

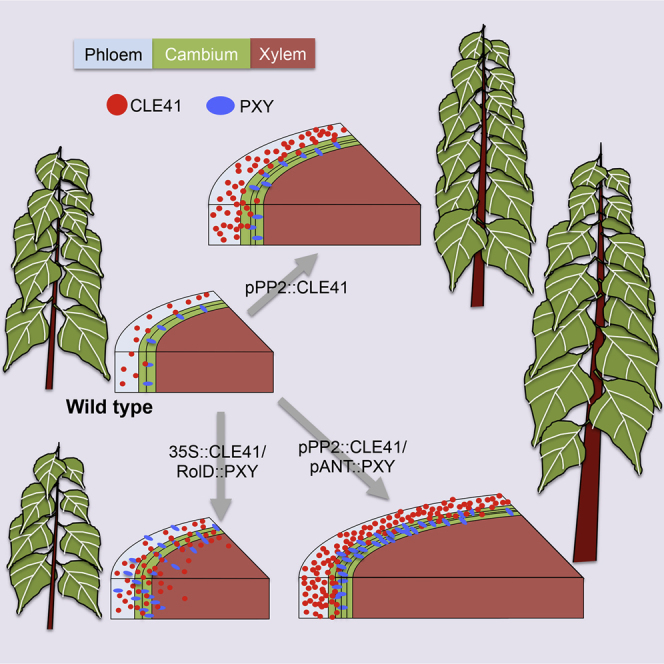
Summary
The woody tissue of trees is composed of xylem cells that arise from divisions of stem cells within the cambial meristem. The rate of xylem cell formation is dependent upon the rate of cell division within the cambium and is controlled by both genetic and environmental factors [1, 2]. In the annual plant Arabidopsis, signaling between a peptide ligand CLE41 and a receptor kinase PXY controls cambial cell divisions [3–5]; however, the pathway regulating secondary growth in trees has not been identified. Here, we show that an aspen receptor kinase PttPXY and its peptide ligand PttCLE41 are functional orthologs and act to control a multifunctional pathway that regulates both the rate of cambial cell division and woody tissue organization. Ectopic overexpression of PttPXY and PttCLE41 genes in hybrid aspen resulted in vascular tissue abnormalities and poor plant growth. In contrast, precise tissue-specific overexpression generated trees that exhibited a 2-fold increase in the rate of wood formation, were taller, and possessed larger leaves compared to the controls. Our results demonstrate that the PXY-CLE pathway has evolved to regulate secondary growth and manipulating this pathway can result in dramatically increased tree growth and productivity.
Graphical Abstract
Highlights
-
•
PXY receptor kinase and CLE peptide signal to regulate radial growth in trees
-
•
Engineering PXY-CLE expression can lead to large increases in wood formation
-
•
Manipulating PXY-CLE also leads to increased tree height and leaf size
-
•
The results can be used to generate trees that are more productive
Etchells et al. show that altering the expression of the poplar homologs of the receptor kinase PXY and its peptide ligand CLE41 results in increased cambial cell division in hybrid aspen. 2-fold increases in the rate of wood formation were observed, demonstrating that engineering PXY/CLE41 signaling offers a means to increase tree productivity.
