

Abstract
The cell adhesion glycoproteins, TgrB1 and TgrC1, are essential for Dictyostelium development and allorecognition, but it has been impossible to determine whether their pleiotropic roles are due to one common function or to distinct functions in separate pathways. Mutations in the respective genes, tgrB1 and tgrC1, abrogate both development and allorecognition and the defects cannot be suppressed by activation of the cyclic AMP dependent protein kinase PKA, a central regulator of Dictyostelium development. Here we report that mutations in genes outside the known PKA pathway partially suppress the tgrC1-null developmental defect. We separated the pleiotropic roles of tgrC1 by testing the effects of a suppression mutation, stcinsA under different conditions. stcAins modified only the developmental defect of tgrC1– but not the allorecognition defect, suggesting that the two functions are separable. The suppressor mutant phenotype also revealed that tgrC1 regulates stalk differentiation in a cell-autonomous manner and spore differentiation in a non-cell-autonomous manner. Moreover, stcAins did not modify the developmental defect of tgrB1–, but the less robust phenotype of tgrB1– obscures the possible role of stcA relative to tgrB1.
Introduction
The social soil amoeba Dictyostelium discoideum is used as a model system for the study of development, cell type differentiation and the evolution of multicellularity. During vegetative growth, the amoebae propagate as single cells. Upon starvation, they stop dividing, aggregate into multicellular structures that contain thousands of cells and develop into fruiting bodies composed of two cell types, the reproductive spores and the somatic stalks. The developmental process takes about 24 hours. In the first quarter of development, the cells entrain each other using extracellular cAMP. They then begin to aggregate and after 8–10 hours they form mounds, while starting to differentiate into the two major cell types. At 16 hours, the mound is transformed into a slug, in which the different cell types have sorted out into designated regions along the long axis of the slug – prestalk cells mostly in the front and prespore cells in the back. Eventually, the slug erects itself and the prestalk cells descend through the prespore cell mass as both cell types undergo terminal differentiation into stalks and spores.
TgrC1 has several important functions in development. TgrC1 is a single-pass transmembrane glycoprotein with a large extracellular domain and a short cytoplasmic tail. It was first identified as an EDTA-insensitive cell-cell adhesion molecule [1,2]. Adhesion is a critical component in multicellularity because it holds the cells together and often participates in signaling. TgrC1 expression starts at the mound stage and peaks at the slug stage [1]. It accounts for about 50% of the total adhesion at the slug stage [3] and its adhesion partner is TgrB1, another single-pass transmembrane glycoprotein [4]. Mutants lacking tgrC1 function are developmentally arrested at the loose aggregate stage and exhibit defects in differentiation of both cell types, suggesting that TgrC1 has a role in controlling differentiation [5]. Self-recognition is an important social feature of Dictyostelium. When strains with different genetic backgrounds develop together, the cells tend to sort out in aggregates and undergo further development in clonal aggregates [6]. TgrB1 and TgrC1 are necessary and sufficient to exclusively mediate this process. When two strains have matching TgrB1/TgrC1, they aggregate and co-develop regardless of their genetic backgrounds. Similarly, when two strains have non-matching TgrB1/TgrC1, they sort out and develop clonally even if the rest of the genome is identical [7]. TgrC1 and TgrB1 participate in adhesion and in signaling, both in development and in social recognition, but it is not clear whether the functions are overlapping or distinct.
We used a genetic screen for suppressors of the tgrC1 - phenotype to begin to address that question. We surmised that such suppressors would uncover genetic components in specific pathways. If such suppressors would modify only one of the phenotypes but not another, or if they modify only the tgrC1 - phenotype but not the tgrB1 - phenotype, then the functions must be distinct. We could use such suppressors as probes to dissect the multiple functions of TgrC1 under different conditions, and such suppressors may also provide an opportunity to differentiate the functions of TgrB1 and TgrC1 in development. We generated random mutations in a tgrC1 - strain and selected for strains that carried a second-site mutation and were able to sporulate. We identified three suppressors of tgrC1 that partially restored the function in development and thus in cell-type differentiation. We further characterized one of the suppressors, stcA ins, and tested its effect on self recognition and on cell-type differentiation to reveal the regulatory functions of TgrC1. The results led us to propose that TgrC1 utilizes different pathways to mediate differentiation and self recognition, and that the self recognition function is separable from the differentiation function. Moreover, TgrC1 regulates the differentiation of the two cell types differently. It regulates stalk formation in a cell-autonomous way and sporulation in a non-cell-autonomous way. We also found that TgrC1 and TgrB1 have distinct roles in signal transduction. We propose that TgrC1 participates in a critical control point in Dictyostelium development, allowing the cells to integrate inputs from both differentiation signals and from self recognition signals and to mount the proper responses.
