellular biomass (26) than cells grown at high O2, which are devoid of ICM. Yet it is not known how much more membrane is needed for ICM development or if there is a specific mechanism to increase fatty acid and phospholipid levels in response to low O2. Observations of de novo ICM synthesis have shown that nascent ICM invaginations can be seen before pigment-protein complexes are evident on their surface, suggesting that phospholipid accumulation may not be obligately linked with synthesis of this specialized membrane (9).
In this study, we ask whether an increase in membrane lipid abundance is linked to ICM assembly. In wild-type cells, we find that fatty acid content is correlated with the level of BChl. However, accumulation of these two ICM components is genetically separable, since we identify a mutant that is devoid of pigment-protein complexes but contains high, near-anaerobic levels of fatty acid at low O2 tensions.
MATERIALS AND METHODS
Bacterial strains and growth conditions.
The bacterial strains used in this study are described in Table 1. R. sphaeroides 2.4.1 strains were grown at 30°C in a succinate-based minimal medium (SIS) (27), unless otherwise noted. For growth in the presence of O2, 500-ml cultures were bubbled with 1% CO2, the indicated percentage of O2, and the balance in N2. For anaerobic growth, 500-ml cultures were bubbled with 5% CO2 and 95% N2. In addition, anaerobic photosynthetic cultures were illuminated with an incandescent light with an intensity of 3, 10, or 100 W/m2 measured through a red glass filter, and anaerobic dark cultures were grown in SIS supplemented with 10% Luria-Bertani medium and 0.3% dimethyl sulfoxide (DMSO). For analysis, cultures were grown to an optical density at 600 nm (OD600) of ∼0.5 to 0.9; anaerobic photosynthetic cultures were grown to an OD600 of ∼0.9 to 1.5 to achieve similar cell densities to high-O2 cultures at the lower OD range.
TABLE 1.
R. sphaeroides strains used in this study
| Strain | Relevant properties | Reference |
|---|---|---|
| 2.4.1 | Wild-type strain | 55 |
| PPS1 (ΔPpsR) | 2.4.1 ppsR::ΩKmr | 40 |
| PRRB78 | 2.4.1 derivative encoding L78P substitution in PrrB, with ΩSmrSpr at SmaI site downstream of prrB | 39 |
| PUFB1 | 2.4.1 ΔpufB::Knr | 41 |
| PRRA1 (ΔPrrA) | 2.4.1 prrA:: ΩSmrSpr | 43 |
| APP11 (ΔAppA) | 2.4.1 ΔappA::Tpr | 42 |
| JZ1678 (ΔFnrL) | 2.4.1 fnrL::ΩSmrSpr | 19 |
SC and BChl analyses.
To assess spectral complex (SC) assembly, aliquots of cell cultures were assayed by visible spectroscopy on an Olis DW-2/2000 spectrophotometer. All spectra were scaled to an absorbance of 1 at 680 nm to normalize for cell density and then staggered vertically for presentation of multiple curves on one axis. BChl content was estimated by readings of absorption at 775 nm of material extracted from cells into 7:2 acetone-methanol, using an extinction coefficient of 75 mM−1 cm−1 (28).
Fatty acid and lipid phosphorus analysis.
Cell harvesting, extraction of lipids with chloroform-methanol, esterification, and gas chromatography-mass spectrometry (GC-MS) analysis were performed as previously described (29). For lipid phosphorus measurements, dried lipid extractions from 2.5-ml samples were digested with perchloric acid and assayed for phosphorus content (30). Cell plating experiments were used to make a standard curve of OD600 versus CFU to normalize fatty acid, lipid phosphorus, and BChl content per cell. For the experiment shown in Fig. 2A, insufficient cell plating data were collected to make a curve, so cell counts from plating within that experiment were used to normalize.
FIG 2.
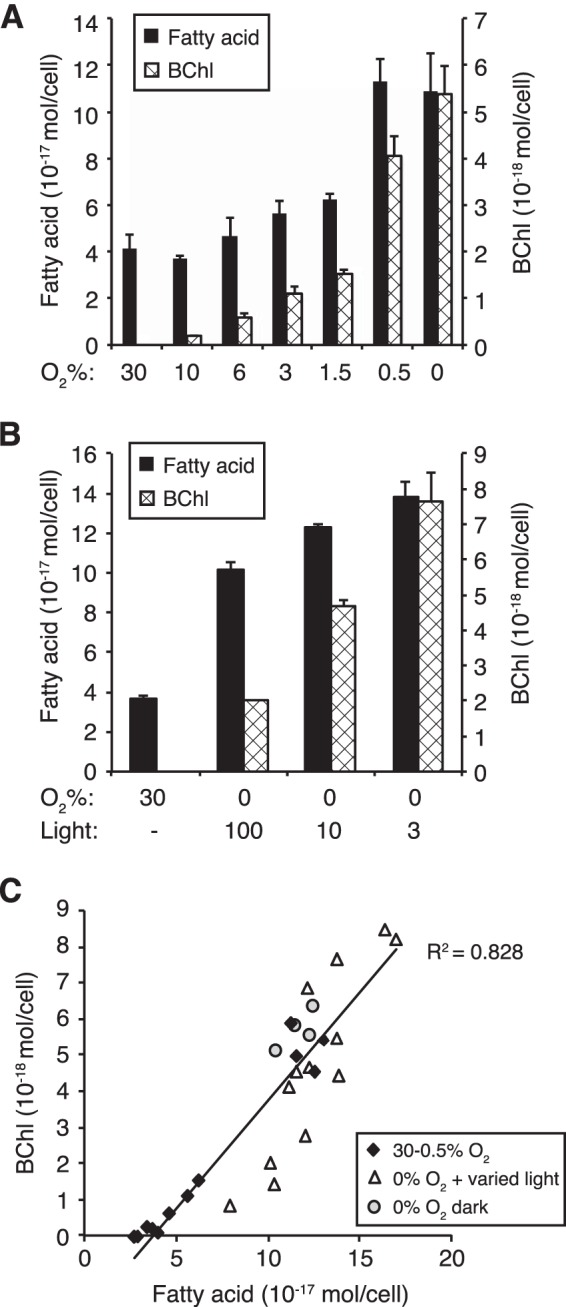
Fatty acid and BChl contents of R. sphaeroides cells when sparged with varied O2 and illuminated with varied light intensity. (A) Fatty acid and BChl contents for cells grown in the dark at the indicated O2 tensions. Cells grown without O2 were given DMSO as an electron acceptor. (B) Fatty acid and BChl contents for cells grown at high O2 tension (30%) compared to cells grown anaerobically at 3, 10, or 100 W/m2 incident light. For panels A and B, one representative experiment is shown; error bars represent standard deviations between two technical replicates. (C) Scatter plot of fatty acid versus BChl content representing individual biological replicates of cells grown under the conditions shown in panels A (diamonds and circles) and B (triangles ). The R2 value for the linear correlation was calculated with Microsoft Excel.
