

Abstract
The majority of individuals exposed to hepatitis C virus (HCV) establish a persistent infection, which is a leading cause of chronic liver disease, cirrhosis and hepatocellular carcinoma. Major progress has been made during the past twenty-five years in understanding the HCV life cycle and immune responses against HCV infection. Increasing evidence indicates that host genetic factors can significantly influence the outcome of HCV infection and the response to interferon alpha-based antiviral therapy. The arrival of highly effective and convenient treatment regimens for patients chronically infected with HCV has improved prospects for the eradication of HCV worldwide. Clinical trials are evaluating the best anti-viral drug combination, treatment doses and duration. The new treatments are better-tolerated and have shown success rates of more than 95%. However, the recent breakthrough in HCV treatment raises new questions and challenges, including the identification of HCV-infected patients and to link them to appropriate health care, the high pricing of HCV drugs, the emergence of drug resistance or naturally occurring polymorphism in HCV sequences which can compromise HCV treatment response. Finally, we still do not have a vaccine against HCV. In this concise review, we will highlight the progress made in understanding HCV infection and therapy. We will focus on the most significant unsolved problems and the key future challenges in the management of HCV infection.
Keywords: Pathogenesis, Host genetics, Direct-acting antivirals, Drug resistance, Vaccine, Hepatitis C virus
Core tip: Twenty-five years after the discovery of hepatitis C virus (HCV) as the major cause of non-A, non-B post-transfusion hepatitis, we have entered a new era in HCV treatment that indicates the prospect of eradication of this important human pathogen. In this article, we will discuss the promising opportunities ahead and key future challenges in the era of new hepatitis C treatments, i.e., barriers in identifying HCV infected individuals, access to new HCV drugs, emergence of drug resistance, and the current status of HCV vaccine development.
INTRODUCTION
Hepatitis C virus (HCV) is a member of the Flaviviridae family, which also includes classical flaviviruses such as those of yellow fever and dengue. HCV is an enveloped virus with a single stranded RNA of positive polarity. The virus has a restricted host range, naturally infecting only humans and chimpanzees, though the origin of HCV still remains elusive. HCV is classified in the genus Hepacivirus of the Flaviviridae family, and the closest genetic relative to HCV is a non-primate hepacivirus, which infects horses[1]. Phylogenetic and sequence analysis of entire viral genomes splits HCV into seven major genotypes. HCV genotypes have been further classified into 67 confirmed and 20 provisional subtypes[2]. The HCV genotype 1 is the most prevalent genotype worldwide (46% of all HCV cases), followed by genotype 3 (30%). Genotypes 2, 4 and 6 are responsible for 23% of all HCV cases and genotype 5 is responsible for less than 1% of all HCV cases. At present, HCV genotype 7 has been isolated only in a patient from Central Africa[3]. Global distribution of HCV genotypes shows geographic variations, which reflect differences in mode of transmission and ethnic variability. In a recently conducted meta-analysis, the number of people with anti-HCV antibodies has been estimated at 185 million in 2005, or 2.8% of the human population, with an estimation of 130-170 million people chronically infected[4]. HCV transmission occurs through blood-to-blood contact. In the early 1990s, introduction of modern anti-HCV screening tests, including the detection of HCV-specific antibodies and HCV RNA[5], almost completely eliminated transmission of HCV through blood transfusions and organ transplants. Injection drug use is currently the primary transmission route for HCV, which usually occurs when blood-contaminated needles and syringes are shared. Unsafe medical procedures, including the reuse of single-use medical devices, remain a major mode of HCV transmission in developing countries[6].
HCV has often been referred to as the “silent virus,” as most HCV infections are clinically silent until the disease reaches a late stage, which often occurs several decades after initial infection. Chronic HCV infection is among the most common causes of cirrhosis and hepatocellular carcinoma, and the most frequent indication for liver transplantation[7]. Recurrence of HCV infection after liver transplantation is universal and a leading cause of graft failure[8]. Efforts to develop direct-acting antivirals (DAAs) for HCV treatment have long been hampered by the absence of an efficient cell culture system for propagation of HCV. Intensive research efforts over the last two decades have resulted in the development of HCV subgenomic replicons, capable of autonomous replication[9], and robust infectious cell culture models for HCV infection[10-12] that not only provide the opportunity to dissect mechanisms of the viral life cycle, but also facilitate the development of large-scale, high-throughput screening assays to identify antiviral targets and to develop highly effective anti-HCV compounds. In this article, we summarize the current state of knowledge and future perspectives for the management of HCV infection.
NATURAL HISTORY OF HCV INFECTION AND ANTIVIRAL IMMUNE RESPONSE
Approximately 25 percent of patients exposed to hepatitis C surmount the infection naturally, but the remaining 75% face persistent or life-long HCV infection. Chronic HCV infection can cause severe liver disease, including cirrhosis, hepatic decompensation and hepatocellular carcinoma (HCC), with an interval of 20-30 years after being exposed to HCV[7]. The World Health Organization’s Global Burden of Disease 2000 project estimated in 2002 that the attributable cirrhosis and liver cancer deaths due to HCV infection globally were 211000 and 155000 respectively[13]. In addition, chronic HCV infection is associated with several extrahepatic manifestations, including mixed cryoglobulinemia vasculitis, type 2 diabetes, lymphoproliferative disorders, renal disease and rheumatic disorders[14]. Considerable research effort has been devoted to understanding the heterogeneous clinical outcome of HCV infection. Comparative immunological studies in HCV-infected patients and experimentally infected chimpanzees demonstrated that clearance of HCV infection is associated with a strong and sustained HCV-specific CD4+ and CD8+ T cell response. Antibody-mediated depletion of either T cell population in chimpanzees provided further evidence that T cell-mediated immunity is crucial for clearance. During the chronic phase of HCV infection, HCV-specific T cells are down-regulated and display an exhausted and dysfunctional phenotype. Chronic liver inflammation, induced by HCV, promotes the generation of T regulatory cells, which contributes to further suppression of the HCV-specific T cell response[15]. HCV infection induces a strong B cell response and antibodies target epitopes within structural and non-structural HCV proteins. Neutralizing antibodies (nAb) arise during HCV infection and the majority of nAb targets epitopes on the envelope glycoproteins[16]. The relevance of neutralizing antibodies in HCV clearance is still unclear. It is important to know that HCV particles interact with serum lipoproteins to form so-called lipo-viro-particles (LVP). Although the overall architectural design of LVP is still unidentified, LVP can facilitate virus entry into hepatocytes and protect the virion from antibody-mediated neutralization[17]. The majority of chronically infected patients have high-titre and cross-reactive neutralizing antibodies[18], suggesting that neutralizing antibodies are unable to clear the infection. Long-term persistence of these antibodies in chronic HCV infection, however, may regulate viral replication and modulate chronic disease. Finally, the host’s immune system is confronted with a highly mutable virus (mutation rate: 10-5-10-4 nucleotides per replication cycle) due to an error-prone viral RNA polymerase that lacks a proofreading activity. Thus, apart from the described genotypes, HCV circulates in infected individuals as a collection of closely related but distinct genomes, called the “quasispecies”[19]. High genetic variability confers an important advantage for HCV, facilitating escape from neutralizing antibodies and cytotoxic T cell recognition.
Human hepatocytes are the primary target cell for HCV infection. A highly sensitive detection system for HCV RNA in the liver demonstrated that the proportion of infected hepatocytes ranges from 1% to 54% and correlated positively with HCV RNA viremia[20]. The first line of immune defense in HCV infection comprises activation of cell-intrinsic innate immunity following HCV recognition. Local production of interferons (IFNs) triggers the expression of hundreds of IFN-stimulated genes disrupting HCV genome replication and spread in the liver parenchyma[21]. Knowledge of viral dynamics and evolution during the early phase of acute HCV infection is still limited because the majority of HCV infections are asymptomatic. The Baltimore Before-and-After Acute Study of Hepatitis enrolled and followed up monthly HCV-negative injection drug users to address these important issues. In this study, high initial HCV RNA viremia level strongly predicted spontaneous clearance of HCV infection[22]. Thus, it is likely that high level HCV replication makes the virus more visible to the innate immune system and, hence, rapidly activates innate immune signaling that result not only in efficient innate anti-viral effector functions but also supports the development of anti-HCV T cell immunity. However, HCV has developed efficient strategies to circumvent innate immune signaling and effector functions. The HCV NS3/NS4A protein - a serine protease responsible for the proteolytic cleavage of the HCV polyprotein precursor - is a key component of the HCV evasion strategy. For example, NS3/NS4A targets and cleaves the mitochondrial antiviral signaling protein, resulting in disruption of innate antiviral signaling and attenuation of IFN production. Furthermore, HCV E2 and NS5A proteins inactivate the double-stranded RNA-dependent protein kinase R, which is a critical mediator of the antiviral effects exerted by IFNs[21].