Results and Discussion
The PXY-CLE Signaling Pathway Is Conserved in Trees and Acts to Regulate Secondary Growth
Wood is composed of xylem cells that arise from divisions of stem cells that reside within the vascular meristem, known as the cambium or procambium. One mechanism that promotes cell division in vascular meristems of Arabidopsis involves phloem-specific expression of CLE41 that encodes a peptide ligand known as TDIF. TDIF is perceived by a receptor kinase, PXY (also known as TDR), that is expressed in the adjacent stem cells of the procambium [3–6]. PXY controls both the orientation [3, 4] and rate of cell division in procambial stem cells [7, 8] and inhibits their differentiation into xylem [5, 9]. Consequently, while ectopically overexpressing CLE41 in Arabidopsis increases the number of cells in vascular bundles, these increases are accompanied by repression of xylem differentiation and loss of vascular organization [3, 5, 10]. Furthermore, output from the pathway is regulated by a negative feedback loop in which CLE41 expression results in downregulation of PXY [3]. To determine whether PXY-CLE41 signaling is conserved in poplar, we cloned putative orthologs of PXY and CLE41 genes from the hybrid aspen (Populus tremula × P. tremuloides), referred to hereafter as PttPXY and PttCLE41, respectively. When overexpressed in Arabidopsis, 35S::PttCLE41 lines demonstrated a loss of vascular organization, increased numbers of cells per vascular bundle, and decreased plant height (Figures S1A, S1B, S1E, and S1F). The 35S::PttPXY construct complemented the Arabidopsis pxy mutant phenotype (Figures S1C, S1D, S1G, and S1H), and this complemented line also restored the ability of the plants to respond to overexpression of the AtCLE41 ligand (Figures S1G and S1H). As such, both PttCLE41 and PttPXY clones act as functional orthologs of their respective Arabidopsis genes. Furthermore, expression of PttPXY in Arabidopsis plants already engineered for tissue-specific AtCLE41 overexpression resulted in increased plant biomass (Figure S1I).
Ectopic Expression of PttCLE41 or PttPXY Leads to Abnormal Vascular Tissue Development in Trees
We investigated the consequence of constitutively overexpressing these genes in trees by making use of the 35S promoter that is known to give widespread expression in hybrid aspen [11]. We used our 35S::PttPXY and 35S::PttCLE41 constructs (see above) individually or overexpressed both genes together in a single binary plasmid that contained 35S::PttCLE41 and rolD::PttPXY cassettes. To varying degrees, all independent lines (n = 15) of 35S::PttCLE41 hybrid aspen had intercalated xylem and phloem (Figure 1A). 35S::PttPXY lines (n = 10) also demonstrated disrupted organization in parts of the xylem, but to a much lesser extent than seen in 35S::PttCLE41 (Figure 1A). 7 out of 15 35S::PttCLE41-rolD::PttPXY lines appeared normal, whereas the remaining 8 exhibited varying degrees of tissue disruption (Figure 1A). None of these lines led to significant increases in tree growth; in fact, 35S::PttCLE41 lines were significantly shorter than wild-type (Figures S2A and S2B), exhibiting various growth abnormalities (Figure 1B).
Figure 1.

Phenotypes of Hybrid Aspen Ectopically Overexpressing PttCLE41 and/or PttPXY Genes
(A) Sections from tissue-culture-grown plantlets 3 weeks post-rooting. Where two images are shown in the upper panel, they were selected to show the range of phenotypes observed. Scale bars represent 200 μM (upper panels) and 50 μM (lower panels). Red asterisks show examples of organized files of cells. The xylem (x) and phloem (ph) are indicated.
(B) Representative greenhouse-grown plants 3 months after transfer to soil.
(C) Phenotypes of hybrid aspen with targeted overexpression of PttCLE41 and PttPXY. Left-hand panels show sections from tissue-culture-grown plantlets 3 weeks post-rooting while greenhouse-grown plants 3 months after transfer to soil are shown on the right. Scale bars represent 200 μM. The xylem (x) and phloem (ph) are also indicated. Arrows highlight the disrupted xylem. See also Figure S1.
