

Abstract
Since the discovery of the “nude” mouse over 40 years ago, investigators have attempted to model human tumor growth in immunodeficient mice. The field has advanced significantly over the ensuing years due to improvements in the murine recipient of human tumors. These improvements include the discovery of the scid mutation and development of targeted mutations in the recombination activating genes 1 and 2 (Rag1null, Rag2null) that severely cripple the adaptive immune response of the murine host. More recently, mice deficient in adaptive immunity have been crossed with mice bearing targeted mutations designed to weaken the innate immune system, ultimately leading to the development of immunodeficient mice bearing a targeted mutation in the IL2 receptor common gamma chain gene (IL2rγnull). The IL2rγnull mutation has been used to develop several immunodeficient strains of mice, including the NOD-scid IL2rγnull (NSG) strain. Using NSG mice as human xenograft recipients, it is now possible to grow almost all types of primary human tumors in vivo, including most solid tumors and hematological malignancies that maintain characteristics of the primary tumor in the patient. Programs to optimize patient-specific therapy using patient-derived xenograft (PDX) tumor growth in NSG mice have been established at several institutions, including The Jackson Laboratory. Moreover, NSG mice can be engrafted with functional human immune systems permitting for the first time the potential to study primary human tumors in vivo in the presence of a human immune system.
Introduction
The use of human tumor cell lines has led to increased understanding of the molecular events that lead to malignancy and fueled the promise of rapid advances in cancer therapy. However, many of the mechanisms responsible for tumorigenicity are altered during cell culture, and drugs that show efficacy in vitro against human tumor cell lines are often ineffective when translated to primary tumors in patients. This can be due to many factors, including genetic alterations that occur through multiple passages of the cell line, lack of appropriate human tumor stromal-associated cell populations that are important for tumor growth and maintenance, lack of an immunologic environment in which the tumor is exposed to in vivo, and issues associated with vascularization and the three-dimensional structure of the tumor in vivo as compared to a two-dimensional cell layer in vitro. In attempts to overcome these issues, human tumor cell lines and solid human tumors were first engrafted into nude mice and later into early models of scid mice, but these early models have shown limited success for investigating the pathogenesis of tumors and the efficacy of drugs on their growth and survival.
Therefore, a longstanding goal for cancer investigators has been to engraft in an animal model primary patient-derived human tumors (referred to as patient-derived xenografts, or PDX) that maintain the genetic, phenotypic, and functional characteristics of the primary tumor after transplantation. To accomplish this, primary human tumors have been engrafted into numerous strains of immunodeficient mice, but until recently there has been varying and often poor success depending on the nature of the primary tumor and on the immunodeficient recipient used. This lack of success has resulted in a bottleneck in the preclinical evaluation of drugs and in their therapeutic efficacy on primary human tumors in vivo, leaving a major knowledge gap on drug efficacy prior to clinical translation. Recently, the development of immunodeficient mice bearing a targeted mutation in the IL2 receptor common gamma chain gene (IL2rγnull) has permitted many of the primary human tumor types that previously could not be grown in immunodeficient mice to engraft and maintain the characteristics of the original primary patient tumor, and in certain cases exhibit the ability to metastasize in a manner similar to that seen in the primary tumor in patients.
Here, we present a short historical perspective of immunodeficient mouse models for cancer research and discuss the recent “state of the art” advances in the field. We will focus on NOD-PrkdcscidIl2rgtmiWjl (NSG) mice as models for PDX tumors and as avatars (representing the individual patient primary tumor) for design and evaluation of patient-specific therapy. Moreover, NSG mice can be engrafted with a functional human immune system, presenting for the first time the promising possibility for studying primary human tumor - immune system interactions in vivo and for testing the therapeutic efficacy of immunomodulatory drugs on human tumors without putting patients at risk. Finally, we will describe limitations in the currently available models and opportunities for overcoming these challenges.
Development of immunodeficient mouse models for the in vivo investigation of human tumors
Advancements in the use of immunodeficient mice for translational biomedical research in multiple fields of science has been extensively reviewed (Shultz et al. 2007; Shultz et al. 2012), and advancements in using immunodeficient mice for cancer research and therapy has closely paralleled and has been dependent on advancements and improvements in the immunodeficient murine hosts (Table 1).
Table 1. Engraftment of human tumors and immune systems in strains of immunodeficient mice.
| Common Strain Name | Mutant gene Nomenclature | Solid Tumor Cell Lines | Primary Solid Tumors | Leukemias | Human Immune Systems | References |
|---|---|---|---|---|---|---|
| Nudeaa | Foxn1nu | + | +/- | - | - | (Fogh et al. 1977; Sharkey and Fogh 1984; Giovanella and Fogh 1985; Price and Zhang 1990) |
| C.B17-scid BALB/c-scid | Prkdcscid | + | + | +/- | +/- | (Reddy et al. 1987; Kamel-Reid et al. 1989; Charley et al. 1990; Mueller and Reisfeld 1991; Cesano et al. 1992; Taylor et al. 1992; Itoh et al. 1993; Flavell 1996; O'Reilly et al. 1996; Sakakibara et al. 1996; Uckun 1996; Visonneau et al. 1998) |
| NOD-scid NOD-Rag1null | Prkdcscid Rag1tm1Mom | ++ | ++ | + | + | (Baersch et al. 1997; Bonnet and Dick 1997; Hudson et al. 1998; Wang et al. 1998; Ailles et al. 1999; Fusetti et al. 2000; Pilarski et al. 2000; Appelbaum et al. 2001; Frost et al. 2004; Huang et al. 2004; Eisterer et al. 2005; Xue et al. 2005; Pearce et al. 2006; Li et al. 2007) |
| NSG | Prkdcscid Il2rgtm1Wjl | +++ | +++ | ++ | +++ | (Shultz et al. 2007; Shultz et al. 2012) |
Includes studies of nude mice on non-inbred and a variety of inbred strain background
Athymic Foxn1nu (nude) mice
The first pivotal breakthrough in the use of immunodeficient mice in the study of human cancer was the discovery of congenitally athymic nude mice in the 1960s (Table 1). The nude mutation prevents development of functional T cells and provided an early model for engraftment of human cell lines derived from solid tumors (Fogh et al. 1977). However, the presence of an intact humoral adaptive immune system and an intact innate immune system, including high NK cell activity limits engraftment with most primary solid human tumors and prevents engraftment of human normal or malignant hematopoietic cells.
C.B17-Prkdcscid (scid) mice
The next major breakthrough in the field of PDX modeling was the description of a spontaneous mutation in C.B17 mice termed “scid” (Prkdcscid, protein kinase DNA activated catalytic polypeptide) (Bosma et al. 1983; Blunt et al. 1996). The scid mutation largely prevents the development of mature T and B lymphocytes of the adaptive immune system. The term “SCID” has now been adapted to refer to all severely immunodeficient strains of mice, including those expressing the Rag1null or Rag2null mutations. Direct comparisons have shown that C.B17-scid mice could be engrafted with a larger range of human solid tumors than nude mice (Phillips et al. 1989), and for the first time permitted the engraftment of low levels of human hematopoietic cells as well as certain hematological malignancies that did not grow in nude mice (Table 1).
Although more receptive as hosts for human cells and tissues as compared to nude mice, mature T and B cells develop in some C.B17-scid mice upon aging (Shultz et al. 2007). Additionally, C.B17-scid mice have an intact innate immune system including moderate NK cell activity that reduces the ability of human hematopoietic cells and PDX tumors to engraft and grow. Furthermore, it has been shown that C.B17-scid mouse NK cells are cytotoxic for human HSC and tumor initiating cells (TIC) (Greiner et al. 1998; Sotiropoulou et al. 2006; Tian et al. 2006), further limiting their utility for the study of tumor growth and therapy.
NOD.C.B17-Prkdcscid (NOD-scid) mice
To reduce NK cell activity and other innate immune components in the murine host, we (and others) backcrossed the scid mutation onto several strain backgrounds, including the NOD/Lt strain background (Hesselton et al. 1995; Shultz et al. 1995). The NOD strain background confers intrinsic defects in innate immunity, including lowered NK cell activity, reduced levels of macrophage activation, abnormal dendritic cell development and function, and an absence of hemolytic complement. Combining these innate immune defects with the ablation of adaptive immunity using the Prkdcscid, Rag1null, or Rag2null mutations led to a murine host more receptive for human hematopoietic cells and primary tumors. When compared directly with C.B17-scid mice, NOD-scid mice exhibit an increased ability to support engraftment with solid human cancers and with transplantable lymphomas and leukemias that either failed to grow or grew only poorly in C.B17-scid mice (Williams et al. 1993; Hudson et al. 1998).
One of the major regulators of human cell engraftment in SCID mice, in addition to host adaptive and innate immune systems, is the signal regulatory protein alpha (Sirpa) gene (Takenaka et al. 2007). Sirpa is expressed on macrophages and other myeloid cell populations. The ligand for SIRPα is CD47, which is expressed on most human hematopoietic and non-hematopoietic cells. SIRPα binding to CD47 leads to a “do not eat me” signal to the macrophage, which in the absence of appropriate receptor-ligand interaction will lead to phagocytosis of the engrafted human cells by mouse macrophages. The NOD strain has a more human-like polymorphism in the Sirpα locus as compared to other mouse strains, enhancing the ability of human solid tumors, hematopoietic cells, and hematological malignancies to engraft in NOD-scid mice (Takenaka et al. 2007). However, many cancers and hematological malignancies still fail to engraft efficiently and grow in NOD-scid mice, largely due to remaining NK cell activity and other residual innate immune function.
NOD-Prkdcscid IL2rgnull (NSG) mice
A major leap forward in the engraftment of primary human cells, tissues, and tumors was the development of immunodeficient mice bearing a targeted mutation in the IL2-receptor common gamma chain gene (IL2rgnull). The IL2rg gene is responsible for high affinity signaling for the IL2, IL4, IL7, IL9, IL15, and IL21 receptors, and lack of signaling through these receptors cripples both the adaptive and innate immune system (Cao et al. 1995; DiSanto et al. 1995; Ohbo et al. 1996). When combined with the scid, Rag1null, or Rag2null mutations, a mouse completely deficient in adaptive immunity and severely deficient in innate immunity is generated that is highly receptive to engraftment of human cells, tissues, and primary tumors (Shultz et al. 2007; Shultz et al. 2012).
