

Abstract
Hepatitis C virus (HCV) induces persistent infection and causes chronic liver disease in most infected patients. Vigorous HCV-specific CD4+ and CD8+ T cell responses against HCV multiple epitopes are necessary for spontaneous viral clearance during the acute phase, but the virus appears to have multiple strategies to evade these defenses. There are relatively few studies on the role of immune responses during the chronic phase of infection. CD4+ T cell responses appear to protect against liver injury and may be important to clearance during interferon and ribavirin based therapy. Classic cytotoxic T cells (CTL) may primarily damage the liver in chronic HCV, but there may be subpopulations of T cells that protect against liver inflammation. Resolution of these outstanding questions is important to the development of a prophylactic vaccine as well as improving therapeutic options for those with chronic infection.
Keywords: Hepatitis C virus, CD4 T cells, CD8+ T cells, chronic infection, liver injury, persistence, immune escape, immunopathogenesis
Introduction
Hepatitis C virus (HCV) is a leading cause of chronic liver disease worldwide [1], with more than 170 million individuals with chronic infection [2]. Since the advent of highly effective screening of blood and blood products to eliminate HCV, transmission is primarily through intravenous drug use; although cases do continue to occur through iatrogenic, occupational, vertical and sexual routes of exposure. Acute HCV infection is typically subclinical, although if detected it is highly amenable to therapy (approximately 80 % chance of sustained virologic response (SVR), or cure). Spontaneous resolution of HCV infection occurs in approximately 20% of infected individuals, although this rate may vary widely depending on host factors such as age and gender, with young females having a higher likelihood of spontaneous resolution than other groups [3]. It is estimated that on average 20 % of those with chronic HCV will develop liver cirrhosis over a 20-year period after infection, although the range is higly variable; and of those who develop cirrhosis between 0 and 3 % will develop hepatocellular carcinoma per year [4]. The rate of disease progression to cirrhosis is highly variable and the host and viral factors that modify the course are poorly understood. Alcohol ingestion is a major cofactor [5], but how other factors, such as male gender and age at acquisition, impact liver injury are at present not understood. Although current therapies can cure HCV infection in approximately one half of infected persons [6], therapy is difficult and not tolerated by many patients.
Characteristics of HCV
HCV belongs to the hepacivirus in the Flaviviridae family [7]. Other members of this family, such as yellow fever virus, cause acute hepatitis but not chronic infection; since HCV appears to infect only humans and chimpanzees, the lack of a closely related virus that can be studied in a small animal model has hindered studies of pathogenesis. HCV has 6 major genotypes and more than 50 subtypes [8]. HCV is a positive-sense single-stranded RNA virus with a genome of 9600 nucleotides encoding a single open reading frame (ORF) encoding a polyprotein of approximately 3,000 amino acids that is processed during and after translation into at least 10 proteins [9]; three structural proteins and seven non-structural proteins (Figure 1). For full details of the current concepts regarding replication and function of the viral proteins, the reader is referred to several recent reviews [7; 9; 10; 11], although the functions are summarized in Table 1. In brief, the viral genome encodes for structural proteins (core, which forms the viral nucleocapsid, and the envelope glycoproteins E1 and E2, which are glycosylated membrane proteins) as well as the non-structural (NS) proteins, which have essential functions in viral replication. NS3 contains protease, RNA helicase and nucleoside triphosphatase (NTPase) activities, all of which are essential to viral replication. NS5B is an RNA-dependent RNA polymerase (RdRp) of HCV, and a key enzyme for viral transcription and replication [7; 10; 11; 12; 13]. Inhibition of NS3 and NS5b function are key targets for rationale drug design of new antiviral agents. However, all proteins can be targets of the immune response and, as we will discuss, several viral proteins appear to play some role in evading host immune defenses.
Figure 1.

The HCV genome and gene products. HCV is positive-sense single-stranded RNA virus. This viral genome contains a single open reading frame (ORF) encoding a polyprotein, processed into at least 10 proteins; three structural proteins and seven non-structural proteins.
Table 1.
Functions of HCV structural and non-structural proteins (NS)
| HCV protein | Function |
|---|---|
| Core | Nucleocapcid |
| E1 and E2 | Envelpe glycosylate membrane proteins
Entry into cell; binding to CD81 Contain HVR** |
| p7 | Viroporin
Ion channel |
| NS2 | NS2/3 protease |
| NS3 | Serine protease
RNA helicase NTPase** |
| NS4A | Cofactor for NS3 protease activity |
| NS4B | Hydrophoblic protein
Integral membrane protein Formation of the HCV RNA replication complex * |
| NS5A | Polyphosphorylated protein
Formation of the HCV RNA replication complex* Responsible for IFN sensitivity of HCV* |
| NS5B | RdRp**
Key enzyme for viral transcription and replication |
A proposed function
HVR; hyper variable region, NTPase; Nucleoside triphosphatase, RdRp; RNA-dependent RNA polymerase
The major site of HCV replication appears to be the hepatocyte, although it is not known how many hepatocytes within the infected liver support productive replication. HCV has also been reported to infect numerous other cell types, including B cells, dendritic cells (DC), and other peripheral blood mononuclear cells [14; 15; 16; 17; 18; 19]. However, true replication within these cells, as opposed to passive adsorption of virus, is not universally accepted; and whether or not this affects function of these cells is similarly not clear. HCV may bind to B cells and drive clonal proliferation, which may account for cryoglobuminemia commonly seen in HCV [20; 21; 22], as well as possibly being related to lymphoproliferative disorders [23].
An HCV replication system has only recently been established that can produce virus capable of both in vitro and in vivo infection [24]. Unfortunately at present this system is limited to a subclone of the hepatocellular carcinoma line Huh-7, and does not produce high titers of virus. This has limited studies of the interaction between host immune cells and the target of infection. There is no small animal model of HCV infection, although immunodeficient mice manipulated to have destruction of the native liver can be transplanted with human hepatocytes, which can then be infected with HCV [25]. The utility of this model is limited by the high mortality of the mouse strain, lack of an adaptive immune system and, more significantly, the fact that in order to understand interactions of the immune system and HCV these mice would have to be transplanted with immune cells from the same donor as the hepatocytes. Chimpanzees are the only species other than man that can be infected with HCV, and thus have been invaluable in studies of the early events following infection. However, very few naïve animals have been available for study in the past, and due to declining availability of these animals, it is possible that this model may no longer be available for research.
Immune responses in acute HCV infection
Following infection, HCV RNA appears in the peripheral blood within 1–2 weeks after primary infection, and triggers multiple arms of the immune response, including both innate and adaptive immunity [26; 27; 28].
The interferon system
HCV RNA triggers production of type I interferon (IFN)α/β in infected cells (reviewed in [11; 29]). A key role in the IFNα/β system is played by IFN regulatory factor (IRF)-3, which activates the IFNα/β promoter; both IFNα and IFNβ are recognized by the IFN receptor, thus providing a positive feedback loop in which there is activation of multiple IFN-responsive genes. Gene expression analysis in acutely infected chimpanzees revealed that HCV triggers expression of type I IFN and IFN-induced genes during the early phase of infection in the liver [30]. Another study in chimpanzees showed upregulation of type I IFN induced double-stranded RNA-dependent protein kinase (PKR), 2′–5′ oligoadenylate synthetase (OAS) and Mx genes upon acute HCV infection, which collectively serve to inhibit replication of viruses and induce apoptosis in infected hepatocytes [31]. IFN may also upregulate the expression of HCV antigens on the surface of infected hepatocytes by modulating the generation of immunoproteosomes, which are essential to presentation of antigens to the immune system [32]. HCV appears to use several mechanisms to subvert the endogenous IFN response, as will be discussed later.
Natural killer (NK) and NKT cell
NK cells and NKT cells play an important role in the first lines of defense against viral infection. These cells may comprise up to 20 to 30 % of intrahepatic lymphocytes in the normal liver and can contribute to the control of viral replication through cytolysis of infected cells, production of cytokines that might inhibit viral replication, such as IFNγ, and activation of both DC and T cells [33; 34; 35]. It has been difficult to measure NK function in acutely infected persons given the subclinical nature of most acute HCV infection. Most studies have focused on measurement of NK cell function in chronic HCV infection and compared these to individuals who have previously cleared HCV. Genetic polymorphisms in the activating and inhibitory receptors expressed on these cells have been associated with the likelihood of spontaneous clearance in some, but not all, cohorts of patients with HCV [36]. There has been no clear consensus about whether the frequency or function of NK cells in chronic HCV infection, in part because of different methodologies used in the published studies. Several studies have reported that the frequency of NK cells in both peripheral blood [37; 38; 39; 40] and liver [41; 42] in chronic HCV compared to controls is decreased. However, Corado et al. reported that there were no differences in the frequency of CD3-CD56+ cells [43]. Functional studies of NK cell have also been conflicting. There is in vitro data that suggests that binding of the E2 protein to CD81 on NK cells inhibits NK cell function [44; 45]. This is consistent with the observation that patients with chronic HCV infection had lower levels of cytotoxicity of NK cells against sensitive cell lines (e.g. K562) than normal controls [43]. This has been confirmed by some [39; 40], but not all [46] groups. Even if one focuses on the CD56dim population, which comprises the NK subgroup with cytotoxic effector function, it is not clear that there are clear differences in the cytolytic capabilities of these cells when the number of effector cells is taken into account [37; 46]. While NKT cells are abundant in the liver, nothing is known about their functional role in acute HCV infection.
Dendritic cell (DC) function
DCs play an important role as antigen presenting cells (APCs) and link innate and adaptive immune responses. Two major subsets of DC are well known: myeloid DCs (mDCs, DC1), which predominantly secret IL12 and TNFα following pro-inflammatory stimuli and drive naïve CD4 cells to a Th1 phenotype; and plasmacytoid DCs (pDCs, DC2), which secret large amount of IFNα in viral infections and drive naïve CD4 T cells toward a Th2 phenotype[47; 48]. In vitro, HCV structural and non-structural proteins influence the capacity and differentiation of DC [49]. DCs generated from uninfected individuals and stimulated with HCV core and E1 protein are impaired in their capacity to stimulate allogenic T cell responses [50], and both DC expressing HCV core and NS3 protein have impaired differentiation [51].
Despite these in vitro studies, studies on DC function and outcome of HCV infection have produced contradictory results. In general, studies have compared individuals with a remote history of spontaneous resolution of HCV and individuals with chronic HCV infection, and compared these to either each other or to non-infected controls. Some studies have shown that mDCs generated from peripheral blood of chronically infected patients have impaired capacity to stimulate allogenic T cell responses and secrete IL12 [18; 52; 53]. In addition, reduced frequencies and impaired capacity of peripheral blood DCs in both acute and chronic HCV infection have been seen [54; 55]. However, not all studies have seen differences either in frequency [56] or function of pDC [57]. In some studies, pDC and mDC are be reduced in number but appear to be functional once differences in cell number are corrected for [58; 59]. Moreover, mDC from chronically infected chimpanzees are phenotypically and functionally normal [60]. These conflicting results may be due to methodologic issues, such as a failure to differentiate mDC and pDC in studies of whole blood, and definitive answers about whether DC dysfunction contributes to persistence in HCV await resolution.
Adaptive immune responses
Adaptive immune responses include humoral immune responses and cellular immune responses. Development of neutralizing antibodies is a hallmark of clearance in many viral infections and induction of these antibodies through immunization with viral subunits or inactivated virus is a classic strategy for the induction of protective immunity. HCV-specific antibodies (Ab) usually become detectable in the serum within several weeks after primary HCV infection, although the range is highly variable. There may be a delayed appearance of antibodies in individuals with persistent infection [61; 62]. Moreover, HCV-specific antibodies appear after cellular immune responses and aminotransferase elevations [63]. The first detectable antibodies against HCV antigens in serum usually target NS3 protein (anti-c33 Ab) and core protein (anti-capsid Ab or anti-22c Ab). Later, the specific antibodies against NS4 protein and envelope glycoproteins (E1 and E2) appear [64]. In particular, the hypervariable region 1 (HVR-1) of HCV E2 glycoprotein is thought to be a major target for neutralizing antibodies. Whether or not the typical antibodies measured in these studies conferred or was a marker of sterilizing immunity is controversial. In vivo studies in chimpanzees showed that the HCV-specific antibodies, specific for the HVR and with in vitro neutralizing ability, had protective effects against HCV infection [65]. On the other hand, other studies demonstrated that anti-HVR antibodies could not confer protective immunity against reinfection with homologous or heterologous strains [66; 67]. Moreover, spontaneous clearance of HCV also was reported in absence of humoral immune responses in a- or hypo-gammaglobulinemic patients [68; 69].
The recent development of assays based on the use of surrogate HCV particles has permitted re-assessment of humoral immune responses against conformational, as opposed to linear, HCV epitopes. The later studies suggest that antibodies may indeed have some role in spontaneous resolution of HCV and might be useful components of prophylactic vaccines. Two technologies have been used to identify neutralizing antibodies. Virus-like particles (VLPs) are HCV structural proteins (core/E1/E2 proteins) assembled into a structural mimic of the native HCV viral particle [70; 71]. Use of VLPs as immiunogens resulted in production of antibodies that prevented HCV entry in vitro [72; 73] and in vivo [74]. Pseudotyped retroviral particles (pps) are infectious virus, usually members of the lentivirus family, which are decorated with the HCV envelope glycoprotein on the surface. These HCV pps can be used to detect antibodies that inhibit infection and to demonstrate cross-genotype neutralization of HCV by antibodies in patients with chronic HCV infection [75; 76; 77; 78; 79]. The studies with HCV pp in humans and chimpanzees have demonstrated that HCV-specific neutralizing antibodies can be found in the late phase of chronic HCV infection. Moreover, these antibodies had cross-reactive activity, which meant they could response against both homologous and heterologous viral variants or HCV pps [76; 77; 79; 80; 81; 82]. A seminal study using a human liver-chimeric mouse model, in which immunodeficient mice have reconstitution with human hepatocytes, demonstrated the ability of antibodies to provide not only protection from homologous HCV isolated and heterologous HCV strains [83].