Results
Suppressor mutations partially rescue the developmental defect of tgrC1-
We used restriction enzyme mediated insertion (REMI) to perform a screen for suppressors of the tgrC1 - phenotype. During development in a pure population, tgrC1 - is arrested at the loose aggregate stage. It does not form fruiting bodies and produces very few spores [5]. We therefore selected for suppressor mutations that produced viable spores that could germinate, grow in association with bacteria and develop into fruiting bodies. We screened a total of 8,000 insertion mutants and identified three independent suppressor strains in which we identified three genes that have not been characterized before. We named them stcA (DDB_G0267562), stcB (DDB_G0270410) and stcC (DDB_G0274171), where stc stands for suppressor of t gr C 1 -. The screen was not carried to saturation. To make sure that the suppression was due to the mutations we discovered, we recapitulated the mutations in a fresh tgrC1 - background and observed the same suppression phenotype (S1 Fig and data not shown).
We developed the three suppressor strains and compared their development on non-nutrient agar with those of the laboratory wild type, AX4, and the parental tgrC1 - cells (Fig 1A). All three suppressor strains developed past the loose aggregate stage, but they all had developmental defects compared with the wild type. After the loose aggregate stage, the aggregate size of all strains was small compared to AX4. In addition, they all formed fruiting bodies eventually, but none of them formed fruiting bodies at 24h and their development was rather unsynchronized. We quantified the sporulation efficiency of these strains and found that in all three cases it was significantly higher than the parental tgrC1 - strain but lower than that of the wild type AX4 cells (Fig 1B). In summary, the three mutations we discovered were able to partially suppress the developmental defect of tgrC1 -, as indicated by advanced morphology and improved sporulation. We further characterized the effects of the mutation in the stcA gene, which encodes a putative protein of 557 amino acids without any distinctive features. The insertion site in stcA occurred at base pair 1718, toward the 3’ end of the gene (Fig 1C). We tested the expression of the disrupted stcA mRNA by RT-PCR and found that the region upstream of the insertion site was expressed at levels comparable or slightly higher than the wild type and the parental tgrC1 - strain (S2A Fig). The region overlapping the insertion site was equally abundant in the wild type and the parental tgrC1 - strain, but no RT-PCR product was observed in the tgrC1 - stcA ins mutant, validating that this region was disrupted by the insertion (S2B Fig). These results indicate that the insertion disrupted the gene and the resulting mRNA, suggesting that the protein may have been disrupted as well.
Fig 1. tgrC1− development is partially restored by suppressor mutations.

A. We developed the suppressor strains (tgrC1−stcAins, tgrC1−stcBins and tgrC1−stcCins), the laboratory wild-type strain AX4 and the parental tgrC1− strain on non-nutrient agar. We photographed the cells with light microscopy from above at 16 hour, 24 hour and 72 hour as indicated (strain genotypes are indicated on the left). Scale bar, 1 mm. B. We determined the sporulation efficiency of the same strains at 72 hour of development on black filters. The results are shown as the average (n = 3) and s.d. of the fraction of cells that made spores normalized to the wild type AX4 (%, y-axis). Strain genotypes are indicated on the bottom. C. An illustration of the stcA gene and the insertional mutation. The thick red line represents the two exons and the thin angled lines represent the single intron. The triangle represents the pBSR1 plasmid, which was inserted at the 3’-end of the gene. The inserted plasmid is not drawn to scale. Numbers (bp) below the lines indicate the beginning and the end of the gene model, the predicted splicing borders and the insertion site.
tgrC1 - has a cell-autonomous defect in spore differentiation, and a non-cell-autonomous defect in stalk differentiation
The tgrC1 - mutation affects several developmental processes, including aggregation and cell-type differentiation and some of these effects are non-cell autonomous [5,7–9]. In order to examine the mode of suppression in the tgrC1 - stcA ins strain, we mixed equal proportions of tgrC1 - with AX4 or with tgrC1 - stcA ins, developed them and determined the proportions of the strains in the resulting spores (Fig 2A). When tgrC1 - cells were mixed with the wild type AX4, about 10% of the spores in the fruiting bodies had the tgrC1 - genotype, consistent with previous findings that the tgrC1 - phenotype has a weak non-cell autonomous property. In the mix of tgrC1 - and tgrC1 - stcA ins we found about 70% tgrC1 - spores, suggesting that the stcA ins mutation enhances sporulation in a non-cell-autonomous way in both tgrC1 - and tgrC1 - stcA ins. One possibility is that tgrC1 - formed more spores than its fair share in the chimera by avoiding contribution to stalks. To examine that possibility we labeled the strains separately with either Green- or Red-fluorescence proteins. We mixed the cells at equal proportions, developed them, and tracked their location in the mixed aggregates during development. At 16 hours of development, we observed that the wild-type AX4 cells formed slugs and left the tgrC1 - aggregates behind, while the tgrC1 - stcA ins cells remained mixed with tgrC1 - and formed chimeric slugs. Interestingly, the cells were not evenly mixed in the chimeric slugs—the prestalk region in the slug anterior contained predominantly tgrC1 - stcA ins cells (Fig 2B). Following development of the tgrC1 - stcA ins and tgrC1 - chimerae to fruiting body formation revealed that the stalks were predominately composed of tgrC1 - stcA ins cells and the spore-containing sori consisted of mostly tgrC1 - cells (Fig 2C). These observations suggest that tgrC1 - formed more spores than its fair share due to its inability to form stalks. This was not the case in the mixes with the wild type, because the wild type segregated from the tgrC1 - cells. Therefore, the inability of tgrC1 - stcA ins to segregate from tgrC1 - contributed to the increased proportion of tgrC1 - in the spores. In addition, the absence of tgrC1 - from the stalks of the chimeric fruiting bodies suggests that the stalk-differentiation defect of tgrC1 - is cell-autonomous.