Three major strains of immunodeficient IL2rγnull mice have been developed, and their characteristics have been reviewed (Shultz et al. 2007; Shultz et al. 2012). These immunodeficient IL2rγnull mice differ in strain background (NOD vs. BALB/c vs. BALB/c × 129 mixed background), the mutation used to induce their immunodeficiency in the adaptive immune system (scid or Rag1null or Rag2null), and in the mutation in the IL2rγ gene itself (complete null or truncated). These variables result in differences in their ability to be engrafted with human cells and tissues, particularly in engraftment of human hematopoietic cells, immune systems, and primary malignancies (Shultz et al. 2012). NODShi.Cg-Prkdcscid Il2rgtm1Sug (NOG) mice have also been successfully used as hosts for primary human tumors (Nakamura and Suemizu 2008). The major difference between the NSG and NOG strains is that the Il2rg targeted mutation used to develop the NSG strain is a complete null so that no IL2rg is expressed and cytokines cannot bind while the Il2rg mutation in the NOG strain produces a protein that is expressed and will bind cytokines but cannot signal. For the purposes of this chapter, we will focus on the use of NSG mice for the study of engrafted human PDX tumors. Direct comparisons revealed that many more human cancers, particularly hematological malignancies, can be engrafted and studied in NSG mice than can be engrafted in NOD-scid mice (Agliano et al. 2008).
NSG mice engrafted with human PDX tumors as models for testing drug efficacy
Almost all classes and types of primary solid or hematological human tumors transplanted into NSG mice have grown, providing new in vivo models for evaluation of tumorigenicity, identification of tumor initiating cells, and in vivo analyses of therapeutic efficacy of drugs on primary human tumors without putting patients at risk.
Solid tumors
A wide variety of solid human tumors can be grown in NSG mice (Table 2). In many cases tumor growth is dependent on the site of implantation (See protocol 1).
Table 2. Engraftment of human primary tumors in NSG Mice.
Includes studies in NRG mice
The characteristics of the solid tumors grown in NSG mice appear to closely recapitulate the genetic and phenotypic characteristics of the primary tumor in patients. This provides the opportunity to test drug therapy on the primary tumor in vivo and to correlate the genetics of the primary tumor with drug efficacy. Aggressive cancers are thought to induce stroma more efficiently and the stroma is considered an important target for drug therapy (Engels et al. 2012). A major advance in the use of NSG mice as recipients of solid PDX tumors is the ability to maintain the stromal cell population following transplantation into the primary recipient. In NSG recipients the stromal component of a subcutaneously transplanted non-small cell lung carcinoma remained intact, and was similar morphologically to the stromal components of the primary tumor (Simpson-Abelson et al. 2008). This included the passenger T cells present in the stromal compartment of the PDX lung tumor. These T cells remained quiescent, as was observed in the primary tumor. Interestingly, when tumor-bearing mice were injected with human recombinant IL12, the human passenger leukocytes (which were predominately CD3+ T cells) became activated, acquired a memory/effector phenotype including production of IFNγ, and migrated from the tumor to the liver, lung, and spleen of the NSG host (Simpson-Abelson et al. 2008).
Hematological tumors
A wide variety of human hematological malignancies can be grown in NSG mice (Table 2). The ability to use engrafted human hematological malignancies for preclinical analyses of drug efficacy is exemplified by the use of an anti-CD47 antibody for the treatment of human acute lymphoblastic leukemia (ALL) (Chao et al. 2011a). Although most ALL patients have a favorable prognosis using standard chemotherapy, a subset of high risk patients express high levels of CD47, resulting in protection from phagocytosis. (Subramanian et al. 2006). Transplantation of ALL from the high CD47 tumor-expressing patients led to robust engraftment of CD47+ tumor cells in NSG mice. Treatment of tumor-bearing mice with a blocking anti-human CD47 antibody led to tumor cell phagocytosis by host mouse macrophages (Chao et al. 2011a). Similar therapeutic effects were observed using a blocking anti-human CD47 antibody for treatment of human acute myelogenous leukemia (AML) (Majeti et al. 2009) and aggressive metastatic leiomyosarcoma tumors (Edris et al. 2012b) engrafted in NSG mice. Anti-human CD47 antibody acted synergistically with anti-human CD20 antibody (Rituximab) in NSG mice engrafted with non-Hodgkin's lymphoma. Interestingly, many human tumors express CD47, including solid tumors such as ovarian, breast, colon, bladder, glioblastoma, hepatocellular carcinoma, and prostate. Anti-human CD47 antibody has therapeutic effects on the growth of these tumors in NSG mice (Willingham et al. 2012). This latter report illustrates the power that the ability to engraft multiple types of human PDX tumors into NSG mice permits for evaluation of drug efficacy.
Immunodeficient mice engrafted with human PDX tumors as models for immunotherapy
One of the major goals for the use of humanized mice is to test immunotherapeutic approaches on primary human tumors in vivo. One of the simplest approaches described was to test the efficacy of IL27 on pediatric AML. Leukemia dissemination was severely hampered by recombinant human IL27 treatment of PDX AML engrafted in NSG mice, and genetic analyses of the tumors revealed a significantly reduced expression of their angiogenic and spreading-related genes (Zorzoli et al. 2012). In an example of cellular therapy, NSG mice were first engrafted with human CD34+ HSC to generate a human hematopoietic and immune system (Liu et al. 2012)., The engrafted mice were injected with human neuroblastoma cells and infused with ex vivo expanded NKT cells. NKT cell migration into the tumor was significantly inhibited by neutralization of the chemokine CCL20 secreted by tumor-associated macrophages, which was increased in states of hypoxia at the tumor site. Injection of NKT cells transduced with human IL15 protected the NKT cells from hypoxia, and dramatically enhanced their anti-metastatic activity. These in vivo analyses demonstrated a new mechanism of immune escape by tumors that can be reversed by adoptive cellular therapy with IL15-transduced NKT cells (Liu et al. 2012).
In a test of T cell immunotherapy, Provasi et. al used zinc finger nucleases to disrupt endogenous TCR β and α chains in primary human CD8 T cells, and then transduced cells with a TCR specific for the Wilms 1 (WT1) tumor antigen expressed by leukemic blasts (Provasi et al. 2012). The CD8 T cells did not cause graft-versus-host reactivity in irradiated recipients but were able to mediate specific anti-tumor activity and lead to long term survival of NSG mice engrafted with WT1+ PDXs. (Provasi et al. 2012). Another approach to engrafting human CD8 T cells having TCR specificity against tumor antigens was to transduce human HSCs with lentivirus encoding a HLA-A2-restricted TCR specific for the MART-1 melanoma antigen (Vatakis et al. 2011). Using a modified human liver/thymus immune system model commonly referred to as the BLT (bone marrow, liver, thymus) humanized mouse model (Shultz et al. 2012), human fetal liver CD34+ cells were transduced with the MART-1-specific TCR and injected into BLT mice engrafted with autologous fetal liver and thymus tissues. This led to the engraftment of human CD8 TCR transgenic-expressing T cells, which cleared human MART-1-expressing melanoma tumor cell lines in vivo. The TCR transgenic T cells required proper thymic selection in the HLA-A2 autologous thymus fragment as they could not kill non-HLA-A2 MART-1-expressing melanoma cells (Vatakis et al. 2011).
Although these validations of CD8 TCR-mediated anti-tumor activity provided in vivo evidence of the efficacy of these approaches for tumor therapy, the need to provide a transgenic TCR specific for each tumor antigen of interest is a major caveat. A new approach has been validated in tumor-bearing NSG mice as follows. A chimeric antigen receptor (CAR) was constructed in which a biotin-binding immune receptor consisting of an extracellular avidin is linked to a human CD3 signaling domain to activate T cells (Urbanska et al. 2012). Tumors can then be targeted with, for example, a tumor specific antibody that is conjugated with biotin to target the CD8 T cells transduced with the CAR to the tumor site where they become activated and acquire cytotoxic activity. This CAR approach has been shown to have therapeutic efficacy against human EpCam+ tumor cells (Urbanska et al. 2012). Extending this approach, human T cells were transduced with a CAR directed to the malignant pleural mesothelioma tumor antigen mesothelin (mesoCAR) and the cells were subsequently transduced with the chemokine receptor CCR2b to localize the T cells to the tumor (Moon et al. 2011). The malignant pleural mesotheliomas secreted high levels of CCL2, the chemokine for the CCR2b receptor. Infusion of the transduced CCR2b mesoCAR+ T cells into NSG mice bearing established tumors resulted in targeted T cell infiltration into the tumor and significantly reduced tumor size (Moon et al. 2011).
Finally, to study human tumor immunotherapy in the presence of a complete human immune system, NSG mice must be engrafted with HSCs that can generate a functional human immune system while at the same time support the growth of a human tumor. To begin to establish this system, NSG mice were engrafted with human CD34+ HSC and injected with human breast cancer cell lines (Wege et al. 2011). The human immune system developed and tumor cells were detectable in different lymphoid and non-lymphoid organs. NK cell accumulation and activation was observed in the tumors, and was enhanced by treatment with a recombinant human IL15/IL15-alpha receptor complex. This model combines MHC-mismatched tumor cells with human immune systems for the study of human immune system/tumor interactions.
The in vivo analysis of these human-specific tumor therapies was possible because 1) primary human tumors grow efficiently in NSG mice, and 2) NSG mice can be engrafted with functional human immune systems. Continued exploitation of the NSG model for modeling PDX tumor growth and analyses of therapies promises great potential for preclinical evaluation of multiple immunotherapeutic approaches without putting patients at risk.
Immunodeficient mice engrafted with human PDX tumors as models for identification and directed therapy towards tumor initiating cells
Cancers exhibit remarkable heterogeneity, and there are numerous hypotheses that this heterogeneity as well as resistance to therapy is due to the presence of tumor initiating cells (TIC). This cell population, also referred to as cancer stem cells (CSC), is thought to be responsible for propagation of the tumor in vivo (Visvader 2011). Immunodeficient mice have been widely used to study human HSC, which are defined functionally by their ability to recapitulate growth of the entire hematopoietic system in vivo (Dick 2008). Similarly, immunodeficient mice have been used to identify TIC based on functionality. These cells have been defined as malignant cells that fulfills the classic stem cell criteria (i.e., the ability to undergo self-renewal and the developmental potential to recapitulate all the cell types found in a given tissue).” (Krumbach et al. 2011). This can only be accomplished in vivo, and immunodeficient mice have been used to define human TIC for a number of human malignancies (Table 3). Many human TIC have been defined using NOD-scid mice, and more recently NSG mice (Kelly et al. 2007; Quintana et al. 2008; Ishizawa et al. 2010).
