

Abstract
Considered an EPPO A2 quarantine pest, Bursaphelenchus xylophilus is the causal agent of the pine wilt disease and the most devastating plant parasitic nematode attacking coniferous trees in the world. In the early stages of invasion, this nematode has to manage host defence mechanisms, such as strong oxidative stress. Only successful, virulent nematodes are able to tolerate the basal plant defences, and furthermore migrate and proliferate inside of the host tree. In this work, our main objective was to understand to what extent B. xylophilus catalases are involved in their tolerance to oxidative stress and virulence, using as oxidant agent the reactive oxygen species hydrogen peroxide (H2O2). After 24 hours of exposure, high virulence isolates of B. xylophilus could withstand higher H2O2 concentrations in comparison with low virulence B. xylophilus and B. mucronatus, corroborating our observation of Bxy-ctl-1 and Bxy-ctl-2 catalase up-regulation under the same experimental conditions. Both catalases are expressed throughout the nematode intestine. In addition, transgenic strains of Caenorhabditis elegans overexpressing B. xylophilus catalases were constructed and evaluated for survival under similar conditions as previously. Our results suggest that catalases of high virulence B. xylophilus were crucial for nematode survival under prolonged exposure to in vitro oxidative stress, highlighting their adaptive response, which could contribute to their success in host conditions.
Introduction
From 1905 until the present time, pine wilt disease (PWD) has been one of the most threatening diseases for worldwide forests [1]. PWD is considered a complex disease resulting from the interaction of three main elements: the pathogenic agent Bursaphelenchus xylophilus (PWN, pine wood nematode), the insect-vector Monochamus spp. (responsible for PWN dissemination), and the host tree, mostly from Pinus spp. [2–3]. In a B. xylophilus population, there is intra-specific variability in the ability to cause PWD (virulence) [4]. The existence of different levels of virulence among PWN isolates, and their relation to PWN reproductive ability in vitro and in vivo were already reported [5–6]. In this context, the low virulence nematodes, often designated as avirulent, show lower reproduction rates and increased generation times that eventually affect their host invasion and colonization [7–9].
In the early stage of PWD, PWN has to cope with different levels of plant immune responses. The first, non-specific, host tree reaction to nematode invasion (or other biotic/abiotic stress) is oxidative burst, an excessive production of reactive oxygen species (ROS). The predominant ROS in plant oxidative burst is H2O2 [10–11]. H2O2 is relatively stable, cell wall diffusible, and found to be transversal in different plant-pathogens systems, being a fundamental signal for inducing plant resistance (i.e. involved in cell-wall reinforcement or induction of defence-related genes in healthy adjacent tissues) [12]. H2O2 is particularly important since it is extremely reactive and can lead to the formation of radical hydroxyl OH-, for which no specific scavenger exists [12]. Generally, parasitic nematodes respond to an oxidant threat by increasing their protective antioxidant enzyme levels [13]. The major antioxidant enzyme families considered are: superoxide dismutases (SOD, EC 1.15.1.1) for conversion of superoxide anion to H2O2, glutathione peroxidases (GPX, EC 1.11.1.9), catalases (CTL, E.C. 1.11.1.6) and peroxiredoxins (PRDX, EC 1.11.1.15) for the conversion of H2O2 into water [14]. Furthermore, many parasites employ many enzyme groups under oxidative stress conditions [14]. In plant parasitic nematodes, Molinari and Miacola [15] showed the presence of many antioxidant enzymes and the relation to its lifestyle stages of Meloidogyne incognita and M. hapla, Globodera rostochiensis and G. pallida, Heterodera schachtii and H. carotae, and Xiphinema index. Robertson et al. [16] described an important role of PRDX and GPX in G. rostochiensis. Bellafiore et al. [17] reported the presence of several detoxifying enzymes, in particular several glutathione S-transferases (GST, EC 2.5.1.18) in the secretome of M. incognita as means for controlling the global oxidative status and potential nematode virulence. PWN also has an efficient antioxidant system to reduce the deleterious effects of tree oxidative burst [18], mostly present in the nematode secretome [19] and expressed in the cuticle [20].
Pine oxidative burst was found intrinsically related with the production of ROS in the first stages of B. xylophilus invasion [21–22]. He et al. [23] evaluated the content of H2O2 of P. thunbergii, P. massoniana and P. taeda at the early stage of B. xylophilus invasion, and found that H2O2 content was higher in the susceptible pines than the resistant P. taeda. In this study, we specifically analyzed the tolerance of B. xylophilus isolates with different levels of virulence (high and low virulence) under in vitro oxidative stress conditions by assessing the effects of H2O2 exposure to nematode survival, catalase activity, catalase gene expression and their spatial localization. Additionally, we expressed B. xylophilus in the nematode model C. elegans to explore the efficiency of B. xylophilus catalases in promoting survival.
