

Abstract
The efficiency of immune-mediated clearance of cancer cells is hampered by immunosuppressive mediators in the malignant microenvironment, including NADPH oxidase–derived reactive oxygen species. We aimed at defining the effects of histamine, an inhibitor of the myeloid NADPH oxidase/NOX2, on the development of Ag-presenting dendritic cells (DCs) from myeloid precursors and the impact of these mechanisms for tumor growth. Histamine was found to promote the maturation of human DCs from monocytes by increasing the expression of HLA-DR and costimulatory molecules, which resulted in improved induction of Th cells with Th0 polarity. Experiments using wild-type and NOX2-deficient myelomonoblastic cells showed that histamine facilitated myeloid cell maturation only in cells capable of generating reactive oxygen species. Treatment of mice with histamine reduced the growth of murine EL-4 lymphomas in parallel with an increment of tumor-infiltrating DCs in NOX2-sufficient mice but not in NOX2-deficient (gp91phox−/−) mice. We propose that strategies to target the myeloid NADPH oxidase may facilitate the development of endogenous DCs in cancer.
Introduction
In several forms of cancer, poorly differentiated myeloid cells accumulate in the blood and in the malignant microenvironment as the result of increased myelopoiesis, along with a deficient differentiation of myeloid progenitors (1–3). The accumulating progenitor cells are suppressive to several aspects of T cell–mediated immunity and do not efficiently develop into Ag-presenting dendritic cells (DCs), thus impeding presentation of tumor-derived Ags to T cells (1–3). Several studies demonstrated that accumulation of immature myeloid cells and the absence of mature DCs result in accelerated tumor growth and poor outcome in human and murine cancers (1, 3–6). Therefore, understanding the regulation of myeloid cell differentiation, in particular with regard to the development of DCs, may provide novel therapeutic targets (2).
Recently, histamine was ascribed a role in myeloid cell differentiation based on studies of mice with genetic disruption of the unique histamine-forming histidine decarboxylase (HDC−/− mice). Yang et al. (7) observed an increased accumulation of immature CD11b+/Gr1+ myeloid cells during cancer progression in HDC−/− mice compared with HDC-sufficient mice and reported that HDC−/− mice were strikingly susceptible to chemically induced cancer. These investigators also reported that the exogenous administration of histamine to HDC−/− mice prevented the accumulation of immature myeloid cells (7). However, the detailed mechanisms by which histamine deficiency results in the accumulation of immature myeloid cells are largely unknown.
In addition to the role of histamine in myeloid cell differentiation, it has been ascribed immune-stimulatory properties by virtue of its inhibitory action on the production and release of immunosuppressive NADPH oxidase–derived reactive oxygen species (ROS) from phagocytes, such as monocytes and neutrophilic granulocytes (8–11). ROS are formed upon the assembly of the membrane-bound myeloid gp91phox subunit NOX2 with other membrane and cytosolic components (12). By reducing the formation and extracellular release of ROS, histamine was found to preserve NK cell function and, thus, promote NK cell–mediated clearance of malignant cells under conditions of phagocyte-induced oxidative stress (9, 10). These results formed the basis for the inclusion of histamine in cancer immunotherapy protocols (13). A histamine salt (histamine dihydrochloride) was recently approved for therapeutic use in acute myeloid leukemia within the European Union and in Israel (14, 15).
High levels of NADPH oxidase/NOX2-derived ROS were suggested to impede the maturation of immature myeloid cells into macrophages and DCs (16). For the current study, we sought to define whether the ROS-inhibitory properties of histamine translate into facilitated differentiation of myeloid cells, including the development of DCs. Our results imply that histamine promotes myeloid cell differentiation, including the development of monocyte-derived DCs, by targeting the NADPH oxidase/NOX2 and that these properties have implications for tumor growth.