T cells in acute HCV infection
CD4+ T cells have multiple effector functions in antiviral responses, both via secreting antiviral cytokines and via activating viral specific B cells and CD8+ T cells. A strong preponderence of evidence demonstrates that in the acute phase of HCV infection, vigorous, broadly directed and sustained HCV-specific CD4+ and CD8+ T cell are closely associated with a self-limited course of infection [26; 84; 85; 86; 87; 88]. HCV-specific T cell responses and the induction of IFNγ in peripheral blood and liver coincide with a decrease in HCV RNA titers [26; 30; 89], although there is a notable lag between the onset of viremia and the onset of T cell responses [26; 89; 90]. There is no “immunodominant” response that is the critical determinant of protective immunity, as the immune response is broadly directed within the individual and the population. For example, Day et al. demonstrated immune responses against 10 of 37 defined epitopes using a proliferative assay in individuals with resolved HCV infection, whereas a responses against more than one epitope was never detected in those with chronic infection [85]. If the HCV-specifc CD4+ T cell responses are inefficient (weak, narrow and short-lasting or lost), persistent infection develops [85; 91]. The definitive role of CD4+ T cell responses in acute HCV was shown in a recent study in which the generation of HCV-specific CD4+ T cell responses was shown to be critical for successful control of HCV; without CD4+ T cell responses, HCV-specific CD8+ T cell and neutralizing antibodies may develop yet fail to control viremia [92].
Similarly, in both human and chimpanzee studies, HCV-specific CD8+ T cell responses in acute spontaneously resolving HCV also vigorous and target multiple epitopes [87; 88], whereas HCV-specific CD8+ responses among those individuals with evolution to chronic infection are lower in frequency and target only a few epitopes [93; 94; 95]. A key proof of principle that T cell responses may be critical to vaccine development was a recent chimpanzee study in which elicitation of peripheral and intrahepatic CD8+ T cell responses conferred protection against HCV and, of significance, against challenge with a heterologous isolate [96]. Elegant experiments in the chimpanzee model, in which it is possible to deplete specific cell populations, demonstrate the critical role of CD4+ and CD8+ T cells in the primary protective immunity against HCV [97; 98; 99]. In these studies, CD8+ T cells are the primary effector cells in protective immunity. However, without sustained CD4+ T cell help, CD8+ T cells cannot keep pace with the viral replication and ultimately escape mutations develop [98; 99].
CD8+ T cells have both cytolytic and non-cytolytic effector functions, the latter of which is mediated by production of cytokines such as IFNγ and TNFα[33]. Recent evidence in other viral systems implicates non-cytolytic mechanisms as a major effector mechanism of clearance [33], although the mechanisms whereby CD8+ T cells might control HCV replication are poorly understood. Drawing upon clear cut data in animal models of HBV infection [33], most investigators have focused on IFNγ as a key cytokine both necessary and sufficient for inhibiting HCV replication. Certainly in vitro data suggests that IFNγ, as well as IFNα/β, can inhibit HCV replication [34; 100]. However, IFNγ expression is abundant in within the liver, even in chronic HCV, and there is no evidence that the level of expression is related to HCV viral load; indeed, the level of IFNγ expression correlates positively with the degree of inflammation within liver tissue [101; 102]. A recent study examined the effect of IFNγ delivery directly into the liver via a gene therapy vector [103] and observed no effect on viral load. Whether other cytokines that might be produced by HCV-specific T cells play a role in controlling HCV replication awaits further study.
Immune escape
HCV has developed sophisticated escape mechanisms to evade host immune systems and it is likely that several different mechanisms are operational. In addition to interference with the endogenous IFN system and the potential for DC dysfunction, there is evidence for viral evasion of the adaptive immune response.
Subversion of innate immunity
Multiple lines of evidence suggest that HCV specifically interferes with the endogenous IFN and Toll-like receptor (TLR) response (summarized in Table 2 and reviewed in [11; 29; 104]). Retinoic-acid-inducible gene I (RIG-I) is an essential molecule for the IFNβ promoter [105] and leads to phosphorylation and activation of IRF-3, which promotes type I IFN production in infected cells in turn [11; 29; 104]. HCV NS3/4A serine protease blocks activation of virus induced IRF-3 [106] and inhibits type I IFN induction by disruption of RIG-I signaling [105; 107; 108; 109]. NS3/4A protease also inhibits TLR3 signaling through cleavage of the TRIF protein [110; 111] (Figure 2). Recent in vivo studies confirm that IRF-3 activation in the setting of chronic HCV is limited [112], which suggests that the endogenous IFN response is not fully operational.
Table 2.
Interactions of HCV with components of innate host defense
| HCV protein | Effect | References |
|---|---|---|
| Core | Inhibits activation or translocation of STAT-1 | [113; 114; 115; 116] |
| Increases degradation STAT-1 | [115; 117] | |
| Induces expression SOCS-3 | [113; 118; 119] | |
| Blocks DNA binding by ISGF3 | [120] | |
| Inhibits SOCS-1 | [218] | |
| E2 | Inhibits PKR activity | [128] |
| Inhibits NK cell function | [44; 45] | |
| NS3/4A | Inihibits TLR3 functon | [110; 111] |
| Block RIG-I signaling | [105; 107; 108; 109] | |
| NS5 | Represses activation of PKR | [121; 122; 123] |
| Blocks 2′-5′ OAS | [124] | |
| Increases IL8 | [125] |
STAT; signal transducer and activator of transcription, SOCS; suppressor of cytokine signaling, ISGF; interferon-stimulated gene factor, PKR; double-stranded RNA-dependent protein kinase, NK cell; natural killer cell, TLR; Toll-like receptor, RIG-I; retinoic-acid-inducible gene I, OAS; oligodenylate synthetase, IL; interleukin
Figure 2.
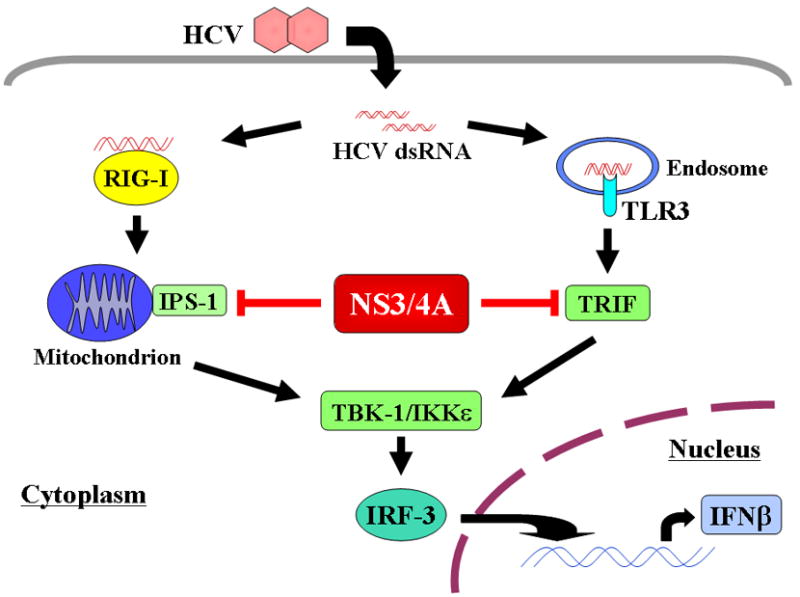
HCV RNA induces activation of IRF-3 via both the RIG-I and TLR3 pathways. Activated IRF-3 leads to IFNβ production and secretion from infected cell. HCV NS3/4A serine protease blocks these two pathways by cleavage of IPS-1 and TRIF.
Newly synthesized IFNβ binds to its cognate receptor and activates the expression of numerous IFN-stimulated genes (ISG) via the JAK/STAT pathway. Jak1 and Tyk2 are members of Janus kinase (JAK) family and phosphorylate signal transducer and activator of transcription (STAT) proteins. Phosphorylated STAT-1 and STAT-2 recruit IRF-9 to form a complex known as interferon-stimulated gene factor 3 (ISGF3). The ISGF3 complex locates to the cell nucleus and binds to IFN-stimulated response elements (IRES) in the promoter regions of ISG. This results in the expression of PKR, 2′-5′ OAS, ISG-p56, IRF-7 and other genes [11; 29; 104] (Figure 3). HCV core protein inhibits activation or translocation of STAT-1[113; 114; 115; 116] and increases degradation of STAT-1 [115; 117]. HCV core protein also inhibits the function of ISGF3 by inducing expression of suppressor of cytokine signaling 3 (SOCS-3) [104; 113; 118; 119] and blocks DNA binding by ISGF3 [120]. NS5A represses activation of PKR [121; 122; 123] through binding to PKR and inhibiting formulation of the PKR dimer [122]. NS5A also blocks the PKR-dependent activation of IRF-1 [123]. In addition, NS5A interacts with 2′-5′ OAS [124] and induces interleukin (IL) 8 [125], a chemokine that inhibits IFN activity in vivo and in vitro [126; 127]. The envelope protein E2 also binds PKR and inhibits its kinase activity [128]. Therefore, collectively HCV uses multiple pathways to disrupt a key mechanism of antiviral defenses.
Figure 3.

Newly synthesized IFNβ binds to its cognate receptor and activates the expression of numerous IFN-stimulated genes (ISG) via JAK/STAT pathway. HCV core protein inhibits this pathway via several mechanisms. HCV core protein inihibits activation or translocation of STAT-1 and formulation of ISGF3 by inducing SOCS-3. HCV core protein also blocks DNA binding by ISGF3. NS5A inhibits activation of PKR and 2′–5′ OAS. E2 inhibits activation of PKR.
“Exhaustion”/anergy
A key question in HCV immunopathogenesis is why T cells fail. As previously discussed, there is controversial data about whether T cell failure might be due to DC dysfunction, although data from acute infection in the chimpanzee or in humans is limited. Data from other model viral infections, such as lymphoctic chorinomeningitis virus (LCMV), suggests that T cell exhaustion might result from a high rate of viral replication that exceeds the capacity of host immunity [129]. One mechanism of persistence in HCV is failure to maintain CD4+ T cells during early infection [91], which may be on the basis of high viral loads. Others have shown that HCV-specific CD8+ T cells in chronic infection are functionally impaired with respect to IFNγ production or anergic [93; 130; 131] and, consistent with this loss of function, exhibit phenotypic changes characteristic of early stages of differentiation [132]. In the chimpanzee, HCV-specific CD8+ T cells in the acute phase do not produce IFNγ having a so-called “stunned” phenotype. In a later phase of infection, T cells recovered their ability to secret IFNγ, coinciding with substantial decrease of viral titer and resolution of hepatitis [26]. Whether HCV-specific T cell responses in the liver during chronic infection are functional or not in vivo is a matter of active scientific debate. Several studies have shown an impaired capacity of HCV-specific T cells to proliferate and secrete IFNγ against HCV proteins [93; 133]. However, other studies have shown IFNγ production both in vitro and ex vivo [134; 135; 136] and complete exhaustion with deletion of HCV-specific CD8+ T cells does not occur because CD8+ T cell responses can be restored with stimulation by IL2 and HCV antigens [137].
Several recent studies suggest that the PD-1/PD-1 ligand (PD-1L) pathway is up-regulated in those with chronic HCV and blockade of PD-1/PD-1L pathway had a beneficial effect on CD8+ T cells, restoring their ability leading to viral clearance, such as lymphocyte proliferation, secretion of cytokines and killing of infected cells [138; 139; 140]. However, a recent study demonstrated that expression of PD-1 was independent of whether HCV resolved spontaneously or became chronic [141].
Sequence mutations
Mutational escape from adaptive immune response has been thought to be one of major viral evasion strategies used by HCV. HCV replicates at a high rate, estimated at approximately 1010 to 1012 viral genome per day, even in the chronic phase of infection [142]. The mutation rate of HCV is estimated to be 1.5–2.0 × 10−3 base substitutions per genome site per year, due to the instrinic lack of proof reading of the RNA dependent RNA polymerase [143]. These characteristics of HCV contribute to the marked genomic diversity of HCV, with the potential to induce viral variants that can evade immune recognition [143; 144]. Erickson et al. first demonstrated immune escape from CD8+ T cell in the chimpanzee model [145]. Mutation occurred within multiple MHC class I restricted epitopes but not flanking regions of the HCV genome within the first few months of chronic infection. Mutations within CTL epitopes have also been seen in longitudinal studies of acutely infected patients and in single source outbreaks [146; 147; 148; 149] and by population based approaches, which sequence multiple strains and compare the observed patterns of viral variability with the known epitopes recognized in the context of that population’s HLA [147; 148; 150]. HCV mutations also affect virus specific CD8+ T cell responses by decreasing binding affinity between epitope and MHC molecule [149], decreasing T cell receptor (TCR) recognition [151] and impairing proteosomal processing of HCV antigens [152]. Mutational escape from CD4+ T cell responses has also been demonstrated [149; 153].
Regulatory T cell (Treg)
Recently, several studies have examined the role of regulatory T cells (Treg) in HCV infection and whether alterations in these T cells might influence the likelihood of chronicity. These cells maintain self-tolerance and limit deleterious immune responses [154; 155; 156]. Although there are likely multiple populations of Treg cells [157; 158], the majority of studies in HCV have focused on the CD4+CD25+ population (or “natural” Treg). The frequency of CD4+CD25+ Tregs was found to increase in the peripheral blood of chronically HCV infected individuals in comparison with resolved individuals or uninfected individuals. Depletion of CD4+CD25+ Tregs from peripheral blood led to peptide-specific IFNγ production and proliferation by HCV-specific CD8+ T cells [159; 160; 161; 162]. However, these studies may be limited by the fact that CD25 is expressed on activated T cells as well; definitive identification may require staining of forkhead box protein 3 (foxp3), for which reagents have only recently become available, or other markers such as CD127 [157; 158]. Indeed, a recent study in chimpanzees showed no difference in CD4+CD25+foxp3+ cells in the periphery or liver irrespective of the outcome of infection [163].
Immune responses in chronic infection: friend or foe?