Fig 2. The tgrC1 - sporulation defect is non-cell autonomous and the stalk development defect is cell-autonomous.

A. We mixed tgrC1− with tgrC1−stcAins or AX4, as indicated, at 1:1 ratios, and developed the mixture for 72 hours. We collected the spores, germinated them in association with Klebsiella aerogenes to allow single plaque formation and recorded the fraction (%) of tgrC1− plaques relative to the total number of plaques. The average fraction and the s.d. are shown (y-axis; n = 3). The overall sporulation efficiency of the tgrC1− and tgrC1−stcAins strains was similar to that of tgrC1−stcAins alone (Fig 1B). The overall sporulation efficiency of the mix between tgrC1− and AX4 was about half of that of AX4 alone (Fig 1B). We mixed GFP-labeled tgrC1− (green text) with RFP-labeled AX4 or tgrC1−stcAins (red text) and developed the mixture of cells on non-nutrient agar. We photographed the structures at 16 hour (B) and single fruiting bodies of GFP-labeled tgrC1− and RFP-labeled tgrC1−stcAins at 48 hour (C). Scale bar, 0.2 mm.
stcA ins does not modify the phenotype of tgrB1 -
The observation that tgrC1 and tgrB1 function in a common signaling pathway [4,7,8] prompted us to test whether the stcA ins mutation could suppress the tgrB1 - phenotype as well. We generated the stcA ins mutation in tgrB1 - cells and followed the developmental morphology and sporulation efficiency of the resulting tgrB1 - stcA ins strain in comparison with the parental tgrB1 - strain and the wild type, AX4. We observed no significant difference between the morphologies of tgrB1 - stcA ins and tgrB1 - strains at any time point (Fig 3A). We also found no significant difference in the sporulation efficiency (Fig 3B). The different behavior of the stcA ins mutation in the tgrB1 - background, compared to the tgrC1 - background, suggests that stcA has a limited effect on the TgrB1-controlled developmental pathway.
Fig 3. stcAins does not modify the tgrB1− phenotype.
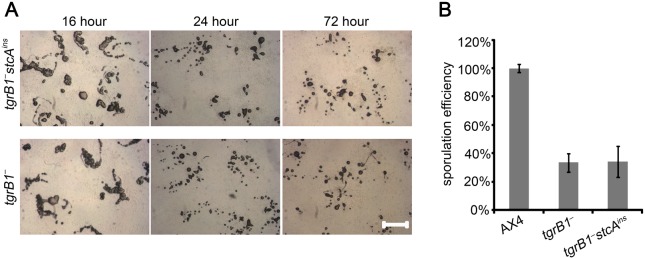
A. We developed tgrB1− and tgrB1−stcAins on non-nutrient agar and photographed the structures from above with light microscopy at 16 hour, 24 hour and 72 hour as indicated on the top. Strain genotypes are indicated on the left. Scale bar, 0.2 mm. B. We measured the sporulation efficiency of tgrB1− and tgrB1−stcAins at 72 hour. The results are shown as the average and s.d. of the fraction of cells (%, y-axis; n = 3) that made spores scaled to the wild type AX4. Strain genotypes are indicated on the bottom.