HOST GENETIC FACTORS IN HCV INFECTION
The genetic background of the host has an important impact on the natural course of HCV infection. CD8+ T cells are the major effector cells that mediate viral clearance. CD8+ T cells recognize viral peptides bound to HLA class I molecules on virus-infected cells. HLA genes display a high degree of genetic variation among individuals, which is reflected in the variations in binding and presentation of viral epitopes. HLA-B27, HLA-B57 and HLA-A3 alleles have been significantly associated with spontaneous clearance of HCV infection. The protective role of these alleles has been linked to viral epitopes, which do not allow immune escape mutations because of profound negative effects on viral replication fitness, resulting in a highly crippled virus[23,24]. Genome-wide association studies that allow the detection of associations between mapped single nucleotide polymorphisms (SNP) and traits have become the standard approach to discovering the genetic basis of human disease. In 2009, a major breakthrough in the understanding of host genomics in HCV infection has been the discovery of several SNPs upstream of the interleukin-28B (IL28B) locus, in particular the SNP rs12979860, which can predict both spontaneous recovery from HCV infection and therapy-induced viral clearance in patients infected with genotype 1[25,26]. The IL28B gene encodes the cytokine IFN-lambda3 (IFN-λ3), which belongs to the type III IFN family (IFN-λ). IFN-λ is rapidly induced during HCV infection and has antiviral activity against HCV[27]. Patients carrying rs12979860 CC genotype had a clearance rate three times higher compared to patients carrying the CT or TT genotype[25]. Interestingly, the frequency of the favorable CC genotype differs markedly across ethnic groups, reaching over 90% in certain North and Eastern Asian populations, an intermediate frequency in Europe, and the lowest frequencies in Africans[25]. Understanding the mechanism of IL28B polymorphism in HCV control is still limited. IL28B polymorphism appears to affect IFN-λ3 expression, with the unfavorable genotypes resulting in reduced IFN-λ3 expression. Patients with the unfavorable genotypes also had a lower induction of innate immunity genes, suggesting that IL28B polymorphism may regulate innate immune functions[21].
ANTIVIRAL TREATMENTS
Until 2011, the standard-of-care (SOC) treatment for chronic hepatitis C was the combination of weekly pegylated interferon-alpha (pegIFNα) and daily doses of ribavirin (RBV) in a 24- or 48-wk course. PegIFNα/RBV dual therapy is associated with several important side effects, including anemia, depression and nausea, which can lead to discontinuation of therapy. Cure of chronic HCV infection is tantamount to the sustained virological response (SVR), which is defined as undetectable HCV RNA in the blood at the end of treatment and again six months later[28]. SVR rates vary according the HCV genotype involved, with SVR rates of 70%-90% for genotypes 2, 3, 5 and 6, but with less than 50% for genotypes 1 and 4[26,29]. In addition to its substantial role as a predictive factor for spontaneous HCV clearance, the IL28B genetic background has been reported as the strongest predictor of response to SOC treatment among patients infected with HCV genotypes 1 and 4[26,30]. Analyses of SOC treatment outcomes in the largest cohort, with more than 1000 patients infected with HCV genotype 1, demonstrated that patients carrying the favorable rs12979860 CC genotype were associated with a more than twofold greater chance of achieving SVR than patients with the unfavorable TT genotype[26]. The discovery of IL28B genetic polymorphism, as a factor predictive of SOC treatment, rapidly stimulated the development of a commercial test to define the IL28B genotype status in HCV genotype 1-infected patients. Although implementation of advanced diagnostic tools - which facilitate personalized medicine approaches - has been long awaited by clinicians, the rapid move in HCV therapy toward DAAs has weakened the relevance of IL28B genotyping in clinical prediction and management of chronic HCV infection.
In 2011, the arrival of first-generation DAAs profoundly changed the landscape of HCV therapy and SVR rates. Though virtually every step of the HCV life cycle - including receptor binding and virus release - can be a target for drug development, DAA targeting of key steps of viral replication and subsequent viral polyprotein processing succeeded in clinical trials. The first available oral DAAs telaprevir (Incivek, Vertex) and boceprevir (Victrelis, Merck) were linear ketoamide inhibitors, which form a reversible but covalent complex with the HCV NS3/4A serine protease catalytic site. Adding one of two NS3/4A inhibitors to dual pegIFN/RBV therapy increased SVR rates up to 75% in treatment-naïve patients and up to 64% for previous non-responders to pegIFN/RBV dual therapy[31-34]. Analysis of SVR rates in the context of IL28B genotype demonstrated that the rs12979860 CC genotype IL28B genotype remained predictive of a favorable response in triple therapy patients[35]. Though the introduction of HCV protease inhibitors was a major milestone in HCV therapy, there are considerable drawbacks of these first-generation protease inhibitors: (1) The unfavorable pharmacokinetic profile of protease inhibitors, which necessitates doses on a thrice-a-day basis; (2) Drug interactions with other medicaments, since HCV protease inhibitors are metabolized by the liver via the cytochrome P450 3A; (3) Protease treatment-related adverse events, including severe skin rashes/pruritus, anemia and dysgeusia; and (4) the treatment option that is limited to HCV genotype 1 infected patients. Drug development has been focused on HCV genotype 1 because of its high prevalence in Europe and the United States, and the low SVR rates in HCV genotype 1 infected patients following pegIFNα/RBV dual therapy[3]. Enormous efforts have been made to overcome these shortcomings, resulting in the development of numerous so-called second-wave protease inhibitors with pan-genotypic effect, improved pharmacokinetic profiles, and tolerability. In 2013, simeprevir (Olysio, Janssen), a once daily administered second-wave protease inhibitor, was approved in combination with pegIFNα/RBV dual therapy. This triple combination increased SVR rates up to 85% in treatment-naïve HCV genotype 1 infected patients, without worsening the known side effects associated with pegIFNα/RBV dual therapy[36,37].
Viral polymerases are prime targets for the development of antiviral drugs since their enzymatic sites are highly conserved between different genotypes. In addition, mutations in the active site of viral polymerases are rarely well tolerated, because they are often associated with reduced viral replication. However, the clinical use of numerous developed nucleoside and nucleotide NSB5 polymerase inhibitors has been halted for toxicity reasons. In 2014, the marketing approval of the first nucleotide NS5B polymerase inhibitor Sofosbuvir (Sovaldi, Gilead Sciences) represented a major milestone in the treatment of chronic hepatitis C. Considered safe and well-tolerated with pan-genotypic activity and a high barrier to resistance, Sofosbuvir - once daily in combination with pegIFNα/RBV dual therapy for 12 wk - improved SVR rates to 82%-100% in treatment naïve patients infected with genotypes 1, 4, 5 or 6[38].
A further step forward toward the next generation of HCV treatment represented the first DAA only regimes (pegIFNα-free, RBV-free). Daclatasvir (Daklinza, Bristol-Myers Squibb) and ledipasvir (Gilead) are inhibitors of the HCV NS5A protein, which play an important role in HCV replication and assembly. Both molecules possess high potency, with a broad coverage of genotypes[39]. The combination of ledipasvir and sofosbuvir in a once-daily, single-tablet regime (Harvoni, Gilead) resulted in high rates of SVR (93%-99%) in treatment naïve HCV genotype 1 patients and previous non-responders to pegIFNα/RBV dual therapy[40-43]. In October 2014, Harvoni was approved for the treatment of patients with chronic HCV genotype 1. The combination sofosbuvir and daclatasvir once daily in a two-tablet regime in patients infected with HCV genotypes 1, 2 or 3 revealed SVR rates ranging from 89% to 100% in previously treated or untreated chronic HCV infection[44] and received also marketing authorization. Impressive SVR rates (> 90%) have been also reported for the dual regime of daclatasvir (Daklinza, Bristol-Myers Squibb) and the second-wave NS3/NS4A protease inhibitor asunaprevir (Sunvepra, Bristol-Myers Squibb)[45]. The first regulatory approval for this combination in patients with HCV genotype 1 infection has been obtained in Japan[46]. Similarily, a combined regimen of simeprevir (Olysio, Janssen) and sofosbuvir (Sovaldi, Gilead Sciences) was efficacious, well tolerated and approved for HCV genotype 1[47]. The so-called 3D combination containing the protease inhibitor ABT-450 with ritonavir (ABT-450/r, AbbVie), the NS5A inhibitor ombitasvir (ABT-267, AbbVie), the nonnucleoside polymerase inhibitor dasabuvir (ABT-333, AbbVie) was associated with cure rates of 99% and is expected to gain approval in 2015[48]. Grazoprevir (MK-5172, Merck) a second-generation protease inhibitor in combination with the NS5A inhibitor elbasvir (MK-8742, Merck) in a single tablet and once-daily regimen[49,50] demonstrated also impressive SVR rates and is expected to file for regulatory approval.
The arrival of potent DAAs has revolutionized chronic hepatitis C treatment, and all-oral pegIFNα-free and RBV-free therapy, achieved by combining two or three DAAs, is no longer science fiction. DAAs promise a highly effective, pan-genotypic, well-tolerated HCV therapy with once-daily single-tablet regimens and shorter courses of treatment (8-12 wk or probably less). The rapid and dramatic reduction in plasma HCV RNA levels observed during DAA treatment (negative HCV-RNA 2-3 wk after starting DAA therapy) will probably also facilitate the management and clinical care of patients with chronic HCV infection. At present, DAA treatments have not been sufficiently studied in genotypes other than genotype 1 and patients who are more difficult to treat, such as patients with advanced fibrosis and cirrhosis or severe liver disease, patients with HIV or HBV co-infection, patients with an indication for liver transplantation and recipients, and patients with renal failure and other co-morbidities. Results from these clinical trials are impatiently awaited to evaluate SVR rates, risk of drug-drug interactions, and rates of side effects in these subsets of patients. Finally, recent advances in high-throughput technologies assessing simultaneously inhibitor potency and specificity may guide to the development of anti-viral drugs with a high safety profile. Analysis of first-generation NS3 protease inhibitors using a high-throughput, super-family wide specificity profiling revealed that telaprevir (Incivek, Vertex) - but not boceprevir (Victrelis, Merck) - potently inhibited two human proteases that are exclusively expressed in the skin[51] suggesting that the serious skin reactions associated with telaprevir (Incivek, Vertex) is mediated by an off-target inhibition of a human protein.