Many studies confirm that HCV-specific immune responses in the periphery are weak and directed against a limited series of epitopes in chronic infection compared to those responses observed in spontaneous resolution of HCV. However, immune responses appear to compartmentalize very early to the liver tissue, thus complicating studies of pathogenesis of chronic liver injury. Many studies have demonstrated that in the setting of chronic infection both HCV-specific CD4+ and CD8+ T cells are present in the infected liver [133; 134; 135; 136; 164; 165; 166; 167; 168; 169; 170; 171]. The ratio of intrahepatic HCV-specific T cells to peripheral T cells may be 100:1 or more. Little is known about the process whereby HCV-specific T cells home to the liver, although there is likely a series of events triggered by the endogenous IFN response. IFN α/β produced by hepatocytes can induce the production of monocyte chemoattractant protein-1 (MCP-1) by the resident liver macrophages, which leads to the recruitment of other macrophages that produce macrophage inflammatory protein-1 alpha (MIP-1α). MIP-1α recruits NK cells, which in turn serve as a source of IFNγ; this cytokine may be antiviral but also leads to the production of monokine induced by IFNγ (MIG or CXCL9), which recruits T cells to the site of inflammation [172]. IFN-inducible protein-10 (IP-10), which correlates with the degree of inflammation, is also increased in chronic HCV [173]. HCV itself may upregulate critical homing molecules involved in T cell recruitment such as interferon-inducible T cell alpha chemoattractant (I-TAC) [174], the expression of which correlates with the degree of hepatic inflammation [175]. This data is consistent with the observation that HCV transgenic mice, despite constitutive tolerance to HCV proteins, may develop inflammatory liver lesions [176; 177].
Clinical data strongly suggests that patients with depressed cellular immunity, such as those with liver allografts or human immunodeficiency virus, have an accelerated course of HCV, and may develop cirrhosis much more rapidly than non-immunocompromised hosts [178; 179]. Prospective studies done early in the course of HCV infection demonstrate that the magnitude of HCV-specific CD4+ T cell responses clearly determine the rate of progression of chronic liver disease [180]; this was confirmed in later cross sectional analysis of CD4+ T cell responses using IFNβ linked ELISpot [181]. Similarly, the magnitude of the CD4+ T cell response in the liver appears to be inversely proportional to the degree of liver injury [181]. Although most investigators have approached Treg as a potential factor in mediating chronicity, a recent study demonstrated that CD4+CD25+ Treg may have an inverse relationship to liver inflammation [182]. Therefore, CD4+ responses in general appear to be protective in the setting of chronic infection, even if they do not completely prevent disease.
Conflicting reports have been described about the role of HCV-specific CD8+ T cell in the chronic phase of HCV infection. Classic adoptive transfer studies, as well as newer murine models in which HCV expression is induced after the period of neonatal tolerance, strongly suggest that CD8+ T cells mediate liver injury [183; 184]. Clinical studies have been inconsistent with respect to the relationship between the presence of CD8+ T cells and the HCV viral load [166; 185; 186; 187; 188; 189]; however, HCV viral load does not predict the extent of liver injury. Similarly, the presence of an HCV-specific CD8+ T cell response has an inconsistent relationship to the severity of liver injury on biopsy [102; 190]. The inflammatory infiltrate in chronic HCV appears to consist of a large number of T cells that are not specific for HCV [191; 192]; this “bystander activation” may be critical in liver injury [192; 193]. However, there are also novel CD8+ T cell populations within the liver that may inhibit inflammation through production of IL-10 [156; 192] or through TGF-β [194]. Further resolution of this issue awaits larger, prospective trials with cohorts with known duration of HCV infection and/or precise evaluation of the rate of liver injury using serial examinations.
Different NK subsets may have different roles in the pathogenesis of liver injury, but this too is an area that needs further development. Lin et al. found that activated CD56bright correlated with inflammation but not fibrosis [38] and Morishima et al. found that CD56dim correlated inversely with fibrosis stage [37]. Whether low levels of CD56dim cells are the cause of increased fibrosis, by failing to protect the liver, or are simply associated with longstanding disease, as there may be apoptosis and loss of these cells over time, will require prospective studies.
Role of immune responses in outcome following IFN and ribavirin treatment
The current standard of treatment of HCV is a combination therapy with pegylated IFNβ (peginterferon) and ribavirin. Overall sustained virologic response (SVR), defined as the absence of virus in the blood 24 weeks after therapy is ended, in about 50% of chronic infected patients treated with combination therapy [195]. However, SVR rates vary depending on viral (e.g., viral genotype and viral load) and host characteristics (e.g., gender, age, ethnic background and fibrosis stage) [196; 197]. For example, SVR rates are typically 42 to 52% among patients with HCV genotype 1 and 76 to 82% of those with genotype 2 or 3 [195; 198; 199; 200; 201]. Why certain genotypes are more responsive to treatment is not known, and is only beginning to be explored as culture systems for both genotype 1 and non-1 isolates become available. Similarly, it is clear that African Americans have lower SVR rates than Caucasians [202; 203]; African Americans have lower baseline CD4+ responses against HCV than Caucasians but it is not clear whether this is the basis for the observed difference in SVR [204; 205].
In addition to direct antiviral effects, both IFNβ and ribavirin may be immunomodulatory: IFNβ enhances APC maturation and CD4+ Tcell function, but with little effect on CTLs [206; 207] and ribavirin induces a switch from Th2 to Th1 profile [208] and inhibits IL10 production [209]. As new antiviral agents are developed that specifically target HCV proteins [210; 211], it is not known whether IFN and/or ribavirin or any immodulation will still need to be part of the regimen to lead to cure. Surprisingly, there is relatively little data on whether or not T cell responses at baseline or during treatment are associated with SVR. Cramp et al. demonstrated that higher CD4+ T cell proliferation in the early phase of therapy was correlated with SVR [209], with a more recent studies demonstrating that the baseline immune response is the critical determinant of SVR rather than enhancement of immune responses on treatment [205; 212]. In a study of individuals with HIV/HCV co-infection, HCV-specific production of IL-10 was a stronger predictor of response that other well known host factors such as race [213].
Since treatment of acute HCV typically leads to high SVR rates even in genotype 1 infection – at least 80% SVR compared to approximately 55% SVR in the setting of chronic HCV – the host factors associated with this have been studied. Kamal et al. demonstrated that the high rate of SVR in acute HCV is associated with strong T cell responses [214], although this has not been seen in every study [62]. A rapid drop in viral load on treatment is associated with more vigorous T cell responses, although whether this is cause or effect is not known [215]. Other host factors in the IFN-regulatory pathway might be associated with the responses to antiviral treatment [216; 217]. Interferon-sensitive gene 15 (ISG15) and ubiquitin-specific protease 18 (USP18), which are linked biochemically in IFN-regulatory pathway, modulate the IFN responses against HCV [216]. USP18 up-regulation is one of the factors predicting a lack of response to IFN treatment [217].
Conclusions and future directions
In recent years, a more complete understanding of the correlates of protective immunity in HCV has been achieved. There is broad consensus that induction of CD4+ T cells and probably CD8+ T cells targeting multiple viral epitopes will be critical to the development of an effective prophylactic vaccine; however, there are considerable technical challenges in formulating a vaccine that would be effective against the multiple circulating quasispecies. There is much less understanding of how HCV leads to persistent infection in most exposed persons, although it is likely that this virus uses multiple mechanisms to evade the immune system. The area in which there is least consensus is the role of T cells in resolution of HCV following during treatment and the mechanisms of liver injury. Further study of these areas is needed as we enter a new generation of HCV therapeutics that do not target the immune system, and to address the needs of the large number of patients with chronic HCV who face the threat of cirrhosis and liver cancer.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Choo QL, Kuo G, Weiner AJ, Overby LR, Bradley DW, Houghton M. Isolation of a cDNA clone derived from a blood-borne non-A, non-B viral hepatitis genome. Science. 1989;244:359–62. doi: 10.1126/science.2523562. [DOI] [PubMed] [Google Scholar]
- 2.Shepard CW, Finelli L, Alter MJ. Global epidemiology of hepatitis C virus infection. Lancet Infect Dis. 2005;5:558–67. doi: 10.1016/S1473-3099(05)70216-4. [DOI] [PubMed] [Google Scholar]
- 3.Bialek SR, Terrault NA. The changing epidemiology and natural history of hepatitis C virus infection. Clin Liver Dis. 2006;10:697–715. doi: 10.1016/j.cld.2006.08.003. [DOI] [PubMed] [Google Scholar]
- 4.NIH Consensus Statement on Management of Hepatitis C: 2002. NIH Consens State Sci Statements 19 (2002) 1–46.
- 5.Schiff ER, Ozden N. Hepatitis C and alcohol. Alcohol Res Health. 2003;27:232–9. [PMC free article] [PubMed] [Google Scholar]
- 6.Weigand K, Stremmel W, Encke J. Treatment of hepatitis C virus infection. World J Gastroenterol. 2007;13:1897–905. doi: 10.3748/wjg.v13.i13.1897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Penin F, Dubuisson J, Rey FA, Moradpour D, Pawlotsky JM. Structural biology of hepatitis C virus. Hepatology. 2004;39:5–19. doi: 10.1002/hep.20032. [DOI] [PubMed] [Google Scholar]
- 8.Simmonds P, Bukh J, Combet C, Deleage G, Enomoto N, Feinstone S, Halfon P, Inchauspe G, Kuiken C, Maertens G, Mizokami M, Murphy DG, Okamoto H, Pawlotsky JM, Penin F, Sablon E, Shin IT, Stuyver LJ, Thiel HJ, Viazov S, Weiner AJ, Widell A. Consensus proposals for a unified system of nomenclature of hepatitis C virus genotypes. Hepatology. 2005;42:962–73. doi: 10.1002/hep.20819. [DOI] [PubMed] [Google Scholar]
- 9.Lindenbach BD, Rice CM. Unravelling hepatitis C virus replication from genome to function. Nature. 2005;436:933–8. doi: 10.1038/nature04077. [DOI] [PubMed] [Google Scholar]
- 10.Lloyd AR, Jagger E, Post JJ, Crooks LA, Rawlinson WD, Hahn YS, Ffrench RA. Host and viral factors in the immunopathogenesis of primary hepatitis C virus infection. Immunol Cell Biol. 2007;85:24–32. doi: 10.1038/sj.icb.7100010. [DOI] [PubMed] [Google Scholar]
- 11.Thimme R, Lohmann V, Weber F. A target on the move: innate and adaptive immune escape strategies of hepatitis C virus. Antiviral Res. 2006;69:129–41. doi: 10.1016/j.antiviral.2005.12.001. [DOI] [PubMed] [Google Scholar]
- 12.Liang TJ, Rehermann B, Seeff LB, Hoofnagle JH. Pathogenesis, natural history, treatment, and prevention of hepatitis C. Ann Intern Med. 2000;132:296–305. doi: 10.7326/0003-4819-132-4-200002150-00008. [DOI] [PubMed] [Google Scholar]
- 13.Inoue Y, Murakami K, Hmwe S, Aizaki H, Suzuki T. Transcriptomic comparison of human hepatoma huh-7 cell clones with different hepatitis C virus replication efficiencies. Jpn J Infect Dis. 2007;60:173–8. [PubMed] [Google Scholar]
- 14.Pachiadakis I, Pollara G, Chain BM, Naoumov NV. Is hepatitis C virus infection of dendritic cells a mechanism facilitating viral persistence? Lancet Infect Dis. 2005;5:296–304. doi: 10.1016/S1473-3099(05)70114-6. [DOI] [PubMed] [Google Scholar]