stcA ins does not modify the kin-recognition function of tgrC1 -
In addition to a critical role in development, tgrC1 has a role in self recognition [7,8]. It was therefore interesting to test whether the stcA ins mutation had an effect on that role of tgrC1 as well. We generated the stcA ins mutation in an otherwise wild type background and also in the double gene replacement strain tgrB1 QS31 tgrC1 QS31, which segregates from the parental AX4 strain due to the different tgrB1-tgrC1 allotype [7,8]. We labeled the strains with either GFP or RFP and mixed them in equal proportions with several test strains to assess self recognition. In Fig 4A, stcA ins was recapitulated in the tgrC1 - background, and the resulting double mutant tgrC1 - stcA ins behaved like tgrC1 - in chimerae with the wild type – both strains segregated from the wild type, suggesting that the stcA ins mutation does not suppress the self-recognition defect conferred by the tgrC1 - mutation. Similarly, when stcA ins was recapitulated in the AX4 background, the resulting mutant segregated from tgrC1 - (Fig 4B), but associated with AX4 (Fig 4C), a behavior that was indistinguishable from that of the wild type, AX4. We also recapitulated stcA ins in tgrB1 QS31 tgrC1 QS31. The resulting strain was indistinguishable from the parental tgrB1 QS31 tgrC1 QS31, segregating from AX4 in chimerae (Fig 4D). These experiments indicate that the developmental function of tgrC1 is separable from its function in kin recognition, suggesting that tgrC1 has distinct functions in development and in kin recognition.
Fig 4. stcAins does not modify kin recognition.
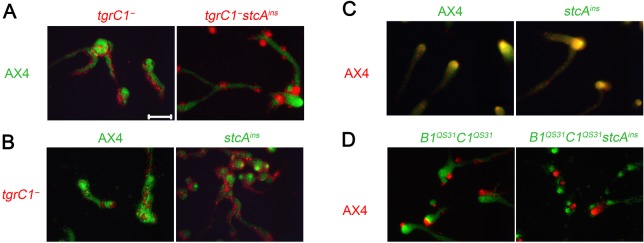
We labeled cells with GFP (green text) or RFP (red text), grew them independently, mixed two strains at 1:1 ratios and allowed them to develop together on non-nutrient agar. Photographs were taken at 10 hour of development. Cells with the genotypes indicated on the left side of each row were mixed with cells of different genotypes as indicated above each panel. Scale bar, 0.2 mm.
Discussion
The cell-cell adhesion glycoprotein TgrC1 [10] is essential for development [5,9] and for kin recognition [7,8]. Inactivation of the tgrC1 gene causes a developmental arrest at the loose aggregate stage and a subsequent inability to differentiate prespore and prestalk cells [5]. Many developmental defects of this type can be suppressed by constitutive activation of PKA. For example, defects in tagC and dhkA cause developmental arrests at different developmental stages, but both can be suppressed by activation of PKA [11]. Surprisingly, the defect of tgrC1 - cannot be suppressed by constitutive expression of PKA [12], suggesting that TgrC1 is involved in a parallel signaling pathway. Our genetic screen identified three suppressor mutations, all of which partially restore the ability of tgrC1 - cells to form fruiting bodies and spores. The suppressor genes have not been identified before, suggesting that they are not directly involved in the well-characterized PKA pathway. These findings support the idea that TgrC1 is involved in a signaling pathway independent of the central PKA pathway in Dictyostelium development.
TgrC1 has a role in cell-cell communications during development [9]. This finding focused our interest on the stcA ins mutation because it distinguished the regulatory roles of tgrC1 in stalk and spore differentiation. Specifically, mixing tgrC1 - cells with AX4 (wild type) cells resulted in the production of very few tgrC1 - spores but mixing tgrC1 - cells with the suppressor strain tgrC1 - stcA ins resulted in a majority of tgrC1 - spores. We also observed that tgrC1 - cells segregate from AX4 cells but not from the suppressor tgrC1 — stcA ins. Based on these findings, we propose that tgrC1 - cells have a non-cell autonomous defect in spore production, probably because they are unable to produce an extracellular signal necessary for that process. We propose that both AX4 and tgrC1 - stcA ins cells have the ability to provide tgrC1 - cells with the missing signal, but the segregation of AX4 from the tgrC1 - cells limits the access of the tgrC1 - cells to the signal. Our findings also suggest that tgrC1 participates in stalk differentiation in a cell-autonomous manner because the tgrC1 - cells did not participate in the stalk in any of the mixing experiments. This observation is consistent with the finding that tgrC1 mRNA is preferentially expressed in prestalk cells at late stages of development [9]. However, the suppression of tgrC1 - by stcA ins is relatively weak, suggesting that other genetic components are involved in controlling differentiation and development. We have not captured these components in our genetic screen either because we did not carry the screen to near saturation or because the insertional mutagenesis method we used cannot reveal these genes.
Mutations that have different effects on cell-type differentiation have been reported before. For example, prestalk-specific expression of pkaRm, a dominant negative regulator of PKA activity, results in a cell-autonomous defect in stalk formation and a non-cell autonomous defect in sporulation [13]. Similarly, inactivation of the prestalk specific gene tagB results in a cell-autonomous defect in stalk formation and a non-cell autonomous defect in sporulation [14]. Subsequent studies have shown that prespore and prestalk cells are engaged in an elaborate signaling cascade during terminal differentiation [15]. Both pkaRm and tagB affect the PKA signaling cascade that regulates terminal differentiation. Since activation of PKA does not suppress the tgrC1 - defect, it is quite possible that tgrC1 and stcA regulate terminal differentiation in a PKA-independent manner.