The SVR response is commonly used to describe the successful treatment of HCV infection. SVR is regarded as being equivalent to long-term viral eradication, though there is still an ongoing debate whether non-detectable serum HCV RNA, following spontaneous clearance or secondary to therapy, represents “true” viral eradication[52]. Reports of HCV reappearance have been described for patients who had developed a SVR following pegIFNα/RBV dual therapy[53]. Although the possibility of re-infection cannot be entirely excluded as a cause of HCV recurrence, HCV RNA sequence comparison studies in patients with late relapse demonstrated the presence of the original HCV sequence before treatment and after relapse[54], suggesting a “true” relapse of the original virus rather than re-infection. Interestingly, Veerapu et al[55] reported trace amounts of HCV RNA that reappeared sporadically in the circulation within eight years in some patients who experienced a SVR after pegIFNα/RBV dual therapy. Although reappearance of HCV RNA seems to be a rare event and did not result in high-level viremia, Veerapu et al[55] demonstrated in subsequent studies that these minimal amounts of HCV RNA can cause infection in the chimpanzee model, indicating the presence of replication-competent virus[56]. It is unclear how HCV achieves low-level persistence for several years after successful pegIFNα/RBV dual therapy. In contrast to HBV and human immunodeficiency virus (HIV), HCV does not integrate into the host genome. Whether HCV persists in the liver in a form that is also refractory to eradication by successful DAA treatment has to be evaluated in long-term follow-up studies.
FUTURE CHALLENGES IN HCV INFECTION
Highly effective DAA-based regimes for the treatment for chronic hepatitis C are available. In addition, current drugs in the anti-HCV pipeline promise further DAA with excellent potency, improved tolerance and safety profiles[57]. The burden of HCV-related cirrhosis and HCC is expected to rise over the next two decades[58], suggesting that advances in HCV therapy have arrived at just the right time. Undoubtedly, introduction of DAA-based treatment regimes will have a long-term effect on HCV prevalence and HCV mortality and morbidity. However, the real impact of DAA-based treatment on the rising burden of HCV-related liver disease is currently difficult to estimate and depends on the number of patients who are receiving treatment. To increase this population, a process of identifying HCV-infected patients and effectively linking them to appropriate care and DAA-based treatment will be essential.
HCV screening
In many countries, testing for HCV is recommended for persons who are at high risk, such as injection drug users, persons who received blood transfusions or organ transplants before July 1992, and HIV-infected patients. However, a national health and nutrition examination survey of United States households from 2001 through 2008 revealed that half of the HCV-infected individuals were unaware of their HCV infection status, indicating limited effectiveness of current HCV testing recommendations[59]. Thus, despite highly effective DAA-based treatment regimes, there might be a modest impact on the rising burden of HCV-related liver disease due to the large pool of unidentified HCV-infected individuals. The so-called “hidden HCV population” may include individuals who deny past risk behaviors for HCV infection, individuals who had been exposed to blood products or invasive procedures in countries with high HCV endemicity or poor precautionary measures to prevent infections, former healthcare workers at risk for occupational exposure to blood or body fluids and, finally, recipients of blood products or organs before 1992 who had not yet been tested for HCV infection. To increase the identification of individuals with chronic hepatitis C and link them to appropriate care and treatment, the United States Centers for Disease Control and Prevention now recommend that adults born during 1945 and 1965 should receive a one-time testing for HCV without prior ascertainment of HCV risk (the so-called “birth-cohort screening”) because several studies have shown that this cohort has the highest prevalence of anti-HCV antibodies[60]. Additional innovative HCV screening approaches and health policies to better identify those chronically infected by HCV are urgently needed. HCV screening programs in developed countries should also consider the epidemiological changes around the world caused by immigration from countries with high HCV prevalence, such as those in Africa[61]. HCV testing requires both an antibody test and HCV RNA follow-up testing. While simple and rapid tests for HCV-specific antibodies have been developed[62], HCV RNA testing still requires a specialized laboratory. Complete testing is critical to ensure that those who are chronically infected receive the care and treatment they need. Simplification of HCV diagnosis is warranted to reduce the number of patients lost before the HCV-RNA follow-up testing. The development of highly sensitive and specific tests for detection of the HCV core antigen, in combination with the detection of anti-HCV specific antibodies[63], may help identify patients with current infection more rapidly, and guide them to therapy in particular in countries where state-of-the-art molecular diagnostic methods are not widely available. The majority of HCV-infected individuals live in low-income or resource-limited regions of the world, where unsafe medical procedures and injections remain risks for HCV and where access to HCV testing is still limited. The key challenge for the next decade is to initiate appropriate HCV screening and counseling programs for countries with political and economic instability.
Cost of DAA-based treatments
Everything has a price, and the price of new HCV medicaments is currently too high, though DAAs are cheap to produce. In high income countries, the list price of Harvoni is US $1125 per pill, which corresponds to US $94000 for a 12-wk course of therapy. Although several countries negotiate price discounts, prolongation of treatment to 24 wk, as recommended in patients with cirrhosis, further explodes health care costs[64]. High HCV treatment costs have stimulated an ethical debate on whom to treat and whom not to. The highest priority is given to patients with advanced liver fibrosis and cirrhosis since, for these patients, the clock is ticking. It is expected that HCV drug prices will decrease over time due to approval processes of several other DAAs, competition from other drug manufacturers, and growing political pressure on drug companies. However, to allow widespread access to HCV therapy in low- and middle-income countries, a significant drop in HCV drug prices is necessary. The Egyptian government has negotiated a deal with Gilead to buy Sovaldi at a 99% discount to the United States price, which would imply a cost of about US $900 if Sovaldi is used as part of a 12-wk treatment course. Similar deals for lower prices are expected by other countries with high HCV prevalence, such as India and China. Hill et al[65] and van de Ven et al[66] calculated that within the next 15 years, with a large-scale manufacture of two or three DAAs, the cost for a 12-wk course could be as low as US $100-$200, indicating that HCV drug prices can be dramatically lowered. High drug pricing has been often justified by the need to compensate for intense research and development. However, it is important to note that essential tools for HCV drug development, such as HCV replicons, have been discovered or developed in public research sectors. It is hoped the current costs of hepatitis C treatment would spur new political debates over patents, pricing for DAAs and government-owned industrial corporations, as well as the establishment of national and global HCV programs facilitating access to HCV treatment, in particular for patients who are not covered by health insurance. The key challenge for the next decade is widespread and affordable access to DAA treatment to everyone infected with HCV, irrespective of liver disease status. A broader implementation of DAAs will have a much larger impact on HCV prevalence and HCV-related morbidity and mortality.
Viral resistance
Selective pressure exerted by antiviral drugs can lead to the emergence of drug-resistant viral variants. In fact, resistant variants are selected rapidly during DAA monotherapy with first generation protease inhibitors (PIs)[67]. PIs were, therefore, approved in a pegIFNα/RBV backbone to minimize the development of viral breakthroughs due to resistance mutations. Mutations alone, or in combination at amino acid positions V36, T54, V55, R155, A156, and V/I170, within the NS3/NS4A sequence, have been associated with resistance to first generation PIs. The pattern of resistance mutation depends on the drug and differs according the viral subtype. Nucleotide-heterogeneity leads to a lower genetic-barrier in HCV genotype 1a vs 1b[68]. A retrospective study determined the frequency of PI-resistant variants in patients, who did not achieve an SVR following a telaprevir-containing pegIFNα/RBV regime. Resistant variants were frequently observed after the failure to achieve an SVR (86% for genotype 1a and 56% for genotype 1b). Selected resistant variants were replaced in the absence of an NS3/NS4A inhibitor by the wild type virus within approximately 16 mo in most patients[69], indicating that in contrast to HIV, DAA-resistant variants are not archived. The recently approved second-wave PI simeprevir is a macrocyclic compound that non-covalently binds to the proteolytic activity of NS3. Although first-generation and second-wave PIs belong to different classes, viral variants carrying the R155K mutation confer marked cross-resistance[68].
In HIV infection, antiviral drug resistance testing before antiretroviral therapy initiation has become an essential part of clinical care. Like HIV, resistance-associated variants are naturally produced during the HCV life cycle and their frequency mainly depends on their replication efficacies relative to other pre-existing variants. Bartels et al[70] reported a low prevalence (< 3% of patients) of naturally occurring resistance variants with decreased sensitivity to first generation PIs. However, the presence of a natural polymorphism Q80K is clinically relevant and frequently found in HCV genotype 1a sequences. For this variant, an approximately tenfold reduction in susceptibility to simeprevir has been observed[71]. HCV genotype 1a Q80K polymorphism is heterogeneously distributed around the world, which probably reflects the different geographical distribution of two genotype 1a clades. A high prevalence of Q80K has been observed in NS3 protease sequences from HCV genotype 1a infected patients in North America compared to those in Europe (48.1% vs 19.4%)[72]. Baseline Q80K mutation has a negative impact on treatment outcome. Patients with Q80K mutation showed reduced SVR rates in combination with PegIFN/RBV compared to patients without baseline Q80K[73]. Thus, Q80K genotyping should be performed in HCV genotype 1a infected patients before treatment with simeprevir is initiated. Natural HCV sequence variations are likely to play an important role in the context of future antiviral drug development with pan-genotypic activity. This is evidenced by the fact that natural polymorphisms in genotypes 2b (i.e., S122R) and 3 (i.e., D168Q) render these virus isolates resistant to simeprevir[72].