- 15.Muratori L, Gibellini D, Lenzi M, Cataleta M, Muratori P, Morelli MC, Bianchi FB. Quantification of hepatitis C virus-infected peripheral blood mononuclear cells by in situ reverse transcriptase-polymerase chain reaction. Blood. 1996;88:2768–74. [PubMed] [Google Scholar]
- 16.Lerat H, Rumin S, Habersetzer F, Berby F, Trabaud MA, Trepo C, Inchauspe G. In vivo tropism of hepatitis C virus genomic sequences in hematopoietic cells: influence of viral load, viral genotype, and cell phenotype. Blood. 1998;91:3841–9. [PubMed] [Google Scholar]
- 17.Garcia F, Jr, Garcia F, Roldan C, Lopez I, Martinez NM, Alvarez M, Bernal MC, Hernandez J, Maroto MC. Detection of HCV and GBV-CHGV RNA in peripheral blood mononuclear cells of patients with chronic type C hepatitis. Microbios. 2000;103:7–15. [PubMed] [Google Scholar]
- 18.Bain C, Fatmi A, Zoulim F, Zarski JP, Trepo C, Inchauspe G. Impaired allostimulatory function of dendritic cells in chronic hepatitis C infection. Gastroenterology. 2001;120:512–24. doi: 10.1053/gast.2001.21212. [DOI] [PubMed] [Google Scholar]
- 19.Sung VM, Shimodaira S, Doughty AL, Picchio GR, Can H, Yen TS, Lindsay KL, Levine AM, Lai MM. Establishment of B-cell lymphoma cell lines persistently infected with hepatitis C virus in vivo and in vitro: the apoptotic effects of virus infection. J Virol. 2003;77:2134–46. doi: 10.1128/JVI.77.3.2134-2146.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Rosa D, Saletti G, De Gregorio E, Zorat F, Comar C, D'Oro U, Nuti S, Houghton M, Barnaba V, Pozzato G, Abrignani S. Activation of naive B lymphocytes via CD81, a pathogenetic mechanism for hepatitis C virus-associated B lymphocyte disorders. Proc Natl Acad Sci U S A. 2005;102:18544–9. doi: 10.1073/pnas.0509402102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Carbonari M, Caprini E, Tedesco T, Mazzetta F, Tocco V, Casato M, Russo G, Fiorilli M. Hepatitis C virus drives the unconstrained monoclonal expansion of VH1-69-expressing memory B cells in type II cryoglobulinemia: a model of infection-driven lymphomagenesis. J Immunol. 2005;174:6532–9. doi: 10.4049/jimmunol.174.10.6532. [DOI] [PubMed] [Google Scholar]
- 22.Charles ED, Green RM, Marukian S, Talal AH, Lake-Bakaar GV, Jacobson IM, Rice CM, Dustin LB. Clonal expansion of IgM+CD27+ B cells in HCV-associated mixed cryoglobulinemia. Blood. 2007 doi: 10.1182/blood-2007-07-101717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Giordano TP, Henderson L, Landgren O, Chiao EY, Kramer JR, El-Serag H, Engels EA. Risk of non-Hodgkin lymphoma and lymphoproliferative precursor diseases in US veterans with hepatitis C virus. JAMA. 2007;297:2010–7. doi: 10.1001/jama.297.18.2010. [DOI] [PubMed] [Google Scholar]
- 24.Lindenbach BD, Meuleman P, Ploss A, Vanwolleghem T, Syder AJ, McKeating JA, Lanford RE, Feinstone SM, Major ME, Leroux-Roels G, Rice CM. Cell culture-grown hepatitis C virus is infectious in vivo and can be recultured in vitro. Proc Natl Acad Sci U S A. 2006;103:3805–9. doi: 10.1073/pnas.0511218103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Mercer DF, Schiller DE, Elliott JF, Douglas DN, Hao C, Rinfret A, Addison WR, Fischer KP, Churchill TA, Lakey JR, Tyrrell DL, Kneteman NM. Hepatitis C virus replication in mice with chimeric human livers. Nat Med. 2001;7:927–33. doi: 10.1038/90968. [DOI] [PubMed] [Google Scholar]
- 26.Thimme R, Oldach D, Chang KM, Steiger C, Ray SC, Chisari FV. Determinants of viral clearance and persistence during acute hepatitis C virus infection. J Exp Med. 2001;194:1395–406. doi: 10.1084/jem.194.10.1395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Abe K, Inchauspe G, Shikata T, Prince AM. Three different patterns of hepatitis C virus infection in chimpanzees. Hepatology. 1992;15:690–5. doi: 10.1002/hep.1840150423. [DOI] [PubMed] [Google Scholar]
- 28.Beach MJ, Meeks EL, Mimms LT, Vallari D, DuCharme L, Spelbring J, Taskar S, Schleicher JB, Krawczynski K, Bradley DW. Temporal relationships of hepatitis C virus RNA and antibody responses following experimental infection of chimpanzees. J Med Virol. 1992;36:226–37. doi: 10.1002/jmv.1890360314. [DOI] [PubMed] [Google Scholar]
- 29.Gale M, Jr, Foy EM. Evasion of intracellular host defence by hepatitis C virus. Nature. 2005;436:939–45. doi: 10.1038/nature04078. [DOI] [PubMed] [Google Scholar]
- 30.Su AI, Pezacki JP, Wodicka L, Brideau AD, Supekova L, Thimme R, Wieland S, Bukh J, Purcell RH, Schultz PG, Chisari FV. Genomic analysis of the host response to hepatitis C virus infection. Proc Natl Acad Sci U S A. 2002;99:15669–74. doi: 10.1073/pnas.202608199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Bigger CB, Brasky KM, Lanford RE. DNA microarray analysis of chimpanzee liver during acute resolving hepatitis C virus infection. J Virol. 2001;75:7059–66. doi: 10.1128/JVI.75.15.7059-7066.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Shin EC, Seifert U, Kato T, Rice CM, Feinstone SM, Kloetzel PM, Rehermann B. Virus-induced type I IFN stimulates generation of immunoproteasomes at the site of infection. J Clin Invest. 2006;116:3006–14. doi: 10.1172/JCI29832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Guidotti LG, Chisari FV. Noncytolytic control of viral infections by the innate and adaptive immune response. Annu Rev Immunol. 2001;19:65–91. doi: 10.1146/annurev.immunol.19.1.65. [DOI] [PubMed] [Google Scholar]
- 34.Frese M, Schwarzle V, Barth K, Krieger N, Lohmann V, Mihm S, Haller O, Bartenschlager R. Interferon-gamma inhibits replication of subgenomic and genomic hepatitis C virus RNAs. Hepatology. 2002;35:694–703. doi: 10.1053/jhep.2002.31770. [DOI] [PubMed] [Google Scholar]
- 35.Brown MG, Dokun AO, Heusel JW, Smith HR, Beckman DL, Blattenberger EA, Dubbelde CE, Stone LR, Scalzo AA, Yokoyama WM. Vital involvement of a natural killer cell activation receptor in resistance to viral infection. Science. 2001;292:934–7. doi: 10.1126/science.1060042. [DOI] [PubMed] [Google Scholar]
- 36.Khakoo SI, Thio CL, Martin MP, Brooks CR, Gao X, Astemborski J, Cheng J, Goedert JJ, Vlahov D, Hilgartner M, Cox S, Little AM, Alexander GJ, Cramp ME, O'Brien SJ, Rosenberg WM, Thomas DL, Carrington M. HLA and NK cell inhibitory receptor genes in resolving hepatitis C virus infection. Science. 2004;305:872–4. doi: 10.1126/science.1097670. [DOI] [PubMed] [Google Scholar]
- 37.Morishima C, Paschal DM, Wang CC, Yoshihara CS, Wood BL, Yeo AE, Emerson SS, Shuhart MC, Gretch DR. Decreased NK cell frequency in chronic hepatitis C does not affect ex vivo cytolytic killing. Hepatology. 2006;43:573–80. doi: 10.1002/hep.21073. [DOI] [PubMed] [Google Scholar]
- 38.Lin AW, Gonzalez SA, Cunningham-Rundles S, Dorante G, Marshall S, Tignor A, Ha C, Jacobson IM, Talal AH. CD56(+dim) and CD56(+bright) cell activation and apoptosis in hepatitis C virus infection. Clin Exp Immunol. 2004;137:408–16. doi: 10.1111/j.1365-2249.2004.02523.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Meier UC, Owen RE, Taylor E, Worth A, Naoumov N, Willberg C, Tang K, Newton P, Pellegrino P, Williams I, Klenerman P, Borrow P. Shared alterations in NK cell frequency, phenotype, and function in chronic human immunodeficiency virus and hepatitis C virus infections. J Virol. 2005;79:12365–74. doi: 10.1128/JVI.79.19.12365-12374.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Par G, Rukavina D, Podack ER, Horanyi M, Szekeres-Bartho J, Hegedus G, Paal M, Szereday L, Mozsik G, Par A. Decrease in CD3-negative-CD8dim(+) and Vdelta2/Vgamma9 TcR+ peripheral blood lymphocyte counts, low perforin expression and the impairment of natural killer cell activity is associated with chronic hepatitis C virus infection. J Hepatol. 2002;37:514–22. doi: 10.1016/s0168-8278(02)00218-0. [DOI] [PubMed] [Google Scholar]
- 41.Deignan T, Curry MP, Doherty DG, Golden-Mason L, Volkov Y, Norris S, Nolan N, Traynor O, McEntee G, Hegarty JE, O'Farrelly C. Decrease in hepatic CD56(+) T cells and V alpha 24(+) natural killer T cells in chronic hepatitis C viral infection. J Hepatol. 2002;37:101–8. doi: 10.1016/s0168-8278(02)00072-7. [DOI] [PubMed] [Google Scholar]
- 42.Kawarabayashi N, Seki S, Hatsuse K, Ohkawa T, Koike Y, Aihara T, Habu Y, Nakagawa R, Ami K, Hiraide H, Mochizuki H. Decrease of CD56(+)T cells and natural killer cells in cirrhotic livers with hepatitis C may be involved in their susceptibility to hepatocellular carcinoma. Hepatology. 2000;32:962–9. doi: 10.1053/jhep.2000.19362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Corado J, Toro F, Rivera H, Bianco NE, Deibis L, De Sanctis JB. Impairment of natural killer (NK) cytotoxic activity in hepatitis C virus (HCV) infection. Clin Exp Immunol. 1997;109:451–7. doi: 10.1046/j.1365-2249.1997.4581355.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Crotta S, Stilla A, Wack A, D'Andrea A, Nuti S, D'Oro U, Mosca M, Filliponi F, Brunetto RM, Bonino F, Abrignani S, Valiante NM. Inhibition of natural killer cells through engagement of CD81 by the major hepatitis C virus envelope protein. J Exp Med. 2002;195:35–41. doi: 10.1084/jem.20011124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Tseng CT, Klimpel GR. Binding of the hepatitis C virus envelope protein E2 to CD81 inhibits natural killer cell functions. J Exp Med. 2002;195:43–9. doi: 10.1084/jem.20011145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Duesberg U, Schneiders AM, Flieger D, Inchauspe G, Sauerbruch T, Spengler U. Natural cytotoxicity and antibody-dependent cellular cytotoxicity (ADCC) is not impaired in patients suffering from chronic hepatitis C. J Hepatol. 2001;35:650–7. doi: 10.1016/s0168-8278(01)00194-5. [DOI] [PubMed] [Google Scholar]
- 47.Liu YJ, Kadowaki N, Rissoan MC, Soumelis V. T cell activation and polarization by DC1 and DC2. Curr Top Microbiol Immunol. 2000;251:149–59. doi: 10.1007/978-3-642-57276-0_19. [DOI] [PubMed] [Google Scholar]
- 48.Liu YJ. Dendritic cell subsets and lineages, and their functions in innate and adaptive immunity. Cell. 2001;106:259–62. doi: 10.1016/s0092-8674(01)00456-1. [DOI] [PubMed] [Google Scholar]
- 49.Szabo G, Dolganiuc A. Subversion of plasmacytoid and myeloid dendritic cell functions in chronic HCV infection. Immunobiology. 2005;210:237–47. doi: 10.1016/j.imbio.2005.05.018. [DOI] [PubMed] [Google Scholar]
- 50.Sarobe P, Lasarte JJ, Casares N, Lopez-Diaz de Cerio A, Baixeras E, Labarga P, Garcia N, Borras-Cuesta F, Prieto J. Abnormal priming of CD4(+) T cells by dendritic cells expressing hepatitis C virus core and E1 proteins. J Virol. 2002;76:5062–70. doi: 10.1128/JVI.76.10.5062-5070.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Dolganiuc A, Kodys K, Kopasz A, Marshall C, Do T, Romics L, Jr, Mandrekar P, Zapp M, Szabo G. Hepatitis C virus core and nonstructural protein 3 proteins induce pro- and anti-inflammatory cytokines and inhibit dendritic cell differentiation. J Immunol. 2003;170:5615–24. doi: 10.4049/jimmunol.170.11.5615. [DOI] [PubMed] [Google Scholar]
- 52.Kanto T, Hayashi N, Takehara T, Tatsumi T, Kuzushita N, Ito A, Sasaki Y, Kasahara A, Hori M. Impaired allostimulatory capacity of peripheral blood dendritic cells recovered from hepatitis C virus-infected individuals. J Immunol. 1999;162:5584–91. [PubMed] [Google Scholar]
- 53.Della Bella S, Crosignani A, Riva A, Presicce P, Benetti A, Longhi R, Podda M, Villa ML. Decrease and dysfunction of dendritic cells correlate with impaired hepatitis C virus-specific CD4+ T-cell proliferation in patients with hepatitis C virus infection. Immunology. 2007;121:283–92. doi: 10.1111/j.1365-2567.2007.02577.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Kanto T, Inoue M, Miyatake H, Sato A, Sakakibara M, Yakushijin T, Oki C, Itose I, Hiramatsu N, Takehara T, Kasahara A, Hayashi N. Reduced numbers and impaired ability of myeloid and plasmacytoid dendritic cells to polarize T helper cells in chronic hepatitis C virus infection. J Infect Dis. 2004;190:1919–26. doi: 10.1086/425425. [DOI] [PubMed] [Google Scholar]
- 55.Ulsenheimer A, Gerlach JT, Jung MC, Gruener N, Wachtler M, Backmund M, Santantonio T, Schraut W, Heeg MH, Schirren CA, Zachoval R, Pape GR, Diepolder HM. Plasmacytoid dendritic cells in acute and chronic hepatitis C virus infection. Hepatology. 2005;41:643–51. doi: 10.1002/hep.20592. [DOI] [PubMed] [Google Scholar]
- 56.Piccioli D, Tavarini S, Nuti S, Colombatto P, Brunetto M, Bonino F, Ciccorossi P, Zorat F, Pozzato G, Comar C, Abrignani S, Wack A. Comparable functions of plasmacytoid and monocyte-derived dendritic cells in chronic hepatitis C patients and healthy donors. J Hepatol. 2005;42:61–7. doi: 10.1016/j.jhep.2004.09.014. [DOI] [PubMed] [Google Scholar]