Material and Methods
B. xylophilus isolates and culturing
Two B. xylophilus isolates with high virulence (Bx Ka4 and Bx T4) and one isolate with low virulence (Bx C14-5) [6] were used in this study. In addition, one isolate of B. mucronatus (Un1), considered a low virulence nematode, was also used. All isolates were supplied by the FFPRI (Forestry and Forest Products Research Institute). The origin of nematode isolates Bx Ka4 and Bx C14-5 is described by Aikawa and Kikuchi [6]. The isolate Bx T4 was collected from dead Pinus densiflora in 1992, in Iwate prefecture, Japan and B. mucronatus Un1 was obtained from Monochamus alternatus, in the Kyoto prefecture.
Nematodes were cultured in Botrytis cinerea on autoclaved barley seed at 25°C. Prior to experiments, nematodes were extracted overnight using Baermann funnel technique at 25°C. Nematodes were washed three times with sterilized distilled water with 10 min. centrifugations at 1,000 rpm between washes, surface cleaned with 3% L-lactic acid for 30s, and washed once with sterilized distilled water [9]. Mix-stages nematodes were used in all experiments.
Oxidative stress tolerance test
H2O2 was used as oxidative agent at concentrations ranging between 15 and 40 mM for Bursaphelenchus isolates. After nematode surface sterilization, the concentration of Bursaphelenchus was adjusted to 150 nematodes per 50 μl of sterile water. Mortality of nematodes was scored after 24h-stress exposure. Nematodes were considered dead if no movements were observed after mechanical stimulation. This experiment was repeated three independent times, with two technical replications for each.
H2O2 neutralization
H2O2 neutralization was inferred using an H2O2 Assay Kit (abcam, Massachusetts, USA). For this purpose, 150 nematodes were exposed for 24h in 15 mM H2O2. Afterwards, nematodes were pelleted at 1,000 g for 15 min and the supernatant was used for analysis following the manufacture’s protocol. This assay was repeated three independent times with two technical replications.
Relative gene expression of catalases under oxidative stress
Relative gene expressions of CTL (catalase) enzymes (Bxy-ctl-1 and Bxy-ctl-2), previously predicted by Vicente et al. [24], were analyzed by qRT-PCR using SYBR green assay. Total RNA was extracted from 24h-stressed nematodes (approximately 2,500) in 15 mM H2O2 using CellAmp Direct RNA Prep Kit for RT-PCR (Takara Bio Inc., Japan) and following manufacture’s instructions. The concentration was quantified using NanoVue plus spectrophotometer (GE Healthcare Life Sciences, USA). Total RNA (adjusted to a concentration of 50 ng/μl) was reverse transcribed using oligo dT primer and PrimeScript RT enzyme from PrimeScript RT reagent Kit (Perfect Real Time, Takara Bio Inc., Japan). qRT-PCR was performed using CFX96 Real-Time (Bio-Rad Laboratories, Inc., California), and SYBR Premix Ex Taq II (Tli RnaseH Plus) kit (Takara Bio Inc., Japan). Actin (Bxy-act-1) was used as a reference gene for calculating relative expression levels of CTLs genes [25]. Primers were designed using Prime 3 software [26] and tested for specificity prior to qRT-PCR (Table 1). For each treatment, two experimental replications were conducted with two technical replications each. Controls with no template added were prepared for each qPCR run. Thermal cycling conditions were: initial denaturation at 95°C for 30 sec; 39 cycles of denaturation at 95°C for 5 sec, annealing and extension at 60°C for 30 sec; followed by the melting curve. A single peak at the melting temperature of the PCR-product confirmed primer specificity.
Table 1. Characterization of Bursaphelenchus xylophilus CTLs, and primers used for CTLs expression analysis.
| Gene and Protein DB | ORF 1 | MW (Da) | SignalP 2 | Secretome 3 (Sec/whole) |
|---|---|---|---|---|
| Bxy-ctl-1 (BUX.s00579.159) | 513 | 58836.42 | NO | NO (ND) |
| Bxy-ctl-2 (BUX.s01109.377) | 272 | 54869.71 | NO | YES (0.26) |
| Primers for qRT-PCR (5’– 3’) | ||||
| Bxy-ctl-1 | For: GCCAGCGTCTTCAGCAAAGT; Rev: CCAAATTCCGTCATCGGTGT | |||
| Bxy-ctl-2 | For: CCGACTTCTTTCAACGGAAC; Rev: CCTTCATCGAGCACCTTTTC | |||
| Bxy-act-1 | For: CATCCTCCGTCTCGACTTGG; Rev: ATGTCACGCACGATTTCACG | |||
| Primers for In-situ hybridization (5’– 3’) | ||||
| Bxy-CTL-1 | For: CTTCCGATCAACTGCCCCTT; Rev: ATTTAGACAAGGGGCCAGCC | |||
| Bxy-CTL-2 | For: AGAGACCCCATCCTCTTCCC; Rev: AGCTTCTCCCTTTGCGTTGA | |||
In-situ hybridization
Single-stranded DNA probes (sense and antisense) labelled with digoxigenin (Roche, United States of America) were used to detect mRNA in mix-stage B. xylophilus as described by De Boer et al. [28]. Table 1 presents the primers used to generate Bxy-CTL-1 and Bxy-CTL-2 probes (Table 1). As positive control for successful hybridization, ENG (cellulase) probe was also prepared as described by Kikuchi et al. [29]. Images were acquired on an Olympus Bx50 light microscope at 10x-40x magnifications.