Pan-genotypic NS5A inhibitors daclatasvir and ledipasvir will play an important role in all-oral DAA combinations, although their specific mechanism of action remains poorly defined. The NS5A protein is organized into three domains and the principal resistance mutations have been mapped on to the first 100 amino acids within the amino-terminal Domain I. The structure of Domain I is dimeric and contains a conserved zinc-binding site required for HCV replication[74]. Patterns of resistance-associated mutations of NS5A inhibitors differ among genotypes. For example, resistance to the NS5A inhibitor daclatasvir is primarily associated with amino acid substitutions at residues M28, Q30, L31, and Y93 for genotype 1a and L31 and Y93 for genotype 1b. The prevalence of natural polymorphism at positions associated with resistance to NS5A inhibitors ranges from 10% to 14%[40,41]. Preexisting NS5A-resistant variants have been associated in some NS5A-based regimes with lower SVR rates. In one study, half of the patients who had had a relapse after treatment with the DAA combination ledipasvir and sofosbuvir (Harvoni) NS5A-resistant variants were already present at baseline[40]. Similarily, the presence of baseline NS5A-resistant variants decreased SVR rates to 76% in a treatment regimen of grazoprevir (MK-5172) and elbasvir (NS5A inhibitor, MK-8742)[49,50].
The pangenotypic inhibitor sofosbuvir is a uridine nucleotide analogue inhibitor of the HCV NS5B polymerase and has a high genetic barrier for resistance. The S282T mutation is the principal mutation that confers decreased susceptibility to sofosbuvir. However, S282T mutation has not been detected in treatment naïve patients and is rarely observed in sofosbuvir-treated patients, since S282T mutation induces a general cost in terms of polymerase efficiency, which may translate into decreased viral fitness[72]. However, low frequency NS5B substitutions at various amino acid positions (i.e., L159F, V321A, C316N) were observed and associated with treatment failure in a subset of patients[75], indicating that further studies are needed to understand the clinical significance of these substitutions.
General drug resistance testing before DAA treatment and following treatment failure is currently not recommended for HCV patients. More complete resistance data and analyses from genotypic and phenotypic resistance assays are needed to determine the clinical impact of potential resistance-associated substitutions and naturally occurring polymorphisms in HCV genotypes that can confer differences in clinical response or complete resistance to DAAs. Based on the results of these studies, a defined drug resistance interpretation system can be developed, and help decide retreatment strategies for those subsets of patients who failed first-line DAA treatments. Finally, medication adherence is expected to be lower in real-word setting which may cause treatment failures due to the emergence or spread of resistant variants.
Vaccine development
Vaccines play a crucial role in controlling infectious diseases and remain the most powerful tool to protect against viral diseases. Eradication of smallpox by worldwide vaccination represents the most crucial achievement[76]. Recent progress in poliovirus eradication further underlines the fundamental role of vaccination in combating viral diseases[77]. Major progress has been made in vaccine development for hepatotropic viruses, allowing the application of efficient vaccines against hepatitis A and B worldwide and, most recently, the first vaccine against hepatitis E virus[78] has been approved in China. However, despite major advances in understanding immunity against HCV, a prophylactic anti-HCV is still missing. Barriers that limit HCV vaccine development are multifaceted and also include limited efforts on part of the pharmaceutical industry. This is illustrated by the fact that only a few promising HCV candidate vaccines entered Phase 1 and Phase 2 clinical trials[79]. One of the major challenges in developing an effective HCV vaccine is the high level of genetic diversity among the different HCV strains and its high mutation rate[2]. The elicitation of a broad and durable neutralizing antibody response, which prevents HCV infection irrespective of the genotype, was first considered the most appealing vaccine strategy; however, due to high variability of HCV envelope glycoproteins, it is a difficult approach. Furthermore, extensive glycosylation of HCV envelope glycoproteins or their interaction with host lipoproteins can attenuate the effect of neutralizing antibodies[80]. Currently, we are beginning to reveal the crystal structures of the HCV envelope glycoprotein E2 alone and in its complex with a neutralizing antibody[81,82]. More of these studies are needed to identify sites in HCV envelope glycoproteins that are targets for neutralizing antibodies. The observation that the humoral immune response alone is insufficient to control HCV infection and that HCV rapidly accumulates mutations in envelope glycoproteins, facilitating escape from neutralizing antibodies[83], has shifted the primary focus in HCV vaccine development to T cell-based vaccines. The objective of an HCV T cell vaccine is to generate a functional and long-lived HCV-specific memory CD4+ and CD8+ T cell response that confers protection from chronic hepatitis C. Studies in humans have shown that HCV-specific memory CD4+ and CD8+ T cells are detectable for up to 20 years after spontaneous viral clearance[84]. To investigate the protective role of HCV-specific T cell memory responses, chimpanzees were re-challenged with HCV, or cohorts of injection drug users at high risk of HCV infection were followed up at close intervals to detect HCV re-infections. These studies demonstrated that resolution of HCV infection does not prevent the risk of HCV re-infection. However, HCV reinfection was characterized by attenuated HCV replication and high rates of spontaneous viral clearance of reinfection. Resolution of HCV reinfection was associated with a rapid recall of HCV-specific T cell immunity, indicating that HCV-specific memory T cells play an important role in protection against secondary HCV infection[85]. Although these findings are considered encouraging, the development of T cell-based HCV vaccines is challenged by the fact that we still do not have defined immunological correlates that predict a protective anti-HCV T cell response.
The most promising HCV vaccine candidates are currently viral vectors, such as adenovirus and vaccinia virus, encoding HCV structural and non-structural proteins[86-90]. Some of these vaccine candidates have been applied in combination with plasmid DNA encoding the same HCV proteins, the so-called “prime-boost strategies”, to enhance the breadth of the elicited CD4+ and CD8+ T cell response. Protective effects of T cell-based vaccines have been tested in chimpanzees, which remain the only reliable model for preclinical studies of HCV vaccines. Folgori et al[86] demonstrated that vaccination with adenoviral vectors and plasmid DNA protected four of five vaccinated chimpanzees from acute hepatitis induced by challenge with a heterologous virus. Though these results are encouraging, it is difficult to draw any definite conclusion regarding the performance of these vaccines in humans, because there are important differences in the outcome of HCV infection in chimpanzees compared with humans. Chimpanzees clear HCV infection more frequently than humans and chronic hepatitis C is less severe in chimpanzees since they do not develop progressive hepatic fibrosis[91]. To advance HCV vaccine development, a Phase I study, assessing the safety and immunogenicity of adenoviral vectors engineered to express viral proteins of HCV genotype 1b (ClinicalTrials.gov NCT01070407), has been tested in healthy volunteers. Vaccination was safe and well-tolerated, and induced long-lived CD4+ and CD8+ T cell response with cross-genotype recognition, indicating the potential of cross-genotypic protection of this vaccine candidate[89]. However, the boosting effect of adenoviral vectors was limited due to the induction of adenovirus-neutralizing antibodies and T cell responses after the first immunization. To circumvent this negative effect, alternative boosting vectors, such as vaccinia virus, are used to maintain a long-term memory response. Recently, Swadling et al[92] tested an HCV T cell vaccination strategy composed on heterologous viral vectors (adenovirus 3 and modified vaccinia virus Ankara) in a phase I human study. This approach generated high levels of both CD8+ and CD4+ HCV-specific T cells targeting multiple HCV antigens irrespective of the host HLA background. Currently, HCV-uninfected active injection drug users are vaccinated with adenoviral vectors and a modified vaccinia virus Ankara in a phase I/II, double-blinded, placebo-controlled study (NTC01436357). Results of this vaccine trial are expected to be available in 2016.
Progress in HCV vaccine development is also hampered by the lack of a small and suitable animal model for the study of protective HCV-specific immunity and efficiency of HCV candidates. Mice and rats are naturally resistant to HCV infection. Engraftment of human hepatocytes into the liver of immunodeficient mice[93] has been proven to be an important model to study the HCV life cycle and the evaluation of anti-HCV drug candidates[94]. However, due to the immunodeficient background needed to prevent transplant rejection, these mice are not suitable to study HCV-specific immunity. A fully immunocompetent mouse model, which is susceptible to HCV infection, is urgently needed to spur testing and prioritizing of HCV vaccine candidates for clinical trials. Current concepts include the development of so-called humanized mouse models, in which human hepatocytes and immune cells are grafted in highly immunodeficient mice. Although various humanized mouse models engrafted with human hematopoietic stem cells have already been developed, dual engraftment of mice remains a difficult challenge[95]. Furthermore, there are a number of limitations in the currently available humanized models. The development and function of certain immune cell types have been shown to be defective or immature. The defects are probably due to the absence of human factors and cytokines required for the differentiation and maturation of immune cells[95,96]. Another approach toward a small animal model for HCV infection and immunity consists in the creation of a transgenic mouse model susceptible to HCV infection. Identification of human factors required for HCV uptake, such as human CD81 and occludin[97], has paved the way for the development of a transgenic mouse model. Expression of human CD81 and occludin in fully immunocompetent inbred mice rendered mice susceptible to HCV infection[98,99]. However, a sustained and prolonged HCV replication was observed only in a profoundly impaired innate immune background[99]. Though in the short term there will be no mouse model that accurately mimics the important hallmarks of HCV infection in humans, these models may give some clues to understanding protective immunity against HCV.