P values for statistical significance were calculated by unpaired t test using Graph Pad QuickCalcs.
TEM.
High-pressure freezing (HPF) and automatic freeze substitution (AFS), followed by plastic embedding, were used to produce thin sections of respective samples of cell suspensions. Cells were pelleted by brief centrifugation with a Quick-Spin minicentrifuge, and 5 μl of the concentrated cell suspension was transferred into an HPF flat specimen carrier and frozen with a Leica EM PACT high-pressure freezer (Leica Microsystems, Inc., Bannockburn, IL) at a typical rate of 1,700°C/s. The pods with compacted frozen cells were transferred under liquid nitrogen to the precooled AFS (EM AFS; Leica), and a protocol for cell fixation, water substitution by acetone, and a gradual warm-up to room temperature was followed (see Table S1 in the supplemental material). After 72 h, the samples were released from the pods, washed three times in acetone, gradually infiltrated with an ascending series of Spurr's low-viscosity embedding medium (Electron Microscopy Sciences, Hatfield, PA) (with 25, 50, and 75%, followed by three 100% washes for 120 min each), and cured at 60°C for 48 h. The polymerized blocks were sectioned to 70-nm thin sections with a Leica Ultracut UCT ultramicrotome, mounted on Formvar-coated 100 mesh Cu transmission electron microscopy (TEM) grids sputter coated with carbon, and poststained for 7 min with aqueous 2% uranyl acetate followed by 3 min of Reynolds' lead citrate (31) prior to TEM imaging. Samples were examined with the Tecnai T-12 TEM (FEI) operating at 120 kV with a LaB6 filament. Images were collected digitally with a 2×2K Ultrascan 1000 CCD (Gatan).
RESULTS
O2 and light regulation of fatty acid and spectral complex content in R. sphaeroides.
Given that R. sphaeroides increases its membrane surface area to synthesize ICM under low-O2 and anaerobic conditions, we wanted to test if these cells contain higher membrane lipid content than high-O2-grown cells. Thus, we sought to quantify membrane lipid content as a function of O2 tension and to determine how membrane content relates to the level of membrane-integral ICM spectral complexes (SCs). To do this, R. sphaeroides 2.4.1 was grown in liquid cultures sparged with high (30%) O2, anaerobically at moderate light intensity (10 W/m2) and in the dark using DMSO as an electron acceptor. In order to relate SC levels to membrane lipid content, we used total extracted BChl as a measure of SC assembly and both fatty acid and lipid phosphorus levels to assess membrane lipid content. We chose to express all of these measurements on a per cell basis in order to gain insight into the physiological changes of an individual cell under our experimental conditions. Since other cellular components of biomass also change in abundance as O2 and light intensity changes (26), this method of reporting provides insight into the physiology of lipid accumulation under the conditions examined.
As expected, the absorbance spectra of these cultures show that SCs are absent in high-O2 cultures and present in anaerobic cultures under light or dark conditions, with characteristic absorbance peaks at 800 and 850 nm (representing LH2) and a shoulder at 875 nm (representing LH1) (Fig. 1A). We also found that cells grown at 30% O2 tension lack detectable BChl, while cells grown under anaerobic conditions either at moderate light intensity or in the dark contain measurable levels of BChl (5 × 10−18 to 6 × 10−18 mol/cell) (Fig. 1B). At high (30%) O2 tension, cells contained 3.5 × 10−17 mol/cell of fatty acid and 1.5 × 10−17 mol/cell of lipid phosphorus (Fig. 1C). The approximately 2:1 ratio of fatty acid to phosphorus supports the conclusion that both assays are measuring phospholipid levels. Anaerobically grown cells contained ∼3 times more fatty acid and lipid phosphorus per cell than those grown at high O2 tension (Fig. 2), consistent with the increased membrane content associated with ICM formation. The relative amounts of the major fatty acid species were the same under all tested growth conditions (Fig. 1D) and were consistent with the fatty acid composition observed in previous studies of R. sphaeroides (32, 33). Since fatty acid and lipid phosphorus levels mirrored each other, we chose to use fatty acid levels to assess total membrane lipid content throughout the rest of this study.
FIG 1.

SC, BChl, and fatty acid contents of R. sphaeroides cells sparged with 30% O2 tension compared to those in cells grown anaerobically in the light or anaerobically in the dark with DMSO as an electron acceptor. (A) Absorbance scans of intact cells to show SC assembly. (B and C) Levels of BChl (B) and total fatty acid (C) in cells grown anaerobically compared to those grown with 30% O2. (D) Percentages of composition of the four predominant fatty acid species palmitic acid (C16:0), palmitoleic acid (C16:1), stearic acid (C18:0), and vaccenic acid (C18:1). Data for panel A are from one representative experiment. Data in panels B to D were averaged from six or more independent cultures; error bars represent standard deviation.
ICM formation is induced by lowered O2 tension, while light intensity also controls the number of ICM invaginations as well as the level and ratio of individual SCs (10, 11, 34, 35). In order to examine if the lipid content correlates with the SC component of ICM, we analyzed cells grown under a range of O2 concentrations (30 to 0%) in the dark. This analysis showed that both the cellular fatty acid and BChl contents began increasing at O2 concentrations below 10%, with a large increase occurring from 1.5% to 0.5%, such that 0.5% O2-grown cells contained levels of both components similar to those in anaerobic cells (Fig. 2A). Averaged over at least 6 independent cultures, there was no statistical difference between the fatty acid and BChl contents in 0.5% O2-grown cells (1.2 × 10−16 mol/cell fatty acid, 5.0 × 10−18 mol/cell BChl) compared to those in cells grown anaerobically under light or dark conditions (Fig. 1B). At external O2 concentrations of between 0.5 and 6%, the degree of increase in fatty acid content over high (30%)-O2 cells was proportional to the amount of BChl present. For example, at 1.5% O2, cells contained 29% of the BChl present in anaerobic cells (1.5 × 10−18 mol/cell versus 5.2 × 10−18 mol/cell), while the increase in fatty acid compared to 30% O2 (2.1 × 10−17 mol/cell) was 31% of the increase observed in anaerobic cells (6.8 × 10−17 mol/cell) (Fig. 2A). When we varied the incident light intensity under anaerobic conditions, we found that these changes also affected lipid and pigment content, with higher cellular lipid and BChl levels as the light intensity decreases (Fig. 2B). This is consistent with previous observations showing that high-light-grown cells contain less BChl and fewer ICM invaginations than low-light-grown cells (35–37). When BChl content was plotted against fatty acid content from these experiments, where we varied oxygen tension or light intensity, we found a positive correlation between the cellular levels of fatty acid and BChl (Fig. 2C) (R2 = 0.83). The data points for cells grown anaerobically in the dark (diamonds and circles) fell closer to this correlation line, while the points for high-light-grown cells in particular fell under the line, likely due at least in part to an increased proportion of LH1 that contains fewer BChl molecules than LH2 (38). These experiments establish that membrane fatty acid content increases proportionally to BChl levels in wild-type cells.