Table 3. Identification of human tumor initiating cells following engraftment in NOD-scid and NOD-scid IL2rγnull mice.
| Solid Tumors | Timor Initiating Cell Phenotype | Reference |
|---|---|---|
| Brain | CD133+ | (Singh et al. 2003) |
| Pancreas | CD44+CD24+ESA+ | (Li et al. 2007) |
| Breast | CD44+CD24—/low & Milk mucin (MM)+ and/or CD271+ | (Al-Hajj et al. 2003; Kim et al. 2012c) |
| Colon | CD133+ CD133+ and CD133— | (O'Brien et al. 2007; Ricci-Vitiani et al. 2007) (Shmelkov et al. 2008) |
| Glioma | high levels of IL13 receptor α2 | (Brown et al. 2012) |
| Head and Neck Squamous Cell Carcinoma | CD44+Lin— | (Prince et al. 2007) |
| Leiomyoma | Leiomyoma-derived side population (LMSP) identified by Hoechst 33342 dye staining | (Ono et al. 2012) |
| Melanoma | CD127+ or CD127— CSC phenotypically diverse TIC | (Quintana et al. 2008) |
| Ovarian | Both CD133+ and CD133— populations | (Stewart et al. 2011) |
| Prostate | CD49fhiTrop2hi | (Goldstein et al. 2010) |
| CD44+ | (Patrawala et al. 2006) | |
| Hematological Tumors | Tumor Initiating Cell Phenotype | Reference |
| Acute Myelogenous Leukemia | CD34+CD38— CD34-CD33+ (NPM mutated AML) | (Bonnet and Dick 1997); (Ishikawa et al. 2007) (Taussig et al. 2010) |
| Acute Lymphoblastic Leukemia | CD34+CD38+CD19+ & CD34+CD38—CD19+ | (Kong et al. 2008) |
| Multiple phenotypes depending on maturation stage of TIC used CD34+CD19—, CD34+CD19+, CD34—CD19+, (intrafemoral injection) | (le Viseur et al. 2008) | |
| Chronic Myelogenous Leukemia | CD34+CD38— | (Jiang et al. 2007) |
| Myeloma | CD34+ | (Pilarski and Belch 2002) |
| CD138+CD38hiCD19—CD45low/— | (Kim et al. 2012a) |
Identification of TIC in solid tumors and hematological malignancies has routinely been accomplished using cell surface phenotypic markers that can be used to sort individual cell subsets for adoptive transfer into immunodeficient recipients. For solid tumors, as an example, human brain TIC express CD133. As few as 100 CD133+ brain tumor cells can develop tumors upon adoptive transfer into NOD-scid mice whereas adoptive transfer of as many as 1×105 CD133-negative brain tumor cells engrafted but failed to form tumors (Singh et al. 2003; Singh et al. 2004). For AML, TIC in the bone marrow were identified as CD34+CD38—CD25+ or CD34+CD38—CD133+CD32+ using NSG mice (Saito et al. 2010a).
Although the existence of “true” TIC remains controversial (Dick 2008), the important question is whether specific drug targeting of TIC will provide better outcomes in developing patient-specific and tumor-specific therapies. This may in part be determined by the specific tumor being studied. For example, two extremes for the frequency of TIC in tumors are melanoma and AML. Primary human melanoma can be grown in NSG mice by injection of as few as 100 unfractionated tumor cells, and indeed, 27% of single cell melanoma transplants can form tumors in NSG mice (Quintana et al. 2008). These results demonstrate that the functional TIC frequency in this population is extremely high. In contrast, engraftment of AML in only 38% of sublethally irradiated newborn NSG mice was observed following injection of 4 million T cell-depleted bone marrow cells containing 80-90% blasts (Ishikawa et al. 2007; Ishikawa et al. 2008). These data suggest that essentially all the melanoma cells in the patient must be eradicated for effective therapy of the tumor, whereas fewer TIC are the key targets for treatment of AML. Identification and targeting of this small TIC cell subpopulation in AML may provide an effective therapeutic approach for curing AML in patients.
The frequency of the TIC population can vary widely between each of the tumor types being studied and the immunodeficient host being used, as demonstrated for melanoma, changing the immunodeficient host increased the detection of the frequency of melanoma TIC from 0.0001-0.1% in NOD-scid mice to 27% in NSG mice. A similar increase in TIC frequency in NSG mice as compared NOD-scid mice for squamous cell lung carcinoma was observed (Ishizawa et al. 2010). In contrast, no differences in TIC frequency in pancreatic adenocarcinoma or head and neck squamous cell carcinoma were observed when limiting numbers of the tumor cells were transplanted into NOD-scid vs. NSG mice (Ishizawa et al. 2010). These data suggest that the detection and frequency of functional TIC will increase dramatically in many, but not all tumors analyzed using NSG mice.
The NSG model is particularly useful for testing the efficacy of therapies aimed at eradicating TIC. In solid tumors, the TIC for high-grade glioma cells has been shown to express high levels of IL13 receptor α2 (Brown et al. 2012), representing a potential target for tumor therapy. Similar information has been obtained for hematological malignancies. The TIC for AML has been identified, and when injected into NSG mice, has been found to engraft in a quiescent form in an osteoblast-rich bone marrow niche that protects it from traditional chemotherapy that reduces the tumor burden, but does not eliminate the TIC in the marrow (Ishikawa et al. 2007). Administration of G-CSF led these TIC to enter the cell cycle and become susceptible to cell cycle-dependent chemotherapy resulting in apoptosis and elimination (Saito et al. 2010b). Although counter-intuitive that a cell cycle inducer in combination with chemotherapy would be important for eliminating AML tumor cells, this study provides preclinical evidence for efficacy of this approach. Furthermore, when AML TIC were defined phenotypically by adoptive transfer into NSG mice and isolated for genomic analyses, a TIC gene signature profile identified novel molecules in the TIC population as targets for potential therapies (Saito et al. 2010a).
Phenotypically identifying TIC can also provide valuable predictive information for clinical outcomes in patients. In breast cancer, two TIC populations have been identified based on their phenotypic characteristics as basal-like (CD271+) or luminal-like (milk mucin+, identified by the M18 antibody that recognizes branched glycans). NSG mice can provide specific information on a patient's tumor that can be used diagnostically, and when combined with genomic analysis of the TIC, provide prognostic information. Identification of TIC can also facilitate new treatment regimens aimed at preventing patient relapse. CD34+ cells were identified as TIC in human chronic myelogenous leukemia (CML) using NSG mice (Zhang et al. 2010). Treatment of patients with a tyrosine kinase inhibitor imatinib mesylate (Gleevec) induces apoptosis in dividing CML cells and drives patients into remission. However, when treatment is withdrawn, there is a significant relapse rate, suggesting that quiescent CML tumor initiating cells survive Gleevec treatment. Using NSG mice, CD34+ CML TIC and progenitor cells were engrafted, and the combination of Gleevec plus a histone deacetylase inhibitor that induces apoptosis in quiescent tumor cells was able to eliminate the tumor in the mice, and no relapse occurred following cessation of treatment (Zhang et al. 2010).
PDX Programs
A number of facilities have established PDX programs for studying patient tumor growth and response to experimental therapy in vivo without putting patients at risk. Figure 1 shows a generic diagram of PDX programs for evaluation of solid human tumors. For the engraftment of human leukemias (not shown), young adult or newborn NSG mice are engrafted intravenously, or in the case of adult recipients, cells may be injected directly into the bone marrow cavity (intrafemoral injection). Recipients may be conditioned with sublethal irradiation prior to engraftment.
Figure 1.
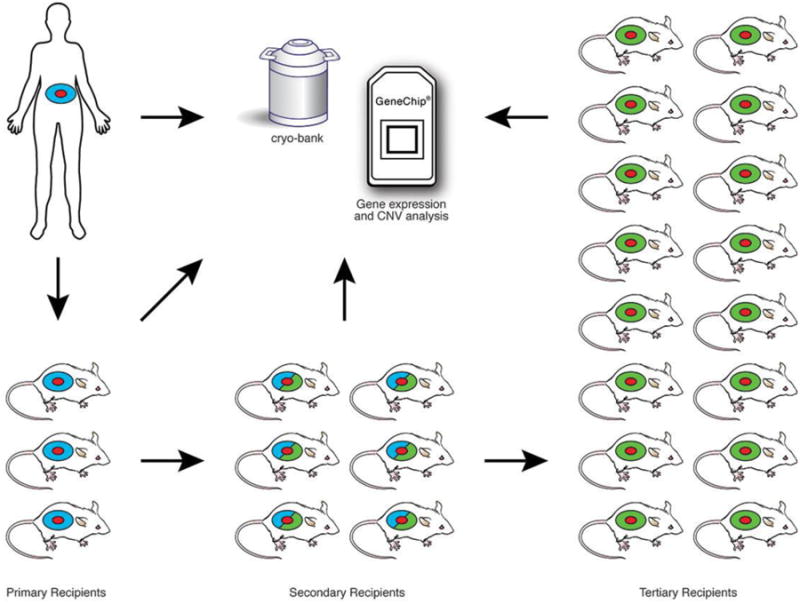
Engraftment and expansion of patient-derived solid tumor xenografts. Young adult NSG mice are engrafted with solid tumors (shown as red) Human stroma accompanying the tumor is shown in blue. Primary solid tumors are engrafted subcutaneously, orthotopically, or under the renal capsule. Established grafts are excised from primary recipients, a portion is cryopreserved, and a portion is analyzed for gene expression, including copy number variation (CNV). The remainder is expanded through serial transplantation in secondary and tertiary NSG recipients to generate cohorts of sufficient size for therapeutic trials. At each stage tumor samples are cryopreserved and gene expression analyzed to compare with the primary tumor analyzed directly ex vivo from the patient. The primary recipients of solid tumors retain human-derived stroma (blue). Tumors in secondary recipients contain a mixture of human-derived and mouse-derived stroma (mixture of blue and green). Tumors in tertiary recipients contain predominantly mouse stroma (green).
The PDX program at The Jackson Laboratory
To establish a PDX resource for the global research community, the Jackson Laboratory Primary Human Tumor Consortium has been formed between The Jackson Laboratory, UC-Davis Comprehensive Cancer Center and over 20 additional cancer centers. This high throughput operation enables the study of PDX tumors under standard conditions. Solid and hematological patient tumor specimens are engrafted into NSG mice and the first generation PDX tumors are compared with the primary tumor for fidelity of histopathology, gene expression and human SNP copy number variance (CNV). The PDX tumors are maintained at low passage so that patient tumor heterogeneity and histology fidelity are maintained and models can be selected for biomarker-driven translational medicine studies. Solid tumor PDX models are passaged as tumor fragments to maintain the tumor and for tissue fragment cryopreservation. By November 2012, the program has engrafted over 600 patient specimens and established 152 PDX models with a 35% tumor model establishment rate across 17 cancer therapeutic areas (N. Goodwin, unpubl.). The top five indications include 34 lung cancer, 26 brain cancer, 15 colorectal cancer, 14 bladder cancer, and 14 pancreatic cancers. The program has the capacity to generate by the 4th in vivo passage generation thousands of PDX tumor-bearing mice for high-throughput drug efficacy studies. Patient tissue sources include solid primary resected tumors, advanced metastases biopsies and pleural effusions, and leukapheresis and bone marrow aspirate specimens from hematological malignancies.