Materials and Methods
Cell isolation and DC generation
PBMCs were prepared from blood donor buffy coats by Ficoll-Paque (Lymphoprep; Nycomed, Oslo, Norway) density centrifugation and were separated into CD14+ monocytes using iMag positive selection beads (BD Biosciences, San Diego, CA), according to the manufacturer’s instructions. The isolation procedures resulted in >98% pure monocytes. In some experiments, monocytes were purified by adherence: PBMCs were cultured in Iscove’s medium containing 10% human AB serum, 2 mM l-glutamine, 100 μg/ml penicillin, and 100 μg/ml streptomycin (complete medium) for 2 h at 4 × 106 cells/ml. Nonadherent cells were carefully washed away, and fresh medium was added. The purified monocytes were differentiated into DCs by culture in complete medium supplemented with IL-4 (500 U/ml) and GM-CSF (600 U/ml) (both from R&D Systems, Abingdon, U.K.), in the presence or absence of 100 μM histamine (Sigma-Aldrich, St. Louis, MO). In control experiments, monocytes were cultured in complete medium in the presence or absence of histamine, without added cytokines. In some experiments, 50 μM ranitidine or AH 20239AA (an inert chemical control to ranitidine; both from GlaxoSmithKline, Solna, Sweden) or 200 U/ml endotoxin-free catalase (Sigma-Aldrich) was added during DC generation. No significant differences were noted for DCs differentiated from monocytes isolated by beads or adherence. Supernatants from DCs cultured for 3 d in the presence or absence of histamine were analyzed for levels of IL-12, IL-6, and TNF using DuoSet ELISA Development Kits (R&D Systems).
Coculture of APCs and T cells
CD3+ T cells were purified from human PBMCs using iMag positive selection beads (BD Biosciences) (>98% purity) and frozen in CryoMaxx S (PAA Laboratories, Pasching, Austria). Thawed CD3+ T cells were stained with CellTrace Violet (Invitrogen) and cocultured with human allogeneic DCs in 96-well round-bottom plates at different DC/T cell ratios. In some experiments, DCs were preincubated with 2 μg/ml anti-CD86 (MAB141) or isotype Abs (both from R&D Systems) for 10 min before T cells were added, diluting the final Ab concentration to 1 μg/ml in the coculture. T cell proliferation was assessed by flow cytometry after 4–5 of DC:T cell coculture. At this time point, supernatants were collected for determination of the levels of IFN-γ, IL-13, IL-17, and IL-10 using the Cytometric Bead Array (R&D Systems) on a LSRFortessa (BD Biosciences), according to the manufacturers’ instructions.
Culture of PLB-985 cells
The human myelomonoblastic cell lines PLB-985 (PLB) (17) and the NOX2-deficient (NOX2-def) clone PLB (18) were kindly provided by Dr. Mary Dinauer (Washington University School of Medicine, St. Louis, MO). The wild-type (WT) PLB and NOX2-def PLB cells were cultured in Iscove’s medium containing 10% human AB serum, 2 mM l-glutamine, 100 U/ml penicillin, and 100 μg/ml streptomycin at 37°C, 5% CO2, in the presence or absence of histamine (100 μM) and GM-CSF (600 U/ml) for 3–6 d. In some experiments, ranitidine (50 μM) was added to the cultures. PLB cells also were differentiated by the addition of 1% DMSO to the culture medium for 3–5 d.
Quantitative reverse-transcription real-time PCR
One hundred myeloid cells were sorted with a BD FACSAria (BD Biosciences) into 96-well plates (Life Technologies) for direct cell lysis, with each well containing 5 μl 1 mg/ml BSA (Thermo Scientific) (19). Three technical replicates from each biological sample were collected and analyzed. Harvested cells were kept at −80°C until subsequent analysis. GrandScript cDNA SuperMix (TATAA Biocenter) was used for reverse transcription. Ten-microliter reactions, including 2 μl 5× GrandScript RT reaction mix, 0.5 μl GrandScript RT enzyme, and 5 μl lysed cells, were mixed. Reverse transcription was performed at 22°C for 5 min, 42°C for 30 min, and terminated at 85°C for 5 min. All samples were diluted with water to 60 μl before real-time PCR. Quantitative real-time PCR was performed with a CFX384 real-time cycler (Bio-Rad). Each 6-μl reaction contained 1× TATAA SYBR GrandMaster Mix (TATAA Biocenter) or 1× iQ Supermix (Bio-Rad), 400 nM of each primer (Sigma-Aldrich) or 1× TaqMan Gene Expression Assay (Life Technologies), and 2 μl diluted cDNA. Assay information and primer sequences are available upon request. The temperature profile for SYBR Green I assays was 95°C for 2 min, followed by 50 cycles of amplification (95°C for 20 s, 60°C for 20 s, and 70°C for 20 s), and 95°C for 3 min, followed by 40 cycles of amplification (95°C for 15 s, 60°C for 1 min) for TaqMan assays. The formation of expected PCR products was confirmed by agarose gel electrophoresis, and all samples containing SYBR Green I were analyzed by melting-curve analysis. Detection of genomic DNA was assessed with reverse-transcription negative samples for each assay. Quantitative real-time PCR data were analyzed as described (20) using GenEx software (MultiD). Briefly, nine potential reference genes were evaluated with NormFinder. RPS10, RPL7, and 18S were chosen as optimal reference genes. Cycle of quantification for the genes analyzed was normalized against the three chosen reference genes to compensate for any eventual difference in sample load. Values were converted to relative quantities in log2 scale.