CONCLUSION
Major progress has been made in HCV research and treatment over the last two decades (Figure 1). Although highly effective HCV drugs will be available and affordable for all countries of the world, this will probably not be the deathblow for HCV. Considerable challenges remain for the next few decades (Table 1) and will require reorientation of funding toward HCV testing and vaccine development. New partnerships between governments and industry should be established to better manage regulatory processes and, most importantly, to limit costs of future treatments. However, what happens to the liver after “getting rid” of the virus? Data collection from large clinical trials with pegIFNα/RBV dual therapy demonstrated that successful treatment of HCV infection is associated with reduced incidence of liver disease progression, including liver failure, cirrhosis and HCC[100]. Nevertheless, it is important to note that a virologic cure does not necessarily reflect a cure from risk of liver disease. Persistent hepatic inflammation and/or progression to cirrhosis have been reported in a small subset of patients following viral clearance. There is also evidence that patients with advanced fibrosis or cirrhosis remain at increased risk of HCC even several years after viral clearance[101,102]. HCV-related cirrhosis or HCC is projected to sharply climb in the next decade in most countries. It is hoped that the recent breakthrough in HCV treatment reaches the patient most in need, on time.
Figure 1.
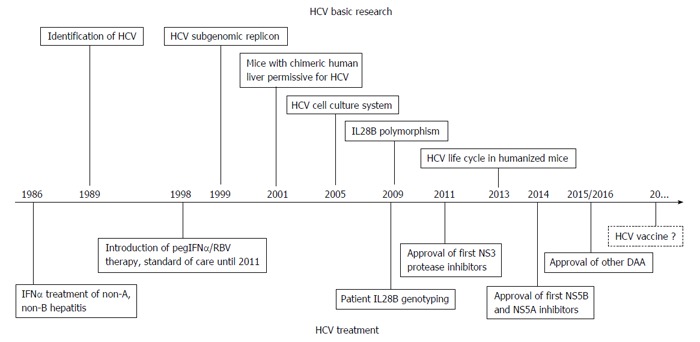
Milestones in hepatitis C virus basic research and treatment. HCV: Hepatitis C virus; DAA: Direct-acting antiviral; IFN: Interferons; IL28B: Interleukin-28B; RBV: Ribavirin; pegIFNα: Pegylated interferon-alpha.
Table 1.
Key elements in future hepatitis C virus management
| Innovative HCV screening programs |
| Education and counseling programs to reduce HCV transmission |
| Development and implementation of a global coordinated HCV action plan |
| Rapid, accurate and cost-effective diagnostic testing methods to detect HCV |
| Affordable prices of DAAs to allow widespread access to HCV treatment |
| HCV drug resistance interpretation tools |
| Detailed understanding of cellular and molecular mechanisms involved in liver regeneration after HCV cure |
| Reinforcement of vaccine research and development |
HCV: Hepatitis C virus; DAAs: Direct-acting antivirals.
Footnotes
P- Reviewer: Gorrell MD, Pan WS S- Editor: Tian YL L- Editor: A E- Editor: Liu SQ
Conflict-of-interest: None.
Open-Access: This article is an open-access article which was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Peer-review started: November 29, 2014
First decision: December 12, 2014
Article in press: January 20, 2015
References
- 1.Pfaender S, Cavalleri JM, Walter S, Doerrbecker J, Campana B, Brown RJ, Burbelo PD, Postel A, Hahn K, Anggakusuma N, et al. Clinical course of infection and viral tissue tropism of hepatitis C virus-like nonprimate hepaciviruses in horses. Hepatology. 2015;61:447–459. doi: 10.1002/hep.27440. [DOI] [PubMed] [Google Scholar]
- 2.Smith DB, Bukh J, Kuiken C, Muerhoff AS, Rice CM, Stapleton JT, Simmonds P. Expanded classification of hepatitis C virus into 7 genotypes and 67 subtypes: updated criteria and genotype assignment web resource. Hepatology. 2014;59:318–327. doi: 10.1002/hep.26744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Messina JP, Humphreys I, Flaxman A, Brown A, Cooke GS, Pybus OG, Barnes E. Global distribution and prevalence of hepatitis C virus genotypes. Hepatology. 2015;61:77–87. doi: 10.1002/hep.27259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Mohd Hanafiah K, Groeger J, Flaxman AD, Wiersma ST. Global epidemiology of hepatitis C virus infection: new estimates of age-specific antibody to HCV seroprevalence. Hepatology. 2013;57:1333–1342. doi: 10.1002/hep.26141. [DOI] [PubMed] [Google Scholar]
- 5.Kuo G, Choo QL, Alter HJ, Gitnick GL, Redeker AG, Purcell RH, Miyamura T, Dienstag JL, Alter MJ, Stevens CE. An assay for circulating antibodies to a major etiologic virus of human non-A, non-B hepatitis. Science. 1989;244:362–364. doi: 10.1126/science.2496467. [DOI] [PubMed] [Google Scholar]
- 6.Alter MJ. HCV routes of transmission: what goes around comes around. Semin Liver Dis. 2011;31:340–346. doi: 10.1055/s-0031-1297923. [DOI] [PubMed] [Google Scholar]
- 7.Hajarizadeh B, Grebely J, Dore GJ. Epidemiology and natural history of HCV infection. Nat Rev Gastroenterol Hepatol. 2013;10:553–562. doi: 10.1038/nrgastro.2013.107. [DOI] [PubMed] [Google Scholar]
- 8.Joshi D, Pinzani M, Carey I, Agarwal K. Recurrent HCV after liver transplantation-mechanisms, assessment and therapy. Nat Rev Gastroenterol Hepatol. 2014;11:710–721. doi: 10.1038/nrgastro.2014.114. [DOI] [PubMed] [Google Scholar]
- 9.Lohmann V, Körner F, Koch J, Herian U, Theilmann L, Bartenschlager R. Replication of subgenomic hepatitis C virus RNAs in a hepatoma cell line. Science. 1999;285:110–113. doi: 10.1126/science.285.5424.110. [DOI] [PubMed] [Google Scholar]
- 10.Wakita T, Pietschmann T, Kato T, Date T, Miyamoto M, Zhao Z, Murthy K, Habermann A, Kräusslich HG, Mizokami M, et al. Production of infectious hepatitis C virus in tissue culture from a cloned viral genome. Nat Med. 2005;11:791–796. doi: 10.1038/nm1268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zhong J, Gastaminza P, Cheng G, Kapadia S, Kato T, Burton DR, Wieland SF, Uprichard SL, Wakita T, Chisari FV. Robust hepatitis C virus infection in vitro. Proc Natl Acad Sci USA. 2005;102:9294–9299. doi: 10.1073/pnas.0503596102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lindenbach BD, Evans MJ, Syder AJ, Wölk B, Tellinghuisen TL, Liu CC, Maruyama T, Hynes RO, Burton DR, McKeating JA, et al. Complete replication of hepatitis C virus in cell culture. Science. 2005;309:623–626. doi: 10.1126/science.1114016. [DOI] [PubMed] [Google Scholar]
- 13.Perz JF, Armstrong GL, Farrington LA, Hutin YJ, Bell BP. The contributions of hepatitis B virus and hepatitis C virus infections to cirrhosis and primary liver cancer worldwide. J Hepatol. 2006;45:529–538. doi: 10.1016/j.jhep.2006.05.013. [DOI] [PubMed] [Google Scholar]
- 14.Jacobson IM, Cacoub P, Dal Maso L, Harrison SA, Younossi ZM. Manifestations of chronic hepatitis C virus infection beyond the liver. Clin Gastroenterol Hepatol. 2010;8:1017–1029. doi: 10.1016/j.cgh.2010.08.026. [DOI] [PubMed] [Google Scholar]
- 15.Rehermann B. Pathogenesis of chronic viral hepatitis: differential roles of T cells and NK cells. Nat Med. 2013;19:859–868. doi: 10.1038/nm.3251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ball JK, Tarr AW, McKeating JA. The past, present and future of neutralizing antibodies for hepatitis C virus. Antiviral Res. 2014;105:100–111. doi: 10.1016/j.antiviral.2014.02.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Lindenbach BD, Rice CM. The ins and outs of hepatitis C virus entry and assembly. Nat Rev Microbiol. 2013;11:688–700. doi: 10.1038/nrmicro3098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bartosch B, Bukh J, Meunier JC, Granier C, Engle RE, Blackwelder WC, Emerson SU, Cosset FL, Purcell RH. In vitro assay for neutralizing antibody to hepatitis C virus: evidence for broadly conserved neutralization epitopes. Proc Natl Acad Sci USA. 2003;100:14199–14204. doi: 10.1073/pnas.2335981100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Farci P. New insights into the HCV quasispecies and compartmentalization. Semin Liver Dis. 2011;31:356–374. doi: 10.1055/s-0031-1297925. [DOI] [PubMed] [Google Scholar]