Since tgrC1 is involved in multiple functions, the phenotype of tgrC1 - at any given time in development is a sum over all its current and previous functions. We would need to integrate the separate functions of adhesion, developmental control and kin-recognition to account for the cells’ behaviors during development, as evidenced by the tgrC1 - spore proportion difference between the chimeric development of tgrC1 - with AX4 and that with tgrC1 - stcA ins. Due to segregation, there is only a small fraction of tgrC1 - cells in the AX4 chimeric aggregates whereas there are 50% of tgrC1 - cells in the tgrC1 - stcA ins chimeric aggregates, partly accounting for the significant difference of the final tgrC1 - spore proportion. In this report we have shown that stalk differentiation of tgrC1 - is defective in a cell-autonomous way. However previous findings have demonstrated that stalk differentiation is partially regulated by the extracellular signaling molecule DIF-1 [16,17]. Others have shown that some tgrC1 - cells could be observed in the stalks of chimeric fruiting bodies of tgrC1 - and AX4 [9]. These findings support a non-cell-autonomous regulation of stalk differentiation. We argue that since AX4 segregates from tgrC1 -, the abundance of tgrC1 - cells in either stalks or sori in the final fruiting bodies is low, as indicated by the small number of spores of tgrC1 - observed in the final spore mass [9]. Therefore, the overall rescue of tgrC1 - stalk differentiation in the chimerae is limited, suggesting that the non-cell-autonomous regulation of stalk differentiation is weak. In addition, differential adhesion and sorting are likely components of cell-type differentiation and morphogenesis. In mixtures with AX4 cells, the tgrC1 - cells are likely carried into the aggregates due to incomplete segregation, whereas in mixtures with tgrC1 - stcA ins, the tgrC1 - cells are included in the aggregate but excluded from the prestalk region. It is likely that the different segregation patterns in the different contexts subject the cells to different signals and hence to different fates in subsequent stages of development. The fact that most of the tgrC1 - cells are absent from the stalks without segregation, and the finding that tgrC1 - stcA ins cells were able to make proper stalks suggest that tgrC1 - has a direct role in stalk differentiation.
PKA is a central regulator of Dictyostelium development and cAMP signals are executed through PKA activation at every stage of development, as demonstrated by the finding that over-expression of PKA-C bypasses the need for cAMP throughout development [18]. In this report, we propose that tgrC1 acts as another important regulator of Dictyostelium development. Unlike the ubiquitous role of PKA throughout development, the findings in this report suggest that tgrC1 integrates important functions such as differentiation, adhesion and kin recognition, into a critical time at which the transition from unicellularity to multicellularity takes place and cell-type differentiation initiates.
One of the most important findings we made is that the role of tgrC1 in kin recognition is separable from its role in development. We provided further evidence of the separation of function by showing that the stcA ins mutation did not modify the segregation behaviors of other related strains. Moreover, stcA ins did not affect the phenotypes of other mutants, suggesting that tgrC1 may have a separate role in kin recognition that is not entirely overlapping with its role in differentiation. Although the function of stcA is not known yet and its sequence does not provide clues in that regard, the mutation and its phenotype are still very informative with respect to the functions of tgrC1, which was the goal of this study.
The TgrB1 and TgrC1 proteins may work as a pair in both development and kin recognition [7,8]. We have shown that a suppressor of tgrC1 - does not alter the phenotype of tgrB1 -. The result is inconclusive because tgrB1 - phenotype is less robust than the tgrC1 - phenotype in terms of both sporulation efficiency and developmental morphology. The insignificant difference of sporulation efficiency between tgrB1 - and tgrB1 - stcA ins could be due to weak suppression compounded with the variability of the tgrB1 - phenotype, or due to the lack of suppression at all.
Materials and Methods
Cell growth, development and transformation
We grew the cells in shaking suspension in HL-5 medium, supplemented with 10μg/mL G418, 10μg/mL Blasticidin S or 20μg/mL uracil, as necessary. We allowed the cells to grow for 24 hours without drugs before the experiments. We harvested the cells at the logarithmic growth phase and developed them on nitrocellulose filters [19] or on non-nutrient agar [20] as previously described. We carried out plasmid transformations as described [6].