Mutations in the PrrBA or PpsR regulatory pathways alter the control of fatty acid and BChl accumulation at high O2 tension.
Since we observed a correlation between BChl and fatty acid levels for wild-type cells under environmental conditions that control ICM synthesis, we tested whether membrane content was altered by mutations that allow SC assembly at high O2 tension, a condition under which they are absent in wild-type cells (9–11). We analyzed mutants with alterations in regulatory pathways that control ICM development in response to O2 deprivation. One mutant (PrrB78) has an amino acid substitution in the PrrB sensor histidine kinase that alters signaling and leads to increased expression of some photosynthesis genes under high-O2 conditions (39). A second mutant lacks the PpsR transcriptional repressor (ΔPpsR), leading to derepression, and thus increased expression, of many photosynthesis genes at high O2 tensions (40). The absorbance spectra show that, in contrast to wild-type cells that lack detectable SCs at 30% O2, these mutants have detectable levels of LH1 (absorbance at 875 nm) and LH2 complexes (absorbances at 800 and 850 nm) under these conditions (Fig. 3A). Quantitation of BChl and fatty acid levels in cells grown at 30% O2 tension showed that each of these mutants had increased fatty acid in addition to the increased BChl (P < 0.03 for both fatty acid and BChl increases for both mutants) but less of each of these components than wild-type cells under anaerobic conditions (Fig. 3B). Thus, when the PrrB78 and ΔPpsR mutants are grown at 30% O2, the coupling between BChl and fatty acid content is maintained. However, the accumulation of both fatty acid and BChl in these mutants is separated from that of normal control by decreased O2 tension.
FIG 3.
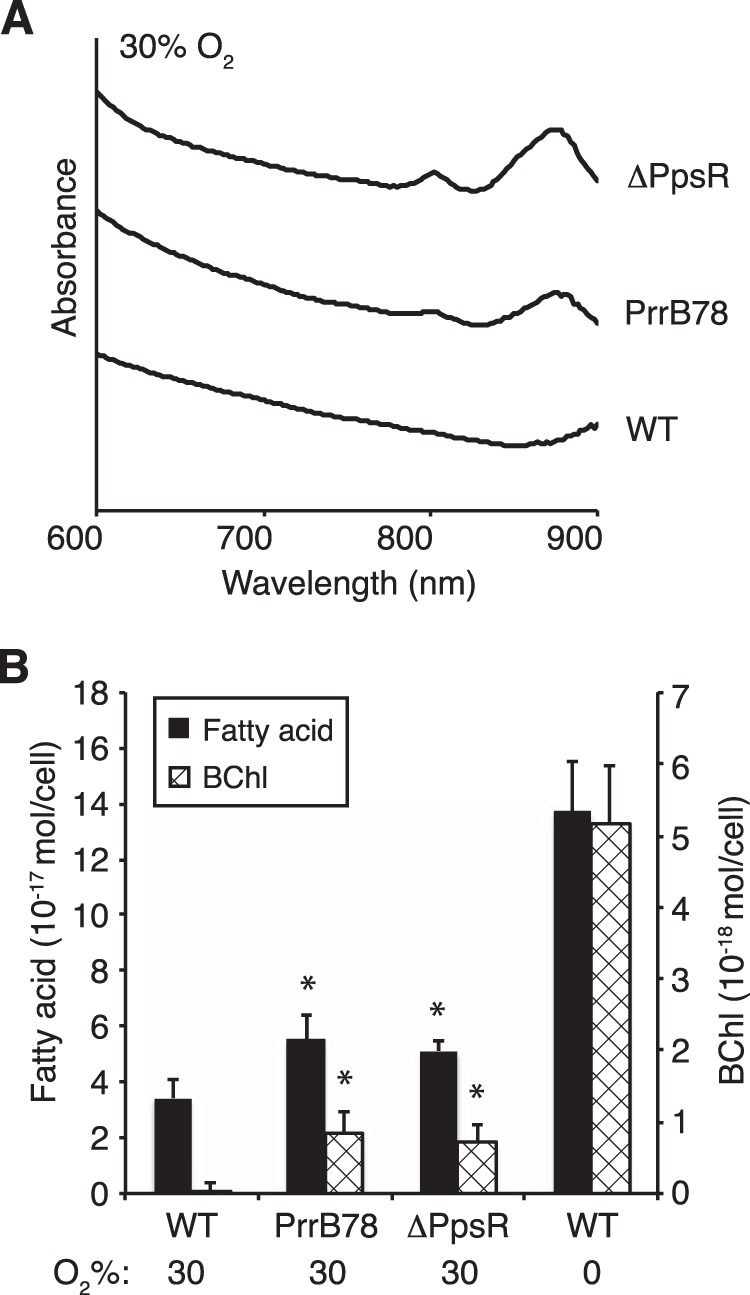
SC, BChl, and fatty acid contents of R. sphaeroides mutants that activate photosynthesis gene expression in the presence of oxygen (PrrB78 and ΔPpsR) compared to wild-type (WT) cells. (A) Absorbance scans of intact cells to show SC assembly. (B) Fatty acid and BChl contents of wild-type, ΔPpsR, and PrrB78 cells grown in the presence of 30% O2 compared to wild-type cells grown anaerobically at 10 W/m2 light. Asterisks indicate that the increases in fatty acid and BChl contents in both mutants relative to wild-type cells under the same condition are statistically significant at P < 0.03. Data represent the average from three biological replicates from independent experiments; error bars indicate standard deviation.
Mutations that uncouple fatty acid and BChl accumulation.