- 20.Wieland S, Makowska Z, Campana B, Calabrese D, Dill MT, Chung J, Chisari FV, Heim MH. Simultaneous detection of hepatitis C virus and interferon stimulated gene expression in infected human liver. Hepatology. 2014;59:2121–2130. doi: 10.1002/hep.26770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Horner SM, Gale M. Regulation of hepatic innate immunity by hepatitis C virus. Nat Med. 2013;19:879–888. doi: 10.1038/nm.3253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Liu L, Fisher BE, Thomas DL, Cox AL, Ray SC. Spontaneous clearance of primary acute hepatitis C virus infection correlated with high initial viral RNA level and rapid HVR1 evolution. Hepatology. 2012;55:1684–1691. doi: 10.1002/hep.25575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Schmidt J, Thimme R, Neumann-Haefelin C. Host genetics in immune-mediated hepatitis C virus clearance. Biomark Med. 2011;5:155–169. doi: 10.2217/bmm.11.19. [DOI] [PubMed] [Google Scholar]
- 24.Neumann-Haefelin C, Oniangue-Ndza C, Kuntzen T, Schmidt J, Nitschke K, Sidney J, Caillet-Saguy C, Binder M, Kersting N, Kemper MW, et al. Human leukocyte antigen B27 selects for rare escape mutations that significantly impair hepatitis C virus replication and require compensatory mutations. Hepatology. 2011;54:1157–1166. doi: 10.1002/hep.24541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Thomas DL, Thio CL, Martin MP, Qi Y, Ge D, O’Huigin C, Kidd J, Kidd K, Khakoo SI, Alexander G, et al. Genetic variation in IL28B and spontaneous clearance of hepatitis C virus. Nature. 2009;461:798–801. doi: 10.1038/nature08463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ge D, Fellay J, Thompson AJ, Simon JS, Shianna KV, Urban TJ, Heinzen EL, Qiu P, Bertelsen AH, Muir AJ, et al. Genetic variation in IL28B predicts hepatitis C treatment-induced viral clearance. Nature. 2009;461:399–401. doi: 10.1038/nature08309. [DOI] [PubMed] [Google Scholar]
- 27.Kelly C, Klenerman P, Barnes E. Interferon lambdas: the next cytokine storm. Gut. 2011;60:1284–1293. doi: 10.1136/gut.2010.222976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ghany MG, Strader DB, Thomas DL, Seeff LB. Diagnosis, management, and treatment of hepatitis C: an update. Hepatology. 2009;49:1335–1374. doi: 10.1002/hep.22759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Manns MP, Wedemeyer H, Cornberg M. Treating viral hepatitis C: efficacy, side effects, and complications. Gut. 2006;55:1350–1359. doi: 10.1136/gut.2005.076646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Asselah T, De Muynck S, Broët P, Masliah-Planchon J, Blanluet M, Bièche I, Lapalus M, Martinot-Peignoux M, Lada O, Estrabaud E, et al. IL28B polymorphism is associated with treatment response in patients with genotype 4 chronic hepatitis C. J Hepatol. 2012;56:527–532. doi: 10.1016/j.jhep.2011.09.008. [DOI] [PubMed] [Google Scholar]
- 31.Bacon BR, Gordon SC, Lawitz E, Marcellin P, Vierling JM, Zeuzem S, Poordad F, Goodman ZD, Sings HL, Boparai N, et al. Boceprevir for previously treated chronic HCV genotype 1 infection. N Engl J Med. 2011;364:1207–1217. doi: 10.1056/NEJMoa1009482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Poordad F, McCone J, Bacon BR, Bruno S, Manns MP, Sulkowski MS, Jacobson IM, Reddy KR, Goodman ZD, Boparai N, et al. Boceprevir for untreated chronic HCV genotype 1 infection. N Engl J Med. 2011;364:1195–1206. doi: 10.1056/NEJMoa1010494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Jacobson IM, McHutchison JG, Dusheiko G, Di Bisceglie AM, Reddy KR, Bzowej NH, Marcellin P, Muir AJ, Ferenci P, Flisiak R, et al. Telaprevir for previously untreated chronic hepatitis C virus infection. N Engl J Med. 2011;364:2405–2416. doi: 10.1056/NEJMoa1012912. [DOI] [PubMed] [Google Scholar]
- 34.Zeuzem S, Andreone P, Pol S, Lawitz E, Diago M, Roberts S, Focaccia R, Younossi Z, Foster GR, Horban A, et al. Telaprevir for retreatment of HCV infection. N Engl J Med. 2011;364:2417–2428. doi: 10.1056/NEJMoa1013086. [DOI] [PubMed] [Google Scholar]
- 35.Poordad F, Bronowicki JP, Gordon SC, Zeuzem S, Jacobson IM, Sulkowski MS, Poynard T, Morgan TR, Molony C, Pedicone LD, et al. Factors that predict response of patients with hepatitis C virus infection to boceprevir. Gastroenterology. 2012;143:608–618.e1-5. doi: 10.1053/j.gastro.2012.05.011. [DOI] [PubMed] [Google Scholar]
- 36.Jacobson IM, Dore GJ, Foster GR, Fried MW, Radu M, Rafalsky VV, Moroz L, Craxi A, Peeters M, Lenz O, et al. Simeprevir with pegylated interferon alfa 2a plus ribavirin in treatment-naive patients with chronic hepatitis C virus genotype 1 infection (QUEST-1): a phase 3, randomised, double-blind, placebo-controlled trial. Lancet. 2014;384:403–413. doi: 10.1016/S0140-6736(14)60494-3. [DOI] [PubMed] [Google Scholar]
- 37.Manns M, Marcellin P, Poordad F, de Araujo ES, Buti M, Horsmans Y, Janczewska E, Villamil F, Scott J, Peeters M, et al. Simeprevir with pegylated interferon alfa 2a or 2b plus ribavirin in treatment-naive patients with chronic hepatitis C virus genotype 1 infection (QUEST-2): a randomised, double-blind, placebo-controlled phase 3 trial. Lancet. 2014;384:414–426. doi: 10.1016/S0140-6736(14)60538-9. [DOI] [PubMed] [Google Scholar]
- 38.Lawitz E, Mangia A, Wyles D, Rodriguez-Torres M, Hassanein T, Gordon SC, Schultz M, Davis MN, Kayali Z, Reddy KR, et al. Sofosbuvir for previously untreated chronic hepatitis C infection. N Engl J Med. 2013;368:1878–1887. doi: 10.1056/NEJMoa1214853. [DOI] [PubMed] [Google Scholar]
- 39.Pawlotsky JM. NS5A inhibitors in the treatment of hepatitis C. J Hepatol. 2013;59:375–382. doi: 10.1016/j.jhep.2013.03.030. [DOI] [PubMed] [Google Scholar]
- 40.Afdhal N, Reddy KR, Nelson DR, Lawitz E, Gordon SC, Schiff E, Nahass R, Ghalib R, Gitlin N, Herring R, et al. Ledipasvir and sofosbuvir for previously treated HCV genotype 1 infection. N Engl J Med. 2014;370:1483–1493. doi: 10.1056/NEJMoa1316366. [DOI] [PubMed] [Google Scholar]
- 41.Afdhal N, Zeuzem S, Kwo P, Chojkier M, Gitlin N, Puoti M, Romero-Gomez M, Zarski JP, Agarwal K, Buggisch P, et al. Ledipasvir and sofosbuvir for untreated HCV genotype 1 infection. N Engl J Med. 2014;370:1889–1898. doi: 10.1056/NEJMoa1402454. [DOI] [PubMed] [Google Scholar]
- 42.Kowdley KV, Gordon SC, Reddy KR, Rossaro L, Bernstein DE, Lawitz E, Shiffman ML, Schiff E, Ghalib R, Ryan M, et al. Ledipasvir and sofosbuvir for 8 or 12 weeks for chronic HCV without cirrhosis. N Engl J Med. 2014;370:1879–1888. doi: 10.1056/NEJMoa1402355. [DOI] [PubMed] [Google Scholar]
- 43.Lawitz E, Poordad FF, Pang PS, Hyland RH, Ding X, Mo H, Symonds WT, McHutchison JG, Membreno FE. Sofosbuvir and ledipasvir fixed-dose combination with and without ribavirin in treatment-naive and previously treated patients with genotype 1 hepatitis C virus infection (LONESTAR): an open-label, randomised, phase 2 trial. Lancet. 2014;383:515–523. doi: 10.1016/S0140-6736(13)62121-2. [DOI] [PubMed] [Google Scholar]
- 44.Sulkowski MS, Gardiner DF, Rodriguez-Torres M, Reddy KR, Hassanein T, Jacobson I, Lawitz E, Lok AS, Hinestrosa F, Thuluvath PJ, et al. Daclatasvir plus sofosbuvir for previously treated or untreated chronic HCV infection. N Engl J Med. 2014;370:211–221. doi: 10.1056/NEJMoa1306218. [DOI] [PubMed] [Google Scholar]
- 45.Manns M, Pol S, Jacobson IM, Marcellin P, Gordon SC, Peng CY, Chang TT, Everson GT, Heo J, Gerken G, et al. All-oral daclatasvir plus asunaprevir for hepatitis C virus genotype 1b: a multinational, phase 3, multicohort study. Lancet. 2014;384:1597–1605. doi: 10.1016/S0140-6736(14)61059-X. [DOI] [PubMed] [Google Scholar]
- 46.Poole RM. Daclatasvir + asunaprevir: first global approval. Drugs. 2014;74:1559–1571. doi: 10.1007/s40265-014-0279-4. [DOI] [PubMed] [Google Scholar]
- 47.Lawitz E, Sulkowski MS, Ghalib R, Rodriguez-Torres M, Younossi ZM, Corregidor A, DeJesus E, Pearlman B, Rabinovitz M, Gitlin N, et al. Simeprevir plus sofosbuvir, with or without ribavirin, to treat chronic infection with hepatitis C virus genotype 1 in non-responders to pegylated interferon and ribavirin and treatment-naive patients: the COSMOS randomised study. Lancet. 2014;384:1756–1765. doi: 10.1016/S0140-6736(14)61036-9. [DOI] [PubMed] [Google Scholar]
- 48.Ferenci P, Bernstein D, Lalezari J, Cohen D, Luo Y, Cooper C, Tam E, Marinho RT, Tsai N, Nyberg A, et al. ABT-450/r-ombitasvir and dasabuvir with or without ribavirin for HCV. N Engl J Med. 2014;370:1983–1992. doi: 10.1056/NEJMoa1402338. [DOI] [PubMed] [Google Scholar]