This approach is based on the recognition that tumors, such as non-small cell lung cancer are in fact quite heterogeneous tumors with multiple complex mechanisms underlying tumor development. Moreover, the genomic characterization of PDX tumors in immunodeficient mice has shown that the patient's primary tumor genomic profile is recapitulated in the PDX tumor derived from the explants (Monsma et al. 2012). Combining genomics with targeted cancer therapy is now possible to provide guidance for patient-specific therapies targeted to the particular molecularly-defined pathways that underlie tumor pathogenesis (Gandara et al. 2012).
In a study of human bladder cancer the PDX program at The Jackson Laboratory has teamed up with investigators at the University of California Davis. Bladder cancer is particularly difficult to treat in the clinic. By engrafting PDX-derived bladder cancers both subcutaneously and in the bladder cell wall, solid and invasive tumor models were established in NSG mice. The tumor-bearing mice were treated with nanoparticles coated with an amino acid sequence that recognized a bladder cancer-specific ligand allowing targeting to bladder cancer cells. These nanoparticles could deliver 3-times the systemic toxic dose of the drug with minimal side effects, and demonstrated therapeutic efficacy against both the solid and invasive bladder tumors established in NSG mice, resulting in prolonged overall survival as compared to control treated tumors. Moreover, the specificity of the nanoparticles was demonstrated in NSG mice engrafted with bladder tumors versus H232A lung cancer cells implanted in the contralateral flank. These data led directly to a clinical trial using the newly formulated nanoparticles (Lin et al. 2012).
Avatar mice
Can NSG mice engrafted with PDX tumors truly become Avatar's for primary patient tumors in predicting how the tumor will respond to therapy in the clinic? In fact, this is already happening. In a recent study out of Johns Hopkins, primary solid tumors as well as metastatic tumors from patients with refractory advanced cancers were transplanted into immunodeficient mice. These recipients were used to test various therapeutics to determine the best available approaches for treating individual patients (Hidalgo et al. 2011). In 14 patients in which therapies guided by results in PDX-bearing immunodeficient mice were developed, treatments were initiated based on the PDX results in 11 of the patients, which proved to have therapeutic efficacy. In 2 patients, no effective treatment in the PDX-bearing immunodeficient mice was identified, and 1 patient died prior to initiation of treatment therapies developed in the PDX-bearing immunodeficient mice. PDX mice have also proved valuable in recent studies of metastasis. Engraftment of primary human stage III cutaneous melanomas into NSG mice has shown that metastasis in the recipient mice correlates with patient-specific clinical outcomes. Human melanomas that metastasized efficiently in the mice eventually progressed to advanced stage IV disease in the patients (Quintana et al. 2012). These findings using PDX mice as avatars suggest that precision medicine using immunodeficient mice as avatars for study of patient tumors may provide personalized medicine for therapeutic decisions and cancer treatment in the clinic.
Remaining challenges and future directions
A note of caution: growth of a tumor following transplantation of the primary tumor does not always mean that the engrafted tumor is representative of the original transplanted tissue. Infections from a number of microorganisms are considered as risk factors for development of cancer in humans and approximately 20% of human cancers have been associated with infection by viruses, bacteria and parasites (Mazza 2010). Infectious agents carried by the primary tumor may induce tumors different than the transplanted primary tumor of human or even mouse origin. In a case of hepatocellular carcinoma, it has been reported that the tumors that grew at the implantation site were in many cases not the primary tumor, but were EBV-induced lymphoblastoid tumors that resulted from the outgrowth of the passenger B cells (Chen et al. 2012).
There are remaining challenges and opportunities. Novel NSG stocks under development transgenically express human HLA class I and class II molecules and microenvironmental factors, lack mouse MHC class I and II molecules, and have additional deficits in innate immunity. These stocks will enable novel approaches such as co-engraftment with PDX tumors and patient T cell populations (Shultz et al. 2012). A number of NSG stocks of mice have been developed that express human growth factors (Shultz et al. 2012). Many of these factors are species specific. For example, many myelomas are dependent on IL6 for their growth in vivo. Mouse IL6 will not support the growth of human myeloma cells (Peters et al. 1996), and most primary human myeloma cells fail to engraft in NSG mice. Engraftment with fetal human bone supports the growth of primary human myeloma grafts perhaps through the production of human IL6 (Kim et al. 2012a). Development of a NSG host that transgenically expresses human IL6 may permit better growth of IL6-dependent myelomas. AML engraftment efficiencies were also improved in NSG mice that transgenically expressed human IL3, GM-CSF and SCF (Wunderlich et al. 2010). Similar approaches may be used for enhancement of human breast cancers that require prolactin as mouse prolactin has a low affinity for human prolactin receptors (Utama et al. 2006). Furthermore, ubiquitously expressed GFP in NSG mice is being evaluated as a model for discriminating human-derived tumor stroma from mouse stroma. The maintenance of human tumor stroma is critical as a drug target in cancer (Ahmed et al. 2008) as well as functioning as a mediator in drug resistance (Sebens and Schafer 2012). The role of tumor-stromal interactions is described in a recent study demonstrating that CML stem and progenitor cells are protected from TKI-mediated inhibition in the presence of BM stromal cells (Zhang et al. 2012a). Finally, patient-derived stem cell populations, such as mesenchymal stem cells, might be able to replace tumor-associated stromal cells that are lost during multiple in vivo passages of the tumors (Fig. 1). These emerging stocks of NSG mice are providing novel tools for engraftment of primary human tumors and will contribute to the advancement of patient-specific therapy.
Acknowledgments
This work was supported by National Institutes of Health Cancer Core Grant CA034196, Diabetes Endocrinology Research Center grant DK32520, AI46629 and a fellowship to VH from the JDRF. The contents of this publication are solely the responsibility of the authors and do not necessarily represent the official views of the National Institutes of Health. We thank Barbara Tennent, Carol Bult, and Greg Cox for critical review of the manuscript. DLG is a consultant for Viacord Inc. and The Jackson Laboratory. LDS is a consultant for Viacord Inc.
References
- Agliano A, Martin-Padura I, Mancuso P, Marighetti P, Rabascio C, Pruneri G, Shultz LD, Bertolini F. Human acute leukemia cells injected in NOD/LtSz-scid/IL-2Rgamma null mice generate a faster and more efficient disease compared to other NOD/scid-related strains. Int J Cancer. 2008;123:2222–2227. doi: 10.1002/ijc.23772. [DOI] [PubMed] [Google Scholar]
- Agliano A, Martin-Padura I, Marighetti P, Gregato G, Calleri A, Prior C, Redrado M, Calvo A, Bertolini F. Therapeutic effect of lenalidomide in a novel xenograft mouse model of human blastic NK cell lymphoma/blastic plasmacytoid dendritic cell neoplasm. Clinical Cancer Res. 2011;17:6163–6173. doi: 10.1158/1078-0432.CCR-11-0212. [DOI] [PubMed] [Google Scholar]
- Ahmed F, Steele JC, Herbert JM, Steven NM, Bicknell R. Tumor stroma as a target in cancer. Curr Cancer Drug Targets. 2008;8:447–453. doi: 10.2174/156800908785699360. [DOI] [PubMed] [Google Scholar]
- Ailles LE, Gerhard B, Kawagoe H, Hogge DE. Growth characteristics of acute myelogenous leukemia progenitors that initiate malignant hematopoiesis in nonobese diabetic/severe combined immunodeficient mice. Blood. 1999;94:1761–1772. [PubMed] [Google Scholar]
- Akunuru S, Palumbo J, Zhai QJ, Zheng Y. Rac1 targeting suppresses human non-small cell lung adenocarcinoma cancer stem cell activity. PLoS One. 2011;6:e16951. doi: 10.1371/journal.pone.0016951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke MF. Prospective identification of tumorigenic breast cancer cells. Proc Natl Acad Sci U S A. 2003;100:3983–3988. doi: 10.1073/pnas.0530291100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Appelbaum FR, Rowe JM, Radich J, Dick JE. Acute myeloid leukemia. Hematology Am Soc Hematol Educ Program. 2001:62–86. doi: 10.1182/asheducation-2001.1.62. [DOI] [PubMed] [Google Scholar]
- Baersch G, Mollers T, Hotte A, Dockhorn-Dworniczak B, Rube C, Ritter J, Jurgens H, Vormoor J. Good engraftment of B-cell precursor ALL in NOD-SCID mice. Klin Padiatr. 1997;209:178–185. doi: 10.1055/s-2008-1043947. [DOI] [PubMed] [Google Scholar]
