

Abstract
We have modified and optimized the technique of organotypic slice culture in order to study the mechanisms regulating growth and pattern formation in regenerating axolotl limb blastemas. Blastema cells maintain many of the behaviors that are characteristic of blastemas in vivo when cultured as slices in vitro, including rates of proliferation that are comparable to what has been reported in vivo. Because the blastema slices can be cultured in basal medium without fetal bovine serum, it was possible to test the response of blastema cells to signaling molecules present in serum, as well as those produced by nerves. We also were able to investigate the response of blastema cells to experimentally regulated changes in BMP signaling. Blastema cells responded to all of these signals by increasing the rate of proliferation and the level of expression of the blastema marker gene, Prrx-1. The organotypic slice culture model provides the opportunity to identify and characterize the spatial and temporal co-regulation of pathways in order to induce and enhance a regenerative response.
Introduction
Research on regenerating body parts has focused on identifying the signaling pathways involved in initiating and regulating this fascinating and biologically important process. Much of that research has focused on regenerating salamander limbs, and has involved either describing the process of regeneration, or inhibiting regeneration (e.g. by denervating the limb), and then attempting to rescue regeneration. In recent years, the gain-of-function assay for limb regeneration (the Accessory Limb Model, ALM) has identified a number of signals and pathways that are necessary and sufficient for induction of blastema formation and subsequent regeneration of an ectopic limb [1–3]. These experimental approaches have provided insights into the mechanisms of regeneration; however, their utility has been limited by the fact that they all involve regeneration in vivo. Thus it is not possible to control the spatial and temporal activation or inhibition of specific pathways without the variability associated with working on a live animal. Attempts to deal with this complexity have involved using in vitro culture techniques with either monolayer cultures of dissociated blastema cells, or with blastema explant cultures [4–7]. These studies have been of limited utility because of the limited viability of explants, and the loss of normal blastema cell behaviors (e.g. proliferation) after enzymatic dissociation. To address this challenge, we have adapted organotypic slice culture techniques that have been used widely and successfully in the field of neurobiology [8], and have investigated the response of blastema cells to nerve signals and to BMP2.
Because regeneration is a stepwise process [1,9,10], there are critical signaling events at each step that are required for progression to the next step. Consequently, if the appropriate signaling does not occur at any one of the steps, regeneration will fail to occur [1,9]. Thus each step represents a possible barrier to regeneration, and the failure to overcome at least one of these barriers likely accounts for regenerative failure in mammals [9]. Identifying the steps and discovering how to provide the appropriate signals to progress beyond each step will be required in order to induce regeneration in humans. One or more of the early signals involved in the initiation of regeneration are provided by nerves [1,11–13], which have been the focus of research efforts for decades.
The nerve has long been recognized to be important in regeneration [11–18]. Historically, most experimental work on the role of the nerve has involved in vivo studies designed to rescue regeneration of amputated limbs that have been denervated. Limbs that have been denervated fail to initiate regeneration, or fail to progress through the early stages of regeneration [12] to the point where they become independent of the nerve [16]. If the nerves are allowed to regenerate to a point where the supply of nerves exceeds a threshold level, reinjured limbs will be able to regenerate [14,19,20]. Thus attempts to identify the pro-regenerative signals provided by nerves have focused on the later stages of regeneration when the regeneration blastema transitions to becoming independent of nerve signals [21], or on the stage when the regenerating nerve reaches the threshold for the required nerve signaling [7,20,22]. Although denervation experiments have repeatedly demonstrated that a nerve is required (loss-of-function), they have not identified the signals that are required to initiate regeneration and allow for progression through to the late stages at which regeneration is no longer nerve-dependent. The variability associated with in vivo experiments (e.g. the age of the animal which affects the timing of the regenerative response, and the degree of nerve regeneration and reinnervation of limbs following denervation) is a challenge to understanding the precise regulation of the nerve signals that are sufficient for inducing regeneration.
To address the challenges associated with studies of regeneration in vivo, there have been repeated attempts to isolate and study blastema cells in vitro. As with in vivo studies, these experiments have provided insights into factors that affect the behavior of blastema cells, particularly with regards to proliferation [4,7,23–25]. Although blastema tissue can be dissociated and blastema cells can be maintained in monolayer culture, the cells quickly lose properties associated with blastema cells in vivo. Cultured blastema cells typically have low rate of proliferation and are highly variable; e.g. growth fractions ranging from 0.2–10% [7,26,27] compared to proliferation in vivo; e.g. labeling index of 20% to 40% [13,28–30]. The difficulty in maintaining normal behavior of blastema cells in vitro has been attributed to their sensitivity to culture conditions such as osmolarity and temperature [5,26,31,32]. Short-term culturing of blastema explants has been reported in studies pertaining to the effect of hormones and growth factors on protein synthesis and cell division [4,23,26,33,34]. The utility of this approach is limited by variability associated with the size of the explants, as well as differences in the viability of cells in the center of the explants compared to cells at the periphery. Finally, dissociated and re-aggregated blastema cells have been cultured in fibrin clots in which they maintain the property of producing high levels of matrix degrading enzymes; however, after a week they lose the ability to signal to other blastema cells in order to induce supernumerary limb patterns when grafted back into a blastema in vivo [35].
In order to overcome the challenges of studying the role of the nerve during regeneration in vivo, we initiated a systematic assessment of techniques for culturing blastema cells. As reported by others, we observed that the proliferation rates of dissociated blastema cells quickly decreased when the cells were cultured on a range of substrates and culture media, and the cultures could not be maintained and passaged over an extended period of time (data not presented). We therefore reasoned that the behavior of blastema cells in vivo was dependent on cell-cell and cell-matrix interactions established during blastema formation. In order to maintain the in vivo organization of blastema cells and matrix, we optimized the technique of organotypic slice culture (OSC) that is commonly used in neurobiology. The technique uses a vibrating blade (vibratome) to make serial sections of unfixed blastemas that are of uniform thickness and maintain the original tissue architecture. Multiple sections can be collected from the same blastema, and thus different culture conditions can be tested experimentally without the variability associated with comparing blastema tissues from different animals (e.g. variability in stages of blastemas). We report here that blastema cells in OSC maintain in vivo levels of proliferation and tissue architecture, and are responsive to signals from co-cultured dorsal root ganglia explants (DRG), and to exogenous BMP2.
Materials and Methods
Ethics statement
This study was carried out in accordance with the recommendations in the Guide for Care and Use of Laboratory Animals of the National Institutes of Health. The experimental work was conducted in accordance with procedures approved by the Institutional Animal Care and Use Committee of the University of California Irvine (IACUC protocol #2007–2705).
Animal procedures and collection of DRG and blastemas
Axolotls (Ambystoma mexicanum) measuring 15–20 cm from snout to tail tip that were spawned at UC Irvine or the Ambystoma Genetic Stock Center at the University of Kentucky were housed on a 12-hour light/dark cycle and fed ad libitum. Axolotls were anesthetized using 0.1% ethyl 3-aminobenzoate methanesulfonate salt (MS222, Sigma) at pH 7.0. Limb regeneration leading to formation of medium-late bud stage blastemas was induced by amputation at the level of the mid-humerus.