- 49.Lawitz E, Gane E, Pearlman B, Tam E, Ghesquiere W, Guyader D, Alric L, Bronowicki JP, Lester L, Sievert W, et al. Efficacy and safety of 12 weeks versus 18 weeks of treatment with grazoprevir (MK-5172) and elbasvir (MK-8742) with or without ribavirin for hepatitis C virus genotype 1 infection in previously untreated patients with cirrhosis and patients with previous null response with or without cirrhosis (C-WORTHY): a randomised, open-label phase 2 trial. Lancet. 2015;385:1075–1086. doi: 10.1016/S0140-6736(14)61795-5. [DOI] [PubMed] [Google Scholar]
- 50.Sulkowski M, Hezode C, Gerstoft J, Vierling JM, Mallolas J, Pol S, Kugelmas M, Murillo A, Weis N, Nahass R, et al. Efficacy and safety of 8 weeks versus 12 weeks of treatment with grazoprevir (MK-5172) and elbasvir (MK-8742) with or without ribavirin in patients with hepatitis C virus genotype 1 mono-infection and HIV/hepatitis C virus co-infection (C-WORTHY): a randomised, open-label phase 2 trial. Lancet. 2015;385:1087–1097. doi: 10.1016/S0140-6736(14)61793-1. [DOI] [PubMed] [Google Scholar]
- 51.Bachovchin DA, Koblan LW, Wu W, Liu Y, Li Y, Zhao P, Woznica I, Shu Y, Lai JH, Poplawski SE, et al. A high-throughput, multiplexed assay for superfamily-wide profiling of enzyme activity. Nat Chem Biol. 2014;10:656–663. doi: 10.1038/nchembio.1578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Seeff LB. Sustained virologic response: is this equivalent to cure of chronic hepatitis C? Hepatology. 2013;57:438–440. doi: 10.1002/hep.25964. [DOI] [PubMed] [Google Scholar]
- 53.Radkowski M, Gallegos-Orozco JF, Jablonska J, Colby TV, Walewska-Zielecka B, Kubicka J, Wilkinson J, Adair D, Rakela J, Laskus T. Persistence of hepatitis C virus in patients successfully treated for chronic hepatitis C. Hepatology. 2005;41:106–114. doi: 10.1002/hep.20518. [DOI] [PubMed] [Google Scholar]
- 54.Hara K, Rivera MM, Koh C, Demino M, Page S, Nagabhyru PR, Rehermann B, Liang TJ, Hoofnagle JH, Heller T. Sequence analysis of hepatitis C virus from patients with relapse after a sustained virological response: relapse or reinfection? J Infect Dis. 2014;209:38–45. doi: 10.1093/infdis/jit541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Veerapu NS, Raghuraman S, Liang TJ, Heller T, Rehermann B. Sporadic reappearance of minute amounts of hepatitis C virus RNA after successful therapy stimulates cellular immune responses. Gastroenterology. 2011;140:676–685.e1. doi: 10.1053/j.gastro.2010.10.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Veerapu NS, Park SH, Tully DC, Allen TM, Rehermann B. Trace amounts of sporadically reappearing HCV RNA can cause infection. J Clin Invest. 2014;124:3469–3478. doi: 10.1172/JCI73104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Pawlotsky JM. New hepatitis C therapies: the toolbox, strategies, and challenges. Gastroenterology. 2014;146:1176–1192. doi: 10.1053/j.gastro.2014.03.003. [DOI] [PubMed] [Google Scholar]
- 58.Davis GL, Alter MJ, El-Serag H, Poynard T, Jennings LW. Aging of hepatitis C virus (HCV)-infected persons in the United States: a multiple cohort model of HCV prevalence and disease progression. Gastroenterology. 2010;138:513–521, 521.e1-6. doi: 10.1053/j.gastro.2009.09.067. [DOI] [PubMed] [Google Scholar]
- 59.Denniston MM, Klevens RM, McQuillan GM, Jiles RB. Awareness of infection, knowledge of hepatitis C, and medical follow-up among individuals testing positive for hepatitis C: National Health and Nutrition Examination Survey 2001-2008. Hepatology. 2012;55:1652–1661. doi: 10.1002/hep.25556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Smith BD, Morgan RL, Beckett GA, Falck-Ytter Y, Holtzman D, Ward JW. Hepatitis C virus testing of persons born during 1945-1965: recommendations from the Centers for Disease Control and Prevention. Ann Intern Med. 2012;157:817–822. doi: 10.7326/0003-4819-157-9-201211060-00529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Asselah T, Perumalswami PV, Dieterich D. Is screening baby boomers for HCV enough? A call to screen for hepatitis C virus in persons from countries of high endemicity. Liver Int. 2014;34:1447–1451. doi: 10.1111/liv.12599. [DOI] [PubMed] [Google Scholar]
- 62.Shivkumar S, Peeling R, Jafari Y, Joseph L, Pant Pai N. Accuracy of rapid and point-of-care screening tests for hepatitis C: a systematic review and meta-analysis. Ann Intern Med. 2012;157:558–566. doi: 10.7326/0003-4819-157-8-201210160-00006. [DOI] [PubMed] [Google Scholar]
- 63.Brandão CP, Marques BL, Marques VA, Villela-Nogueira CA, Do Ó KM, de Paula MT, Lewis-Ximenez LL, Lampe E, Sá Ferreira JA, Villar LM. Simultaneous detection of hepatitis C virus antigen and antibodies in dried blood spots. J Clin Virol. 2013;57:98–102. doi: 10.1016/j.jcv.2013.02.014. [DOI] [PubMed] [Google Scholar]
- 64.European Association for Study of Liver. EASL Clinical Practice Guidelines: management of hepatitis C virus infection. J Hepatol. 2014;60:392–420. doi: 10.1016/j.jhep.2013.11.003. [DOI] [PubMed] [Google Scholar]
- 65.Hill A, Khoo S, Fortunak J, Simmons B, Ford N. Minimum costs for producing hepatitis C direct-acting antivirals for use in large-scale treatment access programs in developing countries. Clin Infect Dis. 2014;58:928–936. doi: 10.1093/cid/ciu012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.van de Ven N, Fortunak J, Simmons B, Ford N, Cooke GS, Khoo S, Hill A. Minimum target prices for production of direct-acting antivirals and associated diagnostics to combat hepatitis C virus. Hepatology. 2015;61:1174–1182. doi: 10.1002/hep.27641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Sarrazin C, Kieffer TL, Bartels D, Hanzelka B, Müh U, Welker M, Wincheringer D, Zhou Y, Chu HM, Lin C, et al. Dynamic hepatitis C virus genotypic and phenotypic changes in patients treated with the protease inhibitor telaprevir. Gastroenterology. 2007;132:1767–1777. doi: 10.1053/j.gastro.2007.02.037. [DOI] [PubMed] [Google Scholar]
- 68.Kieffer TL, George S. Resistance to hepatitis C virus protease inhibitors. Curr Opin Virol. 2014;8:16–21. doi: 10.1016/j.coviro.2014.04.008. [DOI] [PubMed] [Google Scholar]
- 69.Sullivan JC, De Meyer S, Bartels DJ, Dierynck I, Zhang EZ, Spanks J, Tigges AM, Ghys A, Dorrian J, Adda N, et al. Evolution of treatment-emergent resistant variants in telaprevir phase 3 clinical trials. Clin Infect Dis. 2013;57:221–229. doi: 10.1093/cid/cit226. [DOI] [PubMed] [Google Scholar]
- 70.Bartels DJ, Zhou Y, Zhang EZ, Marcial M, Byrn RA, Pfeiffer T, Tigges AM, Adiwijaya BS, Lin C, Kwong AD, et al. Natural prevalence of hepatitis C virus variants with decreased sensitivity to NS3.4A protease inhibitors in treatment-naive subjects. J Infect Dis. 2008;198:800–807. doi: 10.1086/591141. [DOI] [PubMed] [Google Scholar]
- 71.Lenz O, Verbinnen T, Fevery B, Tambuyzer L, Vijgen L, Peeters M, Buelens A, Ceulemans H, Beumont M, Picchio G, et al. Virology analyses of HCV isolates from genotype 1-infected patients treated with simeprevir plus peginterferon/ribavirin in Phase IIb/III studies. J Hepatol. 2014:Epub ahead of print. doi: 10.1016/j.jhep.2014.11.032. [DOI] [PubMed] [Google Scholar]
- 72.Poveda E, Wyles DL, Mena A, Pedreira JD, Castro-Iglesias A, Cachay E. Update on hepatitis C virus resistance to direct-acting antiviral agents. Antiviral Res. 2014;108:181–191. doi: 10.1016/j.antiviral.2014.05.015. [DOI] [PubMed] [Google Scholar]
- 73.Forns X, Lawitz E, Zeuzem S, Gane E, Bronowicki JP, Andreone P, Horban A, Brown A, Peeters M, Lenz O, et al. Simeprevir with peginterferon and ribavirin leads to high rates of SVR in patients with HCV genotype 1 who relapsed after previous therapy: a phase 3 trial. Gastroenterology. 2014;146:1669–1679.e3. doi: 10.1053/j.gastro.2014.02.051. [DOI] [PubMed] [Google Scholar]