- Bagnara D, Kaufman MS, Calissano C, Marsilio S, Patten PE, Simone R, Chum P, Yan XJ, Allen SL, Kolitz JE, et al. A novel adoptive transfer model of chronic lymphocytic leukemia suggests a key role for T lymphocytes in the disease. Blood. 2011;117:5463–5472. doi: 10.1182/blood-2010-12-324210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bankert RB, Balu-Iyer SV, Odunsi K, Shultz LD, Kelleher RJ, Jr, Barnas JL, Simpson-Abelson M, Parsons R, Yokota SJ. Humanized mouse model of ovarian cancer recapitulates patient solid tumor progression, ascites formation, and metastasis. PLoS One. 2011;6:e24420. doi: 10.1371/journal.pone.0024420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blunt T, Gell D, Fox M, Taccioli GE, Lehmann AR, Jackson SP, Jeggo PA. Identification of a nonsense mutation in the carboxyl-terminal region of DNA-dependent protein kinase catalytic subunit in the scid mouse. Proceedings of the National Academy of Sciences of the United States of America. 1996;93:10285–10290. doi: 10.1073/pnas.93.19.10285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonapace L, Bornhauser BC, Schmitz M, Cario G, Ziegler U, Niggli FK, Schafer BW, Schrappe M, Stanulla M, Bourquin JP. Induction of autophagy-dependent necroptosis is required for childhood acute lymphoblastic leukemia cells to overcome glucocorticoid resistance. J Clin Invest. 2010;120:1310–1323. doi: 10.1172/JCI39987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonnet D, Dick JE. Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nat Med. 1997;3:730–737. doi: 10.1038/nm0797-730. [DOI] [PubMed] [Google Scholar]
- Boonyaratanakornkit JB, Yue L, Strachan LR, Scalapino KJ, LeBoit PE, Lu Y, Leong SP, Smith JE, Ghadially R. Selection of tumorigenic melanoma cells using ALDH. J Invest Dermatol. 2010;130:2799–2808. doi: 10.1038/jid.2010.237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bosma GC, Custer RP, Bosma MJ. A severe combined immunodeficiency mutation in the mouse. Nature. 1983;301:527–530. doi: 10.1038/301527a0. [DOI] [PubMed] [Google Scholar]
- Brown CE, Starr R, Aguilar B, Shami AF, Martinez C, D'Apuzzo M, Barish ME, Forman SJ, Jensen MC. Stem-like tumor-initiating cells isolated from IL13Ralpha2 expressing gliomas are targeted and killed by IL13-zetakine-redirected T Cells. Clin Cancer Res. 2012;18:2199–2209. doi: 10.1158/1078-0432.CCR-11-1669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao X, Shores EW, Hu-Li J, Anver MR, Kelsall BL, Russell SM, Drago J, Noguchi M, Grinberg A, Bloom ET, et al. Defective lymphoid development in mice lacking expression of the common cytokine receptor gamma chain. Immunity. 1995;2:223–238. doi: 10.1016/1074-7613(95)90047-0. [DOI] [PubMed] [Google Scholar]
- Castro Alves C, Terziyska N, Grunert M, Gundisch S, Graubner U, Quintanilla-Martinez L, Jeremias I. Leukemia-initiating cells of patient-derived acute lymphoblastic leukemia xenografts are sensitive toward TRAIL. Blood. 2012;119:4224–4227. doi: 10.1182/blood-2011-08-370114. [DOI] [PubMed] [Google Scholar]
- Cesano A, Hoxie JA, Lange B, Nowell PC, Bishop J, Santoli D. The severe combined immunodeficient (SCID) mouse as a model for human myeloid leukemias. Oncogene. 1992;7:827–836. [PubMed] [Google Scholar]
- Chao MP, Alizadeh AA, Tang C, Jan M, Weissman-Tsukamoto R, Zhao F, Park CY, Weissman IL, Majeti R. Therapeutic antibody targeting of CD47 eliminates human acute lymphoblastic leukemia. Cancer Res. 2011a;71:1374–1384. doi: 10.1158/0008-5472.CAN-10-2238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chao MP, Alizadeh AA, Tang C, Myklebust JH, Varghese B, Gill S, Jan M, Cha AC, Chan CK, Tan BT, et al. Anti-CD47 antibody synergizes with rituximab to promote phagocytosis and eradicate non-Hodgkin lymphoma. Cell. 2010;142:699–713. doi: 10.1016/j.cell.2010.07.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chao MP, Tang C, Pachynski RK, Chin R, Majeti R, Weissman IL. Extranodal dissemination of non-Hodgkin lymphoma requires CD47 and is inhibited by anti-CD47 antibody therapy. Blood. 2011b;118:4890–4901. doi: 10.1182/blood-2011-02-338020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charley MR, Tharp M, Locker J, Deng JS, Goslen JB, Mauro T, McCoy P, Abell E, Jegasothy B. Establishment of a human cutaneous T-cell lymphoma in C.B-17 SCID mice. J Invest Dermatol. 1990;94:381–384. doi: 10.1111/1523-1747.ep12874500. [DOI] [PubMed] [Google Scholar]
- Chen K, Ahmed S, Adeyi O, Dick JE, Ghanekar A. Human solid tumor xenografts in immunodeficient mice are vulnerable to lymphomagenesis associated with Epstein-Barr virus. PloS one. 2012;7:e39294. doi: 10.1371/journal.pone.0039294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiu PP, Jiang H, Dick JE. Leukemia-initiating cells in human T-lymphoblastic leukemia exhibit glucocorticoid resistance. Blood. 2010;116:5268–5279. doi: 10.1182/blood-2010-06-292300. [DOI] [PubMed] [Google Scholar]
- Clappier E, Gerby B, Sigaux F, Delord M, Touzri F, Hernandez L, Ballerini P, Baruchel A, Pflumio F, Soulier J. Clonal selection in xenografted human T cell acute lymphoblastic leukemia recapitulates gain of malignancy at relapse. J Exp Med. 2011;208:653–661. doi: 10.1084/jem.20110105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colombo F, Baldan F, Mazzucchelli S, Martin-Padura I, Marighetti P, Cattaneo A, Foglieni B, Spreafico M, Guerneri S, Baccarin M, et al. Evidence of distinct tumour-propagating cell populations with different properties in primary human hepatocellular carcinoma. PLoS One. 2011;6:e21369. doi: 10.1371/journal.pone.0021369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cournoyer S, Nyalendo C, Addioui A, Belounis A, Beaunoyer M, Aumont A, Teira P, Duval M, Fernandes K, Fetni R, et al. Genotype analysis of tumor-initiating cells expressing CD133 in neuroblastoma. Genes Chromosomes Cancer. 2012;51:792–804. doi: 10.1002/gcc.21964. [DOI] [PubMed] [Google Scholar]
- Dick JE. Stem cell concepts renew cancer research. Blood. 2008;112:4793–4807. doi: 10.1182/blood-2008-08-077941. [DOI] [PubMed] [Google Scholar]
- Dieter SM, Ball CR, Hoffmann CM, Nowrouzi A, Herbst F, Zavidij O, Abel U, Arens A, Weichert W, Brand K, et al. Distinct types of tumor-initiating cells form human colon cancer tumors and metastases. Cell Stem Cell. 2011;9:357–365. doi: 10.1016/j.stem.2011.08.010. [DOI] [PubMed] [Google Scholar]
- DiSanto JP, Muller W, Guy-Grand D, Fischer A, Rajewsky K. Lymphoid development in mice with a targeted deletion of the interleukin 2 receptor gamma chain. Proceedings of the National Academy of Sciences of the United States of America. 1995;92:377–381. doi: 10.1073/pnas.92.2.377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Distler E, Wolfel C, Kohler S, Nonn M, Kaus N, Schnurer E, Meyer RG, Wehler TC, Huber C, Wolfel T, et al. Acute myeloid leukemia (AML)-reactive cytotoxic T lymphocyte clones rapidly expanded from CD8(+) CD62L((high)+) T cells of healthy donors prevent AML engraftment in NOD/SCID IL2Rgamma(null) mice. Exp Hematol. 2008;36:451–463. doi: 10.1016/j.exphem.2007.12.011. [DOI] [PubMed] [Google Scholar]
- Edris B, Espinosa I, Muhlenberg T, Mikels A, Lee CH, Steigen SE, Zhu S, Montgomery KD, Lazar AJ, Lev D, et al. ROR2 is a novel prognostic biomarker and a potential therapeutic target in leiomyosarcoma and gastrointestinal stromal tumour. J Pathol. 2012a;227:223–233. doi: 10.1002/path.3986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edris B, Weiskopf K, Volkmer AK, Volkmer JP, Willingham SB, Contreras-Trujillo H, Liu J, Majeti R, West RB, Fletcher JA, et al. Antibody therapy targeting the CD47 protein is effective in a model of aggressive metastatic leiomyosarcoma. Proc Natl Acad Sci U S A. 2012b;109:6656–6661. doi: 10.1073/pnas.1121629109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eisterer W, Jiang X, Christ O, Glimm H, Lee KH, Pang E, Lambie K, Shaw G, Holyoake TL, Petzer AL, et al. Different subsets of primary chronic myeloid leukemia stem cells engraft immunodeficient mice and produce a model of the human disease. Leukemia : official journal of the Leukemia Society of America, Leukemia Research Fund, UK. 2005;19:435–441. doi: 10.1038/sj.leu.2403649. [DOI] [PubMed] [Google Scholar]
- Engels B, Rowley DA, Schreiber H. Targeting stroma to treat cancers. Seminars in cancer biology. 2012;22:41–49. doi: 10.1016/j.semcancer.2011.12.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flavell DJ. Modelling human leukemia and lymphoma in severe combined immunodeficient (SCID) mice: practical applications. Hematol Oncol. 1996;14:67–82. doi: 10.1002/(SICI)1099-1069(199606)14:2<67::AID-HON567>3.0.CO;2-U. [DOI] [PubMed] [Google Scholar]
- Fogh J, Fogh JM, Orfeo T. One hundred and twenty-seven cultured human tumor cell lines producing tumors in nude mice. J Natl Cancer Inst. 1977;59:221–226. doi: 10.1093/jnci/59.1.221. [DOI] [PubMed] [Google Scholar]
- Frost P, Moatamed F, Hoang B, Shi Y, Gera J, Yan H, Gibbons J, Lichtenstein A. In vivo antitumor effects of the mTOR inhibitor CCI-779 against human multiple myeloma cells in a xenograft model. Blood. 2004;104:4181–4187. doi: 10.1182/blood-2004-03-1153. [DOI] [PubMed] [Google Scholar]
- Fusetti L, Pruneri G, Gobbi A, Rabascio C, Carboni N, Peccatori F, Martinelli G, Bertolini F. Human myeloid and lymphoid malignancies in the non-obese diabetic/severe combined immunodeficiency mouse model: frequency of apoptotic cells in solid tumors and efficiency and speed of engraftment correlate with vascular endothelial growth factor production. Cancer research. 2000;60:2527–2534. [PubMed] [Google Scholar]