We also asked whether membrane fatty acid content was altered when SC synthesis was blocked genetically. One way to do this was to analyze the PUFB1 strain, which has a deletion disrupting expression of the pufKBALMX operon (41). This lesion prevents expression of the LH1 structural polypeptides and the RC structural polypeptides, blocking assembly of LH1 complexes (41). PUFB1 also has a greater than 95% decrease in levels of LH2 complexes under anaerobic conditions (41). Although PUFB1 is unable to grow photosynthetically, it grows anaerobically in the dark with DMSO as an electron acceptor (41). When we analyzed the PUFB1 mutant and wild-type cells, as expected, they lacked detectable levels of SCs at 30% O2 (Fig. 4A). However, we unexpectedly found that the fatty acid content of PUFB1 was 1.5-fold higher at high (30%) O2 tension than that of wild-type cells grown under the same conditions (4.6 × 10−17 versus 3.0 × 10−17 mol/cell, respectively; P = 0.0007) (Fig. 4B). Under anaerobic dark conditions, PUFB1 showed an almost complete loss of peaks in the near-infrared range (Fig. 4A) and 33 times less BChl (1.6 × 10−19 mol/cell) than was found in wild-type cells (5.4 × 10−18 mol/cell), indicating a nearly complete lack of SCs. In addition, the fatty acid content in anaerobically grown PUFB1 cells did not show a statistically significant increase compared to that in cells grown at high (30%) O2 tension, although it was elevated 1.9-fold compared to the level in wild-type cells that lack SCs (at high O2) (5.7 × 10−17 versus 3.0 × 10−17 mol/cell, respectively; P = 0.002) (Fig. 4B). Thus, disruption of the pufKBALMX operon results in higher fatty acid content under both high-O2 and anaerobic conditions compared to the level in wild-type aerobic cells. This demonstrates that PUFB1 is able to increase its lipid content relative to the wild type even though it does not assemble appreciable levels of SCs.
FIG 4.

Fatty acid and SC levels in the PUFB1 mutant. (A) Absorption spectra of wild-type (WT) and PUFB1 cells grown at 30% O2 tension (top) and anaerobically in the dark (bottom). (B) Fatty acid content of cells under the same conditions shown in panel A. Data represent the average from three biological replicates; error bars indicate standard deviation. ∗, P = 0.0025; ∗∗, P = 0.0035; NS, not significant.
The properties of PUFB1 suggested that there were genetic alterations in which lipid accumulation could be separated from assembly of ICM SCs. To test this hypothesis further, we examined BChl and fatty acid levels in cells containing additional mutations in known regulators of photosynthesis genes. We reasoned that if O2-dependent increases in fatty acid levels require one or more of these regulatory pathways, then we should be able to identify a mutant(s) in which SC assembly is disrupted but fatty acid accumulation is seen or vice versa. For these experiments, we analyzed previously characterized strains in which there is inactivation of either the PrrA transcriptional regulator (ΔPrrA), AppA, an antirepressor that regulates PpsR transcriptional activity (ΔAppA), or the FnrL transcriptional regulator (ΔFnrL) (Table 1) (19, 42, 43). The mutations in each of these strains reduce expression of some photosynthesis genes and lower SC assembly at low O2: as a result, all of these mutants are unable to grow photosynthetically (19, 42, 43). When grown anaerobically in the dark, unlike wild-type cells, the ΔAppA and ΔPrrA cells lack appreciable levels of SCs (42, 43). The ΔFnrL mutant is unable to grow anaerobically in the dark; when grown at low (2%) O2, it assembles SCs but to a lesser extent than wild-type cells (19).
We used growth at low (0.5%) O2 to compare fatty acid and BChl levels in these mutants to those in wild-type cells, since all three strains grow under these conditions, and our experiments showed that wild-type cells accumulated high levels of fatty acid and BChl under these conditions (Fig. 2A). We found that at 0.5% O2, both the ΔPrrA and ΔAppA mutants lacked detectable SCs (Fig. 5A) and had only trace amounts of BChl (Fig. 5B), similar to what had previously been observed under anaerobic conditions (42, 43). The ΔFnrL mutant, however, assembled SCs at 0.5% O2 (Fig. 5A) with 71% of the total BChl (3.5 × 10−18 mol/cell versus 5.4 × 10−18 mol/cell in the wild type) (Fig. 5B). When wild-type cells were grown at low (0.5%) O2, the increase in fatty acid content was 3.1-fold relative to the level in cells grown at high (30%) O2 (Fig. 5C). In contrast, the ΔPrrA mutant had only a small 1.3-fold increase in fatty acid content when grown at 0.5% O2 relative to when grown at 30% O2, while the ΔAppA and ΔFnrL mutants had 1.9- and 2.5-fold, respectively, increases in fatty acid content at 0.5% O2 relative to 30% O2 (Fig. 5C). When these mutants were grown at high (30%) O2, all three strains had fatty acid levels similar to those of wild-type cells (Fig. 5C).
FIG 5.

SC, BChl, and fatty acid levels of R. sphaeroides mutants (ΔPrrA, ΔAppA, and ΔFnrL) that have decreased expression of photosynthesis genes. (A and B) Absorption spectra (A) and BChl content (B) of cells grown at 0.5% O2 tension in the dark. WT, wild type. (C) Fatty acid content of cells grown at 30% O2 and 0.5% O2 tension in the dark. Data in panels B and C represent the average from three or more biological replicates from independent experiments; error bars indicate standard deviation. (D) Scatter plot of fatty acid versus BChl content of individual biological replicates of wild-type and mutant cells grown at 0.5% O2 tension. The solid line indicates the correlation between fatty acid and BChl calculated for wild-type cells under the growth conditions shown in Fig. 2 (30 to 0.5% O2, anaerobic in the dark, and anaerobic at 3 different light intensities).
When comparing the fatty acid and BChl contents of mutant cells, it follows that the ΔPrrA and ΔFnrL mutants maintain the correlation between fatty acid and BChl levels: PrrA cells (circles) have low levels of both fatty acid and BChl at 0.5% O2, while the ΔFnrL cells (squares) have high levels of both fatty acid and BChl (Fig. 5D). In contrast, the ΔAppA cells grown at 0.5% O2 (triangles) contain increased fatty acid but low BChl levels (Fig. 5D), suggesting that fatty acid and BChl accumulation are uncoupled by the loss of AppA. Furthermore, despite the low levels of BChl in the ΔAppA mutant, there was no detectable SC assembly (Fig. 5A). In sum, we conclude based on these data that low-O2-induced fatty acid accumulation is not absolutely dependent on assembly of active SCs. The significance of these observations to the pathways that control O2-dependent increases in membrane lipids is discussed below (see Discussion).