- 74.Tellinghuisen TL, Marcotrigiano J, Rice CM. Structure of the zinc-binding domain of an essential component of the hepatitis C virus replicase. Nature. 2005;435:374–379. doi: 10.1038/nature03580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Donaldson EF, Harrington PR, O’Rear JJ, Naeger LK. Clinical evidence and bioinformatics characterization of potential hepatitis C virus resistance pathways for sofosbuvir. Hepatology. 2015;61:56–65. doi: 10.1002/hep.27375. [DOI] [PubMed] [Google Scholar]
- 76.Tomori O. From smallpox eradication to the future of global health: innovations, application and lessons for future eradication and control initiatives. Vaccine. 2011;29 Suppl 4:D145–D148. doi: 10.1016/j.vaccine.2011.09.003. [DOI] [PubMed] [Google Scholar]
- 77.Estívariz CF, Pallansch MA, Anand A, Wassilak SG, Sutter RW, Wenger JD, Orenstein WA. Poliovirus vaccination options for achieving eradication and securing the endgame. Curr Opin Virol. 2013;3:309–315. doi: 10.1016/j.coviro.2013.05.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Zhu FC, Zhang J, Zhang XF, Zhou C, Wang ZZ, Huang SJ, Wang H, Yang CL, Jiang HM, Cai JP, et al. Efficacy and safety of a recombinant hepatitis E vaccine in healthy adults: a large-scale, randomised, double-blind placebo-controlled, phase 3 trial. Lancet. 2010;376:895–902. doi: 10.1016/S0140-6736(10)61030-6. [DOI] [PubMed] [Google Scholar]
- 79.Liang TJ. Current progress in development of hepatitis C virus vaccines. Nat Med. 2013;19:869–878. doi: 10.1038/nm.3183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Anjum S, Wahid A, Afzal MS, Albecka A, Alsaleh K, Ahmad T, Baumert TF, Wychowski C, Qadri I, Penin F, et al. Additional glycosylation within a specific hypervariable region of subtype 3a of hepatitis C virus protects against virus neutralization. J Infect Dis. 2013;208:1888–1897. doi: 10.1093/infdis/jit376. [DOI] [PubMed] [Google Scholar]
- 81.Khan AG, Whidby J, Miller MT, Scarborough H, Zatorski AV, Cygan A, Price AA, Yost SA, Bohannon CD, Jacob J, et al. Structure of the core ectodomain of the hepatitis C virus envelope glycoprotein 2. Nature. 2014;509:381–384. doi: 10.1038/nature13117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Kong L, Giang E, Nieusma T, Kadam RU, Cogburn KE, Hua Y, Dai X, Stanfield RL, Burton DR, Ward AB, et al. Hepatitis C virus E2 envelope glycoprotein core structure. Science. 2013;342:1090–1094. doi: 10.1126/science.1243876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.von Hahn T, Yoon JC, Alter H, Rice CM, Rehermann B, Balfe P, McKeating JA. Hepatitis C virus continuously escapes from neutralizing antibody and T-cell responses during chronic infection in vivo. Gastroenterology. 2007;132:667–678. doi: 10.1053/j.gastro.2006.12.008. [DOI] [PubMed] [Google Scholar]
- 84.Takaki A, Wiese M, Maertens G, Depla E, Seifert U, Liebetrau A, Miller JL, Manns MP, Rehermann B. Cellular immune responses persist and humoral responses decrease two decades after recovery from a single-source outbreak of hepatitis C. Nat Med. 2000;6:578–582. doi: 10.1038/75063. [DOI] [PubMed] [Google Scholar]
- 85.Abdel-Hakeem MS, Shoukry NH. Protective immunity against hepatitis C: many shades of gray. Front Immunol. 2014;5:274. doi: 10.3389/fimmu.2014.00274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Folgori A, Capone S, Ruggeri L, Meola A, Sporeno E, Ercole BB, Pezzanera M, Tafi R, Arcuri M, Fattori E, et al. A T-cell HCV vaccine eliciting effective immunity against heterologous virus challenge in chimpanzees. Nat Med. 2006;12:190–197. doi: 10.1038/nm1353. [DOI] [PubMed] [Google Scholar]
- 87.Habersetzer F, Honnet G, Bain C, Maynard-Muet M, Leroy V, Zarski JP, Feray C, Baumert TF, Bronowicki JP, Doffoël M, et al. A poxvirus vaccine is safe, induces T-cell responses, and decreases viral load in patients with chronic hepatitis C. Gastroenterology. 2011;141:890–899.e1-4. doi: 10.1053/j.gastro.2011.06.009. [DOI] [PubMed] [Google Scholar]
- 88.Park SH, Shin EC, Capone S, Caggiari L, De Re V, Nicosia A, Folgori A, Rehermann B. Successful vaccination induces multifunctional memory T-cell precursors associated with early control of hepatitis C virus. Gastroenterology. 2012;143:1048–1060.e4. doi: 10.1053/j.gastro.2012.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Barnes E, Folgori A, Capone S, Swadling L, Aston S, Kurioka A, Meyer J, Huddart R, Smith K, Townsend R, et al. Novel adenovirus-based vaccines induce broad and sustained T cell responses to HCV in man. Sci Transl Med. 2012;4:115ra1. doi: 10.1126/scitranslmed.3003155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Zubkova I, Duan H, Wells F, Mostowski H, Chang E, Pirollo K, Krawczynski K, Lanford R, Major M. Hepatitis C virus clearance correlates with HLA-DR expression on proliferating CD8+ T cells in immune-primed chimpanzees. Hepatology. 2014;59:803–813. doi: 10.1002/hep.26747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Jo J, Lohmann V, Bartenschlager R, Thimme R. Experimental models to study the immunobiology of hepatitis C virus. J Gen Virol. 2011;92:477–493. doi: 10.1099/vir.0.027987-0. [DOI] [PubMed] [Google Scholar]
- 92.Swadling L, Capone S, Antrobus RD, Brown A, Richardson R, Newell EW, Halliday J, Kelly C, Bowen D, Fergusson J, et al. A human vaccine strategy based on chimpanzee adenoviral and MVA vectors that primes, boosts, and sustains functional HCV-specific T cell memory. Sci Transl Med. 2014;6:261ra153. doi: 10.1126/scitranslmed.3009185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Meuleman P, Libbrecht L, De Vos R, de Hemptinne B, Gevaert K, Vandekerckhove J, Roskams T, Leroux-Roels G. Morphological and biochemical characterization of a human liver in a uPA-SCID mouse chimera. Hepatology. 2005;41:847–856. doi: 10.1002/hep.20657. [DOI] [PubMed] [Google Scholar]
- 94.Meuleman P, Leroux-Roels G. The human liver-uPA-SCID mouse: a model for the evaluation of antiviral compounds against HBV and HCV. Antiviral Res. 2008;80:231–238. doi: 10.1016/j.antiviral.2008.07.006. [DOI] [PubMed] [Google Scholar]
- 95.Ito R, Takahashi T, Katano I, Ito M. Current advances in humanized mouse models. Cell Mol Immunol. 2012;9:208–214. doi: 10.1038/cmi.2012.2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Rongvaux A, Willinger T, Martinek J, Strowig T, Gearty SV, Teichmann LL, Saito Y, Marches F, Halene S, Palucka AK, et al. Development and function of human innate immune cells in a humanized mouse model. Nat Biotechnol. 2014;32:364–372. doi: 10.1038/nbt.2858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Scheel TK, Rice CM. Understanding the hepatitis C virus life cycle paves the way for highly effective therapies. Nat Med. 2013;19:837–849. doi: 10.1038/nm.3248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Dorner M, Horwitz JA, Robbins JB, Barry WT, Feng Q, Mu K, Jones CT, Schoggins JW, Catanese MT, Burton DR, et al. A genetically humanized mouse model for hepatitis C virus infection. Nature. 2011;474:208–211. doi: 10.1038/nature10168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Dorner M, Horwitz JA, Donovan BM, Labitt RN, Budell WC, Friling T, Vogt A, Catanese MT, Satoh T, Kawai T, et al. Completion of the entire hepatitis C virus life cycle in genetically humanized mice. Nature. 2013;501:237–241. doi: 10.1038/nature12427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Lee YA, Friedman SL. Reversal, maintenance or progression: what happens to the liver after a virologic cure of hepatitis C? Antiviral Res. 2014;107:23–30. doi: 10.1016/j.antiviral.2014.03.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Morgan RL, Baack B, Smith BD, Yartel A, Pitasi M, Falck-Ytter Y. Eradication of hepatitis C virus infection and the development of hepatocellular carcinoma: a meta-analysis of observational studies. Ann Intern Med. 2013;158:329–337. doi: 10.7326/0003-4819-158-5-201303050-00005. [DOI] [PubMed] [Google Scholar]
- 102.Yamashita N, Ohho A, Yamasaki A, Kurokawa M, Kotoh K, Kajiwara E. Hepatocarcinogenesis in chronic hepatitis C patients achieving a sustained virological response to interferon: significance of lifelong periodic cancer screening for improving outcomes. J Gastroenterol. 2014;49:1504–1513. doi: 10.1007/s00535-013-0921-z. [DOI] [PubMed] [Google Scholar]