- 57.Decalf J, Fernandes S, Longman R, Ahloulay M, Audat F, Lefrerre F, Rice CM, Pol S, Albert ML. Plasmacytoid dendritic cells initiate a complex chemokine and cytokine network and are a viable drug target in chronic HCV patients. J Exp Med. 2007;204:2423–37. doi: 10.1084/jem.20070814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Longman RS, Talal AH, Jacobson IM, Albert ML, Rice CM. Presence of functional dendritic cells in patients chronically infected with hepatitis C virus. Blood. 2004;103:1026–9. doi: 10.1182/blood-2003-04-1339. [DOI] [PubMed] [Google Scholar]
- 59.Longman RS, Talal AH, Jacobson IM, Rice CM, Albert ML. Normal functional capacity in circulating myeloid and plasmacytoid dendritic cells in patients with chronic hepatitis C. J Infect Dis. 2005;192:497–503. doi: 10.1086/431523. [DOI] [PubMed] [Google Scholar]
- 60.Rollier C, Drexhage JA, Verstrepen BE, Verschoor EJ, Bontrop RE, Koopman G, Heeney JL. Chronic hepatitis C virus infection established and maintained in chimpanzees independent of dendritic cell impairment. Hepatology. 2003;38:851–8. doi: 10.1053/jhep.2003.50426. [DOI] [PubMed] [Google Scholar]
- 61.Alter HJ, Purcell RH, Shih JW, Melpolder JC, Houghton M, Choo QL, Kuo G. Detection of antibody to hepatitis C virus in prospectively followed transfusion recipients with acute and chronic non-A, non-B hepatitis. N Engl J Med. 1989;321:1494–500. doi: 10.1056/NEJM198911303212202. [DOI] [PubMed] [Google Scholar]
- 62.Rahman F, Heller T, Sobao Y, Mizukoshi E, Nascimbeni M, Alter H, Herrine S, Hoofnagle J, Liang TJ, Rehermann B. Effects of antiviral therapy on the cellular immune response in acute hepatitis C. Hepatology. 2004;40:87–97. doi: 10.1002/hep.20253. [DOI] [PubMed] [Google Scholar]
- 63.Heller T, Rehermann B. Acute hepatitis C: a multifaceted disease. Semin Liver Dis. 2005;25:7–17. doi: 10.1055/s-2005-864778. [DOI] [PubMed] [Google Scholar]
- 64.Orland JR, Wright TL, Cooper S. Acute hepatitis C. Hepatology. 2001;33:321–7. doi: 10.1053/jhep.2001.22112. [DOI] [PubMed] [Google Scholar]
- 65.Farci P, Alter HJ, Wong DC, Miller RH, Govindarajan S, Engle R, Shapiro M, Purcell RH. Prevention of hepatitis C virus infection in chimpanzees after antibody-mediated in vitro neutralization. Proc Natl Acad Sci U S A. 1994;91:7792–6. doi: 10.1073/pnas.91.16.7792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Farci P, Alter HJ, Govindarajan S, Wong DC, Engle R, Lesniewski RR, Mushahwar IK, Desai SM, Miller RH, Ogata N, et al. Lack of protective immunity against reinfection with hepatitis C virus. Science. 1992;258:135–40. doi: 10.1126/science.1279801. [DOI] [PubMed] [Google Scholar]
- 67.Lai ME, Mazzoleni AP, Argiolu F, De Virgilis S, Balestrieri A, Purcell RH, Cao A, Farci P. Hepatitis C virus in multiple episodes of acute hepatitis in polytransfused thalassaemic children. Lancet. 1994;343:388–90. doi: 10.1016/s0140-6736(94)91224-6. [DOI] [PubMed] [Google Scholar]
- 68.Post JJ, Pan Y, Freeman AJ, Harvey CE, White PA, Palladinetti P, Haber PS, Marinos G, Levy MH, Kaldor JM, Dolan KA, Ffrench RA, Lloyd AR, Rawlinson WD. Clearance of hepatitis C viremia associated with cellular immunity in the absence of seroconversion in the hepatitis C incidence and transmission in prisons study cohort. J Infect Dis. 2004;189:1846–55. doi: 10.1086/383279. [DOI] [PubMed] [Google Scholar]
- 69.Bjoro K, Froland SS, Yun Z, Samdal HH, Haaland T. Hepatitis C infection in patients with primary hypogammaglobulinemia after treatment with contaminated immune globulin. N Engl J Med. 1994;331:1607–11. doi: 10.1056/NEJM199412153312402. [DOI] [PubMed] [Google Scholar]
- 70.Steinmann D, Barth H, Gissler B, Schurmann P, Adah MI, Gerlach JT, Pape GR, Depla E, Jacobs D, Maertens G, Patel AH, Inchauspe G, Liang TJ, Blum HE, Baumert TF. Inhibition of hepatitis C virus-like particle binding to target cells by antiviral antibodies in acute and chronic hepatitis C. J Virol. 2004;78:9030–40. doi: 10.1128/JVI.78.17.9030-9040.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Baumert TF, Wellnitz S, Aono S, Satoi J, Herion D, Tilman Gerlach J, Pape GR, Lau JY, Hoofnagle JH, Blum HE, Liang TJ. Antibodies against hepatitis C virus-like particles and viral clearance in acute and chronic hepatitis C. Hepatology. 2000;32:610–7. doi: 10.1053/jhep.2000.9876. [DOI] [PubMed] [Google Scholar]
- 72.Denis J, Majeau N, Acosta-Ramirez E, Savard C, Bedard MC, Simard S, Lecours K, Bolduc M, Pare C, Willems B, Shoukry N, Tessier P, Lacasse P, Lamarre A, Lapointe R, Lopez Macias C, Leclerc D. Immunogenicity of papaya mosaic virus-like particles fused to a hepatitis C virus epitope: evidence for the critical function of multimerization. Virology. 2007;363:59–68. doi: 10.1016/j.virol.2007.01.011. [DOI] [PubMed] [Google Scholar]
- 73.Vietheer PT, Boo I, Drummer HE, Netter HJ. Immunizations with chimeric hepatitis B virus-like particles to induce potential anti-hepatitis C virus neutralizing antibodies. Antivir Ther. 2007;12:477–87. [PubMed] [Google Scholar]
- 74.Elmowalid GA, Qiao M, Jeong SH, Borg BB, Baumert TF, Sapp RK, Hu Z, Murthy K, Liang TJ. Immunization with hepatitis C virus-like particles results in control of hepatitis C virus infection in chimpanzees. Proc Natl Acad Sci U S A. 2007;104:8427–32. doi: 10.1073/pnas.0702162104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Bartosch B, Dubuisson J, Cosset FL. Infectious hepatitis C virus pseudo-particles containing functional E1-E2 envelope protein complexes. J Exp Med. 2003;197:633–42. doi: 10.1084/jem.20021756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Bartosch B, Bukh J, Meunier JC, Granier C, Engle RE, Blackwelder WC, Emerson SU, Cosset FL, Purcell RH. In vitro assay for neutralizing antibody to hepatitis C virus: evidence for broadly conserved neutralization epitopes. Proc Natl Acad Sci U S A. 2003;100:14199–204. doi: 10.1073/pnas.2335981100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Logvinoff C, Major ME, Oldach D, Heyward S, Talal A, Balfe P, Feinstone SM, Alter H, Rice CM, McKeating JA. Neutralizing antibody response during acute and chronic hepatitis C virus infection. Proc Natl Acad Sci U S A. 2004;101:10149–54. doi: 10.1073/pnas.0403519101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Hsu M, Zhang J, Flint M, Logvinoff C, Cheng-Mayer C, Rice CM, McKeating JA. Hepatitis C virus glycoproteins mediate pH-dependent cell entry of pseudotyped retroviral particles. Proc Natl Acad Sci U S A. 2003;100:7271–6. doi: 10.1073/pnas.0832180100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Meunier JC, Engle RE, Faulk K, Zhao M, Bartosch B, Alter H, Emerson SU, Cosset FL, Purcell RH, Bukh J. Evidence for cross-genotype neutralization of hepatitis C virus pseudo-particles and enhancement of infectivity by apolipoprotein C1. Proc Natl Acad Sci U S A. 2005;102:4560–5. doi: 10.1073/pnas.0501275102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Perotti M, Mancini N, Diotti RA, Tarr AW, Ball JK, Owsianka A, Adair R, Patel AH, Clementi M, Burioni R. Identification of a broadly cross-reacting and neutralizing human monoclonal antibody directed against the hepatitis C virus E2 protein. J Virol. 2008;82:1047–52. doi: 10.1128/JVI.01986-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Meunier JC, Russell RS, Goossens V, Priem S, Walter H, Depla E, Union A, Faulk KN, Bukh J, Emerson SU, Purcell RH. Isolation and characterization of broadly neutralizing human monoclonal antibodies to the e1 glycoprotein of hepatitis C virus. J Virol. 2008;82:966–73. doi: 10.1128/JVI.01872-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Johansson DX, Voisset C, Tarr AW, Aung M, Ball JK, Dubuisson J, Persson MA. Human combinatorial libraries yield rare antibodies that broadly neutralize hepatitis C virus. Proc Natl Acad Sci U S A. 2007;104:16269–74. doi: 10.1073/pnas.0705522104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Law M, Maruyama T, Lewis J, Giang E, Tarr AW, Stamataki Z, Gastaminza P, Chisari FV, Jones IM, Fox RI, Ball JK, McKeating JA, Kneteman NM, Burton DR. Broadly neutralizing antibodies protect against hepatitis C virus quasispecies challenge. Nat Med. 2008;14:25–7. doi: 10.1038/nm1698. [DOI] [PubMed] [Google Scholar]
- 84.Diepolder HM, Zachoval R, Hoffmann RM, Wierenga EA, Santantonio T, Jung MC, Eichenlaub D, Pape GR. Possible mechanism involving T-lymphocyte response to non-structural protein 3 in viral clearance in acute hepatitis C virus infection. Lancet. 1995;346:1006–7. doi: 10.1016/s0140-6736(95)91691-1. [DOI] [PubMed] [Google Scholar]
- 85.Day CL, Lauer GM, Robbins GK, McGovern B, Wurcel AG, Gandhi RT, Chung RT, Walker BD. Broad specificity of virus-specific CD4+ T-helper-cell responses in resolved hepatitis C virus infection. J Virol. 2002;76:12584–95. doi: 10.1128/JVI.76.24.12584-12595.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Missale G, Bertoni R, Lamonaca V, Valli A, Massari M, Mori C, Rumi MG, Houghton M, Fiaccadori F, Ferrari C. Different clinical behaviors of acute hepatitis C virus infection are associated with different vigor of the anti-viral cell-mediated immune response. J Clin Invest. 1996;98:706–14. doi: 10.1172/JCI118842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Lechner F, Wong DK, Dunbar PR, Chapman R, Chung RT, Dohrenwend P, Robbins G, Phillips R, Klenerman P, Walker BD. Analysis of successful immune responses in persons infected with hepatitis C virus. J Exp Med. 2000;191:1499–512. doi: 10.1084/jem.191.9.1499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Gruner NH, Gerlach TJ, Jung MC, Diepolder HM, Schirren CA, Schraut WW, Hoffmann R, Zachoval R, Santantonio T, Cucchiarini M, Cerny A, Pape GR. Association of hepatitis C virus-specific CD8+ T cells with viral clearance in acute hepatitis C. J Infect Dis. 2000;181:1528–36. doi: 10.1086/315450. [DOI] [PubMed] [Google Scholar]
- 89.Thimme R, Bukh J, Spangenberg HC, Wieland S, Pemberton J, Steiger C, Govindarajan S, Purcell RH, Chisari FV. Viral and immunological determinants of hepatitis C virus clearance, persistence, and disease. Proc Natl Acad Sci U S A. 2002;99:15661–8. doi: 10.1073/pnas.202608299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Bowen DG, Walker CM. Adaptive immune responses in acute and chronic hepatitis C virus infection. Nature. 2005;436:946–52. doi: 10.1038/nature04079. [DOI] [PubMed] [Google Scholar]
- 91.Gerlach JT, Diepolder HM, Jung MC, Gruener NH, Schraut WW, Zachoval R, Hoffmann R, Schirren CA, Santantonio T, Pape GR. Recurrence of hepatitis C virus after loss of virus-specific CD4(+) T-cell response in acute hepatitis C. Gastroenterology. 1999;117:933–41. doi: 10.1016/s0016-5085(99)70353-7. [DOI] [PubMed] [Google Scholar]
- 92.Kaplan DE, Sugimoto K, Newton K, Valiga ME, Ikeda F, Aytaman A, Nunes FA, Lucey MR, Vance BA, Vonderheide RH, Reddy KR, McKeating JA, Chang KM. Discordant role of CD4 T-cell response relative to neutralizing antibody and CD8 T-cell responses in acute hepatitis C. Gastroenterology. 2007;132:654–66. doi: 10.1053/j.gastro.2006.11.044. [DOI] [PubMed] [Google Scholar]
- 93.Wedemeyer H, He XS, Nascimbeni M, Davis AR, Greenberg HB, Hoofnagle JH, Liang TJ, Alter H, Rehermann B. Impaired effector function of hepatitis C virus-specific CD8+ T cells in chronic hepatitis C virus infection. J Immunol. 2002;169:3447–58. doi: 10.4049/jimmunol.169.6.3447. [DOI] [PubMed] [Google Scholar]
- 94.Koziel MJ, Dudley D, Afdhal N, Choo QL, Houghton M, Ralston R, Walker BD. Hepatitis C virus (HCV)-specific cytotoxic T lymphocytes recognize epitopes in the core and envelope proteins of HCV. J Virol. 1993;67:7522–32. doi: 10.1128/jvi.67.12.7522-7532.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Lauer GM, Barnes E, Lucas M, Timm J, Ouchi K, Kim AY, Day CL, Robbins GK, Casson DR, Reiser M, Dusheiko G, Allen TM, Chung RT, Walker BD, Klenerman P. High resolution analysis of cellular immune responses in resolved and persistent hepatitis C virus infection. Gastroenterology. 2004;127:924–36. doi: 10.1053/j.gastro.2004.06.015. [DOI] [PubMed] [Google Scholar]