Transgenic C. elegans construction
C. elegans culturing and handling were carried out at 20°C as described by Brenner [30]. Strains used in this experiments were N2 (Bristol strain) and DP38 unc-119(ed3)III. To make reporter constructs, all PCRs were performed with Takara PrimeSTAR GXL DNA polymerase (Takara Bio Inc., Japan). B. xylophilus ctl-1 and -2 cDNAs were fused with C. elegans ctl-1 or -3 promoters and ligated into the gfp vector pPD95.77 (kindly provided by A. Fire, Stanford University) with In-Fusion HD Cloning system (Takara Bio Inc., Japan). We cloned a 1,940 bp sequence upstream from the Ce-ctl-3 start codon for Ce-Pctl-3, and a 1,600 bp sequence between Ce-ctl-3 and Ce-ctl-1 CDS (coding sequences) for Ce-Pctl-1. Fusion genes created in this experiments were Pctl-1::Bxy-ctl-1::gfp and Pctl-3::Bxy-ctl-2::gfp, and PCR primer sequences are listed in Table 2. Each reporter construct (100 μg/ml) was co-injected with an equal concentration of pDP#MM016B into the gonadal arms of unc-119(ed3) adult hermaphrodites to obtain KHA149 {unc-119(ed3)III; chuEx149[Pctl-1::Bxy-ctl-1::gfp, pDP#MM016B]}, KHA150 {unc-119(ed3)III; chuEx150[Pctl-1::Bxy-ctl-1::gfp, pDP#MM016B]}, KHA151 {unc-119(ed3)III; chuEx151[Pctl-3::Bxy-ctl-2::gfp, pDP#MM016B]}, and KHA152 {unc-119(ed3)III; chuEx152[Pctl-3::Bxy-ctl-2::gfp, pDP#MM016B]}. Successful transgenic C. elegans were further confirmed by visualization of fluorescence expression patterns with a Nikon SMZ800 dissection microscope equipped with a fluorescence filter and a ZEISS LSM710 confocal laser-scanning microscope.
Table 2. List of primers used in C. elegans constructs.
| Genes | Primer 5’– 3’ |
|---|---|
| Cectl-1Prom_Ifs | For: CGACTCTAGAGGATCATTGTTTGATATTCAAACTTTTGTA; |
| Rev: CGTCATTTTGGTTCTGAAATTTTAGTTAGG | |
| Cectl-3Prom_Ifs | For: CGACTCTAGAGGATCCTTCGTCACACTTCTATGGAATCC; |
| Rev: GCCATTTTGAAGATTTACTGTTGAATTTCCG | |
| Bxy-ctl-1_Ifs | For: CAAACCAAAATGACGGACTTTGCGGCCAATC; |
| Rev: CCAATCCCGGGGATCTTGACATCCTCTTGCAATCTCCAC | |
| Bxy-ctl-2_Ifs | For: AATCTTCAAAATGGCAAACAACAAGAAGACTGC; |
| Rev: CCAATCCCGGGGATCTCAAATGGGCCTTGACCTTCTTGG |
Oxidative stress tolerance in C. elegans
Synchronized L1 stage C. elegans were obtained by treating egg-containing/or gravid adults with sodium hypochlorite [30] and allowed to grow on NGM plates seeded with E. coli OP50 at 20°C for 48 hours until the late L4 stage. L4 stage C. elegans were washed with M9 buffer and adjusted to 20 nematodes per 50 μl of M9 buffer. Nematodes were transferred to a 96-well plate containing each concentration of H2O2 (0 to 250 μM). The plate was incubated at 20°C for 24 hours, and then examined for animal viability under a stereomicroscope. Since our transgenic nematodes used in this experiment retained transgenes as an extrachromosomal DNA array, non-transgenic uncoordinated nematodes appeared at a certain proportion (around 30%). We excluded non-transgenic uncoordinated animals when performing this experiment. All tests were carried out at 20°C. This experiment was repeated three independent times, and in each experimental repetition, two technical replications were used.