Details for the culture of dorsal root ganglia (DRG) have been reported previously [36]. Briefly DRG were collected post-euthanasia by surgically removing the spinal nerves that innervated either the forelimb (spinal nerves 3, 4 and 5) or the hind limb (spinal nerves 15, 16 and 17). DRG were cultured individually in 12 well Nunc nunclon plates with 60% L-15, 5% FBS, 1% Insulin, Transferrin, and Selenium (ITS), and gentamicin/amphotericin B (Sigma). Each DRG was attached to the bottom of the culture well by embedding it in a small drop (about 3 μl) of growth factor reduced matrigel (BD Biosciences). Only DRG explants that exhibited neurite outgrowth within the first 24 hours of culture were used for subsequent experiments.
Organotypic Slice Culture (OSC)
Medium-late bud stage blastemas (mesenchyme and epithelium) were removed surgically from animals, embedded in a 4% agarose gel, and sectioned at a thickness of 250 micrometers using a vibratome. Blastema slices were then cultured on transwell membrane inserts (3.0μm pore) with 6 well plates (BD Falcon) in 60% L-15 media containing 1% Gentamicin, 1% penicillin/streptomycin/amphotericin B, and 1% ITS media supplement (Sigma) in a refrigerated incubator at 19°C. Blastema slices and blastema slice/DRG co-culture were adhered to the transwell membrane with 5μL of growth factor reduced matrigel (BD). Blastema slices receiving additional treatments were cultured in media containing one of the following supplements: 5% Fetal Bovine Serum (FBS, Atlanta Biologicals); 1ng/mL, 10ng/mL, or 100ng/mL recombinant human Bone Morphogenetic Protein 2 (BMP2, Sigma); or 5nM, 50nM, 500nM of the BMP signaling inhibitor, LDN-193189 hydrochloride (Abcam). Half the volume of the culture media was changed every other day. Slices were cultured for 3, 5, or 7 days after being explanted.
Analysis of cell proliferation and viability
To analyze cell proliferation we used a Click-It EdU labeling kit (Life Technologies) according to the manufacturer’s protocol. The labeling index (% of cells that were in the S phase of the cell cycle) was determined by incubating slice cultures in 80μM of EdU for five hours, which corresponds to approximately 10% of the total cell cycle for blastema cells [37]. The growth fraction (% cells that were proliferating) was determined by incubating the slice cultures in EdU for 80 hours, which corresponds to approximately 1.5 cell cycles (1 cell cycle = 53 hours) [13,37]. After EdU labeling, the slice cultures were washed in PBS, fixed in 4% paraformaldehyde, and sectioned at 5μm. EdU labeled cells were then visualized using the Click-It-EdU alexa-fluor 488 or alexa-fluor 594 kit (Life Technologies), and were counted using the cell counter plug-in for ImageJ. In order to compare the labeling index of blastema cells in response to experimental treatments as compared to the in vivo labeling index, we first injected animals with BrdU two hours before removing the blastema for OCS in vitro. The incorporation of BrdU was visualized by using immunohistochemistry. The total number of cells was visualized by staining with DAPI.
To determine the relative number of viable and dying cells, we used a TUNEL assay kit (Roche) to identify cells that were undergoing cell death. Slides were processed as indicated above and then processed in accordance with the manufacturer’s protocol. For each experiment, the sample size (N) represents the number of biological replicates (independent experiments). For each sample, cells were counted in a total of three sections that spanned the width or thickness of the slice (anterior to posterior or ventral to dorsal). Standard deviation and standard error were calculated using Excel. An F-test was performed to determine equal or unequal variance (i.e. F-value >0.05 or <0.05, respectively). P-values were then calculated using a t-test assuming equal or unequal variance based on the F-value.
Immunohistochemistry
Blastema slices were fixed for three hours at room temperature in 4% paraformaldehyde, dehydrated in graded alcohol followed by xylene, embedded in paraplast, and sectioned at 5μm. For immunohistochemistry sections were de-paraffinized, rehydrated in TBST, and incubated with anti-acetylated α–tubulin (Abcam, catalog # T7451, diluted 1:250) and anti-RT-97 (DSHB, diluted 1:250) overnight at 4°C. Sections were then washed with TBST, incubated with anti-mouse 594 (Abcam, diluted 1:250) for 4 hours, washed with TBST, and mounted with ProLong Gold Antifade mounting medium with DAPI. Images were obtained using a LSM780 confocal microscope.
The intra-nuclear distribution and translocation of phospho-Smad 1/5/8 was quantified by staining sections with an anti-phospho-Smad 1/5/8 antibody (Millipore, Catalog # AB3848, Lot # 2390361, diluted 1:75). Sections were de-paraffinized and rehydrated in TBST as described above. Heat-induced epitope retrieval was performed at 100°C for 1 hour in Tris/EDTA (pH 9.0). Sections were incubated with anti-phospho-Smad 1/5/8 overnight at 4°C, washed with TBST, and incubated with anti-rabbit 488 (Abcam, diluted 1:250). Twelve regions containing 100–200 mesenchymal cells from three different OSC (N = 3) from each condition tested were selected and analyzed in a double blind manner. The corrected nuclear fluorescence was calculated in order to normalize the intensity of nuclear staining for variation in the area of the nuclei being analyzed as well as for the background fluorescence [38].
Quantitative PCR
Blastema slice RNA was extracted using Trizol (Invitrogen) in conjunction with the Nucleospin RNA XS kit (Macherey-Nagel). Samples were processed to generate cDNA using the Transcriptor First Strand cDNA Kit (Roche). Real-time PCR was performed using a LightCycler 480 (Roche) and the resulting data were analyzed using the Pfaffl method [39]. Sequences for the primers for qPCR (Sigma) were based on primers published previously [40,41].
GAPDH FWD: GACGCTGGTGCAGGCATTGCC
GAPDH REV: ACCATCAGGTCCACAACACGCTGAC
Prrx1 FWD: GGCGAAAGTTTGCTCTTCGG
Prrx1 REV: GGCGAAACTTTGCTCTTCGG
Results
Organotypic slice cultures of axolotl blastemas remain viable for several days
OSC allowed for the culture of multiple sections of uniform dimensions from a single blastema (Fig 1). In response to limb amputation, a blastema (medium-late bud stage) formed in about 10–12 days (Step 1, Fig 1A). The blastema was surgically removed (Step 2), embedded in gelatin and sectioned using a vibratome that resulted in uniform 250μm thick longitudinal sections (Step 3). Depending on the size of the blastema, a typical mid bud blastema from an adult axolotl (10-15cm) yielded 3–4 slices that were similar in shape and size. Adjacent blastema slices maintained the tissue architecture of in vivo blastemas (i.e. mesenchyme surrounded by a wound epithelium) (Fig 1B and 1C). The blastema slices were then cultured under various experimental and control conditions (Step 4).
Fig 1. Organotypic slice culture model for axolotl blastemas.