Flow cytometry
The following fluorochrome-labeled anti-human mAbs were used: anti-CD14 (MΦP9), anti-CD86 (FUN-1), anti–HLA-DR (L243), anti-CD40 (5C3), anti-CD4 (RPA-T4), anti-CD8 (RPA-T8) (all from BD Biosciences), and anti-CD3 (S4.1) (Invitrogen, Carlsbad, CA). The purified anti-histamine H2 receptor Ab (polyclonal rabbit IgG) and antiflavocytochrome b558 (gp91phox; clone 7D5) were from MBL International (Woburn, MA), and the secondary reagent, goat anti-rabbit–PE–Cy5.5, was from Invitrogen. The following fluorochrome-labeled anti-mouse mAbs were purchased from BD Biosciences: anti-CD45 (30-F11), anti-CD11c (HL3), anti-IaIe (2G9), CD3 (145-2311), CD4 (RM4-5), and CD8 (53-6.7). In addition, CD16/CD32 (2.4 G2) (BD Biosciences), DCFDA (Molecular Probes/Invitrogen), and DAPI (Invitrogen) were used for flow cytometry. A minimum of 10,000 gated live (DAPI−) and hematopoietic (CD45+) cells was collected on a three-laser BD FACSAria (405, 488, and 633 nm) or a four-laser BD LSRFortessa (405, 488, 532, and 640 nm; both from BD Biosciences). Data were analyzed using FACSDiva Version 6 software (BD Biosciences) or FlowJo Version 8.4 software (TreeStar).
Staining for intracellular ROS
DCs or monocytes were stained for 30 min at 37°C with 4.7 μM DCFDA in RPMI 1640 medium. Cells were washed and analyzed by flow cytometry in the presence of DAPI to exclude dead cells.
Detection of ROS by chemiluminescence
Superoxide anion production by myeloid cells was determined by the use of isoluminol-ECL, as described elsewhere (21). In brief, myeloid cells were suspended at a concentration of 106/ml in Krebs-Ringer glucose buffer supplemented with isoluminol (10 μg/ml; Sigma-Aldrich) and HRP (4 U/ml; Boehringer Mannheim, Mannheim, Germany) and incubated in a 96-well plate at 37°C for 5 min in the presence or absence of 100 μM histamine or N-methyl-histamine dihydrochloride (an H2-type histamine receptor [H2R]-selective histamine derivate; Sigma-Aldrich), as well as in the presence or absence of 10 μM ranitidine or AH 20239AA. fMLF (10−7 M; Sigma-Aldrich) or Trp-Lys-Tyr-Met-Val-d-Met (10−7 M, R&D Systems) was added as an NADPH oxidase–triggering stimulus, and light emission was recorded continuously using a FLUOstar Omega plate reader (BMG, Ortenberg, Germany).
Confocal microscopy
DCs were generated from human monocytes, as described above, in the presence or absence of histamine. After 48 h, cells were replated in μ-Slide VI 0.4 chambers (Ibidi, München, Germany) for an additional 24 h. The cells were stained using 3.7% paraformaldehyde containing 100 μg/ml l-α-phosphatidylcholine (Sigma-Aldrich) and 5 U/ml Alexa Fluor 568 phalloidin (Invitrogen) for 20 min at 4°C. After extensive washing, cells were mounted in ProLong Gold Antifade reagent with DAPI (Invitrogen), and photomicrographs were obtained from nine randomly chosen DCs, in cultures with or without histamine, using a Zeiss LSM700 confocal microscope. DC diameters were acquired using ZEN Blue software (Zeiss) and measured at their most elongated point.