Data analysis
Statistical analysis was performed using SPSS version 11.5. One-way ANOVA analysis was performed in order to test if the different B. xylophilus and B. mucronatus isolates were equally tolerant to H2O2 treatments for the parameters: nematode survival percentage, H2O2 neutralization, and C. elegans survival percentage. Post hoc multi-comparison Duncan test (significance level of 0.05) was used to group significantly different Bursaphelenchus sp. isolates in the following parameters: nematode survival percentage, H2O2 neutralization, and C. elegans survival percentage. Levene’s Test of Homogeneity was performed a priori to infer ANOVA assumption of homogeneity of variance.
Relative gene expressions of CTLs were analysed using the 2-ΔΔCT method [25]. The data were analysed with CT (cycle threshold) values in normal and stress conditions and using Eq. (1), where ΔΔCT = (CT,Target—CT,Actin)normal-(CT,Target—CT,Actin)stress. The fold change of CTL genes, normalized to β-act and relative to the expression at normal conditions, were calculated for each sample using Eq. (1). Data represent the mean ± standard deviation (SD). Statistical differences at 95% and 99% confidence levels were inferred by Student t-test by comparison between relative expression of both CTLs for each B. xylophilus isolate and the control treatment without H2O2.
Results
Oxidative stress tolerance and H2O2 neutralisation
B. xylophilus (high virulence: Bx Ka4 and Bx T4; low virulence: Bx C14-5) and B. mucronatus (low virulence: Bm Un1) isolates were tested in increasing concentrations of H2O2 (0, 15, 20, 30 and 40 mM). After a 24h-exposure to this oxidant, nematode survival was examined (Fig 1). A clear difference between high and low virulence isolates was observed, even in the lowest H2O2 concentration (15 mM). The isolate Bx Ka4 seemed to be the most tolerant to H2O2 at all concentrations followed by Bx T4. Statistical differences (p < 0.05) between Bx Ka4 and Bx T4 were found in all H2O2 concentrations. Low virulence isolates (Bm Un1 and Bx C14-5) were sensitive to H2O2, having low survival even in 15 mM H2O2. No statistical differences (p > 0.05) were found between isolates.
Fig 1. Survival percentage of Bursaphelenchus sp. isolates (high virulence B. xylophilus Ka4 and B. xylophilus T4, and low virulence B. xylophilus C14-5 and B. mucronatus Un1) after 24 hours exposition to H2O2 conditions (H2O2 concentrations ranging from 0 to 40 mM).

Error bars represent standard deviation. Different letters above the columns indicate significant differences (p < 0.05) between Bursaphelenchus sp. isolates survival percentages in each H2O2 treatment, according to post-hoc Duncan’s test.
In terms of H2O2 neutralization (Fig 2), high virulence Bx Ka4 and Bx T4 were more efficient at H2O2 reduction (p < 0.05) in comparison with the low virulence Bm Un1 and Bx C14-5 (no statistical differences between each pair).
Fig 2. H2O2 neutralization of of Bursaphelenchus sp. isolates (high virulence B. xylophilus Ka4 and B. xylophilus T4, and low virulence B. xylophilus C14-5 and B. mucronatus Un1) after 24 hours exposition to H2O2.

Error bars represent standard deviation. Different letters above the columns indicate significant differences (p < 0.05) between Bursaphelenchus sp. isolates H2O2 neutralization, according to post-hoc Duncan’s test.
Gene expression analysis
B. xylophilus catalases (Bxy-ctl-1 and Bxy-ctl-2; Table 1) were previously analyzed and found to be conserved among Nematoda [24]. The CDS of both CTLs were similar between high and low virulence B. xylophilus isolates (data not shown). Because the lack of B. mucronatus genome in databases prevented the analyses of gene expression for the Bm Un1 isolate, our studies about levels of virulence will only focus on the Bx Ka4, Bx T4 and Bx C14-5 isolates. Relative gene expression of the CTLs was analyzed after a 24h-exposure to 15 mM H2O2 (Fig 3). High virulence isolates Bx Ka4 and Bx T4 presented a higher induction of CTL expression than low virulence Bx C14-5. In fact, the relative expression of both Bxy-ctl-1 and Bxy-ctl-2 of Bx C14-5 in stress conditions was not significantly different (p > 0.05) from expression in non-stress conditions. In oxidative conditions, Bxy-ctl-1 and Bxy-ctl-2 from both high virulence isolates Bx Ka4 and Bx T4 were significantly induced (0.01< p < 0.05) in comparison with non-stress conditions (Fig 3A and 3B). Both Bxy-ctl-1 and Bxy-ctl-2 of Bx Ka4 were 2.5-fold up-regulated. Bxy-ctl-1 of Bx T4 was significantly induced, nearly 2-fold (Fig 3A). Bxy-ctl-2 was also significantly up regulated (p < 0.05) in stress, nearly 4-fold more than in normal conditions (Fig 3B).