(A1) The limb of an axolotl (Ambystoma mexicanum) was amputated and allowed to regenerate a medium-late bud stage. (A2) The blastema was surgically removed from the animal, (A3) was sectioned using a vibratome (A3) and cultured (A4, B, C). Depending on the size of the blastema, a typical blastema yielded 3–4 slices that appeared similar when cultured (B and C). A region of pooled blood cells in the apical region of adjacent slices is indicated by the arrows in (B and C). The proximal (P) to distal (D) orientation of the sections is indicated. Scale bars = 500μm.
Blastema slices appeared healthy and the cells continued to proliferate in vitro over an extended period of time (Fig 2). After 10 days in culture, the slices maintained a normal morphology with a thickened epithelium surrounding the blastema mesenchyme (Fig 2A). The cells within the slice appeared healthy and regions of high cell density corresponding to pre-chondrogenic condensations were evident in some sections (Fig 2A, arrows). Although the appearance of an OSC blastema was similar to a blastema that was integrated into the amputated limb (e.g. there was an apical epithelium that covered the blastema mesenchyme), there also were differences. At the proximal boundary of the slice, the epidermis began to migrate over the free surface that was created when the blastema was surgically removed from the animal. This phenomenon referred to as “epiboly”, has been described previously in whole-mount explants of amphibian blastemas and limb buds [42,43]. Many of the cells of the blastema mesenchyme incorporated EdU after 10 days of culture, although cells in the regions of high cell density (visualized by DAPI staining of the nuclei) did not (Fig 2B and 2D). The keratinocytes of the apical epithelium (Apical Epithelial Cap, AEC) in this experiment (cultured with 5% FBS) did not incorporate EdU (Fig 2B), which was consistent with the observation that AEC cells withdraw from the cell cycle in vivo [44,45]. Better understanding how the proliferative response of AEC keratinocytes, particularly the basal keratinocytes [2] is regulated in organotypic slice cultures is a goal of future experiments.
Fig 2. Organotypic blastema slices survive and proliferate in vitro.

(A) Hematoxylin and eosin stained section of a blastema slice that had been cultured for 10 days in the presence of 5% FBS. Regions of pre-cartilage condensations are evident (arrows). The blastema epithelium has begun to migrate over the proximal cut end. (B) EdU labeling of a blastema slice after 10 days in vitro. EdU positive nuclei (green) are present in the mesenchyme but absent from the epithelium. (C) A bright field image of a blastema slice (upper right) and DRG (lower left) after 10 days of co-culture. (D) Immunofluorescence of the blastema slice/DRG co-culture illustrated in (C). Proliferating cells in the slices (green) are restricted to the mesenchyme, nuclei are stained with DAPI (blue), and neurofilaments are labeled with RT97 (red). The proximal (P) to distal (D) orientation of the sections is indicated. Scale bars = 500μm.
As reported previously, DRG can be isolated and co-cultured with blastema explants in order to study the interaction between nerves and blastema cells [36]. To investigate the response of OSC blastema cells to signaling from nerves, blastema slices were co-cultured with DRG (Fig 2C and 2D). Both OSC blastema slices and DRG explants appeared healthy after an extended period of time (10 day cultures illustrated in Fig 2C and 2D). As with blastema cultures without DRG explants, many of the mesenchymal cells not associated with regions of high cell density incorporated EdU (Fig 2D, green). As with blastema slices cultured without DRG, the keratinocytes of the AEC did not incorporate EdU. As reported previously [36], the cultured DRG contained large numbers of regenerating neurons with RT97-positive neurofilaments (Fig 2D, red).
As noted above, blastema slice cultures appeared healthy for at least 10 days in culture. During this period, some pyknotic nuclei were observed in cells along the proximal edge of the explant that was created when the blastema was surgically removed from the animal. TUNEL staining was relatively low, and was not significantly different at a given time point for slices cultured with or without FBS or co-cultured DRG (Fig 3). As with the distribution of pyknotic nuclei, most of the TUNEL-positive nuclei were restricted to the region immediately adjacent to the proximal boundary of the explant. After five days in vitro, TUNEL staining remained about the same as observed at three days, and did not differ depending on the culture conditions. After seven days in vitro, TUNEL staining was more variable and the percentage of TUNEL-positive nuclei was higher in slices cultured in basal medium and 5%FBS; however, this increase was not significant. Based on these data, we did not culture slices for longer than seven days in vitro so as to minimize variability associated with cell death. This time period is comparable to the window for experimental work using OSC in neurobiology [8].
Fig 3. Cell death in blastema slices after culture for 3, 5, and 7 days.

The percentage of TUNEL positive cells was not significantly different between treatments at any of the time points, or between the three time points. Error bars represent one Standard Deviation of the Mean, and P-values were determined by t-test with 2 tails assuming unequal variances. The number of biological replicates for 3, 5, and 7 days were as follows: Basal medium: 5, 5, and 3, 5%FBS: 4, 4, and 5, DRG co-culture: 3, 3, and 3.
Expression of Prrx-1 (Paired-related Homeobox 1) in OSC blastema mesenchymal cells was maintained and increased in vitro (Fig 4). Prrx-1 is a transcription factor that is expressed at high levels in developing and regenerating axolotl limb mesenchymal cells [41], but is expressed at low levels that are not detected by in situ hybridization in uninjured skin. Expression of Prrx-1 in the blastema is regulated by interactions between the wound epithelium and the nerve [41], and thus it is a marker for regenerating blastema cells [3,41,46]. As reported previously [47], Prrx-1 expression was detected at low levels by qPCR in uninjured skin (Fig 4). Expression in blastema sections prior to being culture was significantly upregulated an average of 6-fold, which is comparable to the increased level of expression in vivo when comparing blastemas to uninjured skin [3]. Blastema slices cultured in basal medium for 7 days expressed Prrx-1 at a 23-fold higher level relative to uninjured skin (about a 4-fold increase during the culture period. Since Prrx-1 expression is restricted to the distal tip of blastemas in vivo, this increase in expression in vitro could be a result of increased numbers of cells in the slices being induced to express this gene rather than an increase in the level of expression in the distal blastema cells. Better understanding the mechanisms of Prrx-1 expression during regeneration is a goal of future experiments using OSC. Taken together, these data indicate that blastema slices cultured in basal media are viable and maintain expression of a gene that is characteristic of regenerating blastema cells.
Fig 4. Prrx-1 expression in blastema slices.

Fold change in Prrx-1 levels after seven days of culture under different culture conditions. The value for “Uninjured Skin” was determined for samples of skin that had not been cultured. The value for “Pre-culture Slice” was determined for blastema slices that had not been cultured. Error bars represent S.E.M., and P-values were determined by t-test with 2 tails assuming unequal variances. The number of biological replicates for the conditions tested were as follows: Pre-culture slice: N = 5, Basal medium: N = 7, 5%FBS: N = 5, Pre-conditioned DRG: N = 5, 100ng/mL BMP: N = 3. Each biological replicate consisted of four technical replicates. Asterisk (*) = P<0.05.