Histamine treatment of naive C57BL/6 mice
All animal experiments were approved by the Research Animal Ethics Committee in Gothenburg. C57BL/6 mice were obtained from Charles River Laboratories (Sulzfeld, Germany). Naive C57BL/6 mice were treated i.p. with histamine (1500 μg/mouse); after 2, 3, or 7 d, mice were sacrificed and analyzed for DC content in the spleen and peritoneal cavity. Peritoneal cells were collected by injection of 5 ml Iscove’s medium with 2% FCS into the peritoneal cavity and retrieving the fluid. Single-cell suspensions of splenocytes were prepared, and erythrocytes were lysed using RBC lysing buffer (Sigma-Aldrich). Splenocytes and peritoneal cells were stained with DC markers and analyzed using flow cytometry.
Isolation of Gr1+ myeloid cells
Bone marrow cells were isolated from naive C57BL/6 mice or B6.129S6-Cybbtm1din (NOX2-knockout [KO]) mice (Jackson Laboratory, Bar Harbor, ME). RBCs were lysed, and Gr1+ cells were isolated using Gr1-biotinylated Abs, followed by streptavidin-conjugated MACS beads (Miltenyi Biotec, Lund, Sweden), according to the manufacturer’s instructions. The purity was routinely >95%.
EL-4 mouse model
The EL-4 thymoma cell line (kindly provided by Dr. Ingo Schmitz, Otto-von-Guericke-University, Magdeburg, Germany) was cultured in DMEM (Invitrogen) supplemented with 10% FBS (Sigma-Aldrich), 100 U/ml penicillin, and 100 μg/ml streptomycin at 37°C, 5% CO2, prior to injection in mice.
C57BL/6 mice or NOX2-KO mice were injected s.c. with 2 × 105 or 3 × 105 EL-4 lymphoma cells and treated with histamine (i.p.; 1500 μg/mouse or 75 mg/kg) three times a week, starting the day before tumor inoculation. Tumor size was measured every second day (calculated as length × width); after 2 wk, mice were sacrificed, and tumors and spleens were collected. Single-cell suspensions of splenocytes were prepared and depleted from erythrocytes using RBC Lysing buffer (Sigma-Aldrich). Tumors were dissociated into single-cell suspensions by enzymatic digestion using the Tumor Dissociation Kit in combination with mechanical dissociation using a gentleMACS Dissociator (both from Miltenyi Biotec). Single-cell suspensions of tumors and spleens were stained with DC markers and T cell markers and analyzed using flow cytometry.
Statistical analysis
For statistical analyses, we used the two-tailed paired or unpaired t test. For multiple comparisons, one-way ANOVA, followed by the Bonferroni multiple-comparison test, was used.
Results
Histamine promotes the development of DCs from human monocytes
We first determined the effects of histamine on the differentiation of DCs from monocyte precursors. Human monocytes were cultured with IL-4 and GM-CSF in the presence or absence of histamine, added at the onset of culture. After 3–6 d, DCs generated in the presence of histamine expressed significantly higher levels of maturation markers, such as the Ag-presenting receptor HLA-DR and the costimulatory molecules CD86 and CD40, as measured by flow cytometry (Fig. 1A–D, Supplemental Fig. 1A). The enhanced expression of HLA-DR and CD86 after culture in the presence of histamine was paralleled by an increase in expression at the mRNA level, as analyzed by quantitative reverse-transcription real-time PCR (Supplemental Fig. 1B). Histamine did not significantly upregulate the expression of these surface receptors when monocytes were cultured in the absence of growth factors (Fig. 1A–D, Supplemental Fig. 1A). Analysis of the kinetics of acquisition of maturation markers revealed that the presence of histamine enhanced CD86 expression on monocyte-derived DCs within 2 d of culture (Supplemental Fig. 1C). Monocytes cultured for 3 d with IL-4/GM-CSF acquired morphological features of DCs, regardless of the presence of histamine. A tendency toward a more stretched-out morphology was noted for DCs that had matured in the presence of histamine (Fig. 1E, Supplemental Fig. 1D, 1E).