Since the ΔAppA mutant contained ∼2-fold more fatty acid when grown at 0.5% O2 than when grown at 30% O2, but did not contain appreciable amounts of assembled SCs, we asked whether the cells formed ICM-like structures. We used transmission electron microscopy (TEM) to compare the morphologies of wild-type, ΔAppA, ΔPrrA, and ΔFnrL cells. TEM micrographs of wild-type and ΔFnrL cells grown at 0.5% O2 showed the presence of ICM (Fig. 6A and D), while the ΔPrrA mutant was devoid of ICM (Fig. 6B and C). These findings correlate with the fatty acid and BChl levels we observed under these conditions (Fig. 5B to D). In contrast to wild-type and ΔFnrL cells, the ΔAppA mutant did not have any apparent ICM when grown at 0.5% O2 (Fig. 6E and F). To examine if the increased fatty acid content in the AppA mutant grown at 0.5% O2 reflected an increase in lipid, we measured lipid phosphorus levels for this strain. A 2.5-fold increase in lipid phosphorus from 2.0 × 10−17 mol/cell at 30% O2 to 4.9 × 10−17 mol/cell at 0.5% O2 was observed. In sum, these data suggest the ΔAppA mutant has increased phospholipid content at 0.5% O2 that does not accumulate in structures that appear by TEM to be similar to ICM, nor is it visible in these images.
FIG 6.

TEM micrographs of thin sections of the R. sphaeroides wild type (WT) and mutant (ΔPrrA, ΔAppA, and ΔFnrL) cells grown at 0.5% O2 tension. The arrows in the wild-type and ΔFnrL panels indicate examples of the lighter-colored spherical ICM structures. The larger white cellular structures represent polyhydroxybutyrate granules, which are present in all strains but not in each cross-section of a cell. A representative image is shown for each strain (A, B, D, and E), with additional lower-magnification images included for ΔPrrA (C) and ΔAppA (F) to show more cells.
DISCUSSION
The goal of this study was to analyze the environmental and genetic systems that control lipid production in the facultative bacterium R. sphaeroides. As O2 tensions decrease, R. sphaeroides increases its membrane surface area to synthesize an ICM system that contains lipids, SCs, and other bioenergetic enzymes, which enable cells to grow via photosynthesis (10, 11, 34, 37). In this study, we used fatty acid levels to assess membrane lipid content under conditions that promote ICM synthesis. We chose this approach since R. sphaeroides is not known to accumulate or contain genes annotated to encode enzymes for, neutral lipid synthesis (such as triacylglycerols [TAGs] or wax esters) (44). To further validate this approach, we found the expected 2:1 ratio of fatty acid to lipid phosphorus. Previous studies have demonstrated that there are not significant differences in the phospholipid or fatty acid composition between aerobic and anaerobic cells (32, 45), therefore, we focused on the difference in absolute levels of fatty acids. Our fatty acid analysis method also allowed us to assess fatty acid composition, which was not found to vary significantly between the conditions or strains used in this study.
We show here that conditions that promote ICM synthesis also cause cells to increase their total lipid content. At low O2 tension (0.5%), as well as anaerobically in the dark or at moderate light intensity, this increase in lipid content is approximately 3-fold compared to that in cells grown at high O2, which are devoid of ICM. We also found that a decrease in O2 tension is sufficient for this increased lipid accumulation, indicating that light is not needed for this response. Below we summarize the new information we have obtained by analyzing the environmental and genetic controls that allow this bacterium to increase its membrane surface area.
Uncoupling of membrane lipid and SC accumulation.
Our data demonstrate a correlation between fatty acid and BChl levels under physiological conditions that control ICM synthesis (Fig. 2C), indicating there is a direct relationship between the amount of membrane lipid and assembled SCs. This correlation is not surprising, since the increased membrane surface area is thought to house the SCs and other integral membrane enzymes of the ICM. However, the systems that link fatty acid levels to those of ICM SCs are still unresolved. For example, it is possible that increased expression of an ICM protein(s) is sufficient to drive increased membrane synthesis. This is illustrated by observations in Escherichia coli, where overexpression of individual integral membrane proteins, including fumarate reductase (46), the glycerol acyl transferase PlsB (47), or the ATP synthase b subunit (48), is sufficient to induce formation of intracellular membranes. Indeed, we found that when synthesis of SCs was increased by mutation under high O2 tension, when they are not normally present, membrane lipid content also increased (Fig. 3A and B). Thus, it is possible that increased membrane content at low O2 is an indirect consequence of inserting a large amount of SCs or other integral membrane proteins into the ICM.
However, when assembly of SCs was blocked at low O2 in the ΔAppA mutant, this strain contained ∼2-fold-higher fatty acid and lipid phosphorus levels than when grown at high O2 (Fig. 5A and C). This observation suggests that there is a mechanism to increase membrane content in response to low O2 tension that is independent of SC insertion into the ICM. This notion is also supported by observations of the de novo ICM assembly that occurs when cells are shifted from aerobic to anaerobic growth conditions (9). Under these conditions, new ICM invaginations form before BChl accumulation is detected. Furthermore, these nascent ICM invaginations appear smooth when analyzed by freeze fracture electron microscopy, suggesting they are enriched in phospholipid, in contrast to mature photosynthetic membranes that contain a high level of particulate membrane-bound pigment-protein complexes (9).
Potential regulators of membrane lipid accumulation.
The R. sphaeroides transcriptional networks regulating gene expression in response to decreased O2 tension contain many operons encoding proteins that are necessary for the assembly of functional photosynthetic membranes (12). In this study, we used mutations in each of these pathways to test their role in the accumulation of membrane lipids at low O2 tension.