Fig 3. Relative gene expression of Bxy-ctl-1 (A) and Bxy-ctl-2 (B) of B. xylophilus (high virulence isolates Ka4 and T4; low virulence isolate C14-5) after 24 hours exposition to H2O2.
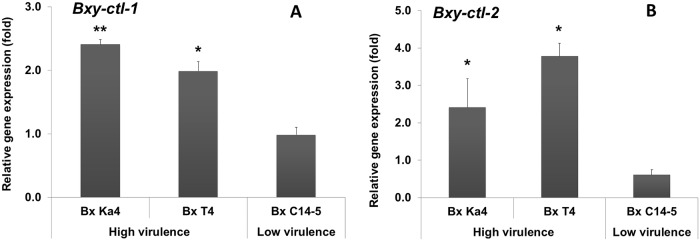
* and ** indicate, respectively, statistical differences at 95% and 99% confidence levels compared to a normalized value of 1.00 for control treatment without H2O2. Error bars represent standard deviation.
Bxy-ctl-1 and Bxy-ctl-2 messenger RNAs presented the same expression pattern in the nematode’s intestine (Fig 4A and 4B). Bxy-eng-1, profusely expressed in B. xylophilus oesophageal glands, was used as a positive control (Fig 4D) [29]. No hybridization was observed for the control sense probe of Bxy-ctl-1 (Fig 4C), Bxy-ctl-2 or Bxy-egn-1 (data not shown).
Fig 4. mRNA expression patterns of Bxy-ctl-1 (A), Bxy-ctl-2 (B), and Bxy-eng-1 (D) in B. xylophilus Ka4.

No expression signal was observed with Bxy-ctl-1 sense probe (C). Light microscope images of B. xylophilus head region. Scale bars, 100 μm.
Transgenic C. elegans and OS tolerance
Three catalase genes exist in tandem in the C. elegans genome (Ce-ctl-3, -1, and -2), while two genes exist separately in the B. xylophilus genome (WormBase, http://www.wormbase.org/). Because Bxy-ctl-1 and Bxy-ctl-2 sequences were highly homologous to Ce-ctl-1 and Ce-ctl-3 respectively, and Ce-Pctl-1 is identical to Ce-Pctl-2, we constructed transgenic C. elegans overexpressing Bx-ctl-1 or Bxy-ctl-2 with C. elegans promoters Ce-Pctl-1 or Ce-Pctl-3. We created two independent transgenic lines for each construct; KHA149 and 150 for Ce-Pctl-1::Bx-ctl-1::gfp, and KHA151 and 152 for Ce-Pctl-3::Bx-ctl-2::gfp.
Under the control of C. elegans promoters, B. xylophilus Bxy-ctl-1 and Bxt-ctl-2 showed different spatial expression patterns; ctl-1 was mainly detected in the cytosol of the intestine, and ctl-2 was detected as high levels along the nervous system and pharynx (Fig 5A and 5B). These expression patterns were the same with those previously reported [31]. Although these transgenic C. elegans were extrachromosomal lines with unstable transgene transmission, the expression patterns were similar between the transgenic lines KHA149 and 150, and between KHA151 and 152 (data not shown). We used KHA149 and 151 for further experiments.
Fig 5. Expression patterns of Bxy-ctl-1::gfp (A) and Bxy-ctl-2::gfp (B) in, respectively, transgenic C. elegans KHA149 and KHA151.

Differential interference contrast (DIC) microscope images and, DIC and fluorescence-merged images (DIC+FL) of C. elegans head, vulva and tail region. Scale bars, 100 μm.
To gauge C. elegans (wild-type and transgenic) tolerance to oxidative stress conditions, they were exposed to H2O2 concentrations between 50 and 500 μM, substantially lower than the concentrations used for B. xylophilus. The survival of wild-type C. elegans N2 decreased significantly (p < 0.05) with increasing concentrations of H2O2 (Fig 6). Interestingly, both of the transgenic C. elegans overexpressing B. xylophilus catalases were resistant to H2O2 stress conditions. KHA149 and KHA151, expressing Bxy-ctl-1 and Bxy-ctl-2 respectively, were not statistically different (p > 0.05) in the H2O2 conditions tested.
Fig 6. Survival percentage of wild-type (N2) and transgenic (KHA149 and KHA151) C. elegans after 24 hours exposition to H2O2 (H2O2 concentrations ranging between 0 and 500 μM).

Error bars represent standard deviation. Different letters above the columns indicate significant differences (p < 0.05) between C. elegans N2, KHA149 and KHA151 survival percentages in each H2O2 treatment, according to post-hoc Duncan’s test.