- 96.Folgori A, Capone S, Ruggeri L, Meola A, Sporeno E, Ercole BB, Pezzanera M, Tafi R, Arcuri M, Fattori E, Lahm A, Luzzago A, Vitelli A, Colloca S, Cortese R, Nicosia A. A T-cell HCV vaccine eliciting effective immunity against heterologous virus challenge in chimpanzees. Nat Med. 2006;12:190–7. doi: 10.1038/nm1353. [DOI] [PubMed] [Google Scholar]
- 97.Nascimbeni M, Mizukoshi E, Bosmann M, Major ME, Mihalik K, Rice CM, Feinstone SM, Rehermann B. Kinetics of CD4+ and CD8+ memory T-cell responses during hepatitis C virus rechallenge of previously recovered chimpanzees. J Virol. 2003;77:4781–93. doi: 10.1128/JVI.77.8.4781-4793.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Grakoui A, Shoukry NH, Woollard DJ, Han JH, Hanson HL, Ghrayeb J, Murthy KK, Rice CM, Walker CM. HCV persistence and immune evasion in the absence of memory T cell help. Science. 2003;302:659–62. doi: 10.1126/science.1088774. [DOI] [PubMed] [Google Scholar]
- 99.Shoukry NH, Grakoui A, Houghton M, Chien DY, Ghrayeb J, Reimann KA, Walker CM. Memory CD8+ T cells are required for protection from persistent hepatitis C virus infection. J Exp Med. 2003;197:1645–55. doi: 10.1084/jem.20030239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Lanford RE, Guerra B, Lee H, Averett DR, Pfeiffer B, Chavez D, Notvall L, Bigger C. Antiviral effect and virus-host interactions in response to alpha interferon, gamma interferon, poly(i)-poly(c), tumor necrosis factor alpha, and ribavirin in hepatitis C virus subgenomic replicons. J Virol. 2003;77:1092–104. doi: 10.1128/JVI.77.2.1092-1104.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Napoli J, Bishop GA, McCaughan GW. Increased intrahepatic messenger RNA expression of interleukins 2, 6, and 8 in human cirrhosis. Gastroenterology. 1994;107:789–98. doi: 10.1016/0016-5085(94)90128-7. [DOI] [PubMed] [Google Scholar]
- 102.Bonilla N, Barget N, Andrieu M, Roulot D, Letoumelin P, Grando V, Trinchet JC, Ganne-Carrie N, Beaugrand M, Deny P, Choppin J, Guillet J, Ziol M. Interferon gamma-secreting HCV-specific CD8+ T cells in the liver of patients with chronic C hepatitis: relation to liver fibrosis--ANRS HC EP07 study. J Viral Hepat. 2006;13:474–81. doi: 10.1111/j.1365-2893.2005.00711.x. [DOI] [PubMed] [Google Scholar]
- 103.Shin EC, Protzer U, Untergasser A, Feinstone SM, Rice CM, Hasselschwert D, Rehermann B. Liver-directed gamma interferon gene delivery in chronic hepatitis C. J Virol. 2005;79:13412–20. doi: 10.1128/JVI.79.21.13412-13420.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Bode JG, Brenndorfer ED, Haussinger D. Subversion of innate host antiviral strategies by the hepatitis C virus. Arch Biochem Biophys. 2007;462:254–65. doi: 10.1016/j.abb.2007.03.033. [DOI] [PubMed] [Google Scholar]
- 105.Foy E, Li K, Sumpter R, Jr, Loo YM, Johnson CL, Wang C, Fish PM, Yoneyama M, Fujita T, Lemon SM, Gale M., Jr Control of antiviral defenses through hepatitis C virus disruption of retinoic acid-inducible gene-I signaling. Proc Natl Acad Sci U S A. 2005;102:2986–91. doi: 10.1073/pnas.0408707102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Foy E, Li K, Wang C, Sumpter R, Jr, Ikeda M, Lemon SM, Gale M., Jr Regulation of interferon regulatory factor-3 by the hepatitis C virus serine protease. Science. 2003;300:1145–8. doi: 10.1126/science.1082604. [DOI] [PubMed] [Google Scholar]
- 107.Vilasco M, Larrea E, Vitour D, Dabo S, Breiman A, Regnault B, Riezu JI, Eid P, Prieto J, Meurs EF. The protein kinase IKKepsilon can inhibit HCV expression independently of IFN and its own expression is downregulated in HCV-infected livers. Hepatology. 2006;44:1635–47. doi: 10.1002/hep.21432. [DOI] [PubMed] [Google Scholar]
- 108.Johnson CL, Owen DM, Gale M., Jr Functional and therapeutic analysis of hepatitis C virus NS3.4A protease control of antiviral immune defense. J Biol Chem. 2007;282:10792–803. doi: 10.1074/jbc.M610361200. [DOI] [PubMed] [Google Scholar]
- 109.Binder M, Kochs G, Bartenschlager R, Lohmann V. Hepatitis C virus escape from the interferon regulatory factor 3 pathway by a passive and active evasion strategy. Hepatology. 2007;46:1365–74. doi: 10.1002/hep.21829. [DOI] [PubMed] [Google Scholar]
- 110.Li K, Foy E, Ferreon JC, Nakamura M, Ferreon AC, Ikeda M, Ray SC, Gale M, Jr, Lemon SM. Immune evasion by hepatitis C virus NS3/4A protease-mediated cleavage of the Toll-like receptor 3 adaptor protein TRIF. Proc Natl Acad Sci U S A. 2005;102:2992–7. doi: 10.1073/pnas.0408824102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Meylan E, Curran J, Hofmann K, Moradpour D, Binder M, Bartenschlager R, Tschopp J. Cardif is an adaptor protein in the RIG-I antiviral pathway and is targeted by hepatitis C virus. Nature. 2005;437:1167–72. doi: 10.1038/nature04193. [DOI] [PubMed] [Google Scholar]
- 112.Lau DT, Fish PM, Sinha M, Owen DM, Lemon SM, Gale M., Jr Interferon regulatory factor-3 activation, hepatic interferon-stimulated gene expression, and immune cell infiltration in hepatitis C virus patients. Hepatology. 2008;47:799–809. doi: 10.1002/hep.22076. [DOI] [PubMed] [Google Scholar]
- 113.Bode JG, Ludwig S, Ehrhardt C, Albrecht U, Erhardt A, Schaper F, Heinrich PC, Haussinger D. IFN-alpha antagonistic activity of HCV core protein involves induction of suppressor of cytokine signaling-3. FASEB J. 2003;17:488–90. doi: 10.1096/fj.02-0664fje. [DOI] [PubMed] [Google Scholar]
- 114.Melen K, Fagerlund R, Nyqvist M, Keskinen P, Julkunen I. Expression of hepatitis C virus core protein inhibits interferon-induced nuclear import of STATs. J Med Virol. 2004;73:536–47. doi: 10.1002/jmv.20123. [DOI] [PubMed] [Google Scholar]
- 115.Lin W, Kim SS, Yeung E, Kamegaya Y, Blackard JT, Kim KA, Holtzman MJ, Chung RT. Hepatitis C virus core protein blocks interferon signaling by interaction with the STAT1 SH2 domain. J Virol. 2006;80:9226–35. doi: 10.1128/JVI.00459-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Kawamura H, Govindarajan S, Aswad F, Machida K, Lai MM, Sung VM, Dennert G. HCV core expression in hepatocytes protects against autoimmune liver injury and promotes liver regeneration in mice. Hepatology. 2006;44:936–44. doi: 10.1002/hep.21360. [DOI] [PubMed] [Google Scholar]
- 117.Lin W, Choe WH, Hiasa Y, Kamegaya Y, Blackard JT, Schmidt EV, Chung RT. Hepatitis C virus expression suppresses interferon signaling by degrading STAT1. Gastroenterology. 2005;128:1034–41. doi: 10.1053/j.gastro.2005.02.006. [DOI] [PubMed] [Google Scholar]
- 118.Kawaguchi T, Yoshida T, Harada M, Hisamoto T, Nagao Y, Ide T, Taniguchi E, Kumemura H, Hanada S, Maeyama M, Baba S, Koga H, Kumashiro R, Ueno T, Ogata H, Yoshimura A, Sata M. Hepatitis C virus down-regulates insulin receptor substrates 1 and 2 through up-regulation of suppressor of cytokine signaling 3. Am J Pathol. 2004;165:1499–508. doi: 10.1016/S0002-9440(10)63408-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Yoshida T, Hanada T, Tokuhisa T, Kosai K, Sata M, Kohara M, Yoshimura A. Activation of STAT3 by the hepatitis C virus core protein leads to cellular transformation. J Exp Med. 2002;196:641–53. doi: 10.1084/jem.20012127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.de Lucas S, Bartolome J, Carreno V. Hepatitis C virus core protein down-regulates transcription of interferon-induced antiviral genes. J Infect Dis. 2005;191:93–9. doi: 10.1086/426509. [DOI] [PubMed] [Google Scholar]
- 121.Gale MJ, Jr, Korth MJ, Tang NM, Tan SL, Hopkins DA, Dever TE, Polyak SJ, Gretch DR, Katze MG. Evidence that hepatitis C virus resistance to interferon is mediated through repression of the PKR protein kinase by the nonstructural 5A protein. Virology. 1997;230:217–27. doi: 10.1006/viro.1997.8493. [DOI] [PubMed] [Google Scholar]
- 122.Gale M, Jr, Blakely CM, Kwieciszewski B, Tan SL, Dossett M, Tang NM, Korth MJ, Polyak SJ, Gretch DR, Katze MG. Control of PKR protein kinase by hepatitis C virus nonstructural 5A protein: molecular mechanisms of kinase regulation. Mol Cell Biol. 1998;18:5208–18. doi: 10.1128/mcb.18.9.5208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Pflugheber J, Fredericksen B, Sumpter R, Jr, Wang C, Ware F, Sodora DL, Gale M., Jr Regulation of PKR and IRF-1 during hepatitis C virus RNA replication. Proc Natl Acad Sci U S A. 2002;99:4650–5. doi: 10.1073/pnas.062055699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Taguchi T, Nagano-Fujii M, Akutsu M, Kadoya H, Ohgimoto S, Ishido S, Hotta H. Hepatitis C virus NS5A protein interacts with 2′,5′-oligoadenylate synthetase and inhibits antiviral activity of IFN in an IFN sensitivity-determining region-independent manner. J Gen Virol. 2004;85:959–69. doi: 10.1099/vir.0.19513-0. [DOI] [PubMed] [Google Scholar]
- 125.Polyak SJ, Khabar KS, Paschal DM, Ezelle HJ, Duverlie G, Barber GN, Levy DE, Mukaida N, Gretch DR. Hepatitis C virus nonstructural 5A protein induces interleukin-8, leading to partial inhibition of the interferon-induced antiviral response. J Virol. 2001;75:6095–106. doi: 10.1128/JVI.75.13.6095-6106.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Polyak SJ, Khabar KS, Rezeiq M, Gretch DR. Elevated levels of interleukin-8 in serum are associated with hepatitis C virus infection and resistance to interferon therapy. J Virol. 2001;75:6209–11. doi: 10.1128/JVI.75.13.6209-6211.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Khabar KS, Al-Zoghaibi F, Al-Ahdal MN, Murayama T, Dhalla M, Mukaida N, Taha M, Al-Sedairy ST, Siddiqui Y, Kessie G, Matsushima K. The alpha chemokine, interleukin 8, inhibits the antiviral action of interferon alpha. J Exp Med. 1997;186:1077–85. doi: 10.1084/jem.186.7.1077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Taylor DR, Shi ST, Romano PR, Barber GN, Lai MM. Inhibition of the interferon-inducible protein kinase PKR by HCV E2 protein. Science. 1999;285:107–10. doi: 10.1126/science.285.5424.107. [DOI] [PubMed] [Google Scholar]
- 129.Fuller MJ, Hildeman DA, Sabbaj S, Gaddis DE, Tebo AE, Shang L, Goepfert PA, Zajac AJ. Cutting edge: emergence of CD127high functionally competent memory T cells is compromised by high viral loads and inadequate T cell help. J Immunol. 2005;174:5926–30. doi: 10.4049/jimmunol.174.10.5926. [DOI] [PubMed] [Google Scholar]
- 130.Gruener NH, Lechner F, Jung MC, Diepolder H, Gerlach T, Lauer G, Walker B, Sullivan J, Phillips R, Pape GR, Klenerman P. Sustained dysfunction of antiviral CD8+ T lymphocytes after infection with hepatitis C virus. J Virol. 2001;75:5550–8. doi: 10.1128/JVI.75.12.5550-5558.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Urbani S, Boni C, Missale G, Elia G, Cavallo C, Massari M, Raimondo G, Ferrari C. Virus-specific CD8+ lymphocytes share the same effector-memory phenotype but exhibit functional differences in acute hepatitis B and C. J Virol. 2002;76:12423–34. doi: 10.1128/JVI.76.24.12423-12434.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Appay V, Dunbar PR, Callan M, Klenerman P, Gillespie GM, Papagno L, Ogg GS, King A, Lechner F, Spina CA, Little S, Havlir DV, Richman DD, Gruener N, Pape G, Waters A, Easterbrook P, Salio M, Cerundolo V, McMichael AJ, Rowland-Jones SL. Memory CD8+ T cells vary in differentiation phenotype in different persistent virus infections. Nat Med. 2002;8:379–85. doi: 10.1038/nm0402-379. [DOI] [PubMed] [Google Scholar]
- 133.Spangenberg HC, Viazov S, Kersting N, Neumann-Haefelin C, McKinney D, Roggendorf M, von Weizsacker F, Blum HE, Thimme R. Intrahepatic CD8+ T-cell failure during chronic hepatitis C virus infection. Hepatology. 2005;42:828–37. doi: 10.1002/hep.20856. [DOI] [PubMed] [Google Scholar]
- 134.Koziel MJ, Dudley D, Afdhal N, Grakoui A, Rice CM, Choo QL, Houghton M, Walker BD. HLA class I-restricted cytotoxic T lymphocytes specific for hepatitis C virus. Identification of multiple epitopes and characterization of patterns of cytokine release. J Clin Invest. 1995;96:2311–21. doi: 10.1172/JCI118287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.He XS, Rehermann B, Lopez-Labrador FX, Boisvert J, Cheung R, Mumm J, Wedemeyer H, Berenguer M, Wright TL, Davis MM, Greenberg HB. Quantitative analysis of hepatitis C virus-specific CD8(+) T cells in peripheral blood and liver using peptide-MHC tetramers. Proc Natl Acad Sci U S A. 1999;96:5692–7. doi: 10.1073/pnas.96.10.5692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Cooper S, Erickson AL, Adams EJ, Kansopon J, Weiner AJ, Chien DY, Houghton M, Parham P, Walker CM. Analysis of a successful immune response against hepatitis C virus. Immunity. 1999;10:439–49. doi: 10.1016/s1074-7613(00)80044-8. [DOI] [PubMed] [Google Scholar]