For some mutants, the normal linkage between fatty acid and BChl levels was maintained at 0.5% O2. Specifically, the ΔPrrA mutant had low levels of both lipid and pigment, while the ΔFnrL mutant has high levels of both lipid and pigment under these conditions. In contrast, the ΔAppA mutant has no detectable SCs but high levels of fatty acid at 0.5% O2, suggesting that this lesion uncouples the linkage between SCs and membrane lipid content. This observation suggests that while AppA is necessary for SC assembly, it is not necessary for increased lipid levels at low O2 since ΔAppA cells still exhibit an increase in fatty acid at 0.5% O2 compared to cells grown at 30% O2. Since AppA acts as an antirepressor of the transcription factor PpsR (18), this suggests that depression of the transcriptional target genes of PpsR is not necessary for increased fatty acid at low O2. It is also possible that deletion of AppA leads to derepression of a regulator of fatty acid biosynthesis independent of PpsR. However, fatty acid levels are not significantly increased in the ΔAppA mutant when it is sparged with 30% O2. Thus, we do not think it is likely that the increase in fatty acid levels seen in the ΔAppA mutant is due to direct derepression of a regulator of fatty acid synthesis. Finally, our data suggest that FnrL is not required for either increased membrane lipid or SC synthesis at low O2, while the PrrBA pathway is required for both. In sum, we conclude that transcriptional regulation by FnrL and the AppA-PpsR pathway are dispensable for low-O2-induced fatty acid accumulation, since the ΔAppA and ΔFnrL mutant strains still increased fatty acid content at least 2-fold under low O2 tension compared to high O2 tension (Fig. 5C), while the PrrBA pathway is required. We can infer from this that one or more genes within the PrrA regulon either directly or indirectly are necessary for cells to increase membrane content for ICM development. To date there are not any members of this regulon that are known or predicted to participate in or regulate fatty acid and phospholipid biosynthesis.
Our finding that at 0.5% O2 the ΔFnrL mutant assembles SCs, has 71% of the BChl and 83% of the fatty acid as wild-type cells, and forms ICM was somewhat unexpected since previous analysis found that this mutant has much lower SC levels than the wild type at 2% O2 tension (49) and formed many fewer ICM invaginations than wild-type cells in poorly aerated cultures (50). These differences may be due to the different O2 tensions used between our studies and previous experiments and may indicate that loss of FnrL changes the sensitivity of cells to decreased O2 for induction of ICM SC and lipid synthesis. However, from our data at 0.5% O2, we conclude that FnrL is not strictly essential for low-O2-induced lipid accumulation.
Relevance to lipid production in other microbes.
Our data demonstrate that R. sphaeroides has a mechanism to increase lipid content in response to decreased O2 tension, which is at least partially dependent on an intact PrrBA pathway. This is significant and relevant to both ICM assembly and the potential engineering of microbes for production of lipid biofuels and chemicals, because it identifies a transcriptional regulatory pathway that can increase microbial lipid content. In most microbes that accumulate storage lipids, such as triacylglycerols (TAGs), accumulation is normally induced by nutrient limitation or other stresses (3–5). Under such conditions, the gene expression changes contributing to increased lipid storage have not been elucidated. For example, in the oleaginous microalgae Cytotella cryptica under silicon starvation conditions that lead to TAG accumulation, the first dedicated enzyme in fatty acid biosynthesis, acetyl coenzyme A (acetyl-CoA) carboxylase (ACCase), has elevated gene expression and enzyme activity (51, 52). However, overexpression of ACCase alone, in this and other microalgae, does not lead to increased TAG accumulation (52). Other studies in microalgae, yeast, and bacteria have used similar approaches with more success, targeting deletion and/or overexpression of enzymes both singly and in combination to increase the yield of TAGs or other fatty-acid-derived fuels (7, 53). While the success of individual studies varies, this approach requires the manipulation of many different genes to maximize product yield. Instead, an approach that altered activity of a transcriptional regulator would be advantageous, as it could potentially alter the expression of many relevant genes affecting a metabolic pathway with one genetic manipulation and also be used in organisms where the enzymes of fatty acid biosynthesis and degradation are not well characterized. In support of this idea, a recent study demonstrated that in E. coli, cells engineered to produce free fatty acids, homologous expression of the FadR transcription factor, a negative regulator of fatty acid degradation and positive regulator of biosynthesis, increased fatty acid yield 7.5-fold (54). Elucidating additional regulatory networks controlling fatty acid and lipid accumulation in other microbes will facilitate achieving of the increased product yields necessary for economical large-scale production of biofuels and chemicals.
Our studies in R. sphaeroides identify a novel oxygen-regulated mechanism of lipid accumulation that is dependent, at least in part, on the PrrA global transcriptional regulator. Future studies will be directed at determining the relationship of the PrrBA pathway and other yet to be discovered pathways to increased lipid accumulation and how the knowledge gained from studying these systems might be applied to microbial production of renewable fuels and chemicals.
Supplementary Material
ACKNOWLEDGMENTS
This work was supported by DOE Great Lakes Bioenergy Research Center grant (DOE Office of Science BER DE-FC02-07ER64494) to T.J.D. and USDA NIFA fellowship 2011-67012-30702 to K.C.L. Electron microscopy was performed at the Environmental Molecular Science Laboratory (EMSL), a DOE Office of Science user facility sponsored by the Office of Biological and Environmental Research and located at PNNL.
We thank Mark Gomelsky for providing strains APP11 (ΔAppA) and PPS1 (ΔPpsR) and Jill Zeilstra-Ryalls for providing strain JZ1678 (ΔFnrL).
Footnotes
Supplemental material for this article may be found at http://dx.doi.org/10.1128/JB.02510-14.