- Gandara DR, Li T, Lara PN, Jr, Mack PC, Kelly K, Miyamoto S, Goodwin N, Beckett L, Redman MW. Algorithm for codevelopment of new drug-predictive biomarker combinations: accounting for inter- and intrapatient tumor heterogeneity. Clin Lung Cancer. 2012;13:321–325. doi: 10.1016/j.cllc.2012.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerber JM, Smith BD, Ngwang B, Zhang H, Vala MS, Morsberger L, Galkin S, Collector MI, Perkins B, Levis MJ, et al. A clinically relevant population of leukemic CD34(+)CD38(-) cells in acute myeloid leukemia. Blood. 2012;119:3571–3577. doi: 10.1182/blood-2011-06-364182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giovanella BC, Fogh J. The nude mouse in cancer research. Adv Cancer Res. 1985;44:69–120. doi: 10.1016/s0065-230x(08)60026-3. [DOI] [PubMed] [Google Scholar]
- Goardon N, Marchi E, Atzberger A, Quek L, Schuh A, Soneji S, Woll P, Mead A, Alford KA, Rout R, et al. Coexistence of LMPP-like and GMP-like leukemia stem cells in acute myeloid leukemia. Cancer Cell. 2011;19:138–152. doi: 10.1016/j.ccr.2010.12.012. [DOI] [PubMed] [Google Scholar]
- Goldstein AS, Huang J, Guo C, Garraway IP, Witte ON. Identification of a cell of origin for human prostate cancer. Science. 2010;329:568–571. doi: 10.1126/science.1189992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greiner DL, Hesselton RA, Shultz LD. SCID mouse models of human stem cell engraftment. Stem Cells. 1998;16:166–177. doi: 10.1002/stem.160166. [DOI] [PubMed] [Google Scholar]
- Hesselton RM, Greiner DL, Mordes JP, Rajan TV, Sullivan JL, Shultz LD. High levels of human peripheral blood mononuclear cell engraftment and enhanced susceptibility to human immunodeficiency virus type 1 infection in NOD/LtSz-scid/scid mice. J Infect Dis. 1995;172:974–982. doi: 10.1093/infdis/172.4.974. [DOI] [PubMed] [Google Scholar]
- Hidalgo M, Bruckheimer E, Rajeshkumar NV, Garrido-Laguna I, De Oliveira E, Rubio-Viqueira B, Strawn S, Wick MJ, Martell J, Sidransky D. A pilot clinical study of treatment guided by personalized tumorgrafts in patients with advanced cancer. Mol Cancer Ther. 2011;10:1311–1316. doi: 10.1158/1535-7163.MCT-11-0233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang SY, Tien HF, Su FH, Hsu SM. Nonirradiated NOD/SCID-human chimeric animal model for primary human multiple myeloma: a potential in vivo culture system. The American journal of pathology. 2004;164:747–756. doi: 10.1016/S0002-9440(10)63162-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hudson WA, Li Q, Le C, Kersey JH. Xenotransplantation of human lymphoid malignancies is optimized in mice with multiple immunologic defects. Leukemia. 1998;12:2029–2033. doi: 10.1038/sj.leu.2401236. [DOI] [PubMed] [Google Scholar]
- Ishikawa F, Saito Y, Shultz LD. Modeling human leukemia using immune-compromised mice. Mouse Models of Human Blood Cancers. 2008:121–132. [Google Scholar]
- Ishikawa F, Yoshida S, Saito Y, Hijikata A, Kitamura H, Tanaka S, Nakamura R, Tanaka T, Tomiyama H, Saito N, et al. Chemotherapy-resistant human AML stem cells home to and engraft within the bone-marrow endosteal region. Nat Biotechnol. 2007;25:1315–1321. doi: 10.1038/nbt1350. [DOI] [PubMed] [Google Scholar]
- Ishizawa K, Rasheed ZA, Karisch R, Wang Q, Kowalski J, Susky E, Pereira K, Karamboulas C, Moghal N, Rajeshkumar NV, et al. Tumor-initiating cells are rare in many human tumors. Cell Stem Cell. 2010;7:279–282. doi: 10.1016/j.stem.2010.08.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Itoh T, Shiota M, Takanashi M, Hojo I, Satoh H, Matsuzawa A, Moriyama T, Watanabe T, Hirai K, Mori S. Engraftment of human non-Hodgkin lymphomas in mice with severe combined immunodeficiency. Cancer. 1993;72:2686–2694. doi: 10.1002/1097-0142(19931101)72:9<2686::aid-cncr2820720927>3.0.co;2-t. [DOI] [PubMed] [Google Scholar]
- Jiang X, Saw KM, Eaves A, Eaves C. Instability of BCR-ABL gene in primary and cultured chronic myeloid leukemia stem cells. Journal of the National Cancer Institute. 2007;99:680–693. doi: 10.1093/jnci/djk150. [DOI] [PubMed] [Google Scholar]
- Kamel-Reid S, Letarte M, Sirard C, Doedens M, Grunberger T, Fulop G, Freedman MH, Phillips RA, Dick JE. A model of human acute lymphoblastic leukemia in immune-deficient SCID mice. Science. 1989;246:1597–1600. doi: 10.1126/science.2595371. [DOI] [PubMed] [Google Scholar]
- Kelly PN, Dakic A, Adams JM, Nutt SL, Strasser A. Tumor growth need not be driven by rare cancer stem cells. Science. 2007;317:337. doi: 10.1126/science.1142596. [DOI] [PubMed] [Google Scholar]
- Khabele D, Fadare O, Liu AY, Wilson AJ, Wass E, Osteen K, Crispens MA. An orthotopic model of platinum-sensitive high grade serous fallopian tube carcinoma. Int J Clin Exp Pathol. 2012;5:37–45. [PMC free article] [PubMed] [Google Scholar]
- Kikushige Y, Ishikawa F, Miyamoto T, Shima T, Urata S, Yoshimoto G, Mori Y, Iino T, Yamauchi T, Eto T, et al. Self-renewing hematopoietic stem cell is the primary target in pathogenesis of human chronic lymphocytic leukemia. Cancer Cell. 2011;20:246–259. doi: 10.1016/j.ccr.2011.06.029. [DOI] [PubMed] [Google Scholar]
- Kim D, Park CY, Medeiros BC, Weissman IL. CD19(-)CD45(low/-)CD38(high)/CD138(+) plasma cells enrich for human tumorigenic myeloma cells. Leukemia. 2012a doi: 10.1038/leu.2012.140. Epub ahead of print. [DOI] [PubMed] [Google Scholar]
- Kim J, Villadsen R, Sorlie T, Fogh L, Gronlund SZ, Fridriksdottir AJ, Kuhn I, Rank F, Wielenga VT, Solvang H, et al. Tumor initiating but differentiated luminal-like breast cancer cells are highly invasive in the absence of basal-like activity. Proc Natl Acad Sci U S A. 2012b;109:6124–6129. doi: 10.1073/pnas.1203203109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim RS, Avivar-Valderas A, Estrada Y, Bragado P, Sosa MS, Aguirre-Ghiso JA, Segall JE. Dormancy signatures and metastasis in estrogen receptor positive and negative breast cancer. PloS one. 2012c;7:e35569. doi: 10.1371/journal.pone.0035569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kong Y, Yoshida S, Saito Y, Doi T, Nagatoshi Y, Fukata M, Saito N, Yang SM, Iwamoto C, Okamura J, et al. CD34+CD38+CD19+ as well as CD34+CD38-CD19+ cells are leukemia-initiating cells with self-renewal capacity in human B-precursor ALL. Leukemia. 2008;22:1207–1213. doi: 10.1038/leu.2008.83. [DOI] [PubMed] [Google Scholar]
- Krumbach R, Schuler J, Hofmann M, Giesemann T, Fiebig HH, Beckers T. Primary resistance to cetuximab in a panel of patient-derived tumour xenograft models: activation of MET as one mechanism for drug resistance. Eur J Cancer. 2011;47:1231–1243. doi: 10.1016/j.ejca.2010.12.019. [DOI] [PubMed] [Google Scholar]
- Kupas V, Weishaupt C, Siepmann D, Kaserer ML, Eickelmann M, Metze D, Luger TA, Beissert S, Loser K. RANK is expressed in metastatic melanoma and highly upregulated on melanoma-initiating cells. J Invest Dermatol. 2011;131:944–955. doi: 10.1038/jid.2010.377. [DOI] [PubMed] [Google Scholar]
- le Viseur C, Hotfilder M, Bomken S, Wilson K, Rottgers S, Schrauder A, Rosemann A, Irving J, Stam RW, Shultz LD, et al. In childhood acute lymphoblastic leukemia, blasts at different stages of immunophenotypic maturation have stem cell properties. Cancer Cell. 2008;14:47–58. doi: 10.1016/j.ccr.2008.05.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li C, Heidt DG, Dalerba P, Burant CF, Zhang L, Adsay V, Wicha M, Clarke MF, Simeone DM. Identification of pancreatic cancer stem cells. Cancer Res. 2007;67:1030–1037. doi: 10.1158/0008-5472.CAN-06-2030. [DOI] [PubMed] [Google Scholar]
- Lin TY, Li YP, Zhang H, Luo J, Goodwin N, Gao T, White R, KLam KS, Pan CX. Tumor-targeting multifunctional micelles for imaging and chemotherapy of advanced bladder cancer. Nanomedicine. 2012 doi: 10.2217/nnm.12.150. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu D, Song L, Wei J, Courtney AN, Gao X, Marinova E, Guo L, Heczey A, Asgharzadeh S, Kim E, et al. IL-15 protects NKT cells from inhibition by tumor-associated macrophages and enhances antimetastatic activity. J Clin Invest. 2012;122:2221–2233. doi: 10.1172/JCI59535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma J, Lin JY, Alloo A, Wilson BJ, Schatton T, Zhan Q, Murphy GF, Waaga-Gasser AM, Gasser M, Stephen Hodi F, et al. Isolation of tumorigenic circulating melanoma cells. Biochem Biophys Res Commun. 2010;402:711–717. doi: 10.1016/j.bbrc.2010.10.091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Majeti R, Chao MP, Alizadeh AA, Pang WW, Jaiswal S, Gibbs KD, Jr, van Rooijen N, Weissman IL. CD47 is an adverse prognostic factor and therapeutic antibody target on human acute myeloid leukemia stem cells. Cell. 2009;138:286–299. doi: 10.1016/j.cell.2009.05.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin-Padura I, Marighetti P, Agliano A, Colombo F, Larzabal L, Redrado M, Bleau AM, Prior C, Bertolini F, Calvo A. Residual dormant cancer stem-cell foci are responsible for tumor relapse after antiangiogenic metronomic therapy in hepatocellular carcinoma xenografts. Lab Invest. 2012;92:952–966. doi: 10.1038/labinvest.2012.65. [DOI] [PubMed] [Google Scholar]
- Mazza JJ. Infectious disease and cancer. WMJ. 2010;109:66–69. [PubMed] [Google Scholar]