Organotypic slice cultured blastema cells responded to FBS and co-culture with DRG
Because blastema cells remained viable when cultured in a minimal basal medium, it was possible to assay for their response to changes in the culture environment. As reported above, there was no significant change in the percentage of TUNEL-positive cells in basal medium when compared to cultures with added FBS or with co-cultured DRG. Expression of Prrx-1 was maintained, and increased over time, in slices cultured in basal medium (Fig 4). Addition of 5% FBS to the basal medium induced a significant increase in the level of Prrx-1 expression after seven days in culture (43-fold relative to uninjured skin; 7-fold relative to an uncultured blastema; nearly 2-fold relative to blastema slices cultured in basal medium). While there was an increase in Prrx-1 expression in slices co-cultured with a DRG compared to slices cultured in a basal medium (1.6-fold), the difference was not statistically significant.
In contrast to Prrx-1 expression that increased during culture in basal medium, proliferation of the blastema mesenchymal cells in basal medium decreased during the initial three days in culture and remained at a relatively low level over the next four days (Fig 5A). The labeling index (LI) of the mesenchymal cells in blastema slices prior to amputation (equivalent to in vivo) was about 23%, which was comparable to values reported previously for blastema cells in vivo [13,28–30]. In basal medium the LI had decreased to about 7% after three days in vitro, and remained constant at this level until the end of the experiment (seven days). This decrease in proliferation was comparable to the rate of proliferation observed in vivo in denervated newt limbs [48] and regressing axolotl ectopic blastemas [1].
Fig 5. Labeling index and growth fraction in blastema slices.
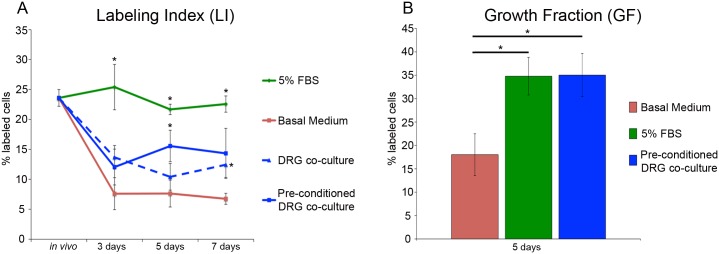
A) Labeling index of blastema mesenchymal cells in blastema slices over a period of seven days in culture under different culture conditions as indicated. Labeling period was for five hours. The value for “in vivo” was determined by injecting animals with BrdU two hours prior to collecting the blastema for sectioning and counting of labeled cells without culture. Error bars represent S.E.M., and P-values were determined by T-test with 2 tails assuming unequal variances. The number of biological replicates for in vivo: N = 22. The number of biological replicates for the following conditions on 3,5, and 7 days were as follows: Basal medium: N = 6, 8, and 9. 5%FBS: N = 7, 7, and 7. DRG co-culture: N = 3, 5, and 6. Pre-conditioned DRG: N = 5, 5, and 4. Asterisk (*) = P<0.05 when compared to the LI in basal medium at equivalent time point. B) Growth Fraction of blastema mesenchymal cells in blastema slices after five days of culture under different culture conditions as indicated. Labeling period was for 80 hours. Error bars represent S.E.M., and P-values were determined by t-test with 2 tails assuming unequal variances. N = 4. Asterisk (*) = P<0.04.
The rate of proliferation was maintained at in vivo levels in cultures that were supplemented with 5% FBS (Fig 5A). The proliferation rate in slices that were co-cultured with a DRG removed from animals the same day as initial culturing of OSC was higher (not statistically significant) than the rate for slices cultured in basal medium during the first five days of culture. After seven days of culture, the LI in blastema/DRG co-cultures was about 2-fold higher compared to the basal medium cultures (p = 0.015).
Gene expression data suggest that DRG might undergo a refractory period or injury response that changes the expression of genes involved with growth and proliferation of blastema cells [36]. It also has been shown that the growth response to DRG was greatest when DRG were surgically grafted 2–3 days prior to whole explant culture of blastemas rather than concurrently, supporting the idea that DRG might require a period of recovery before the growth promoting effect of the nerve is restored [4]. In light of these observations, we hypothesized that DRG might require a recovery period prior to co-culture with DRG in order to restore the growth promoting effect of the nerve. Because previous work has shown that there is a large change in gene expression when comparing DRG in vitro for 5 days to day 0 DRG [36], we decided to pre-condition DRG for 5 days prior to co-culture with OSC. Pre-conditioning DRG led to a higher proliferation rate in OSC at days 3, 5, and 7 when compared to OSC cultured in basal medium (Fig 5A). While the proliferation rate was higher at all three time points, it was at its highest and most significant at day five, when the LI was approximately 15% (Fig 5A).
The decrease in LI in basal medium cultures likely was not a consequence of cell death since the percentage of TUNEL-positive nuclei was not significantly different between the different culture conditions or between the various time points of the experiment (Fig 3). However, it appeared that a significant number of cells withdrew from the cell cycle when cultured in basal medium (Fig 5B), which could account in part for the decrease in the LI. The growth fraction (80 hours of continuous EdU labeling which corresponds to 1.5 cell cycles) of blastema cells cultured in basal medium was significantly lower (about 50% decrease) that that of cells cultured in medium with 5% FBS or co-cultured with pre-conditioned DRG in basal medium (both about 35%) (Fig 5B). Although the growth fraction (about 18%) decreased in the basal medium cultures without FBS or co-cultured with DRG, it was considerably higher and more consistent that what has been reported for monolayer cultures of blastema cells (growth fraction of 0.1%- 9% after 96 hours of labeling [7].
Organotypic slice cultured blastema cells respond to human BMP2
A major advantage of OSC is that the cells remain viable when cultured in minimal basal medium, and it therefore is possible to test the affects of activating specific signaling pathways. Our previous analyses of the transcriptional response of cultured DRG identified BMP2 as a signaling molecule whose expression was specifically upregulated in response to co-culture with a blastema explant [36]. We therefore hypothesized that a regenerating nerve produces BMP2 in response to interacting with blastema mesenchymal cells, and that BMP2 in turns regulates the behavior of the blastema cells.
To test whether cultured blastemas cell respond to exogenous BMP2, we added recombinant human BMP2 to the basal medium of OSC blastema cells at concentrations that span our best estimate for the in vivo concentration of BMP2 [49–53]. We first determined that the axolotl blastema cells responded to the addition of non-axolotl BMP2 by quantifying the changes in the level of phospho-Smad 1/5/8 (p-Smad 1/5/8) immunofluorescence within the nucleus of OSC blastema cells (Fig 6). Treatment with BMP2 at all three concentrations tested (1–100 ng/ml) induced a significant increase (nearly double at 100 ng/ml) in the amount of nuclear-localized p-Smad 1/5/8 relative to basal medium, indicating that human BMP2 activated the canonical BMP signaling pathway in axolotl blastema cells.
Fig 6. Nuclear P-Smad 1/5/8 fluorescence in response to BMP2.

Nuclear Phospho-Smad 1/5/8 staining in blastema mesenchymal cells in blastema slices cultured in the presence or absence of exogenous human BMP2 in amounts as indicated. The corrected nuclear fluorescence was calculated in order to normalize the intensity of nuclear staining for variation in the area of the nuclei being analyzed as well as for the background fluorescence [40]. Error bars represent S.E.M., and P-values were determined by t-test with 2 tails assuming unequal variances.