Discussion
Depending on their lifestyle, sedentary or migratory, plant parasitic nematodes possess different repertoires of enzymatic and non-enzymatic molecules involved in oxidative stress metabolism that are a reflection of their pathogenicity [15] In some organisms, like bacteria, these enzymes are defined as virulence factors [32]. Recent work [18] indicates that B. xylophilus has an enzymatic inventory to counterattack the pine oxidative burst. A total of 12 anti-oxidant enzymes were found in the B. xylophilus secretome, including PRX, CTL, GST, SOD, nucleoredoxin-like protein and thioredoxin [19]. In the present study, we investigated the tolerance of B. xylophilus isolates with different levels of virulence (high and low virulence) under in vitro oxidative stress conditions using H2O2 as an oxidizing agent. We were able to observe a contrasting performance between high and low virulence B. xylophilus and its relative B. mucronatus, corroborating previous results in other plant parasitic nematodes [33–34]. Emphasizing the results of the oxidative stress tolerance and H2O2 neutralization tests, the CTL (Bxy-ctl-1 and Bxy-ctl-2) gene expression indicated that high and low virulence B. xylophilus respond differently towards H2O2 stress. High virulence nematodes showed an up-regulation of CTL genes under induced oxidative stress conditions, corroborating our preliminary results [24]. In addition, since all B. xylophilus isolates share protein sequence similarity between CTLs, the contrasting responses to H2O2 stress may indicate different regulation in the detoxification of ROS. This correlation between catalase levels and in vitro H2O2 tolerance has also been described in other parasitic nematodes [35–36]. Previously, Shinya et al. [20] showed the accumulation of anti-oxidant and detoxifying enzymes on the body surface of the nematode. Here, we were able to determine that both B. xylophilus catalases (Bxy-ctl-1 and Bxy-ctl-2) are expressed in the intestine.
We also accounted for other antioxidant enzymes (SODs and GXPs) under H2O2 stress (S1 Fig), and found that, in our experimental conditions, only catalase expressions were significant. Although we could ascertain the importance of CTLs (Bxy-ctl-1 and Bxy-ctl-2) in the protection against H2O2 induced-stress, we cannot rule out the effect of other antioxidant proteins in their response. Another important observation in this study is the maintenance of CTL induction in high virulence B. xylophilus even after 24h of H2O2 exposure. Kotze [37] observed similar CTL induction in parasitic nematode Haemonchus contortus against in vitro H2O2 stress. This feature is crucial and highlights the adaptive response and antioxidant protection of high virulence B. xylophilus in host conditions. Further studies are still needed to completely understand the role of all protectant molecules (enzymatic or non-enzymatic) of B. xylophilus against the massive ROS production of susceptible pines.
The regulatory pathways of xenobiotic degradation of C. elegans are conserved in B. xylophilus [18]. Hence, We constructed transgenic C. elegans overexpressing both Bxy-ctl-1 and Bxy-ctl-2. Since we previously have shown that Bxy-ctl-1 sequence was more similar to Ce-ctl-1 and Ce-ctl-2, and Bxy-ctl-2 to Ce-ctl-3 [24], we chose C. elegans Pctl-1 to regulate Bxy-ctl-1 and C. elegans Pctl-3 to regulate Bxt-ctl-2. Our results showed that subcellular localization of both catalases is in accordance with C. elegans ctl-1 and ctl-3 promoter expression as shown by Petriv and Rachubinski [38]. Furthermore, Oláhova et al. [31] have also shown a decrease in survival rate of wild-type N2 to 1 mM H2O2 only 10 hours after treatment, which support our selection to test 50–500 μM H2O2 as experimental concentrations. C. elegans contains three catalases, Ce-ctl-1 (cytosolic), Ce-ctl-2 (peroxisomal) and Ce-ctl-3 (neurons, muscles and pharynx), which present different activities, with CTL-2 being responsible for 80% of total catalase activity [31]. Expressing exogenous B. xylophilus catalases, transgenic KHA149 and KHA151 animals were able to withstand the H2O2 stress up to 500 μM, supporting the efficiency of B. xylophilus catalases in H2O2 neutralization.
Supporting Information
Error bars represent standard deviation.