Mutagenesis, selection and identification of the mutated genes
We performed restriction enzyme mediated insertion (REMI) as described [21]. We collected and pooled Blasticidin S resistant cells from four transformations after 6 days of drug selection, spread 5 x 105 cells on each 10 cm SM agar plates in association with Klebsiella aerogenes. Immediately after the amoebae cleared the bacterial lawns, we harvested the cells and vortexed for 1 min in 20 mM potassium phosphate buffer (pH 6.2) with 0.1% NP40 to select for spores. We plated about 100 spores on each 10 cm SM agar plate in association with Klebsiella aerogenes to allow clonal germination, growth and development. We identified single plaques that contained any fruiting bodies as suppressor mutants of tgrC1 -. We performed plasmid rescue as described to identify the mutated genes [22]. We cloned stcA with SpeI, stcB with ClaI, and stcC with EcoRI. We identified the genes by sequencing the insertion sites with SP6 and T7 primers.
Nucleic acid manipulation
We prepared genomic DNA as described [23] and performed Southern blot analysis as described [24]. We made the Southern blot probes by PCR. We used the pBSR1 plasmid [21] as the template for Blasticidin S resistance (BSR) cassette and AX4 genomic DNA for all the gene probes. BSR cassette probe primers: 5’-AGTAGAAGTAGCGACAGAGAAGA-3’, 5’-TGGCTGTTTTACATCTAATGC-3’; stcA probe primers: 5’-CTTATTGTCTTGTTTCATGTCCTGG-3’, 5’-caatatgaccattttcttcaatgtg-3’; stcB probe primers: 5’- GGTGGTTTTTGATGTTGATG-3’, 5’-CATTATTATCATCCTCTTCATCAC-3’; stcC probe primers: 5’-CATTGAATTGCACTGCATTCTC-3’, 5’-GAACCCCACTAATTTAAATTTACTGC-3’. We cloned the insertion sits as described [21].
Construction of new strains and labeling with fluorescence
We used the rescue plasmids to recapitulate the insertional mutations in different backgrounds. We used the pA15/tdTomato and pDXA-GFP2 [8] to label strains with RFP and GFP respectively.
Segregation assay
We performed segregation assays as described [8]. Briefly, we grew the cells in pure populations, harvested them, mixed two strains at 1:1 ratio and developed them on non-nutrient agar. We photographed the developmental structures with fluorescence microscopy and merged the images without further manipulation.
Sporulation efficiency
We grew and developed the cells as indicated above. We collected the spores after 72 hours by vortexing the entire filters for 1 min in 20 mM potassium phosphate buffer (pH 6.2) with 0.1% NP40. We then counted the spores. Sporulation efficiency was calculated as the fraction (percent) of spores relative to the number of cells at the start of development. The sporulation efficiency test of each strain contained three independent biological replicates. Each biological replicate included three technical replicates. We averaged (mean) the sporulation efficiencies of the three technical replicates of each strain for every biological replicate, and then normalized the efficiency of every strain to wild type AX4, which was performed in parallel. The final sporulation efficiency was calculated by averaging the data from the three biological replicates.
RNA extraction and reverse transcription PCR
We performed the experiments as described [8]. We collected RNAs at 20 hours of development for all strains tested. The primer set we used to amplify the stcA fragment upstream the insertion site were: 5’-CCACCACCATTATCAACTAG-3’, 5’-GATCATAAAAACCAGGACATG-3’. The primer set we used to amplify the fragment spanning the insertion site were: 5’-CCACCACCATTATCAACTAG-3’, 5’-TTACTTAAATGCATTAAAATCAA-3’.
Supporting Information
We prepared genomic DNAs from all the strains tested (relevant genotypes are indicated above the lanes; ‘suppressor’ represents the original suppressor strain), digested the DNA samples with SpeI, and separated the fragments on 0.8% agarose gel. After transferring the DNAs to nitrocellulous membranes, we processed the blots with probes against stcA or against the blasticidin S resistance cassette (BSR) as indicated below the lanes. MW – molecular weight markers (Kb). Arrows point to the wild type (WT) and mutant (ins) stcA bands. The SpeI fragment of stcA gene is about 2.2 Kb, and the pBSR1 insertion size is approximately 4.1 Kb.
(TIF)
We developed the strains tgrC1 - stcA ins, tgrC1 - and AX4 for 20 hour, which is the peak expression time of stcA in AX4. We collected RNA samples and reverse transcribed the mRNA. Using the resulting cDNA, we PCR-amplified one fragment 5’ of the insertion site (A) and another fragment across the insertion site (B). Each reaction was performed with and without reverse transcriptase (RT) to control for genomic DNA contamination. The reactions were loaded side by side where “+” indicates reactions with RT, and “-” indicates reactions without RT. The relevant genotypes are indicated above the lanes. MW – molecular weight marker (base pairs). The green boxes below the gene models represent the target RT-PCR regions in each experiment. The dotted lines represent the start and the end of the RT-PCR products. The blue line represents the insertion site.
(TIF)
Data Availability
All relevant data are within the paper.