REFERENCES
- 1.Li Q, Du W, Liu D. 2008. Perspectives of microbial oils for biodiesel production. Appl Microbiol Biotechnol 80:749–756. doi: 10.1007/s00253-008-1625-9. [DOI] [PubMed] [Google Scholar]
- 2.Meng X, Yang J, Xu X, Zhang L, Nie Q, Xian M. 2009. Biodiesel production from oleaginous microorganisms. Renewable Energy 34:1–5. doi: 10.1016/j.renene.2008.04.014. [DOI] [Google Scholar]
- 3.Blatti JL, Michaud J, Burkart MD. 2013. Engineering fatty acid biosynthesis in microalgae for sustainable biodiesel. Curr Opin Chem Biol 17:496–505. doi: 10.1016/j.cbpa.2013.04.007. [DOI] [PubMed] [Google Scholar]
- 4.Alvarez HM, Steinbuchel A. 2002. Triacylglycerols in prokaryotic microorganisms. Appl Microbiol Biotechnol 60:367–376. doi: 10.1007/s00253-002-1135-0. [DOI] [PubMed] [Google Scholar]
- 5.Beopoulos A, Nicaud JM, Gaillardin C. 2011. An overview of lipid metabolism in yeasts and its impact on biotechnological processes. Appl Microbiol Biotechnol 90:1193–1206. doi: 10.1007/s00253-011-3212-8. [DOI] [PubMed] [Google Scholar]
- 6.Kosa M, Ragauskas AJ. 2011. Lipids from heterotrophic microbes: advances in metabolism research. Trends Biotechnol 29:53–61. doi: 10.1016/j.tibtech.2010.11.002. [DOI] [PubMed] [Google Scholar]
- 7.Liang MH, Jiang JG. 2013. Advancing oleaginous microorganisms to produce lipid via metabolic engineering technology. Prog Lipid Res 52:395–408. doi: 10.1016/j.plipres.2013.05.002. [DOI] [PubMed] [Google Scholar]
- 8.Melandri AB, Zannoni D. 1978. Photosynthetic and respiratory electron flow in the dual functional membrane of facultative photosynthetic bacteria. J Bioenerg Biomembr 10:109–138. doi: 10.1007/BF00743056. [DOI] [PubMed] [Google Scholar]
- 9.Chory J, Donohue TJ, Varga AR, Staehelin LA, Kaplan S. 1984. Induction of the photosynthetic membranes of Rhodopseudomonas sphaeroides: biochemical and morphological studies. J Bacteriol 159:540–554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Tavano CL, Donohue TJ. 2006. Development of the bacterial photosynthetic apparatus. Curr Opin Microbiol 9:625–631. doi: 10.1016/j.mib.2006.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kiley PJ, Kaplan S. 1988. Molecular genetics of photosynthetic membrane biosynthesis in Rhodobacter sphaeroides. Microbiol Rev 52:50–69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Zeilstra-Ryalls JH, Kaplan S. 2004. Oxygen intervention in the regulation of gene expression: the photosynthetic bacterial paradigm. Cell Mol Life Sci 61:417–436. doi: 10.1007/s00018-003-3242-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zeilstra-Ryalls J, Gomelsky M, Eraso JM, Yeliseev A, O'Gara J, Kaplan S. 1998. Control of photosystem formation in Rhodobacter sphaeroides. J Bacteriol 180:2801–2809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Arai H, Roh JH, Kaplan S. 2008. Transcriptome dynamics during the transition from anaerobic photosynthesis to aerobic respiration in Rhodobacter sphaeroides 2.4.1. J Bacteriol 190:286–299. doi: 10.1128/JB.01375-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gomelsky L, Moskvin OV, Stenzel RA, Jones DF, Donohue TJ, Gomelsky M. 2008. Hierarchical regulation of photosynthesis gene expression by the oxygen-responsive PrrBA and AppA-PpsR systems of Rhodobacter sphaeroides. J Bacteriol 190:8106–8114. doi: 10.1128/JB.01094-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Comolli JC, Carl AJ, Hall C, Donohue T. 2002. Transcriptional activation of the Rhodobacter sphaeroides cytochrome c2 gene P2 promoter by the response regulator PrrA. J Bacteriol 184:390–399. doi: 10.1128/JB.184.2.390-399.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Potter CA, Ward A, Laguri C, Williamson MP, Henderson PJ, Phillips-Jones MK. 2002. Expression, purification and characterisation of full-length histidine protein kinase RegB from Rhodobacter sphaeroides. J Mol Biol 320:201–213. doi: 10.1016/S0022-2836(02)00424-2. [DOI] [PubMed] [Google Scholar]
- 18.Elsen S, Jaubert M, Pignol D, Giraud E. 2005. PpsR: a multifaceted regulator of photosynthesis gene expression in purple bacteria. Mol Microbiol 57:17–26. doi: 10.1111/j.1365-2958.2005.04655.x. [DOI] [PubMed] [Google Scholar]
- 19.Zeilstra-Ryalls JH, Kaplan S. 1995. Aerobic and anaerobic regulation in Rhodobacter sphaeroides 2.4.1: the role of the fnrL gene. J Bacteriol 177:6422–6431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Dufour YS, Imam S, Koo BM, Green HA, Donohue TJ. 2012. Convergence of the transcriptional responses to heat shock and singlet oxygen stresses. PLoS Genet 8:e1002929. doi: 10.1371/journal.pgen.1002929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Dufour YS, Kiley PJ, Donohue TJ. 2010. Reconstruction of the core and extended regulons of global transcription factors. PLoS Genet 6:e1001027. doi: 10.1371/journal.pgen.1001027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Bruscella P, Eraso JM, Roh JH, Kaplan S. 2008. The use of chromatin immunoprecipitation to define PpsR binding activity in Rhodobacter sphaeroides 2.4.1. J Bacteriol 190:6817–6828. doi: 10.1128/JB.00719-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Karls RK, Wolf JR, Donohue TJ. 1999. Activation of the cycA P2 promoter for the Rhodobacter sphaeroides cytochrome c2 gene by the photosynthesis response regulator. Mol Microbiol 34:822–835. doi: 10.1046/j.1365-2958.1999.01649.x. [DOI] [PubMed] [Google Scholar]
- 24.Onishi JC, Niederman RA. 1982. Rhodopseudomonas sphaeroides membranes: alterations in phospholipid composition in aerobically and phototrophically grown cells. J Bacteriol 149:831–839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lascelles J, Szilagyi JF. 1965. Phospholipid synthesis by Rhodopseudomonas spheroides in relation to the formation of photosynthetic pigments. J Gen Microbiol 38:55–64. doi: 10.1099/00221287-38-1-55. [DOI] [PubMed] [Google Scholar]
- 26.Imam S, Yilmaz S, Sohmen U, Gorzalski AS, Reed JL, Noguera DR, Donohue TJ. 2011. iRsp1095: a genome-scale reconstruction of the Rhodobacter sphaeroides metabolic network. BMC Syst Biol 5:116. doi: 10.1186/1752-0509-5-116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sistrom WR. 1960. A requirement for sodium in the growth of Rhodopseudomonas spheroides. J Gen Microbiol 22:778–785. doi: 10.1099/00221287-22-3-778. [DOI] [PubMed] [Google Scholar]
- 28.Clayton RK. 1966. Spectroscopic analysis of bacteriochlorophylls in vitro and in vivo. Photochem Photobiol 5:669–677. doi: 10.1111/j.1751-1097.1966.tb05813.x. [DOI] [Google Scholar]