- 137.Lauer GM, Ouchi K, Chung RT, Nguyen TN, Day CL, Purkis DR, Reiser M, Kim AY, Lucas M, Klenerman P, Walker BD. Comprehensive analysis of CD8(+)-T-cell responses against hepatitis C virus reveals multiple unpredicted specificities. J Virol. 2002;76:6104–13. doi: 10.1128/JVI.76.12.6104-6113.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Radziewicz H, Ibegbu CC, Fernandez ML, Workowski KA, Obideen K, Wehbi M, Hanson HL, Steinberg JP, Masopust D, Wherry EJ, Altman JD, Rouse BT, Freeman GJ, Ahmed R, Grakoui A. Liver-infiltrating lymphocytes in chronic human hepatitis C virus infection display an exhausted phenotype with high levels of PD-1 and low levels of CD127 expression. J Virol. 2007;81:2545–53. doi: 10.1128/JVI.02021-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Penna A, Pilli M, Zerbini A, Orlandini A, Mezzadri S, Sacchelli L, Missale G, Ferrari C. Dysfunction and functional restoration of HCV-specific CD8 responses in chronic hepatitis C virus infection. Hepatology. 2007;45:588–601. doi: 10.1002/hep.21541. [DOI] [PubMed] [Google Scholar]
- 140.Golden-Mason L, Palmer B, Klarquist J, Mengshol JA, Castelblanco N, Rosen HR. Upregulation of PD-1 expression on circulating and intrahepatic hepatitis C virus-specific CD8+ T cells associated with reversible immune dysfunction. J Virol. 2007;81:9249–58. doi: 10.1128/JVI.00409-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Kasprowicz V, Schulze Zur Wiesch J, Kuntzen T, Nolan BE, Longworth S, Berical A, Blum J, McMahon C, Reyor LL, Elias N, Kwok WW, McGovern BG, Freeman G, Chung RT, Klenerman P, Lewis-Ximenez L, Walker BD, Allen TM, Kim AY, Lauer GM. High level of PD-1 expression on hepatitis C virus (HCV)-specific CD8+ and CD4+ T cells during acute HCV infection, irrespective of clinical outcome. J Virol. 2008;82:3154–60. doi: 10.1128/JVI.02474-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Neumann AU, Lam NP, Dahari H, Gretch DR, Wiley TE, Layden TJ, Perelson AS. Hepatitis C viral dynamics in vivo and the antiviral efficacy of interferon-alpha therapy. Science. 1998;282:103–7. doi: 10.1126/science.282.5386.103. [DOI] [PubMed] [Google Scholar]
- 143.Bukh J, Miller RH, Purcell RH. Genetic heterogeneity of hepatitis C virus: quasispecies and genotypes. Semin Liver Dis. 1995;15:41–63. doi: 10.1055/s-2007-1007262. [DOI] [PubMed] [Google Scholar]
- 144.Bowen DG, Walker CM. Mutational escape from CD8+ T cell immunity: HCV evolution, from chimpanzees to man. J Exp Med. 2005;201:1709–14. doi: 10.1084/jem.20050808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Erickson AL, Kimura Y, Igarashi S, Eichelberger J, Houghton M, Sidney J, McKinney D, Sette A, Hughes AL, Walker CM. The outcome of hepatitis C virus infection is predicted by escape mutations in epitopes targeted by cytotoxic T lymphocytes. Immunity. 2001;15:883–95. doi: 10.1016/s1074-7613(01)00245-x. [DOI] [PubMed] [Google Scholar]
- 146.Cox AL, Mosbruger T, Mao Q, Liu Z, Wang XH, Yang HC, Sidney J, Sette A, Pardoll D, Thomas DL, Ray SC. Cellular immune selection with hepatitis C virus persistence in humans. J Exp Med. 2005;201:1741–52. doi: 10.1084/jem.20050121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Timm J, Lauer GM, Kavanagh DG, Sheridan I, Kim AY, Lucas M, Pillay T, Ouchi K, Reyor LL, Schulze zur Wiesch J, Gandhi RT, Chung RT, Bhardwaj N, Klenerman P, Walker BD, Allen TM. CD8 epitope escape and reversion in acute HCV infection. J Exp Med. 2004;200:1593–604. doi: 10.1084/jem.20041006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Ray SC, Fanning L, Wang XH, Netski DM, Kenny-Walsh E, Thomas DL. Divergent and convergent evolution after a common-source outbreak of hepatitis C virus. J Exp Med. 2005;201:1753–9. doi: 10.1084/jem.20050122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Tester I, Smyk-Pearson S, Wang P, Wertheimer A, Yao E, Lewinsohn DM, Tavis JE, Rosen HR. Immune evasion versus recovery after acute hepatitis C virus infection from a shared source. J Exp Med. 2005;201:1725–31. doi: 10.1084/jem.20042284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Neumann-Haefelin C, Frick DN, Wang JJ, Pybus OG, Salloum S, Narula GS, Eckart A, Biezynski A, Eiermann T, Klenerman P, Viazov S, Roggendorf M, Thimme R, Reiser M, Timm J. Analysis of the evolutionary forces in an immunodominant CD8 epitope in the hepatitis C virus at a population level. J Virol. 2008 doi: 10.1128/JVI.01700-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Meyer-Olson D, Shoukry NH, Brady KW, Kim H, Olson DP, Hartman K, Shintani AK, Walker CM, Kalams SA. Limited T cell receptor diversity of HCV-specific T cell responses is associated with CTL escape. J Exp Med. 2004;200:307–19. doi: 10.1084/jem.20040638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Seifert U, Liermann H, Racanelli V, Halenius A, Wiese M, Wedemeyer H, Ruppert T, Rispeter K, Henklein P, Sijts A, Hengel H, Kloetzel PM, Rehermann B. Hepatitis C virus mutation affects proteasomal epitope processing. J Clin Invest. 2004;114:250–9. doi: 10.1172/JCI20985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Puig M, Mihalik K, Tilton JC, Williams O, Merchlinsky M, Connors M, Feinstone SM, Major ME. CD4+ immune escape and subsequent T-cell failure following chimpanzee immunization against hepatitis C virus. Hepatology. 2006;44:736–45. doi: 10.1002/hep.21319. [DOI] [PubMed] [Google Scholar]
- 154.Sakaguchi S. Naturally arising CD4+ regulatory t cells for immunologic self-tolerance and negative control of immune responses. Annu Rev Immunol. 2004;22:531–62. doi: 10.1146/annurev.immunol.21.120601.141122. [DOI] [PubMed] [Google Scholar]
- 155.Nelson BH. IL-2, regulatory T cells, and tolerance. J Immunol. 2004;172:3983–8. doi: 10.4049/jimmunol.172.7.3983. [DOI] [PubMed] [Google Scholar]
- 156.Accapezzato D, Francavilla V, Paroli M, Casciaro M, Chircu LV, Cividini A, Abrignani S, Mondelli MU, Barnaba V. Hepatic expansion of a virus-specific regulatory CD8(+) T cell population in chronic hepatitis C virus infection. J Clin Invest. 2004;113:963–72. doi: 10.1172/JCI20515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Shevach EM. From vanilla to 28 flavors: multiple varieties of T regulatory cells. Immunity. 2006;25:195–201. doi: 10.1016/j.immuni.2006.08.003. [DOI] [PubMed] [Google Scholar]
- 158.Shevach EM, DiPaolo RA, Andersson J, Zhao DM, Stephens GL, Thornton AM. The lifestyle of naturally occurring CD4+ CD25+ Foxp3+ regulatory T cells. Immunol Rev. 2006;212:60–73. doi: 10.1111/j.0105-2896.2006.00415.x. [DOI] [PubMed] [Google Scholar]
- 159.Cabrera R, Tu Z, Xu Y, Firpi RJ, Rosen HR, Liu C, Nelson DR. An immunomodulatory role for CD4(+)CD25(+) regulatory T lymphocytes in hepatitis C virus infection. Hepatology. 2004;40:1062–71. doi: 10.1002/hep.20454. [DOI] [PubMed] [Google Scholar]
- 160.Sugimoto K, Ikeda F, Stadanlick J, Nunes FA, Alter HJ, Chang KM. Suppression of HCV-specific T cells without differential hierarchy demonstrated ex vivo in persistent HCV infection. Hepatology. 2003;38:1437–48. doi: 10.1016/j.hep.2003.09.026. [DOI] [PubMed] [Google Scholar]
- 161.Boettler T, Spangenberg HC, Neumann-Haefelin C, Panther E, Urbani S, Ferrari C, Blum HE, von Weizsacker F, Thimme R. T cells with a CD4+CD25+ regulatory phenotype suppress in vitro proliferation of virus-specific CD8+ T cells during chronic hepatitis C virus infection. J Virol. 2005;79:7860–7. doi: 10.1128/JVI.79.12.7860-7867.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Rushbrook SM, Ward SM, Unitt E, Vowler SL, Lucas M, Klenerman P, Alexander GJ. Regulatory T cells suppress in vitro proliferation of virus-specific CD8+ T cells during persistent hepatitis C virus infection. J Virol. 2005;79:7852–9. doi: 10.1128/JVI.79.12.7852-7859.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Manigold T, Shin EC, Mizukoshi E, Mihalik K, Murthy KK, Rice CM, Piccirillo CA, Rehermann B. Foxp3+CD4+CD25+ T cells control virus-specific memory T cells in chimpanzees that recovered from hepatitis C. Blood. 2006;107:4424–32. doi: 10.1182/blood-2005-09-3903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Koziel MJ, Dudley D, Wong JT, Dienstag J, Houghton M, Ralston R, Walker BD. Intrahepatic cytotoxic T lymphocytes specific for hepatitis C virus in persons with chronic hepatitis. J Immunol. 1992;149:3339–44. [PubMed] [Google Scholar]
- 165.Bertoletti A, D'Elios MM, Boni C, De Carli M, Zignego AL, Durazzo M, Missale G, Penna A, Fiaccadori F, Del Prete G, Ferrari C. Different cytokine profiles of intraphepatic T cells in chronic hepatitis B and hepatitis C virus infections. Gastroenterology. 1997;112:193–9. doi: 10.1016/s0016-5085(97)70235-x. [DOI] [PubMed] [Google Scholar]
- 166.Nelson DR, Marousis CG, Davis GL, Rice CM, Wong J, Houghton M, Lau JY. The role of hepatitis C virus-specific cytotoxic T lymphocytes in chronic hepatitis C. J Immunol. 1997;158:1473–81. [PubMed] [Google Scholar]
- 167.Schirren CA, Jung MC, Gerlach JT, Worzfeld T, Baretton G, Mamin M, Hubert Gruener N, Houghton M, Pape GR. Liver-derived hepatitis C virus (HCV)-specific CD4(+) T cells recognize multiple HCV epitopes and produce interferon gamma. Hepatology. 2000;32:597–603. doi: 10.1053/jhep.2000.9635. [DOI] [PubMed] [Google Scholar]
- 168.Grabowska AM, Lechner F, Klenerman P, Tighe PJ, Ryder S, Ball JK, Thomson BJ, Irving WL, Robins RA. Direct ex vivo comparison of the breadth and specificity of the T cells in the liver and peripheral blood of patients with chronic HCV infection. Eur J Immunol. 2001;31:2388–94. doi: 10.1002/1521-4141(200108)31:8<2388::aid-immu2388>3.0.co;2-l. [DOI] [PubMed] [Google Scholar]
- 169.Penna A, Missale G, Lamonaca V, Pilli M, Mori C, Zanelli P, Cavalli A, Elia G, Ferrari C. Intrahepatic and circulating HLA class II-restricted, hepatitis C virus-specific T cells: functional characterization in patients with chronic hepatitis C. Hepatology. 2002;35:1225–36. doi: 10.1053/jhep.2002.33153. [DOI] [PubMed] [Google Scholar]
- 170.Leroy V, Vigan I, Mosnier JF, Dufeu-Duchesne T, Pernollet M, Zarski JP, Marche PN, Jouvin-Marche E. Phenotypic and functional characterization of intrahepatic T lymphocytes during chronic hepatitis C. Hepatology. 2003;38:829–41. doi: 10.1053/jhep.2003.50410. [DOI] [PubMed] [Google Scholar]
- 171.Kamal SM, Graham CS, He Q, Bianchi L, Tawil AA, Rasenack JW, Khalifa KA, Massoud MM, Koziel MJ. Kinetics of intrahepatic hepatitis C virus (HCV)-specific CD4+ T cell responses in HCV and Schistosoma mansoni coinfection: relation to progression of liver fibrosis. J Infect Dis. 2004;189:1140–50. doi: 10.1086/382278. [DOI] [PubMed] [Google Scholar]
- 172.Lalor PF, Shields P, Grant A, Adams DH. Recruitment of lymphocytes to the human liver. Immunol Cell Biol. 2002;80:52–64. doi: 10.1046/j.1440-1711.2002.01062.x. [DOI] [PubMed] [Google Scholar]
- 173.Shields PL, Morland CM, Salmon M, Qin S, Hubscher SG, Adams DH. Chemokine and chemokine receptor interactions provide a mechanism for selective T cell recruitment to specific liver compartments within hepatitis C-infected liver. J Immunol. 1999;163:6236–43. [PubMed] [Google Scholar]
- 174.Helbig KJ, Ruszkiewicz A, Semendric L, Harley HA, McColl SR, Beard MR. Expression of the CXCR3 ligand I-TAC by hepatocytes in chronic hepatitis C and its correlation with hepatic inflammation. Hepatology. 2004;39:1220–9. doi: 10.1002/hep.20167. [DOI] [PubMed] [Google Scholar]
- 175.Harvey CE, Post JJ, Palladinetti P, Freeman AJ, Ffrench RA, Kumar RK, Marinos G, Lloyd AR. Expression of the chemokine IP-10 (CXCL10) by hepatocytes in chronic hepatitis C virus infection correlates with histological severity and lobular inflammation. J Leukoc Biol. 2003;74:360–9. doi: 10.1189/jlb.0303093. [DOI] [PubMed] [Google Scholar]
- 176.Honda A, Arai Y, Hirota N, Sato T, Ikegaki J, Koizumi T, Hatano M, Kohara M, Moriyama T, Imawari M, Shimotohno K, Tokuhisa T. Hepatitis C virus structural proteins induce liver cell injury in transgenic mice. J Med Virol. 1999;59:281–9. doi: 10.1002/(sici)1096-9071(199911)59:3<281::aid-jmv4>3.0.co;2-s. [DOI] [PubMed] [Google Scholar]
- 177.Alonzi T, Agrati C, Costabile B, Cicchini C, Amicone L, Cavallari C, Rocca CD, Folgori A, Fipaldini C, Poccia F, Monica NL, Tripodi M. Steatosis and intrahepatic lymphocyte recruitment in hepatitis C virus transgenic mice. J Gen Virol. 2004;85:1509–20. doi: 10.1099/vir.0.19724-0. [DOI] [PubMed] [Google Scholar]