OSC blastema mesenchymal cells responded to increased BMP2 signaling by increasing expression of Prrx-1 to the level observed in response to 5% FBS (Fig 4). In addition, the rate of proliferation increased significantly (double that for cultures in basal medium) at all three concentrations of BMP2 that were tested (Figs 7 and 8A). The average labeling indices for all three concentrations were comparable to each other and to in vivo rates of proliferation [13,28–30]. Since the mean LI did not change in response to increasing dose, and the variability (standard error) was considerable smaller at the lowest dose tested, we concluded that 1 ng/ml BMP2 was an appropriate dose for experiments with OSC axolotl blastema cells.
Fig 7. Proliferation in response to BMP2.

Immunofluorescence showing EdU labeling of blastema slices originating from the same blastema and cultured in either basal medium or 100ng/mL BMP2. EdU positive proliferating cells are green, nuclei are stained with DAPI and are blue. The proximal (P) to distal (D) orientation of the sections is indicated. Scale bar = 1mm.
Fig 8. Labeling index in response to exogenous BMP2 and LDN193189.

A) Labeling index of blastema mesenchymal cells in blastema slices over a period of five days in culture in the presence or absence of exogenous human BMP2 in amounts as indicated. The number of biological replicates = 4. B) Labeling index of blastema mesenchymal cells in blastema slices over a period of 5 days cultured in 5%FBS and in the presence or absence of LDN193189 in amounts as indicated. The number of biological replicates = 3. Error bars in both A and B represent S.E.M. and P-values were determined by t-test with 2 tails assuming unequal variances.
To determine if the BMP2 pathway was indeed involved with proliferation in OSC, we used the BMP2 pathway inhibitor LDN193189 in order to abrogate the effect that 5%FBS has on the proliferation rates of OSC given that BMP is present in FBS [54]. LDN193189 is a small molecule derivative of Dorsomophin that prevents the BMP induced Smad1/5/8 phosphorylation by binding to the ATP binding site in the kinase domain of Type I BMP receptors [55–57]. We found that incubation of OSC in medium containing 5% FBS along with LDN193189 at doses of 5, 50 and 500nM all had a significant effect on the rate of proliferation in slices (Fig 8B). At LDN193189 doses of 50nM and 500nM, the LI was reduced to the level observed in OSC cultured in basal medium (Fig 8B).
Discussion
Maintenance of in vivo blastema cell behavior is dependent on maintenance of cell-cell and cell-matrix interactions
Amphibian cells, including those derived from salamanders, have been used for in vitro experimentation since the advent of modern cell culture [58]. Early experiments using salamander tissues typically employed the technique of whole explant cultures, either using the developing limb bud or regenerating limb blastema [4,24,32,43,59–61]. While these experiments paved the way for the study of regeneration in vitro, the use of explant cultures has its drawbacks; namely, increased cell death, lack of reproducibility owing to the variability of sample source and preparation, and lack of access to the interior mesenchymal tissues for experimental manipulation when blastemas are cultured with the epithelium present [62]. In addition, normal rates of proliferation are not maintained. For example, medium bud blastemas from newts were reported to have a mitotic index of 1.2% (number of cells in the M phase of the cell cycle relative to the total number of cells) that decreased to 0.2% when the blastemas were placed in explant culture [24].
The technique of dissociation, culture and passage of individual cells is an alternative approach to in vitro studies that avoids the drawbacks of explant cultures. Although blastemas can be dissociated and blastema cells can be cultured individually, the cells quickly lose the behavior of blastema cells in vivo, most notably they have a dramatically lower rate of proliferation. Data for the labeling index (number of cells in the S phase of the cell cycle relative to the total number of cells) of blastema cells in vitro are comparable to uninjured skin (2%) in contrast to that of blastema cells in vivo (20%- 40%) [7,13,26–30]. It appears that this decrease in proliferation is in part a consequence of cultured blastema cells exiting the cell cycle when cultured. For example, the percentage of blastema cells that incorporated BrdU after 96 hours of continuous labeling (growth fraction) was highly variable (0.1% to 9%, with a median of 1%) [7] in contrast to a growth fraction of 40% to 80% (3H-thymidine injection every 12 hours for two to four days) for blastema cells in vivo [48]. With OSC (this study), the growth fraction (80 hours labeling which corresponds to approximately 1.5 cell cycles) was 35% after five days in culture (Fig 5B) and as high as 63% after 10 days in culture (N = 3 samples). Consistent with maintaining a growth fraction comparable to blastema cells in vivo, the labeling index of OSC blastema cells cultured with FBS also was comparable to what has been reported in vivo (Fig 5A).
We hypothesize that normal blastema cell behavior is dependent on maintaining normal cell-cell and cell-matrix interactions. OSC allows for cell-cell and cell-matrix interactions to occur, as well as providing conditions for all of the cells to be uniformly exposed to the cell culture media and experimental supplements [8,62]. Consistent with this hypothesis are the findings that blastema cells remain viable with only a low amount of cell death, that they continue to express a blastema marker (Prrx-1), that they continue to proliferate at in vivo levels, that their rate of proliferation is responsive to the presence of nerve signals provided by DRG, and that they can respond to changes in the culture environment (addition of FBS or BMP2).
There are several likely reasons why OSC is an appropriate model for studying the biology of blastema cells. First, a regenerating limb blastema is comprised of a heterogeneous population of progenitor cells [63,64], which has been noted previously as to why culture techniques requiring dissociation are unadvisable [32]. In addition, blastemas are not only heterogeneous in terms of cell types of different lineages, but also in terms of the state of developmental plasticity of blastema cells. The cells in the apical region of a medium-late bud blastema are undifferentiated and developmentally plastic, in contrast to those at more basal regions that are beginning to redifferentiate and have stabilized their positional information [65,66]. The interaction of blastema cells with different positional information controls growth and pattern formation during regeneration [1,67,68], and thus the spatial relationship of blastema cells would be expected to be important in terms of regulating their behaviors. Dissociation of a blastema into single cells disrupts the spatial organization of the blastema; whereas, OSC maintains the normal positional interactions. The stability of positional information is maintained for about a week in posterior blastema cells cultured as a micromass in a fibrin clot and grafted into the anterior of a host blastema in vivo [35]. Testing the hypothesis that the spatial organization of positional information also is maintained in OSC is a goal of future experiments.
Blastema cell gene expression and proliferation are responsive to nerve signals and to BMP2
A distinct advantage to organotypic slice cultures is that cultured blastema cells remain viable without the addition of FBS. Amphibian tissues are well suited for organ culture [69,70], and early experiments utilizing salamander tissues reported proliferation and the long-term survival of cells in basic salt solutions [32,43,59]. OSC blastema cells also survived and proliferated when cultured in basal medium with FBS. The mitogenic activity of FBS led to its use in early studies to identify mitogenic factors, and subsequently to general use in cell culture in order to expand cell populations for passage. It has been noted that FBS evokes a specific physiological response in cultured cells that is reminiscent of wound healing [71]. Cells such as fibroblasts, which are the progenitors of many of the cells found in the blastema [63,64], would encounter the factors found in FBS in the early wound environment created by tissue injury. Thus OSC blastema cells respond to FBS and proliferate at in vivo levels (Fig 5A), even when a nerve is not present. In spite of the response of OSC cultured blastema cells to added FBS, the utility of OSC for studying the behavior of blastema cells is that addition of FBS is not necessary; therefore, it is possible to study the mechanisms by which specific signaling pathways are involved in the regenerative response.