- 29.Lennon CW, Lemmer KC, Irons JL, Sellman MI, Donohue TJ, Gourse RL, Ross W. 2014. A Rhodobacter sphaeroides protein mechanistically similar to Escherichia coli DksA regulates photosynthetic growth. mBio 5(3):e01105-14. doi: 10.1128/mBio.01105-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Rouser G, Fkeischer S, Yamamoto A. 1970. Two dimensional thin layer chromatographic separation of polar lipids and determination of phospholipids by phosphorus analysis of spots. Lipids 5:494–496. doi: 10.1007/BF02531316. [DOI] [PubMed] [Google Scholar]
- 31.Reynolds ES. 1963. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J Cell Biol 17:208–212. doi: 10.1083/jcb.17.1.208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Russell NJ, Harwood JL. 1979. Changes in the acyl lipid composition of photosynthetic bacteria grown under photosynthetic and non-photosynthetic conditions. Biochem J 181:339–345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Donohue TJ, Cain BD, Kaplan S. 1982. Purification and characterization of an N-acylphosphatidylserine from Rhodopseudomonas sphaeroides. Biochemistry 21:2765–2773. doi: 10.1021/bi00540a029. [DOI] [PubMed] [Google Scholar]
- 34.Niederman RA. 2013. Membrane development in purple photosynthetic bacteria in response to alterations in light intensity and oxygen tension. Photosynth Res 116:333–348. doi: 10.1007/s11120-013-9851-0. [DOI] [PubMed] [Google Scholar]
- 35.Chory J, Kaplan S. 1983. Light-dependent regulation of the synthesis of soluble and intracytoplasmic membrane proteins of Rhodopseudomonas sphaeroides. J Bacteriol 153:465–474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Adams PG, Hunter CN. 2012. Adaptation of intracytoplasmic membranes to altered light intensity in Rhodobacter sphaeroides. Biochim Biophys Acta 1817:1616–1627. doi: 10.1016/j.bbabio.2012.05.013. [DOI] [PubMed] [Google Scholar]
- 37.Yen GS, Cain BD, Kaplan S. 1984. Cell-cycle-specific biosynthesis of the photosynthetic membrane of Rhodopseudomonas sphaeroides. Structural implications. Biochim Biophys Acta 777:41–55. doi: 10.1016/0005-2736(84)90495-4. [DOI] [PubMed] [Google Scholar]
- 38.Cogdell RJ, Gall A, Kohler J. 2006. The architecture and function of the light-harvesting apparatus of purple bacteria: from single molecules to in vivo membranes. Q Rev Biophys 39:227–324. doi: 10.1017/S0033583506004434. [DOI] [PubMed] [Google Scholar]
- 39.Eraso JM, Kaplan S. 1995. Oxygen-insensitive synthesis of the photosynthetic membranes of Rhodobacter sphaeroides: a mutant histidine kinase. J Bacteriol 177:2695–2706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Gomelsky M, Kaplan S. 1997. Molecular genetic analysis suggesting interactions between AppA and PpsR in regulation of photosynthesis gene expression in Rhodobacter sphaeroides 2.4.1. J Bacteriol 179:128–134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Davis J, Donohue TJ, Kaplan S. 1988. Construction, characterization, and complementation of a Puf− mutant of Rhodobacter sphaeroides. J Bacteriol 170:320–329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Gomelsky M, Kaplan S. 1995. appA, a novel gene encoding a trans-acting factor involved in the regulation of photosynthesis gene expression in Rhodobacter sphaeroides 2.4.1. J Bacteriol 177:4609–4618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Eraso JM, Kaplan S. 1994. prrA, a putative response regulator involved in oxygen regulation of photosynthesis gene expression in Rhodobacter sphaeroides. J Bacteriol 176:32–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kontur WS, Schackwitz WS, Ivanova N, Martin J, Labutti K, Deshpande S, Tice HN, Pennacchio C, Sodergren E, Weinstock GM, Noguera DR, Donohue TJ. 2012. Revised sequence and annotation of the Rhodobacter sphaeroides 2.4.1 genome. J Bacteriol 194:7016–7017. doi: 10.1128/JB.01214-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Donohue TJ, Cain BD, Kaplan S. 1982. Alterations in the phospholipid composition of Rhodopseudomonas sphaeroides and other bacteria induced by Tris. J Bacteriol 152:595–606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Weiner JH, Lemire BD, Elmes ML, Bradley RD, Scraba DG. 1984. Overproduction of fumarate reductase in Escherichia coli induces a novel intracellular lipid-protein organelle. J Bacteriol 158:590–596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Wilkison WO, Walsh JP, Corless JM, Bell RM. 1986. Crystalline arrays of the Escherichia coli sn-glycerol-3-phosphate acyltransferase, an integral membrane protein. J Biol Chem 261:9951–9958. [PubMed] [Google Scholar]
- 48.Arechaga I, Miroux B, Karrasch S, Huijbregts R, de Kruijff B, Runswick MJ, Walker JE. 2000. Characterisation of new intracellular membranes in Escherichia coli accompanying large scale over-production of the b subunit of F1Fo ATP synthase. FEBS Lett 482:215–219. doi: 10.1016/S0014-5793(00)02054-8. [DOI] [PubMed] [Google Scholar]
- 49.Zeilstra-Ryalls JH, Gabbert K, Mouncey NJ, Kaplan S, Kranz RG. 1997. Analysis of the fnrL gene and its function in Rhodobacter capsulatus. J Bacteriol 179:7264–7273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Fedotova Y, Zeilstra-Ryalls J. 2014. Analysis of the role of PrrA, PpsR, and FnrL in intracytoplasmic membrane differentiation of Rhodobacter sphaeroides 241 using transmission electron microscopy. Photosynth Res 119:283–290. doi: 10.1007/s11120-013-9944-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Roessler PG. 1988. Changes in the activities of various lipid and carbohydrate biosynthetic enzymes in the diatom Cyclotella cryptica in response to silicon deficiency. Arch Biochem Biophys 267:521–528. doi: 10.1016/0003-9861(88)90059-8. [DOI] [PubMed] [Google Scholar]
- 52.Sheehan J, Dunahay T, Bennemann J, Roessler P. 1998. A look back at the U.S. Department of Energy's aquatic species program: biodiesel from algae; close-out report. Technical report. NREL/TP-580-24190. National Renewable Energy Laboratory, Golden, CO. [Google Scholar]
- 53.Janssen HJ, Steinbuchel A. 2014. Fatty acid synthesis in Escherichia coli and its applications towards the production of fatty acid based biofuels. Biotechnol Biofuels 7:7. doi: 10.1186/1754-6834-7-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Zhang F, Ouellet M, Batth TS, Adams PD, Petzold CJ, Mukhopadhyay A, Keasling JD. 2012. Enhancing fatty acid production by the expression of the regulatory transcription factor FadR. Metab Eng 14:653–660. doi: 10.1016/j.ymben.2012.08.009. [DOI] [PubMed] [Google Scholar]
- 55.van Niel CB. 1944. The culture, general physiology, morphology, and classification of the non-sulfur purple and brown bacteria. Bacteriol Rev 8:1–118. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.