Tissue-Specific Expression of PttPXY and PttCLE41 Increases Vascular Cell Division and Retains Normal Vascular Tissue Organization
We hypothesized that the tissue-specific expression of both PttPXY and PttCLE41 might be important both for tissue organization and for maximizing cambial cell division. Transcriptomic data show that in poplar, PXY is expressed predominantly in the cambium and at a low level in the xylem [12]. Poplar microarray data identified the ANTEGUMENTA (ANT) gene as highly expressed only within the division zone [12]. Using an early draft of the Populus trichocarpa genome [13] as a guide, we identified and cloned a putative promoter from hybrid aspen (PttANT), although better annotation of the genome subsequently suggested the PttANT promoter fragment contained sequences both upstream and downstream of the putative transcriptional start site. Analysis of leaves from PttANT::GUS plants showed clear vascular-specific GUS expression, while in the stems, GUS activity was restricted to the dividing cambial zone (Figure 2B), consistent with our initial interpretation of the expression data. We also identified and cloned regulatory sequences from a phloem-specific lectin gene, PHLOEM PROTEIN2 (PP2), from Populus trichocorpa (PtPP2). GUS analysis verified this promoter as vascular tissue-specific in the leaves and giving excellent phloem-specific expression in stems (Figure 2C). These promoters were used to generate three constructs designed to give tissue-specific increases in expression: PttANT::PttPXY, PtPP2::PttCLE41, and PtPP2::PttCLE41-PttANT::PttPXY. In contrast to 35S::PttCLE41 (Figure 1A), PtPP2::PttCLE41 lines demonstrated highly organized vasculature in all 14 lines examined (Figure 1C). 7 out of 15 PttANT::PttPXY lines demonstrated minor disruptions in xylem morphology (Figure 1C, arrow) similar to those observed in 35S::PttPXY trees (Figure 1A); however, all 12 independent PtPP2::PttCLE41-PttANT::PttPXY double overexpression lines analyzed exhibited highly organized vascular tissue comparable to that of wild-type controls (Figure 1C). Strikingly, PtPP2::PttCLE41, PttANT::PttPXY, and PtPP2::PttCLE41-PttANT::PttPXY double overexpression lines clearly demonstrated increases in the number of vascular cells as early as 3 weeks post-rooting in tissue culture (Figure S2C).
Figure 2.

Expression Patterns Derived from PttANT and PtPP2 Promoters
(A–C) GUS-stained and cleared control (A), PttANT::GUS (B), and PtPP2::GUS (C) plant material. Upper panels show leaves; lower panels are transverse stem sections. Scale bars represent 200 μm (upper panels) and 100 μm (lower panels).
Tissue-Specific Expression of PttPXY and PttCLE41 Results in Trees that Grow Faster
We further monitored the growth of these transgenic hybrid aspen trees following transfer to soil and maintenance in the greenhouse. Over a 6-month period, PtPP2::PttCLE41, PttANT::PttPXY, and PtPP2::PttCLE41-PttANT::PttPXY plants grew normally (Figure 1C) and were consistently larger than the control plants, with both greater stem diameter and plant height (Figures 3A and 3B). PtPP2::PttCLE41-PttANT::PttPXY lines gave the largest increase in radial growth, and after 6 months in the greenhouse, they exhibited a 35% increase in stem diameter compared to untransformed controls and a 10% increase compared to PtPP2::PttCLE41, the next best-performing genotype (Figure 3A). The PtPP2::PttCLE41-PttANT::PttPXY lines also demonstrated a 56% increase in height over their wild-type counterparts and a 12% increase in height over the next best-performing transgenic line (PttANT::PttPXY) (Figure 3B). This increase was due to a generally faster growth rate, with PtPP2::PttCLE41-PttANT::PttPXY plants having on average 90 internodes compared to a mean of 60 for control plants (Figure 3C), as well as to an increase in internode length (Figure 3D). While the plants appeared morphologically normal (Figure 1C), the PtPP2::PttCLE41-PttANT::PttPXY lines also exhibited increases in leaf area (Figure 3E), with the average leaf area increasing by almost 2-fold. These increases in growth may reflect PXY/CLE signaling acting on other aspects of plant development or be a consequence of increases in sink strength. Although further work is needed to test these hypotheses and to understand the basis of these developmental changes, they do contribute to a general increase in biomass that is likely to further improve the effectiveness of any biotechnological application of these discoveries.
Figure 3.