- Medyouf H, Gao X, Armstrong F, Gusscott S, Liu Q, Gedman AL, Matherly LH, Schultz KR, Pflumio F, You MJ, et al. Acute T-cell leukemias remain dependent on Notch signaling despite PTEN and INK4A/ARF loss. Blood. 2010;115:1175–1184. doi: 10.1182/blood-2009-04-214718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mirandola L, Yu Y, Jenkins MR, Chiaramonte R, Cobos E, John CM, Chiriva-Internati M. Tracking human multiple myeloma xenografts in NOD-Rag-1/IL-2 receptor gamma chain-null mice with the novel biomarker AKAP-4. BMC Cancer. 2011;11:394. doi: 10.1186/1471-2407-11-394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monsma DJ, Monks NR, Cherba DM, Dylewski D, Eugster E, Jahn H, Srikanth S, Scott SB, Richardson PJ, Everts RE, et al. Genomic characterization of explant tumorgraft models derived from fresh patient tumor tissue. J Transl Med. 2012;10:125. doi: 10.1186/1479-5876-10-125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moon EK, Carpenito C, Sun J, Wang LC, Kapoor V, Predina J, Powell DJ, Jr, Riley JL, June CH, Albelda SM. Expression of a functional CCR2 receptor enhances tumor localization and tumor eradication by retargeted human T cells expressing a mesothelin-specific chimeric antibody receptor. Clin Cancer Res. 2011;17:4719–4730. doi: 10.1158/1078-0432.CCR-11-0351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morisot S, Wayne AS, Bohana-Kashtan O, Kaplan IM, Gocke CD, Hildreth R, Stetler-Stevenson M, Walker RL, Davis S, Meltzer PS, et al. High frequencies of leukemia stem cells in poor-outcome childhood precursor-B acute lymphoblastic leukemias. Leukemia. 2010;24:1859–1866. doi: 10.1038/leu.2010.184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mueller BM, Reisfeld RA. Potential of the scid mouse as a host for human tumors. Cancer Metastasis Rev. 1991;10:193–200. doi: 10.1007/BF00050791. [DOI] [PubMed] [Google Scholar]
- Nakamura M, Suemizu H. Novel metastasis models of human cancer in NOG mice. Current topics in microbiology and immunology. 2008;324:167–177. doi: 10.1007/978-3-540-75647-7_11. [DOI] [PubMed] [Google Scholar]
- O'Brien CA, Pollett A, Gallinger S, Dick JE. A human colon cancer cell capable of initiating tumour growth in immunodeficient mice. Nature. 2007;445:106–110. doi: 10.1038/nature05372. [DOI] [PubMed] [Google Scholar]
- O'Reilly MS, Holmgren L, Chen C, Folkman J. Angiostatin induces and sustains dormancy of human primary tumors in mice. Nat Med. 1996;2:689–692. doi: 10.1038/nm0696-689. [DOI] [PubMed] [Google Scholar]
- Ohbo K, Suda T, Hashiyama M, Mantani A, Ikebe M, Miyakawa K, Moriyama M, Nakamura M, Katsuki M, Takahashi K, et al. Modulation of hematopoiesis in mice with a truncated mutant of the interleukin-2 receptor gamma chain. Blood. 1996;87:956–967. [PubMed] [Google Scholar]
- Ono M, Qiang W, Serna VA, Yin P, Coon JSt, Navarro A, Monsivais D, Kakinuma T, Dyson M, Druschitz S, et al. Role of stem cells in human uterine leiomyoma growth. PloS one. 2012;7:e36935. doi: 10.1371/journal.pone.0036935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patrawala L, Calhoun T, Schneider-Broussard R, Li H, Bhatia B, Tang S, Reilly JG, Chandra D, Zhou J, Claypool K, et al. Highly purified CD44+ prostate cancer cells from xenograft human tumors are enriched in tumorigenic and metastatic progenitor cells. Oncogene. 2006;25:1696–1708. doi: 10.1038/sj.onc.1209327. [DOI] [PubMed] [Google Scholar]
- Pearce DJ, Taussig D, Zibara K, Smith LL, Ridler CM, Preudhomme C, Young BD, Rohatiner AZ, Lister TA, Bonnet D. AML engraftment in the NOD/SCID assay reflects the outcome of AML: implications for our understanding of the heterogeneity of AML. Blood. 2006;107:1166–1173. doi: 10.1182/blood-2005-06-2325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peters M, Jacobs S, Ehlers M, Vollmer P, Mullberg J, Wolf E, Brem G, Meyer zum Buschenfelde KH, Rose-John S. The function of the soluble interleukin 6 (IL-6) receptor in vivo: sensitization of human soluble IL-6 receptor transgenic mice towards IL-6 and prolongation of the plasma half-life of IL-6. The Journal of experimental medicine. 1996;183:1399–1406. doi: 10.1084/jem.183.4.1399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phillips RA, Jewett MA, Gallie BL. Growth of human tumors in immune-deficient scid mice and nude mice. Curr Top Microbiol Immunol. 1989;152:259–263. doi: 10.1007/978-3-642-74974-2_31. [DOI] [PubMed] [Google Scholar]
- Pilarski LM, Belch AR. Clonotypic myeloma cells able to xenograft myeloma to nonobese diabetic severe combined immunodeficient mice copurify with CD34 (+) hematopoietic progenitors. Clin Cancer Res. 2002;8:3198–3204. [PubMed] [Google Scholar]
- Pilarski LM, Hipperson G, Seeberger K, Pruski E, Coupland RW, Belch AR. Myeloma progenitors in the blood of patients with aggressive or minimal disease: engraftment and self-renewal of primary human myeloma in the bone marrow of NOD SCID mice. Blood. 2000;95:1056–1065. [PubMed] [Google Scholar]
- Price JE, Zhang RD. Studies of human breast cancer metastasis using nude mice. Cancer metastasis reviews. 1990;8:285–297. doi: 10.1007/BF00052605. [DOI] [PubMed] [Google Scholar]
- Prince ME, Sivanandan R, Kaczorowski A, Wolf GT, Kaplan MJ, Dalerba P, Weissman IL, Clarke MF, Ailles LE. Identification of a subpopulation of cells with cancer stem cell properties in head and neck squamous cell carcinoma. Proceedings of the National Academy of Sciences of the United States of America. 2007;104:973–978. doi: 10.1073/pnas.0610117104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Provasi E, Genovese P, Lombardo A, Magnani Z, Liu PQ, Reik A, Chu V, Paschon DE, Zhang L, Kuball J, et al. Editing T cell specificity towards leukemia by zinc finger nucleases and lentiviral gene transfer. Nat Med. 2012;18:807–815. doi: 10.1038/nm.2700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quintana E, Piskounova E, Shackleton M, Weinberg D, Eskiocak U, Fullen DR, Johnson TM, Morrison SJ. Human Melanoma Metastasis in NSG Mice Correlates with Clinical Outcome in Patients. Science translational medicine. 2012;4:159ra149. doi: 10.1126/scitranslmed.3004599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quintana E, Shackleton M, Foster HR, Fullen DR, Sabel MS, Johnson TM, Morrison SJ. Phenotypic heterogeneity among tumorigenic melanoma cells from patients that is reversible and not hierarchically organized. Cancer Cell. 2010;18:510–523. doi: 10.1016/j.ccr.2010.10.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quintana E, Shackleton M, Sabel MS, Fullen DR, Johnson TM, Morrison SJ. Efficient tumour formation by single human melanoma cells. Nature. 2008;456:593–598. doi: 10.1038/nature07567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reddy S, Piccione D, Takita H, Bankert RB. Human lung tumor growth established in the lung and subcutaneous tissue of mice with severe combined immunodeficiency. Cancer Res. 1987;47:2456–2460. [PubMed] [Google Scholar]
- Ricci-Vitiani L, Lombardi DG, Pilozzi E, Biffoni M, Todaro M, Peschle C, De Maria R. Identification and expansion of human colon-cancer-initiating cells. Nature. 2007;445:111–115. doi: 10.1038/nature05384. [DOI] [PubMed] [Google Scholar]
- Saito Y, Kitamura H, Hijikata A, Tomizawa-Murasawa M, Tanaka S, Takagi S, Uchida N, Suzuki N, Sone A, Najima Y, et al. Identification of therapeutic targets for quiescent, chemotherapy-resistant human leukemia stem cells. Sci Transl Med. 2010a;2:17ra19. doi: 10.1126/scitranslmed.3000349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saito Y, Uchida N, Tanaka S, Suzuki N, Tomizawa-Murasawa M, Sone A, Najima Y, Takagi S, Aoki Y, Wake A, et al. Induction of cell cycle entry eliminates human leukemia stem cells in a mouse model of AML. Nat Biotechnol. 2010b;28:275–280. doi: 10.1038/nbt.1607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sakakibara T, Xu Y, Bumpers HL, Chen FA, Bankert RB, Arredondo MA, Edge SB, Repasky EA. Growth and metastasis of surgical specimens of human breast carcinomas in SCID mice. Cancer J Sci Am. 1996;2:291–300. [PubMed] [Google Scholar]
- Sanchez PV, Perry RL, Sarry JE, Perl AE, Murphy K, Swider CR, Bagg A, Choi JK, Biegel JA, Danet-Desnoyers G, et al. A robust xenotransplantation model for acute myeloid leukemia. Leukemia. 2009;23:2109–2117. doi: 10.1038/leu.2009.143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sarry JE, Murphy K, Perry R, Sanchez PV, Secreto A, Keefer C, Swider CR, Strzelecki AC, Cavelier C, Recher C, et al. Human acute myelogenous leukemia stem cells are rare and heterogeneous when assayed in NOD/SCID/IL2Rgammac-deficient mice. J Clin Invest. 2011;121:384–395. doi: 10.1172/JCI41495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sebens S, Schafer H. The tumor stroma as mediator of drug resistance--a potential target to improve cancer therapy? Current pharmaceutical biotechnology. 2012;13:2259–2272. doi: 10.2174/138920112802501999. [DOI] [PubMed] [Google Scholar]
- Seitz G, Pfeiffer M, Fuchs J, Warmann SW, Leuschner I, Vokuhl C, Lang P, Handgretinger R, Armeanu-Ebinger S. Establishment of a rhabdomyosarcoma xenograft model in human-adapted mice. Oncol Rep. 2010;24:1067–1072. doi: 10.3892/or.2010.1067. [DOI] [PubMed] [Google Scholar]
- Sharkey FE, Fogh J. Considerations in the use of nude mice for cancer research. Cancer metastasis reviews. 1984;3:341–360. doi: 10.1007/BF00051459. [DOI] [PubMed] [Google Scholar]
- Shmelkov SV, Butler JM, Hooper AT, Hormigo A, Kushner J, Milde T, St Clair R, Baljevic M, White I, Jin DK, et al. CD133 expression is not restricted to stem cells, and both CD133+ and CD133- metastatic colon cancer cells initiate tumors. The Journal of clinical investigation. 2008;118:2111–2120. doi: 10.1172/JCI34401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shultz LD, Brehm MA, Garcia-Martinez JV, Greiner D. Humanized mice for immune system investigation: progress, promise and challenges. Nature Rev Immunol. 2012 doi: 10.1038/nri3311. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shultz LD, Ishikawa F, Greiner DL. Humanized mice in translational biomedical research. Nat Rev Immunol. 2007;7:118–130. doi: 10.1038/nri2017. [DOI] [PubMed] [Google Scholar]