One pathway that likely is involved in the regulation of blastema cell behavior is BMP signaling, which previously has been implicated in controlling regeneration in a number of model systems, including mammals [36,47,72–74]. The OSC model allowed us to determine that axolotl blastema cells responded to a specific, optimal dose of human BMP2 (1 ng/ml) by increasing the expression of a blastema marker gene (Prrx-1) and increasing the rate of proliferation. Furthermore, incubation with the BMP pathway inhibitor, LDN193189, decreased the rate of proliferation similar to one seen in OSC cultured in basal medium. In addition, we have identified a similar response (changes in gene expression and proliferation) to signaling from co-cultured nerves, which has been associated previously with FGF as well as BMP signaling [21,36,47]. The report that an ectopic blastema can be induced by implanting beads soaked in a cocktail of FGF2, FGF8 and GDF5/BMP2/BMP7 in the absence of a deviated nerve provides additional evidence that blastema formation can be induced by the activation of well characterized and highly conserved signaling pathways (e.g. FGF and BMP) [47]. The OSC model provides the opportunity to discover the mechanism for the spatial and temporal co-regulation of these pathways in order to induce and enhance a regenerative response.
Acknowledgments
The authors wish to thank the members of the David Gardiner/Susan Bryant Lab for their help with and encouragement of the research. We are especially grateful to Minhan Dinh, Matt Korn and Karina Cramer in the Cramer lab at UCI for sharing their equipment and expertise in the use of organotypic slice cultures.
Data Availability
All relevant data are within the paper.
Funding Statement
Research funded by a United States Army Multidisciplinary University Research Initiative (MURI) subcontract from Tulane University (TUL 589-09/10), and the National Science Foundation through its support of the Ambystoma Genetic Stock Center at the University of Kentucky, Lexington. JL was funded in part by a research fellowship from the Grass Foundation (www.grassfoundation.org) for a summer research internship at the Marine Biological Laboratory, Woods Hole, Massachusetts. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Endo T, Bryant SV, Gardiner DM (2004) A stepwise model system for limb regeneration. Dev Biol 270: 135–145. 10.1016/j.ydbio.2004.02.016 [DOI] [PubMed] [Google Scholar]
- 2. Satoh A, Graham G, Bryant SV, Gardiner DM (2008) Neurotrophic regulation of epidermal dedifferentiation during wound healing and limb regeneration in the axolotl (Ambystoma mexicanum). Dev Biol 319: 321–335. 10.1016/j.ydbio.2008.04.030 [DOI] [PubMed] [Google Scholar]
- 3. Satoh A, Makanae A, Hirata A, Satou Y (2011) Blastema induction in aneurogenic state and Prrx-1 regulation by MMPs and FGFs in Ambystoma mexicanum limb regeneration. Dev Biol 355: 263–274. 10.1016/j.ydbio.2011.04.017 [DOI] [PubMed] [Google Scholar]
- 4. Globus M, Liversage RA (1975) In vitro studies of limb regeneration in adult Diemictylus viridescens: neural dependence of blastema cells for growth and differentiation. J Embryol Exp Morphol 33: 813–829. [PubMed] [Google Scholar]
- 5. Jabaily JA, Blue P, Singer M (1982) The Culturing of Dissociated Newt Forelimb Regenerate Cells. J Exp Zool 219: 67–73. [Google Scholar]
- 6. Hinterberger TJ, Cameron JA (1983) Muscle and Cartilage Differentiation in Axolotl Limb Regeneration Blastema Cultures. J Exp Zool 226: 399–407. 10.1002/jez.1402260310 [DOI] [PubMed] [Google Scholar]
- 7. Kumar A, Godwin JW, Gates PB, Garza-Garcia AA, Brockes JP (2007) Molecular basis for the nerve dependence of limb regeneration in an adult vertebrate. Science 318: 772–777. 10.1126/science.1147710 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Gähwiler BH, Capogna M, Debanne D, McKinney RA, Thompson SM (1997) Organotypic slice cultures: a technique has come of age. Trends Neurosci 20: 471–477. 10.1016/S0166-2236(97)01122-3 [DOI] [PubMed] [Google Scholar]
- 9. Muller TL, Ngo-Muller V, Reginelli A, Taylor G, Anderson R, Muneoka K (1999) Regeneration in higher vertebrates: Limb buds and digit tips. Semin Cell Dev Biol 10: 405–413. 10.1006/scdb.1999.0327 [DOI] [PubMed] [Google Scholar]
- 10. Knapp D, Schulz H, Rascon CA, Volkmer M, Scholz J, Nacu E, et al. (2013) Comparative transcriptional profiling of the axolotl limb identifies a tripartite regeneration-specific gene program. PLoS ONE 8: e61352–e61352. 10.1371/journal.pone.0061352 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Todd TJ (1823) On the process of reproduction of the members of the aquatic salamander. Q Jour Sci Lit Arts 16: 84–96. [Google Scholar]
- 12. Singer M (1952) The influence of the nerve in regeneration of the amphibian extremity. Q Rev Biol 27: 169–200. 10.1086/398873 [DOI] [PubMed] [Google Scholar]
- 13. Wallace H (1981) Vertebrate limb regeneration. John Wiley & Sons; 1 pp. [Google Scholar]
- 14. Singer M (1946) The nervous system and regeneration of the forelimb of adult Triturus; the influence of number of nerve fibers, including a quantitative study of limb innervation. J Exp Zool 101: 299–337. 10.1002/jez.1401010303 [DOI] [PubMed] [Google Scholar]
- 15. Singer M (1947) The Nervous System and Regeneration of the Forelimb of Adult Triturus. 6. a Further Study of the Importance of Nerve Number, Including Quantitative Measurements of Limb Innervation. J Exp Zool 104: 223–249. [DOI] [PubMed] [Google Scholar]
- 16. Singer M, Craven L (1948) The Growth and Morphogenesis of the Regenerating Forelimb of Adult Triturus Following Denervation at Various Stages of Development. J Exp Zool 108: 279–308. 10.1002/jez.1401080207 [DOI] [PubMed] [Google Scholar]
- 17. Singer M, Egloff FR (1949) The nervous system and regeneration of the forelimb of adult Triturus; the effect of limited nerve quantities on regeneration. J Exp Zool 111: 295–314. [DOI] [PubMed] [Google Scholar]
- 18. Sidman RL, Singer M (1951) Stimulation of forelimb regeneration in the newt, Triturus viridescens, by a sensory nerve supply isolated from the central nervous system. Am J Physiol 165: 257–260. [DOI] [PubMed] [Google Scholar]