Growth Characteristics of Trees with Targeted PttCLE41/PttPXY Overexpression
(A and B) Mean stem diameter (A) and plant height (B) measurements from hybrid aspen grown in soil are shown. Trees rooted in April were measured at 15 weeks (July), 26 weeks (August), and 33 weeks (October).
(C–F) Further analysis of 6-month-old plants: number of internodes (C), length of 50th internode (D), leaf area calculated from measurements of five leaves from around the 50th internode (E), and xylem cell number in a sector, with a central angle of 40°, of a stem transverse section taken from the 50th internode (F).
(G) Graph showing dry weight of 10-cm pieces of sapling stem. Samples were taken from the base, middle (50th internode), and top, except for 35S::PttCLE41, which had less than 50 internodes and a section taken midway between the top and bottom was used instead.
All p values were calculated with an ANOVA and a least significant difference (LSD) post hoc test; n = 15 (A–E) or n = 8 (F and G). Error bars indicate the SE. See also Figure S2 and Table S1.
Tissue-Specific Expression of PttPXY and PttCLE41 Results in Large Increases in Wood and Biomass Formation
To better understand the cause of the increases in stem diameter in PtPP2::PttCLE41-PttANT::PttPXY lines, at 33 weeks, we harvested half of the trees from each line and sectioned stem material in order to perform cell counts for each line as described in Figure S3. In order to examine material from a similar developmental stage and to account for the differing sizes of the trees examined, we carried out the analysis on material from the 50th internode. We observed a dramatic increase in xylem cell numbers that correlated with the increase in stem diameter, with PtPP2::PttCLE41-PttANT::PttPXY lines having the largest number of xylem cells, 189% that of control plants (Figure 3F). Within individual lines, there was also a correlation between cell numbers and PttCLE41 expression and, to a lesser extent, with PttPXY expression (Figure S4). To determine whether it was possible to increase wood formation without altering xylem morphology, we adapted Cellprofiler [14] to measure a number of morphological characteristics of the xylem (Figure S3). The analysis revealed no significant differences in average cell size, average cell lumen size, average cell wall area, and vessel numbers as a proportion of total xylem cells in PtPP2::PttCLE41-PttANT::PttPXY compared to controls lines (Table S1), indicating that the increased wood production did not alter wood morphology.
To determine whether the improved growth characteristics led to increased woody biomass, we allowed the remaining trees to grow for an additional 6-month period, after which we determined dry weight (Figure 3G) and wet weight (Figure S2D) at various points along the stem. Consistent with our previous observations, measurement at the base, at the 50th internode (middle), and at the top of the stem demonstrated that PtPP2::PttCLE41-PttANT::PttPXY lines exhibited significant increases in dry weight in comparison to other lines used in this study. In particular, at the middle and base of trees, the dry weight of PtPP2::PttCLE41-PttANT::PttPXY stem segments were on average more than twice the weight of the control plants.
In order to ensure that the differences observed were reproducible, we clonally propagated material from six independent PtPP2::PttCLE41-PttANT::PttPXY lines. We monitored the growth of these plants weekly, starting shortly after transfer to soil. The diameter of several clones was significantly bigger than wild-type at all stages monitored (Figure 4). There was also variation between clones such that plants from line 2 were both significantly taller and exhibited a significantly larger diameter than plants from line 3 at all five time points examined (Figure 4).
Figure 4.

Growth of Clonally Propagated Plants Derived from Independent Transformants of PtPP2::PttCLE41-PttANT::PttPXY
Diameter (top) and height (bottom) of plants were measured at weekly intervals starting 4 weeks after transfer from tissue culture to soil. Asterisk indicates a p value of less than 0.05 compared to the controls. All p values were calculated with an ANOVA and a LSD post hoc test; n = 6 for the control; n = 5 for PtPP2::PttCLE41-PttANT::PttPXY lines 1, 3, and 9; n = 4 for lines 2 and 4. Error bars indicate the SE. See also Figure S4.