FIGURE 1.

Histamine promotes the development of DCs from monocyte precursors. Human monocytes were cultured for 3–6 d in the absence (Ctl) or presence of histamine (his) in IL-4– and GM-CSF–containing media (DC) or in the absence of growth factors (MO) (n = 29 or 26 for DC and n = 13 or 11 for MO). Representative line graphs of myeloid cell HLA-DR (A) and CD86 expression (B) along with MFI of myeloid cell HLA-DR (C) and CD86 (D) for all donors tested. (E) Representative confocal micrographs of actin staining of a DC cultured in the absence (Ctl) or presence of histamine. ***p < 0.001 paired Student t test.
DC-derived proinflammatory cytokines were analyzed in supernatants from DCs generated from IL-4– and GM-CSF–treated monocytes after 3 d of culture. The levels of IL-12 and TNF-α were low in all DC cultures (<15 pg/ml for IL-12 and 18 ± 9 and 11 ± 3 pg/ml for TNF-α in cultures without versus with histamine [mean ± SD], n = 5). Histamine did not affect the levels of IL-6 in the DC cultures (122 ± 86 and 117 ± 64 pg/ml in cultures without versus with histamine [mean ± SD], n = 5).
Histamine-induced DCs activate T cells with Th0 polarity
We next determined the effects of histamine-induced DCs on T cell responses. DCs, generated from IL-4/GM-CSF–treated monocytes in the presence or absence of histamine, were cultured at different ratios with allogeneic T cells. T cell proliferation and the production of cytokines were determined after 4–5 d of coculture. Histamine-generated DCs were significantly more efficient than were control DCs in inducing T cell proliferation and production of the T cell cytokines IFN-γ and IL-4 (Fig. 2A–D), implying that histamine-induced DCs triggered a Th0-type response in T cells. No differences were noted in the production of IL-10 or IL-17 in these cocultures (data not shown). Negligible T cell proliferation was induced when monocytes cultured in the absence of growth factors were used as APCs (data not shown). Treatment of DCs with neutralizing Abs against CD86, present throughout coculture with T cells, strongly reduced T cell proliferation (Fig. 2E, 2F), whereas an isotype-control Ab did not (data not shown).
FIGURE 2.

Histamine-matured DCs are efficient activators of T cells. DCs developed in the presence (red line) or absence (black line) of histamine were cocultured with allogeneic CellTrace Violet–stained T cells. The percentage of proliferating CD4+ T cells (A) and CD8+ T cells (B) in response to stimulation by different doses of allogeneic DCs generated in the presence or absence of histamine (n = 10). Supernatant content of IFN-γ (C) and IL-4 (D) (n = 5). Percentage of proliferating CD4+ (E) and CD8+ (F) T cells. Results were obtained with neutralizing anti-CD86 Abs (dashed lines) or isotype controls Abs (solid lines) present in the coculture (n = 3). All data are mean ± SEM. The effects of histamine-matured DCs on T cell proliferation and cytokine production, as well as the effect of anti-CD86 Abs, were evaluated using the paired Student t test. *p < 0.05, **p < 0.01, ***p < 0.001.
Prodifferentiating properties of histamine are NADPH oxidase/NOX2 dependent
Earlier studies showed that histamine reduces NADPH oxidase–dependent ROS formation and release in several types of myeloid cells, which was proposed to protect adjacent lymphocytes from ROS-induced inhibition and apoptosis (8, 11). We verified the action of histamine on the extracellular release of ROS by stimulating monocytes with the ROS inducer fMLF (Supplemental Fig. 2A). We next measured the levels of intracellular ROS in DCs by using the fluorescent radical probe DCFDA and observed a significant reduction in intracellular ROS for DCs that had been generated in the presence of histamine (Fig. 3A). The reduction in intracellular ROS after histamine treatment was not accompanied by a reduction in NADPH oxidase expression, as reflected by the median fluorescence intensity (MFI) of the membrane-bound NADPH oxidase subunit NOX2 (the myeloid gp91phox subunit) (the NOX2 expression by DCs differentiated in the absence or presence of histamine was 2761 ± 942 and 3206 ± 1541, respectively [mean ± SD], n = 9). To explore whether scavengers of extracellular ROS mimic the DC maturation induced by histamine, DCs were generated in the presence of catalase, which did not affect the levels of CD86 expressed by DCs (Fig. 3B).