- Shultz LD, Schweitzer PA, Christianson SW, Gott B, Schweitzer IB, Tennent B, McKenna S, Mobraaten L, Rajan TV, Greiner DL, et al. Multiple defects in innate and adaptive immunologic function in NOD/LtSz-scid mice. Journal of immunology. 1995;154:180–191. [PubMed] [Google Scholar]
- Silvestre DC, Pineda JR, Hoffschir F, Studler JM, Mouthon MA, Pflumio F, Junier MP, Chneiweiss H, Boussin FD. Alternative lengthening of telomeres in human glioma stem cells. Stem Cells. 2011;29:440–451. doi: 10.1002/stem.600. [DOI] [PubMed] [Google Scholar]
- Simpson-Abelson MR, Sonnenberg GF, Takita H, Yokota SJ, Conway TF, Jr, Kelleher RJ, Jr, Shultz LD, Barcos M, Bankert RB. Long-term engraftment and expansion of tumor-derived memory T cells following the implantation of non-disrupted pieces of human lung tumor into NOD-scid IL2Rgamma(null) mice. J Immunol. 2008;180:7009–7018. doi: 10.4049/jimmunol.180.10.7009. [DOI] [PubMed] [Google Scholar]
- Singh SK, Clarke ID, Terasaki M, Bonn VE, Hawkins C, Squire J, Dirks PB. Identification of a cancer stem cell in human brain tumors. Cancer Res. 2003;63:5821–5828. [PubMed] [Google Scholar]
- Singh SK, Hawkins C, Clarke ID, Squire JA, Bayani J, Hide T, Henkelman RM, Cusimano MD, Dirks PB. Identification of human brain tumour initiating cells. Nature. 2004;432:396–401. doi: 10.1038/nature03128. [DOI] [PubMed] [Google Scholar]
- Sotiropoulou PA, Perez SA, Gritzapis AD, Baxevanis CN, Papamichail M. Interactions between human mesenchymal stem cells and natural killer cells. Stem Cells. 2006;24:74–85. doi: 10.1634/stemcells.2004-0359. [DOI] [PubMed] [Google Scholar]
- Stewart JM, Shaw PA, Gedye C, Bernardini MQ, Neel BG, Ailles LE. Phenotypic heterogeneity and instability of human ovarian tumor-initiating cells. Proc Natl Acad Sci U S A. 2011;108:6468–6473. doi: 10.1073/pnas.1005529108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subramanian S, Parthasarathy R, Sen S, Boder ET, Discher DE. Species- and cell type-specific interactions between CD47 and human SIRPalpha. Blood. 2006;107:2548–2556. doi: 10.1182/blood-2005-04-1463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takenaka K, Prasolava TK, Wang JC, Mortin-Toth SM, Khalouei S, Gan OI, Dick JE, Danska JS. Polymorphism in Sirpa modulates engraftment of human hematopoietic stem cells. Nat Immunol. 2007;8:1313–1323. doi: 10.1038/ni1527. [DOI] [PubMed] [Google Scholar]
- Taussig DC, Miraki-Moud F, Anjos-Afonso F, Pearce DJ, Allen K, Ridler C, Lillington D, Oakervee H, Cavenagh J, Agrawal SG, et al. Anti-CD38 antibody-mediated clearance of human repopulating cells masks the heterogeneity of leukemia-initiating cells. Blood. 2008;112:568–575. doi: 10.1182/blood-2007-10-118331. [DOI] [PubMed] [Google Scholar]
- Taussig DC, Vargaftig J, Miraki-Moud F, Griessinger E, Sharrock K, Luke T, Lillington D, Oakervee H, Cavenagh J, Agrawal SG, et al. Leukemia-initiating cells from some acute myeloid leukemia patients with mutated nucleophosmin reside in the CD34(-) fraction. Blood. 2010;115:1976–1984. doi: 10.1182/blood-2009-02-206565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor CW, Grogan TM, Lopez MH, Leong SP, Odeleye A, Feo-Zuppardi FJ, Hersh EM. Growth and dissemination of human malignant melanoma cells in mice with severe combined immune deficiency. Lab Invest. 1992;67:130–137. [PubMed] [Google Scholar]
- Tian X, Woll PS, Morris JK, Linehan JL, Kaufman DS. Hematopoietic engraftment of human embryonic stem cell-derived cells is regulated by recipient innate immunity. Stem Cells. 2006;24:1370–1380. doi: 10.1634/stemcells.2005-0340. [DOI] [PubMed] [Google Scholar]
- Uckun FM. Severe combined immunodeficient mouse models of human leukemia. Blood. 1996;88:1135–1146. [PubMed] [Google Scholar]
- Urbanska K, Lanitis E, Poussin M, Lynn RC, Gavin BP, Kelderman S, Yu J, Scholler N, Powell DJ., Jr A universal strategy for adoptive immunotherapy of cancer through use of a novel T-cell antigen receptor. Cancer Res. 2012;72:1844–1852. doi: 10.1158/0008-5472.CAN-11-3890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Utama FE, LeBaron MJ, Neilson LM, Sultan AS, Parlow AF, Wagner KU, Rui H. Human prolactin receptors are insensitive to mouse prolactin: implications for xenotransplant modeling of human breast cancer in mice. J Endocrinol. 2006;188:589–601. doi: 10.1677/joe.1.06560. [DOI] [PubMed] [Google Scholar]
- Valdez KE, Fan F, Smith W, Allred DC, Medina D, Behbod F. Human primary ductal carcinoma in situ (DCIS) subtype-specific pathology is preserved in a mouse intraductal (MIND) xenograft model. J Pathol. 2011;225:565–573. doi: 10.1002/path.2969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vargaftig J, Taussig DC, Griessinger E, Anjos-Afonso F, Lister TA, Cavenagh J, Oakervee H, Gribben J, Bonnet D. Frequency of leukemic initiating cells does not depend on the xenotransplantation model used. Leukemia. 2012;26:858–860. doi: 10.1038/leu.2011.250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vatakis DN, Koya RC, Nixon CC, Wei L, Kim SG, Avancena P, Bristol G, Baltimore D, Kohn DB, Ribas A, et al. Antitumor activity from antigen-specific CD8 T cells generated in vivo from genetically engineered human hematopoietic stem cells. Proc Natl Acad Sci U S A. 2011;108:E1408–1416. doi: 10.1073/pnas.1115050108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Visonneau S, Cesano A, Torosian MH, Miller EJ, Santoli D. Growth characteristics and metastatic properties of human breast cancer xenografts in immunodeficient mice. Am J Pathol. 1998;152:1299–1311. [PMC free article] [PubMed] [Google Scholar]
- Visvader JE. Cells of origin in cancer. Nature. 2011;469:314–322. doi: 10.1038/nature09781. [DOI] [PubMed] [Google Scholar]
- Wang JC, Lapidot T, Cashman JD, Doedens M, Addy L, Sutherland DR, Nayar R, Laraya P, Minden M, Keating A, et al. High level engraftment of NOD/SCID mice by primitive normal and leukemic hematopoietic cells from patients with chronic myeloid leukemia in chronic phase. Blood. 1998;91:2406–2414. [PubMed] [Google Scholar]
- Wege AK, Ernst W, Eckl J, Frankenberger B, Vollmann-Zwerenz A, Mannel DN, Ortmann O, Kroemer A, Brockhoff G. Humanized tumor mice--a new model to study and manipulate the immune response in advanced cancer therapy. Int J Cancer. 2011;129:2194–2206. doi: 10.1002/ijc.26159. [DOI] [PubMed] [Google Scholar]
- Williams SS, Alosco TR, Croy BA, Bankert RB. The study of human neoplastic disease in severe combined immunodeficient mice. Laboratory animal science. 1993;43:139–146. [PubMed] [Google Scholar]
- Willingham SB, Volkmer JP, Gentles AJ, Sahoo D, Dalerba P, Mitra SS, Wang J, Contreras-Trujillo H, Martin R, Cohen JD, et al. The CD47-signal regulatory protein alpha (SIRPa) interaction is a therapeutic target for human solid tumors. Proc Natl Acad Sci U S A. 2012;109:6662–6667. doi: 10.1073/pnas.1121623109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wong K, Brenneman F, Chesney A, Spaner DE, Gorczynski RM. Soluble CD200 is critical to engraft Chronic Lymphocytic Leukemia cells in immunocompromised mice. Cancer Res. 2012 Aug 8; doi: 10.1158/0008-5472.CAN-12-1390. Epub ahead of print. [DOI] [PubMed] [Google Scholar]
- Wunderlich M, Chou FS, Link KA, Mizukawa B, Perry RL, Carroll M, Mulloy JC. AML xenograft efficiency is significantly improved in NOD/SCID-IL2RG mice constitutively expressing human SCF, GM-CSF and IL-3. Leukemia. 2010;24:1785–1788. doi: 10.1038/leu.2010.158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xue SA, Gao L, Hart D, Gillmore R, Qasim W, Thrasher A, Apperley J, Engels B, Uckert W, Morris E, et al. Elimination of human leukemia cells in NOD/SCID mice by WT1-TCR gene-transduced human T cells. Blood. 2005;106:3062–3067. doi: 10.1182/blood-2005-01-0146. [DOI] [PubMed] [Google Scholar]
- Zhang B, Min L, McDonald T, Holyoake T, Moon RT, Campana D, Shultz L, Bhatia R. Microenvironmental protection of CML stem and progenitor cells from tyrosine kinase inhibitors through N-Cadherin and Wnt-beta-Catenin signaling. Blood. 2012a doi: 10.1182/blood-2012-02-412890. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang B, Strauss AC, Chu S, Li M, Ho Y, Shiang KD, Snyder DS, Huettner CS, Shultz L, Holyoake T, et al. Effective targeting of quiescent chronic myelogenous leukemia stem cells by histone deacetylase inhibitors in combination with imatinib mesylate. Cancer Cell. 2010;17:427–442. doi: 10.1016/j.ccr.2010.03.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang WC, Shyh-Chang N, Yang H, Rai A, Umashankar S, Ma S, Soh BS, Sun LL, Tai BC, Nga ME, et al. Glycine decarboxylase activity drives non-small cell lung cancer tumor-initiating cells and tumorigenesis. Cell. 2012b;148:259–272. doi: 10.1016/j.cell.2011.11.050. [DOI] [PubMed] [Google Scholar]
- Zhang X, Cruz FD, Terry M, Remotti F, Matushansky I. Terminal differentiation and loss of tumorigenicity of human cancers via pluripotency-based reprogramming. Oncogene. 2012c doi: 10.1038/onc.2012.237. 2012 Jul 9. [Epub ahead of print] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zorzoli A, Di Carlo E, Cocco C, Ognio E, Ribatti D, Ferretti E, Dufour C, Locatelli F, Montagna D, Airoldi I. Interleukin-27 inhibits the growth of pediatric acute myeloid leukemia in NOD/SCID/Il2rg-/- mice. Clin Cancer Res. 2012;18:1630–1640. doi: 10.1158/1078-0432.CCR-11-2432. [DOI] [PubMed] [Google Scholar]