Conclusions
Trees represent a huge natural resource used for the production of paper, fuel, and materials and are an increasingly important carbon sink [15] that can help to ameliorate anthropogenic increases in atmospheric CO2. Recently, trees have also been the focus of intense interest as a renewable source of plant biomass that may be converted into bioethanol [16] and other chemicals for the rapidly expanding field of industrial biotechnology [17]. The majority of biomass in trees is derived from radial growth that is characterized by growth rings in the wood. The size of each growth ring is intimately linked to the environmental conditions during the growing season that year.
Our data suggest that the PXY-CLE pathway functions in trees to regulate secondary growth and is likely to be central to the way in which trees evolved secondary growth. Together, the analysis demonstrates that by engineering the PXY-CLE pathway, we were able to dramatically increase secondary growth in plants shortly after they were first rooted (Figures 3 and S2C), the earliest point they could be analyzed, and the increase in xylem was maintained in plants grown for up to a year (Figures 4 and S2D). These results indicate that this pattern of growth is likely to continue during the lifetime of the tree, thereby providing a means of dramatically increasing tree productivity that would help to meet the increasing demand for renewable resources.
While tree productivity may benefit from anthropogenic increases in atmospheric CO2, climate models and recent changes in weather pattern strongly suggest that we are entering a period in which large parts of the globe experience more frequent exposure to extreme and changeable weather [18] that is likely to have detrimental effects on growth. It will be important to establish whether manipulating PXY-CLE signaling will enable us to override the environmental cues that normally regulate plant growth and thus enable us to generate trees that are able to maintain high productivity even when exposed to more extreme environmental conditions.
Author Contributions
J.P.E., M.K., and S.R.T. designed the experiments. L.S.M., J.P.E., L.C., and M.K. carried out the experimental work. S.R.T., J.P.E., and M.K. wrote the manuscript.
Acknowledgments
The authors are grateful to Bjorn Sundberg for providing the T89 clone. We thank Joe Ogas, Patrick Gallois, Thomas Nuhse, Stephen High, and Minsung Kim for critical reading of the manuscript. This work was funded by BBSRC (grant number BB/H019928).
Footnotes
This is an open access article under the CC BY license (http://creativecommons.org/licenses/by/4.0/).
Accession Numbers
The GenBank accession numbers for PttPXY and PttCLE41 reported in this paper are KP682331 and KP682332, respectively.
Supplemental Information
References
- 1.Miyashima S., Sebastian J., Lee J.Y., Helariutta Y. Stem cell function during plant vascular development. EMBO J. 2013;32:178–193. doi: 10.1038/emboj.2012.301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ursache R., Nieminen K., Helariutta Y. Genetic and hormonal regulation of cambial development. Physiol. Plant. 2013;147:36–45. doi: 10.1111/j.1399-3054.2012.01627.x. [DOI] [PubMed] [Google Scholar]
- 3.Etchells J.P., Turner S.R. The PXY-CLE41 receptor ligand pair defines a multifunctional pathway that controls the rate and orientation of vascular cell division. Development. 2010;137:767–774. doi: 10.1242/dev.044941. [DOI] [PubMed] [Google Scholar]
- 4.Fisher K., Turner S. PXY, a receptor-like kinase essential for maintaining polarity during plant vascular-tissue development. Curr. Biol. 2007;17:1061–1066. doi: 10.1016/j.cub.2007.05.049. [DOI] [PubMed] [Google Scholar]