(TIF)
Acknowledgments
B. xylophilus isolates were provided by FFPRI, Tsukuba, Japan. C. elegans strains were provided by the Caenorhabditis Genetics Center (University of Minnesota, Minneapolis, MN). The authors thank to Margarida Espada (NemaLab, Universidade de Évora, Évora, Portugal) for the support in the In-situ hybridization experiments; and Dr. Mihoko Kato for linguistic correction of the manuscript.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by the Japan Society for Science Promotion (JSPS - reference P14394) postdoctoral fellowship to CSLV; the Heiwa Nakajima Foundation, international joint research grant; the European Project REPHRAME - Development of improved methods for detection, control and eradication of pine wood nematode in support of EU Plant Health policy, European Union Seventh Framework Programme FP7-KBBE-2010-4; and FEDER Funds through the Operational Programme for Competitiveness Factors - COMPETE and National Funds through FCT - Foundation for Science and Technology under the Strategic Project PEst-C/AGR/UI0115/2011. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Futai K. Pine wood nematode, Bursaphelenchus xylophilus. Ann Rev Phytopathol. 2013; 51: 61–83. 10.1146/annurev-phyto-081211-172910 [DOI] [PubMed] [Google Scholar]
- 2. Mota MM, Vieira P. Pine Wilt Disease: A Worldwide Threat to Forest Ecosystems. 2008. Netherlands: Springer. [Google Scholar]
- 3. Vicente C, Espada M, Vieira P, Mota M. Pine Wilt Disease: a threat to European forestry. Eur J Plant Pathol. 2012; 133: 89–99. [Google Scholar]
- 4. Shinya R, Takeuchi Y, Ichimura K, Takemoto S, Futai K. Establishment of a set of inbred strains of the pine wood nematode, Bursaphelenchus xylophilus (Aphelenchida: Aphelenchoididae), and evidence of their varying levels of virulence. Appl Entomol Zool. 2012; 47: 341–350. [Google Scholar]
- 5. Kiyohara T, Bolla RI. Pathogenic variability among populations of the pinewood nematode, Bursaphelenchus xylophilus. Forest Sci. 1990; 36: 1061–1076. [Google Scholar]
- 6. Aikawa T, Kikuchi T. Estimation of virulence of Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) based on its reproductive ability. Nematol. 2007; 9: 371–377. [Google Scholar]
- 7. Asai E, Futai K. Effects of inoculum density of pinewood nematode on the development of pine wilt disease in Japanese black pine seedlings pretreated with simulated acid rain. Forest Pathol. 2005; 35: 135–144. [Google Scholar]
- 8. Aikawa T, Togashi K, Kosaka H. Different developmental responses of virulent and avirulent isolates of the pine wood nematode, Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae), to the insect vector, Monochamus alternatus (Coleoptera: Cerambycidae). Environm Entomol. 2003; 32: 96–102. [Google Scholar]
- 9. Takemoto S. Population ecology of Bursaphelenchus xylophilus In: Pine Wilt Disease. 2008. Netherlands; Springer. [Google Scholar]
- 10. D’Autréaux B, Toledano MB. ROS as signalling molecules: mechanisms that generate specificity in ROS homeostasis. Nat Rev Mol Cell Biol. 2007; 10: 813–824. [DOI] [PubMed] [Google Scholar]
- 11. Quan LJ, Zhang B, Shi WS, Li HY. Hydrogen peroxide in plants: a versatile molecule of the reactive oxygen species network. J Integrative Plant Biol. 2008; 50: 2–18. [DOI] [PubMed] [Google Scholar]
- 12. Lamb C, Dixon R. The oxidative burst in plant disease resistance. Annu Rev Plant Physiol Plant Mol Biol. 1997; 48: 251–275. [DOI] [PubMed] [Google Scholar]
- 13. Dubreuil G, Deleury E, Magliano M, Jaouannet M, Abad P, et al. Peroxiredoxins from the plant parasitic root-knot nematode, Meloidogyne incognita, are required for successful development within the host. Int J Parasitol. 2011; 41: 385–396. 10.1016/j.ijpara.2010.10.008 [DOI] [PubMed] [Google Scholar]
- 14. Henkle-Dührsen K, Kampkötter A. Antioxidant enzyme families in parasitic nematodes. Mol Biochem Parasitol. 2001; 114: 129–142. [DOI] [PubMed] [Google Scholar]
- 15. Molinari S, Miacola. Antioxidant enzymes in phytoparasitic nematodes. J Nematol. 1997; 29: 153–159. [PMC free article] [PubMed] [Google Scholar]
- 16. Robertson L, Robertson WM, Sobczak M, Helder J, Tetaud E, et al. Cloning, expression and functional characterisation of a peroxiredoxin from the potato cyst nematode Globodera rostochiensis. Mol Biochem Parasitol. 2000; 111: 41–49. [DOI] [PubMed] [Google Scholar]