- 19. Kamrin AA, Singer M (1959) The growth influence of spinal ganglia implanted into the denervated forelimb regenerate of the newt, Triturus. J Morphol 104: 415–439. 10.1002/jmor.1051040303 [DOI] [PubMed] [Google Scholar]
- 20. Salley-Guydon JD, Tassava RA (2006) Timing the commitment to a wound-healing response of denervated limb stumps in the adult newt, Notophthalmus viridescens. Wound Repair Regen 14: 479–483. 10.1111/j.1743-6109.2006.00154.x [DOI] [PubMed] [Google Scholar]
- 21. Mullen LM, Bryant SV, Torok MA, Blumberg B, Gardiner DM (1996) Nerve dependency of regeneration: the role of Distal-less and FGF signaling in amphibian limb regeneration. Dev Suppl 122: 3487–3497. [DOI] [PubMed] [Google Scholar]
- 22. Tassava RA, Goldhammer DJ, Tomlinson BL (1987) Cell-Cycle Controls and the Role of Nerves and the Regenerate Epithelium in Urodele Forelimb Regeneration—Possible Modifications of Basic Concepts. Biochem Cell Biol 65: 739–749. [DOI] [PubMed] [Google Scholar]
- 23. Globus M, Vethamany-Globus S (1977) Transfilter mitogenic effect of dorsal root ganglia on cultured regeneration blastemata, in the newt,Notophthalmus viridescens. Dev Biol 56: 316–328. 10.1016/0012-1606(77)90273-1 [DOI] [PubMed] [Google Scholar]
- 24. Carlone RL, Foret JE (1979) Stimulation of Mitosis in Cultured Limb Blastemata of the Newt, Notophthalmus-Viridescens. J Exp Zool 210: 245–252. 10.1002/jez.1402100207 [DOI] [Google Scholar]
- 25. Boilly B, Albert P (1988) Control of blastema cell proliferation during axolotl limb regeneration: in vitro cell culture study. Monogr Dev Biol 21: 1–8. [PubMed] [Google Scholar]
- 26. Ferretti P, Brockes JP (1988) Culture of Newt Cells From Different Tissues and Their Expression of a Regeneration-Associated Antigen. J Exp Zool 247: 77–91. [DOI] [PubMed] [Google Scholar]
- 27. Kumar A, Godwin JW (2010) Preparation and culture of limb blastema stem cells from regenerating larval and adult salamanders. Cold Spring Harbor Protocols 2010: pdb–prot5367. 10.1101/pdb.prot5367 [DOI] [PubMed] [Google Scholar]
- 28. Maden M (1978) Neurotrophic Control of the Cell-Cycle During Amphibian Limb Regeneration. J Embryol Exp Morphol 48: 169–175. [PubMed] [Google Scholar]
- 29. Tomlinson BL, Globus M, Vethamany-Globus S (1984) Blastema cell cycle in vitro and attempted restimulation of blastema cell cycling in denervated blastemata of the adult newt, Notophthalmus viridescens. J Exp Zool 232: 249–258. 10.1002/jez.1402320212 [DOI] [Google Scholar]
- 30. Goldhammer DJ, Tomlinson BL, Tassava RA (1992) Ganglia Implantation as a Means of Supplying Neurotrophic Stimulation to the Newt Regeneration Blastema—Cell-Cycle Effects in Innervated and Denervated Limbs. J Exp Zool 262: 71–80. 10.1002/jez.1402620110 [DOI] [PubMed] [Google Scholar]
- 31. Fimian WJ (1959) The in vitro cultivation of amphibian blastema tissue. J Exp Zool 140: 125–143. 10.1002/jez.1401400107 [DOI] [PubMed] [Google Scholar]
- 32. Conn ME, Dearlove GE, Dresden MH (1979) Selection of a chemically defined medium for culturing adult newt forelimb regenerates. In Vitro 15: 409–414. 10.2307/4292222 [DOI] [PubMed] [Google Scholar]
- 33. Choo AF, Logan DM, Rathbone MP (1978) Nerve trophic effects: an in vitro assay for factors involved in regulation of protein synthesis in regenerating amphibian limbs. J Exp Zool 206: 347–354. 10.1002/jez.1402060306 [DOI] [PubMed] [Google Scholar]
- 34. Mescher AL, Loh JJ (1981) Newt forelimb regeneration blastemas in vitro: cellular response to explanation and effects of various growth-promoting substances. J Exp Zool 216: 235–245. 10.1002/jez.1402160204 [DOI] [PubMed] [Google Scholar]
- 35. Groell AL, Gardiner DM, Bryant SV (1993) Stability of positional identity of axolotl blastema cells in vitro. Roux's Arch Dev Biol 202: 170–175. [DOI] [PubMed] [Google Scholar]
- 36. Athippozhy A, Lehrberg J, Monaghan JR, Gardiner DM, Voss SR (2014) Characterization of in vitro transcriptional responses of dorsal root ganglia cultured in the presence and absence of blastema cells from regenerating salamander limbs. Regeneration 1: 1–10. 10.1002/reg2.14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Wallace H, Maden M (1976) The cell cycle during amphibian limb regeneration. J Cell Sci 20: 539–547. [DOI] [PubMed] [Google Scholar]
- 38. Gavet O, Pines J (2010) Progressive Activation of CyclinB1-Cdk1 Coordinates Entry to Mitosis. Dev Cell 18: 533–543. 10.1016/j.devcel.2010.02.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Pfaffl MW (2001) A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res 29: e45–e45. 10.1093/nar/29.9.e45 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Satoh A, Gardiner DM, Bryant SV, Endo T (2007) Nerve-induced ectopic limb blastemas in the axolotl are equivalent to amputation-induced blastemas. Dev Biol 312: 231–244. 10.1016/j.ydbio.2007.09.021 [DOI] [PubMed] [Google Scholar]
- 41. Whited JL, Lehoczky JA, Austin CA, Tabin CJ (2011) Dynamic Expression of Two Thrombospondins During Axolotl Limb Regeneration. Dev Dyn 240: 1249–1258. 10.1002/dvdy.22548 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Holtfreter J (1943) Properties and functions of the surface coat in amphibian embryos. J Exp Zool 93: 251–323. [Google Scholar]
- 43. Wilde CE (1950) Studies on the organogenesis in vitro of the urodele limb bud. J Morphol 86: 73–113. [DOI] [PubMed] [Google Scholar]
- 44. Hay ED, Fischman DA (1961) Origin of the blastema in regenerating limbs of the newt Triturus viridescens: An autoradiographic study using tritiated thymidine to follow cell proliferation and migration. Dev Biol 3: 26–59. 10.1016/0012-1606(61)90009-4 [DOI] [PubMed] [Google Scholar]
- 45. Satoh A, Bryant SV, Gardiner DM (2012) Nerve signaling regulates basal keratinocyte proliferation in the blastema apical epithelial cap in the axolotl (Ambystoma mexicanum). Dev Biol 366: 374–381. 10.1016/j.ydbio.2012.03.022 [DOI] [PubMed] [Google Scholar]
- 46. Yokoyama H, Maruoka T, Aruga A, Amano T, Ohgo S, Shiroishi T, et al. (2011) Prx-1 Expression in Xenopus laevis Scarless Skin-Wound Healing and Its Resemblance to Epimorphic Regeneration. J Invest Dermatol 131: 2477–2485. 10.1038/jid.2011.223 [DOI] [PubMed] [Google Scholar]