- 5.Hirakawa Y., Shinohara H., Kondo Y., Inoue A., Nakanomyo I., Ogawa M., Sawa S., Ohashi-Ito K., Matsubayashi Y., Fukuda H. Non-cell-autonomous control of vascular stem cell fate by a CLE peptide/receptor system. Proc. Natl. Acad. Sci. USA. 2008;105:15208–15213. doi: 10.1073/pnas.0808444105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kondo T., Sawa S., Kinoshita A., Mizuno S., Kakimoto T., Fukuda H., Sakagami Y. A plant peptide encoded by CLV3 identified by in situ MALDI-TOF MS analysis. Science. 2006;313:845–848. doi: 10.1126/science.1128439. [DOI] [PubMed] [Google Scholar]
- 7.Etchells J.P., Provost C.M., Mishra L., Turner S.R. WOX4 and WOX14 act downstream of the PXY receptor kinase to regulate plant vascular proliferation independently of any role in vascular organisation. Development. 2013;140:2224–2234. doi: 10.1242/dev.091314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hirakawa Y., Kondo Y., Fukuda H. TDIF peptide signaling regulates vascular stem cell proliferation via the WOX4 homeobox gene in Arabidopsis. Plant Cell. 2010;22:2618–2629. doi: 10.1105/tpc.110.076083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kondo Y., Ito T., Nakagami H., Hirakawa Y., Saito M., Tamaki T., Shirasu K., Fukuda H. Plant GSK3 proteins regulate xylem cell differentiation downstream of TDIF-TDR signalling. Nat. Commun. 2014;5:3504. doi: 10.1038/ncomms4504. [DOI] [PubMed] [Google Scholar]
- 10.Whitford R., Fernandez A., De Groodt R., Ortega E., Hilson P. Plant CLE peptides from two distinct functional classes synergistically induce division of vascular cells. Proc. Natl. Acad. Sci. USA. 2008;105:18625–18630. doi: 10.1073/pnas.0809395105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Nilsson O., Little C.H.A., Sandberg G., Olsson O. Expression of two heterologous promoters, Agrobacterium rhizogenes rolC and cauliflower mosaic virus 35S, in the stem of transgenic hybrid aspen plants during the annual cycle of growth and dormancy. Plant Mol. Biol. 1996;31:887–895. doi: 10.1007/BF00019475. [DOI] [PubMed] [Google Scholar]
- 12.Schrader J., Nilsson J., Mellerowicz E., Berglund A., Nilsson P., Hertzberg M., Sandberg G. A high-resolution transcript profile across the wood-forming meristem of poplar identifies potential regulators of cambial stem cell identity. Plant Cell. 2004;16:2278–2292. doi: 10.1105/tpc.104.024190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Tuskan G.A., Difazio S., Jansson S., Bohlmann J., Grigoriev I., Hellsten U., Putnam N., Ralph S., Rombauts S., Salamov A. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray) Science. 2006;313:1596–1604. doi: 10.1126/science.1128691. [DOI] [PubMed] [Google Scholar]
- 14.Carpenter A.E., Jones T.R., Lamprecht M.R., Clarke C., Kang I.H., Friman O., Guertin D.A., Chang J.H., Lindquist R.A., Moffat J. CellProfiler: image analysis software for identifying and quantifying cell phenotypes. Genome Biol. 2006;7:R100. doi: 10.1186/gb-2006-7-10-r100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Stephenson N.L., Das A.J., Condit R., Russo S.E., Baker P.J., Beckman N.G., Coomes D.A., Lines E.R., Morris W.K., Rüger N. Rate of tree carbon accumulation increases continuously with tree size. Nature. 2014;507:90–93. doi: 10.1038/nature12914. [DOI] [PubMed] [Google Scholar]
- 16.Somerville C. The billion-ton biofuels vision. Science. 2006;312:1277. doi: 10.1126/science.1130034. [DOI] [PubMed] [Google Scholar]
- 17.Raunikar R., Buongiorno J., Turner J.A., Zhu S. Global outlook for wood and forests with the bioenergy demand implied by scenarios of the Intergovernmental Panel on Climate Change. For. Policy Econ. 2010;12:48–56. [Google Scholar]
- 18.Palmer T. Atmospheric science. Record-breaking winters and global climate change. Science. 2014;344:803–804. doi: 10.1126/science.1255147. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.