- 17. Bellafiore S, Shen Z, Rosso MN, Abad P, Shih P, et al. Direct identification of the Meloidogyne incognita secretome reveals proteins with host cell reprogramming potential. PLoS pathogens. 2008; 4: e1000192 10.1371/journal.ppat.1000192 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Kikuchi T, Cotton JA, Dalzell JJ, Hasegawa K, Kanzaki N, et al. Genomic insights into the origin of parasitism in the emerging plant pathogen Bursaphelenchus xylophilus. PLoS Pathogens. 2011; 7:e 1002219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Shinya R, Morisaka H, Kikuchi T, Takeuchi Y, Ueda M, et al. Secretome analysis of pine wood nematode Bursaphelenchus xylophilus reveals the tangled roots of parasitism and its potential for molecular mimicry. PloS one. 2012; 8:e 67377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Shinya R, Morisaka H, Takeuchi Y, Ueda M, Futai K. Comparison of the surface coat proteins of the pine wood nematode appeared during host pine infection and in vitro culture by a proteomic approach. Phytopathol. 2010; 100: 1289–97. [DOI] [PubMed] [Google Scholar]
- 21. Hirao T, Fukatsu E, Watanabe A. Characterization of resistance to pine wood nematode infection in Pinus thunbergii using suppression subtractive hybridization. BMC Plant Biol. 2012; 12:13 10.1186/1471-2229-12-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Santos CSS, Vasconcelos MW. Identification of genes differentially expressed in Pinus pinaster and Pinus pinea after infection with pine wood nematode. Eur J Plant Pathol. 2012; 132: 407–418. [Google Scholar]
- 23. He L, Wu X, Yu L, Ji J, Ye J. The difference of H 2 O 2 and oxidative enzyme in the interaction of different resistance pines and Bursaphelenchus xylophilus. J Nanjing Univ Natur Sci. 2010; 06. [Google Scholar]
- 24. Vicente CSL, Ikuyo Y, Mota M, Hasegawa K. Pinewood nematode-associated bacteria contribute to oxidative stress resistance of Bursaphelenchus xylophilus. BMC Microbiol. 2013; 13: 299 10.1186/1471-2180-13-299 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods. 2001; 25: 402–408. [DOI] [PubMed] [Google Scholar]
- 26. Rozen S, Skaletzki H. Primer3 on the www for general users and for biologist programmers In: Bioinformatics Methods and Protocols: Methods in Molecular Biology Totowa. 2000; Humana Press. [DOI] [PubMed] [Google Scholar]
- 27. Petersen TN, Brunak S, Heijne G, Nielsen H. SignalP 4.0: discriminating signal peptides from transmembrane regions Nat Methods. 2011; 8:785–786. 10.1038/nmeth.1701 [DOI] [PubMed] [Google Scholar]
- 28. De Boer JM, Yan Y, Smant G, Davis EL, Baum TJ. In-situ hybridization to messenger RNA in Heterodera glycines. J. Nematol. 1998; 30: 309–312. [PMC free article] [PubMed] [Google Scholar]
- 29. Kikuchi T, Jones JT, Aikawa T, Kosaka H, Ogura N. A family of glycosyl hydrolase family 45 cellulases from pine wood nematode Bursaphelenchus xylophilus. FEBS letters. 2004; 572: 201–205. [DOI] [PubMed] [Google Scholar]
- 30. Brenner S. The genetics of the nematode Caenorhabditis elegans. Genetics. 1974; 77: 71–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Oláhova M, Taylor SR, Khazaipoul S, Wang J, Morgan BA, et al. A redox-sensitive peroxiredoxin that is important for longevity has tissue- and stress-specific roles in stress resistance. PNAS. 2008; 50: 19839–19844. 10.1073/pnas.0805507105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Guo M, Block A, Bryan CD, Becker DF, Alfano AJ. Pseudomonas syringae catalases are collectively required for plant pathogenesis. J Bacteriol. 2012; 194: 5054–5064. 10.1128/JB.00999-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Molinari S, Rosso L, Ornat LC. The role of antioxidant enzymes in the virulence of root-knot nematodes on resistant tomato. Nematropica. 2005; 35: 88. [Google Scholar]
- 34. Molinari S. Antioxidant enzymes in (a) virulent populations of root-knot nematodes. Nematol. 2009; 11: 689–697. [Google Scholar]
- 35. Ou X, Thomas GR, Chacon MR, Tang L, Selkirk ME. Brugia malayi: differential susceptibility to and metabolism of hydrogen peroxide in adults and microfilariae. Exp Parasitol. 1995; 80: 530–540. [DOI] [PubMed] [Google Scholar]
- 36. Kotze AC, McClure SJ. Haemonchus contortus utilises catalase in defence against exogenous hydrogen peroxide in vitro. Int J Parasitol. 2001; 31: 1563–1571. [DOI] [PubMed] [Google Scholar]
- 37. Kotze AC. Catalase induction protects Haemonchus contortus against hydrogen peroxide in vitro. Int J Parasitol. 2003; 33: 393–400. [DOI] [PubMed] [Google Scholar]
- 38. Petriv OI, Rachubinski RA. Lack of peroxisomal catalase causes a progeric phenotype in Caenorhabditis elegans. J Biol Chem. 2004; 279: 19996–20001. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Error bars represent standard deviation.
(TIF)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.