- 47. Makanae A, Hirata A, Honjo Y, Mitogawa K, Satoh A (2013) Nerve independent limb induction in axolotls. Dev Biol 381: 213–226. 10.1016/j.ydbio.2013.05.010 [DOI] [PubMed] [Google Scholar]
- 48. Goldhammer DJ, Tassava RA (1987) An Analysis of Proliferative Activity in Innervated and Denervated Forelimb Regenerates of the Newt, Notophthalmus-Viridescens. Dev Suppl 100: 619–628. [Google Scholar]
- 49. Mayer H, Scutt AM, Ankenbauer T (1996) Subtle differences in the mitogenic effects of recombinant human bone morphogenetic proteins-2 to -7 on DNA synthesis on primary bone-forming cells and identification of BMP-2/4 receptor. Calcif Tissue Int 58: 249–255. [DOI] [PubMed] [Google Scholar]
- 50. Wong GA, Tang V, El-Sabeawy F, Weiss RH (2003) BMP-2 inhibits proliferation of human aortic smooth muscle cells via p21Cip1/Waf1. Am J Physiol Endocrinol Metab 284: E972–E979. 10.1152/ajpendo.00385.2002 [DOI] [PubMed] [Google Scholar]
- 51. Herrera B, Inman GJ (2009) A rapid and sensitive bioassay for the simultaneous measurement of multiple bone morphogenetic proteins. Identification and quantification of BMP4, BMP6 and BMP9 in bovine and human serum. BMC Cell Biol 10: –20 10.1186/1471-2121-10-20 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Wang Q, Zhao G, Xing S, Zhang L, Yang X (2011) Role of bone morphogenetic proteins in form-deprivation myopia sclera. Mol Vis 17: 647–657. [PMC free article] [PubMed] [Google Scholar]
- 53. Honda Y, Ding X, Mussano F, Wiberg A, Ho C- M, Nishimura I. (2013) Guiding the osteogenic fate of mouse and human mesenchymal stem cells through feedback system control. Sci Rep 3: –3420 10.1038/srep03420 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Kodaira K, Imada M, Goto M, Tomoyasu A, Fukuda T, Kamijo R, et al. (2006). Purification and identification of a BMP-like factor from bovine serum. Biochem Biophys Res Commun. 345:1224–31. [DOI] [PubMed] [Google Scholar]
- 55. Boergermann JH, Kopf J, Yu PB, Knaus P (2010) Dorsomorphin and LDN-193189 inhibit BMP-mediated Smad, p38 and Akt signalling in C2C12 cells. Int J Biochem Cell Biol 42: 1802–1807. 10.1016/j.biocel.2010.07.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Yu PB, Hong CC, Sachidanandan C, Babitt JL, Deng DY, Hoyng SA, et al. (2008) Dorsomorphin inhibits BMP signals required for embryogenesis and iron metabolism. Nat Chem Biol 4: 33–41. 10.1038/nchembio.2007.54 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Wrighton KH, Lin X, Yu PB, Feng X-H (2009) Transforming Growth Factor {beta} Can Stimulate Smad1 Phosphorylation Independently of Bone Morphogenic Protein Receptors. J Biol Chem 284: 9755–9763. 10.1074/jbc.M809223200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Holtfreter J (1929) Über histologische Differenzierungen von isoliertem Material jüngster Amphibienkeime. 1 pp. [Google Scholar]
- 59. Dalton HC (1950) Comparison of White and Black Axolotl Chromatophores in Vitro. J Exp Zool 115: 19. [Google Scholar]
- 60. Oudkhir M, Boilly B, Lassalle B (1986) Cell cycle changes after denervation of the newt limb regenerate, studied in vitro. Experientia 42: 189–190. [Google Scholar]
- 61. Mescher AL, Connell E, Hsu C, Patel C, Overton B (1997) Transferrin is necessary and sufficient for the neural effect on growth in amphibian limb regeneration blastemas. Dev Growth Differ 39: 677–684. 10.1046/j.1440-169X.1997.t01-5-00003.x [DOI] [PubMed] [Google Scholar]
- 62. Lossi L, Alasia S, Salio C, Merighi A (2009) Cell death and proliferation in acute slices and organotypic cultures of mammalian CNS. Prog Neurobiol 88: 221–245. 10.1016/j.pneurobio.2009.01.002 [DOI] [PubMed] [Google Scholar]
- 63. Muneoka K, Fox WF, Bryant SV (1986) Cellular Contribution From Dermis and Cartilage to the Regenerating Limb Blastema in Axolotls. Dev Biol 116: 256–260. 10.1016/0012-1606(86)90062-X [DOI] [PubMed] [Google Scholar]
- 64. Kragl M, Knapp D, Nacu E, Khattak S, Maden M, Epperlein HH, et al. (2009) Cells keep a memory of their tissue origin during axolotl limb regeneration. Nature 460: 60–65. 10.1038/nature08152 [DOI] [PubMed] [Google Scholar]
- 65. McCusker CD, Gardiner DM (2013) Positional information is reprogrammed in blastema cells of the regenerating limb of the axolotl (Ambystoma mexicanum). PLoS ONE 8: e77064–e77064. 10.1371/journal.pone.0077064 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. McCusker C, Lehrberg J, Gardiner D (2014) Position‐specific induction of ectopic limbs in non‐regenerating blastemas on axolotl forelimbs. Regeneration 1: 27–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. French V, Bryant PJ, Bryant SV (1976) Pattern Regulation in Epimorphic Fields. Science 193: 969–981. [DOI] [PubMed] [Google Scholar]
- 68. Bryant SV, Iten LE (1976) Supernumerary limbs in amphibians: Experimental production in Notophthalmus viridescens and a new interpretation of their formation. Dev Biol 50: 212–234. 10.1016/0012-1606(76)90079-8 [DOI] [PubMed] [Google Scholar]
- 69. Monnickendam MA, Millar JL, Balls M (1970) Cell proliferation in vivo and in vitro in visceral organs of the adult newt, Triturus cristatus carnifex. J Morphol 132: 453–459. 10.1002/jmor.1051320406 [DOI] [PubMed] [Google Scholar]
- 70. Monnickendam MA, Balls M (1973) Amphibian Organ-Culture. Experientia 29: 1–17. [DOI] [PubMed] [Google Scholar]
- 71. Iyer VR, Eisen MB, Ross DT, Schuler G, Moore T, Lee JC, et al. (1999) The transcriptional program in the response of human fibroblasts to serum. Science 283: 83–87. [DOI] [PubMed] [Google Scholar]
- 72. Reddien PW, Bermange AL, Kicza AM, Alvarado AS (2007) BMP signaling regulates the dorsal planarian midline and is needed for asymmetric regeneration. Dev Suppl 134: 4043–4051. 10.1242/dev.007138 [DOI] [PubMed] [Google Scholar]
- 73. Yu L, Han M, Yan M, Lee E-C, Lee J, Muneoka K (2010) BMP signaling induces digit regeneration in neonatal mice. Dev Suppl 137: 551–559. 10.1242/dev.042424 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Guimond J-C, Lévesque M, Michaud P-L, Berdugo J, Finnson K, Phillip A, et al. (2010) BMP-2 functions independently of SHH signaling and triggers cell condensation and apoptosis in regenerating axolotl limbs. BMC Dev Biol 10: –15 10.1186/1471-213X-10-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the paper.