Funding Statement
This work was funded by grant number R01 GM084992 from the National Institute of General Medical Sciences (http://www.nigms.nih.gov). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Geltosky JE, Weseman J, Bakke A, Lerner RA (1979) Identification of a cell surface glycoprotein involved in cell aggregation in D. discoideum. Cell 18: 391–398. Available: http://www.ncbi.nlm.nih.gov/pubmed/498273. Accessed 2014 September 8. [DOI] [PubMed] [Google Scholar]
- 2. Wang J, Hou L, Awrey D, Loomis WF, Firtel RA, Siu CH (2000) The membrane glycoprotein gp150 is encoded by the lagC gene and mediates cell-cell adhesion by heterophilic binding during Dictyostelium development. Dev Biol 227: 734–745. Available: http://www.ncbi.nlm.nih.gov/pubmed/11071787. Accessed 2014 September 15. [DOI] [PubMed] [Google Scholar]
- 3. Gao EN, Shier P, Siu CH (1992) Purification and partial characterization of a cell adhesion molecule (gp150) involved in postaggregation stage cell-cell binding in Dictyostelium discoideum. J Biol Chem 267: 9409–9415. Available: http://www.ncbi.nlm.nih.gov/pubmed/1577768. Accessed 2014 September 15. [PubMed] [Google Scholar]
- 4. Chen G, Wang J, Xu X, Wu X, Piao R, Siu CH (2013) TgrC1 mediates cell-cell adhesion by interacting with TgrB1 via mutual IPT/TIG domains during development of Dictyostelium discoideum. Biochem J 452: 259–269. Available: http://www.ncbi.nlm.nih.gov/pubmed/23477311. Accessed 2014 September 15. 10.1042/BJ20121674 [DOI] [PubMed] [Google Scholar]
- 5. Dynes JL, Clark AM, Shaulsky G, Kuspa A, Loomis WF, Firtel RA (1994) LagC is required for cell-cell interactions that are essential for cell-type differentiation in Dictyostelium. Genes Dev 8: 948–958. Available: http://www.ncbi.nlm.nih.gov/pubmed/7926779. Accessed 2014 September 15. [DOI] [PubMed] [Google Scholar]
- 6. Ostrowski EA, Katoh M, Shaulsky G, Queller DC, Strassmann JE (2008) Kin discrimination increases with genetic distance in a social amoeba. PLoS Biol 6: e287 Available: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2586364&tool=pmcentrez&rendertype=abstract. Accessed 2014 September 15. 10.1371/journal.pbio.0060287 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Hirose S, Benabentos R, Ho H-I, Kuspa A, Shaulsky G (2011) Self-recognition in social amoebae is mediated by allelic pairs of tiger genes. Science 333: 467–470. Available: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3142563&tool=pmcentrez&rendertype=abstract. Accessed 2014 September 15. 10.1126/science.1203903 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Benabentos R, Hirose S, Sucgang R, Curk T, Katoh M, Ostrowski EA et al. (2009) Polymorphic members of the lag gene family mediate kin discrimination in Dictyostelium. Curr Biol 19: 567–572. Available: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2694408&tool=pmcentrez&rendertype=abstract. Accessed 2014 September 15. 10.1016/j.cub.2009.02.037 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Kibler K, Svetz J, Nguyen TL, Shaw C, Shaulsky G (2003) A cell-adhesion pathway regulates intercellular communication during Dictyostelium development. Dev Biol 264: 506–521. Available: http://linkinghub.elsevier.com/retrieve/pii/S0012160603005256. Accessed 2014 September 8. [DOI] [PubMed] [Google Scholar]
- 10. Siu C-H, Harris TJC, Wang J, Wong E (2004) Regulation of cell-cell adhesion during Dictyostelium development. Semin Cell Dev Biol 15: 633–641. Available: http://www.ncbi.nlm.nih.gov/pubmed/15561583. Accessed 2014 September 15. [DOI] [PubMed] [Google Scholar]
- 11. Anjard C, Zeng C, Loomis WF, Nellen W (1998) Signal transduction pathways leading to spore differentiation in Dictyostelium discoideum. Dev Biol 193: 146–155. Available: http://www.ncbi.nlm.nih.gov/pubmed/9473320. Accessed 2014 September 15. [DOI] [PubMed] [Google Scholar]
- 12. Iranfar N, Fuller D, Loomis WF (2006) Transcriptional regulation of post-aggregation genes in Dictyostelium by a feed-forward loop involving GBF and LagC. Dev Biol 290: 460–469. Available: http://www.ncbi.nlm.nih.gov/pubmed/16386729. Accessed 2014 September 10. [DOI] [PubMed] [Google Scholar]
- 13. Harwood AJ, Early A, Williams JG (1993) A repressor controls the timing and spatial localisation of stalk cell-specific gene expression in Dictyostelium. Development 118: 1041–1048. Available: http://www.ncbi.nlm.nih.gov/pubmed/8269839. Accessed 2014 September 15. [DOI] [PubMed] [Google Scholar]
- 14. Shaulsky G, Kuspa A, Loomis WF (1995) A multidrug resistance transporter/serine protease gene is required for prestalk specialization in Dictyostelium. Genes Dev 9: 1111–1122. Available: http://www.ncbi.nlm.nih.gov/pubmed/7744252. Accessed 2014 September 15. [DOI] [PubMed] [Google Scholar]