- 178.Charlton M. Recurrence of hepatitis C infection: Where are we now? Liver Transpl. 2005;11:S57–62. doi: 10.1002/lt.20597. [DOI] [PubMed] [Google Scholar]
- 179.Koziel MJ, Peters MG. Viral hepatitis in HIV infection. N Engl J Med. 2007;356:1445–54. doi: 10.1056/NEJMra065142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Kamal SM, Rasenack JW, Bianchi L, Al Tawil A, El Sayed Khalifa K, Peter T, Mansour H, Ezzat W, Koziel M. Acute hepatitis C without and with schistosomiasis: correlation with hepatitis C-specific CD4(+) T-cell and cytokine response. Gastroenterology. 2001;121:646–56. doi: 10.1053/gast.2001.27024. [DOI] [PubMed] [Google Scholar]
- 181.Rosen HR, Miner C, Sasaki AW, Lewinsohn DM, Conrad AJ, Bakke A, Bouwer HG, Hinrichs DJ. Frequencies of HCV-specific effector CD4+ T cells by flow cytometry: correlation with clinical disease stages. Hepatology. 2002;35:190–8. doi: 10.1053/jhep.2002.30293. [DOI] [PubMed] [Google Scholar]
- 182.Ward SM, Fox BC, Brown PJ, Worthington J, Fox SB, Chapman RW, Fleming KA, Banham AH, Klenerman P. Quantification and localisation of FOXP3+ T lymphocytes and relation to hepatic inflammation during chronic HCV infection. J Hepatol. 2007;47:316–24. doi: 10.1016/j.jhep.2007.03.023. [DOI] [PubMed] [Google Scholar]
- 183.Wakita T, Katsume A, Kato J, Taya C, Yonekawa H, Kanegae Y, Saito I, Hayashi Y, Koike M, Miyamoto M, Hiasa Y, Kohara M. Possible role of cytotoxic T cells in acute liver injury in hepatitis C virus cDNA transgenic mice mediated by Cre/loxP system. J Med Virol. 2000;62:308–17. doi: 10.1002/1096-9071(200011)62:3<308::aid-jmv2>3.0.co;2-6. [DOI] [PubMed] [Google Scholar]
- 184.Takaku S, Nakagawa Y, Shimizu M, Norose Y, Maruyama I, Wakita T, Takano T, Kohara M, Takahashi H. Induction of hepatic injury by hepatitis C virus-specific CD8+ murine cytotoxic T lymphocytes in transgenic mice expressing the viral structural genes. Biochem Biophys Res Commun. 2003;301:330–7. doi: 10.1016/s0006-291x(02)03018-8. [DOI] [PubMed] [Google Scholar]
- 185.Hiroishi K, Kita H, Kojima M, Okamoto H, Moriyama T, Kaneko T, Ishikawa T, Ohnishi S, Aikawa T, Tanaka N, Yazaki Y, Mitamura K, Imawari M. Cytotoxic T lymphocyte response and viral load in hepatitis C virus infection. Hepatology. 1997;25:705–12. doi: 10.1002/hep.510250336. [DOI] [PubMed] [Google Scholar]
- 186.Freeman AJ, Pan Y, Harvey CE, Post JJ, Law MG, White PA, Rawlinson WD, Lloyd AR, Marinos G, Ffrench RA. The presence of an intrahepatic cytotoxic T lymphocyte response is associated with low viral load in patients with chronic hepatitis C virus infection. J Hepatol. 2003;38:349–56. doi: 10.1016/s0168-8278(02)00424-5. [DOI] [PubMed] [Google Scholar]
- 187.Rehermann B, Chang KM, McHutchison JG, Kokka R, Houghton M, Chisari FV. Quantitative analysis of the peripheral blood cytotoxic T lymphocyte response in patients with chronic hepatitis C virus infection. J Clin Invest. 1996;98:1432–40. doi: 10.1172/JCI118931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Rehermann B, Chang KM, McHutchinson J, Kokka R, Houghton M, Rice CM, Chisari FV. Differential cytotoxic T-lymphocyte responsiveness to the hepatitis B and C viruses in chronically infected patients. J Virol. 1996;70:7092–102. doi: 10.1128/jvi.70.10.7092-7102.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Wong DK, Dudley DD, Afdhal NH, Dienstag J, Rice CM, Wang L, Houghton M, Walker BD, Koziel MJ. Liver-derived CTL in hepatitis C virus infection: breadth and specificity of responses in a cohort of persons with chronic infection. J Immunol. 1998;160:1479–88. [PubMed] [Google Scholar]
- 190.Rothman AL, Morishima C, Bonkovsky HL, Polyak SJ, Ray R, Di Bisceglie AM, Lindsay KL, Malet PF, Chang M, Gretch DR, Sullivan DG, Bhan AK, Wright EC, Koziel MJ. Associations among clinical, immunological, and viral quasispecies measurements in advanced chronic hepatitis C. Hepatology. 2005;41:617–25. doi: 10.1002/hep.20581. [DOI] [PubMed] [Google Scholar]
- 191.Abrignani S. Antigen-independent activation of resting T-cells in the liver of patients with chronic hepatitis. Dev Biol Stand. 1998;92:191–4. [PubMed] [Google Scholar]
- 192.Abel M, Sene D, Pol S, Bourliere M, Poynard T, Charlotte F, Cacoub P, Caillat-Zucman S. Intrahepatic virus-specific IL-10-producing CD8 T cells prevent liver damage during chronic hepatitis C virus infection. Hepatology. 2006;44:1607–16. doi: 10.1002/hep.21438. [DOI] [PubMed] [Google Scholar]
- 193.Bonorino P, Leroy V, Dufeu-Duchesne T, Tongiani-Dashan S, Sturm N, Pernollet M, Vivier E, Zarski JP, Marche PN, Jouvin-Marche E. Features and distribution of CD8 T cells with human leukocyte antigen class I-specific receptor expression in chronic hepatitis C. Hepatology. 2007;46:1375–86. doi: 10.1002/hep.21850. [DOI] [PubMed] [Google Scholar]
- 194.Alatrakchi N, Graham CS, van der Vliet HJ, Sherman KE, Exley MA, Koziel MJ. Hepatitis C virus (HCV)-specific CD8+ cells produce transforming growth factor beta that can suppress HCV-specific T-cell responses. J Virol. 2007;81:5882–92. doi: 10.1128/JVI.02202-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Di Bisceglie AM, Hoofnagle JH. Optimal therapy of hepatitis C. Hepatology. 2002;36:S121–7. doi: 10.1053/jhep.2002.36228. [DOI] [PubMed] [Google Scholar]
- 196.Poynard T, Marcellin P, Lee SS, Niederau C, Minuk GS, Ideo G, Bain V, Heathcote J, Zeuzem S, Trepo C, Albrecht J. Randomised trial of interferon alpha2b plus ribavirin for 48 weeks or for 24 weeks versus interferon alpha2b plus placebo for 48 weeks for treatment of chronic infection with hepatitis C virus. International Hepatitis Interventional Therapy Group (IHIT) Lancet. 1998;352:1426–32. doi: 10.1016/s0140-6736(98)07124-4. [DOI] [PubMed] [Google Scholar]
- 197.Feld JJ, Hoofnagle JH. Mechanism of action of interferon and ribavirin in treatment of hepatitis C. Nature. 2005;436:967–72. doi: 10.1038/nature04082. [DOI] [PubMed] [Google Scholar]
- 198.Manns MP, McHutchison JG, Gordon SC, Rustgi VK, Shiffman M, Reindollar R, Goodman ZD, Koury K, Ling M, Albrecht JK. Peginterferon alfa-2b plus ribavirin compared with interferon alfa-2b plus ribavirin for initial treatment of chronic hepatitis C: a randomised trial. Lancet. 2001;358:958–65. doi: 10.1016/s0140-6736(01)06102-5. [DOI] [PubMed] [Google Scholar]
- 199.Fried MW, Shiffman ML, Reddy KR, Smith C, Marinos G, Goncales FL, Jr, Haussinger D, Diago M, Carosi G, Dhumeaux D, Craxi A, Lin A, Hoffman J, Yu J. Peginterferon alfa-2a plus ribavirin for chronic hepatitis C virus infection. N Engl J Med. 2002;347:975–82. doi: 10.1056/NEJMoa020047. [DOI] [PubMed] [Google Scholar]
- 200.Hadziyannis SJ, Sette H, Jr, Morgan TR, Balan V, Diago M, Marcellin P, Ramadori G, Bodenheimer H, Jr, Bernstein D, Rizzetto M, Zeuzem S, Pockros PJ, Lin A, Ackrill AM. Peginterferon-alpha2a and ribavirin combination therapy in chronic hepatitis C: a randomized study of treatment duration and ribavirin dose. Ann Intern Med. 2004;140:346–55. doi: 10.7326/0003-4819-140-5-200403020-00010. [DOI] [PubMed] [Google Scholar]
- 201.Zeuzem S, Hultcrantz R, Bourliere M, Goeser T, Marcellin P, Sanchez-Tapias J, Sarrazin C, Harvey J, Brass C, Albrecht J. Peginterferon alfa-2b plus ribavirin for treatment of chronic hepatitis C in previously untreated patients infected with HCV genotypes 2 or 3. J Hepatol. 2004;40:993–9. doi: 10.1016/j.jhep.2004.02.007. [DOI] [PubMed] [Google Scholar]
- 202.Muir AJ, Bornstein JD, Killenberg PG. Peginterferon alfa-2b and ribavirin for the treatment of chronic hepatitis C in blacks and non-Hispanic whites. N Engl J Med. 2004;350:2265–71. doi: 10.1056/NEJMoa032502. [DOI] [PubMed] [Google Scholar]
- 203.Conjeevaram HS, Fried MW, Jeffers LJ, Terrault NA, Wiley-Lucas TE, Afdhal N, Brown RS, Belle SH, Hoofnagle JH, Kleiner DE, Howell CD. Peginterferon and ribavirin treatment in African American and Caucasian American patients with hepatitis C genotype 1. Gastroenterology. 2006;131:470–7. doi: 10.1053/j.gastro.2006.06.008. [DOI] [PubMed] [Google Scholar]
- 204.Sugimoto K, Stadanlick J, Ikeda F, Brensinger C, Furth EE, Alter HJ, Chang KM. Influence of ethnicity in the outcome of hepatitis C virus infection and cellular immune response. Hepatology. 2003;37:590–9. doi: 10.1053/jhep.2003.50103. [DOI] [PubMed] [Google Scholar]
- 205.Rosen HR, Weston SJ, Im K, Yang H, Burton JR, Jr, Erlich H, Klarquist J, Belle SH. Selective decrease in hepatitis C virus-specific immunity among African Americans and outcome of antiviral therapy. Hepatology. 2007;46:350–8. doi: 10.1002/hep.21714. [DOI] [PubMed] [Google Scholar]
- 206.Brinkmann V, Geiger T, Alkan S, Heusser CH. Interferon alpha increases the frequency of interferon gamma-producing human CD4+ T cells. J Exp Med. 1993;178:1655–63. doi: 10.1084/jem.178.5.1655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Luft T, Pang KC, Thomas E, Hertzog P, Hart DN, Trapani J, Cebon J. Type I IFNs enhance the terminal differentiation of dendritic cells. J Immunol. 1998;161:1947–53. [PubMed] [Google Scholar]
- 208.Tam RC, Pai B, Bard J, Lim C, Averett DR, Phan UT, Milovanovic T. Ribavirin polarizes human T cell responses towards a Type 1 cytokine profile. J Hepatol. 1999;30:376–82. doi: 10.1016/s0168-8278(99)80093-2. [DOI] [PubMed] [Google Scholar]
- 209.Cramp ME, Rossol S, Chokshi S, Carucci P, Williams R, Naoumov NV. Hepatitis C virus-specific T-cell reactivity during interferon and ribavirin treatment in chronic hepatitis C. Gastroenterology. 2000;118:346–55. doi: 10.1016/s0016-5085(00)70217-4. [DOI] [PubMed] [Google Scholar]
- 210.Pawlotsky JM. Therapy of hepatitis C: from empiricism to eradication. Hepatology. 2006;43:S207–20. doi: 10.1002/hep.21064. [DOI] [PubMed] [Google Scholar]
- 211.Pawlotsky JM, Gish RG. Future therapies for hepatitis C. Antivir Ther. 2006;11:397–408. [PubMed] [Google Scholar]
- 212.Pilli M, Zerbini A, Penna A, Orlandini A, Lukasiewicz E, Pawlotsky JM, Zeuzem S, Schalm SW, von Wagner M, Germanidis G, Lurie Y, Esteban JI, Haagmans BL, Hezode C, Lagging M, Negro F, Homburger Y, Neumann AU, Ferrari C, Missale G. HCV-specific T-cell response in relation to viral kinetics and treatment outcome (DITTO-HCV project) Gastroenterology. 2007;133:1132–43. doi: 10.1053/j.gastro.2007.06.059. [DOI] [PubMed] [Google Scholar]
- 213.Graham CS, Wells A, Liu T, Sherman KE, Peters M, Chung RT, Bhan AK, Andersen J, Koziel MJ. Relationships between cellular immune responses and treatment outcomes with interferon and ribavirin in HIV/hepatitis C virus co-infection. AIDS. 2006;20:345–51. doi: 10.1097/01.aids.0000206500.16783.2e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Kamal SM, Ismail A, Graham CS, He Q, Rasenack JW, Peters T, Tawil AA, Fehr JJ, Khalifa Kel S, Madwar MM, Koziel MJ. Pegylated interferon alpha therapy in acute hepatitis C: relation to hepatitis C virus-specific T cell response kinetics. Hepatology. 2004;39:1721–31. doi: 10.1002/hep.20266. [DOI] [PubMed] [Google Scholar]
- 215.Tang KH, Herrmann E, Cooksley H, Tatman N, Chokshi S, Williams R, Zeuzem S, Naoumov NV. Relationship between early HCV kinetics and T-cell reactivity in chronic hepatitis C genotype 1 during peginterferon and ribavirin therapy. J Hepatol. 2005;43:776–82. doi: 10.1016/j.jhep.2005.05.024. [DOI] [PubMed] [Google Scholar]
- 216.Randall G, Chen L, Panis M, Fischer AK, Lindenbach BD, Sun J, Heathcote J, Rice CM, Edwards AM, McGilvray ID. Silencing of USP18 potentiates the antiviral activity of interferon against hepatitis C virus infection. Gastroenterology. 2006;131:1584–91. doi: 10.1053/j.gastro.2006.08.043. [DOI] [PubMed] [Google Scholar]
- 217.Chen L, Borozan I, Feld J, Sun J, Tannis LL, Coltescu C, Heathcote J, Edwards AM, McGilvray ID. Hepatic gene expression discriminates responders and nonresponders in treatment of chronic hepatitis C viral infection. Gastroenterology. 2005;128:1437–44. doi: 10.1053/j.gastro.2005.01.059. [DOI] [PubMed] [Google Scholar]
- 218.Miyoshi H, Fujie H, Shintani Y, Tsutsumi T, Shinzawa S, Makuuchi M, Kokudo N, Matsuura Y, Suzuki T, Miyamura T, Moriya K, Koike K. Hepatitis C virus core protein exerts an inhibitory effect on suppressor of cytokine signaling (SOCS)-1 gene expression. J Hepatol. 2005;43:757–63. doi: 10.1016/j.jhep.2005.03.028. [DOI] [PubMed] [Google Scholar]