- 15. Anjard C, Su Y, Loomis WF (2011) The polyketide MPBD initiates the SDF-1 signaling cascade that coordinates terminal differentiation in Dictyostelium. Eukaryot Cell 10: 956–963. Available: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3147415&tool=pmcentrez&rendertype=abstract. Accessed 2014 September 15. 10.1128/EC.05053-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Kay RR, Jermyn KA (1983) A possible morphogen controlling differentiation in Dictyostelium. Nature 303: 242–244. Available: http://www.ncbi.nlm.nih.gov/pubmed/6843672. Accessed 2014 September 15. [DOI] [PubMed] [Google Scholar]
- 17. Thompson CRL, Kay RR (2000) The role of DIF-1 signaling in Dictyostelium development. Mol Cell 6: 1509–1514. [DOI] [PubMed] [Google Scholar]
- 18. Wang B, Kuspa A (1997) Dictyostelium development in the absence of cAMP. Science 277: 251–254. Available: http://www.ncbi.nlm.nih.gov/pubmed/9211856. Accessed 2014 September 15. [DOI] [PubMed] [Google Scholar]
- 19. Shaulsky G, Loomis WF (1993) Cell type regulation in response to expression of ricin A in Dictyostelium. Dev Biol 160: 85–98. Available: http://www.ncbi.nlm.nih.gov/pubmed/8224551. Accessed 2014 September 15. [DOI] [PubMed] [Google Scholar]
- 20. Huang E, Blagg SL, Keller T, Katoh M, Shaulsky G, Thompson CR (2006) bZIP transcription factor interactions regulate DIF responses in Dictyostelium. Development 133: 449–458. Available: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3531922&tool=pmcentrez&rendertype=abstract. Accessed 2014 September 15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Shaulsky G, Escalante R, Loomis WF (1996) Developmental signal transduction pathways uncovered by genetic suppressors. Proc Natl Acad Sci U S A 93: 15260–15265. Available: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=26391&tool=pmcentrez&rendertype=abstract. Accessed 2014 September 15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Kuspa A, Vollrath D, Cheng Y, Kaiser D (1989) Physical mapping of the Myxococcus xanthus genome by random cloning in yeast artificial chromosomes. Proc Natl Acad Sci U S A 86: 8917–8921. Available: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=298401&tool=pmcentrez&rendertype=abstract. Accessed 2014 September 15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Katoh M, Curk T, Xu Q, Zupan B, Kuspa A, Shaulsky G (2006) Developmentally regulated DNA methylation in Dictyostelium discoideum. Eukaryot Cell 5: 18–25. Available: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1360260&tool=pmcentrez&rendertype=abstract. Accessed 2014 September 15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Vollrath D, Davis RW, Connelly C, Hieter P (1988) Physical mapping of large DNA by chromosome fragmentation. Proc Natl Acad Sci U S A 85: 6027–6031. Available: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=281898&tool=pmcentrez&rendertype=abstract. Accessed 2014 September 15. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
We prepared genomic DNAs from all the strains tested (relevant genotypes are indicated above the lanes; ‘suppressor’ represents the original suppressor strain), digested the DNA samples with SpeI, and separated the fragments on 0.8% agarose gel. After transferring the DNAs to nitrocellulous membranes, we processed the blots with probes against stcA or against the blasticidin S resistance cassette (BSR) as indicated below the lanes. MW – molecular weight markers (Kb). Arrows point to the wild type (WT) and mutant (ins) stcA bands. The SpeI fragment of stcA gene is about 2.2 Kb, and the pBSR1 insertion size is approximately 4.1 Kb.
(TIF)
We developed the strains tgrC1 - stcA ins, tgrC1 - and AX4 for 20 hour, which is the peak expression time of stcA in AX4. We collected RNA samples and reverse transcribed the mRNA. Using the resulting cDNA, we PCR-amplified one fragment 5’ of the insertion site (A) and another fragment across the insertion site (B). Each reaction was performed with and without reverse transcriptase (RT) to control for genomic DNA contamination. The reactions were loaded side by side where “+” indicates reactions with RT, and “-” indicates reactions without RT. The relevant genotypes are indicated above the lanes. MW – molecular weight marker (base pairs). The green boxes below the gene models represent the target RT-PCR regions in each experiment. The dotted lines represent the start and the end of the RT-PCR products. The blue line represents the insertion site.
(TIF)
Data Availability Statement
All relevant data are within the paper.