FIGURE 3.

Prodifferentiating properties of histamine are NADPH oxidase/NOX2 dependent. (A) Human monocytes were cultured in the presence or absence of histamine for 3–6 d with IL-4 and GM-CSF (DC) or without growth factors (MO). The levels of intracellular ROS were determined by a flow-based DCFDA assay and are displayed as MFI. The p values were obtained using the Student paired t test. (B) Expression of CD86 (MFI) after culture of human monocytes with IL-4/GM-CSF in the presence or absence of histamine or the extracellular ROS scavenger catalase. (C–E) Results of experiments in which WT PLB cells and NOX2-def PLB cells (NOX2 KO) were cultured for 3–6 d with GM-CSF in the presence or absence of histamine. (C) MFI of CD11b expression. Percentage of CD11b+ cells (D) and CD14+ cells (E). The p values in (B)–(E) were obtained using repeated-measures ANOVA and the Bonferroni post hoc test; the medians and quartiles are indicated by boxes, and the error bars show the minimum to maximum values. (F) Representative line graphs of PLB cell expression of CD11b (MFI). (G) Representative line graphs of CD11b expression (MFI) of WT and NOX2-def cell lines differentiated in the presence or absence of 1% DMSO (one representative experiment of three). *p < 0.05, **p < 0.01, ***p < 0.001.
NOX2 is the principal component of the NADPH oxidase of phagocytes. To clarify the involvement of the NADPH oxidase for the observed prodifferentiating properties of histamine we used the myelomonoblastic cell line PLB (17) and its NOX2-def variant (NOX2-def PLB) (18). WT PLB or NOX2-def PLB cells were cultured in the presence or absence of histamine. In WT PLB cells, histamine triggered significant upregulation of CD11b and CD14 expression. In contrast, histamine did not alter the expression of these markers of maturation in NOX2-def PLB cells (Fig. 3C–F). The induction of DC Ags induced by 1% DMSO was equal in WT and NOX2-def PLB cells, as reflected by the level of CD11b expression (Fig. 3G). Differentiated WT PLB cells, but not NOX2-def PLB cells, produced ROS after maturation, which was inhibited by histamine (Supplemental Fig. 2B, data not shown).
Histamine induces maturation of DCs and myelomonoblastic cells via H2Rs
Histaminergic regulation of NADPH oxidase–dependent ROS production in myeloid cells is reportedly transduced by H2Rs (8, 22). We verified that the inhibitory effects of histamine on ROS formation were mediated by H2Rs by stimulating monocytes with the ROS inducer fMLF and measuring the induction of ROS in the presence or absence of histamine, the H2R-specific antagonist ranitidine, a chemical inert control to ranitidine [AH20239AA, in which the thioether of ranitidine is replaced by an ether, thereby reducing its H2R affinity > 50-fold (8)], and a highly selective H2R agonist (N-methyl-histamine) (Fig. 4A).
FIGURE 4.
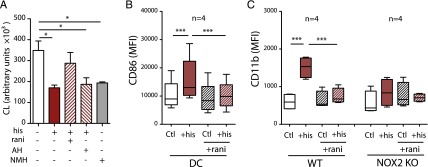
Histamine facilitates maturation of DCs and myelomonoblastic cells via H2Rs. (A) ROS production by human monocytes stimulated with fMLF alone (open bar) or fMLF in the presence of histamine (his; 100 μM), ranitidine (rani; 10 μM), AH20239AA (AH; 10 μM), or N-methyl histamine (NMH; 100 μM) (n = 3). (B) CD86 expression (MFI) on DCs and monocytes cultured in the presence or absence of histamine (his) and/or ranitidine (rani). (C) CD11b expression (MFI) of PLB cells and NOX2-def PLB cells cultured with GM-CSF in the presence or absence of histamine (his) and/or ranitidine (rani). The medians and quartiles are indicated by boxes, and the error bars show the minimum to maximum values. *p < 0.05, ***p < 0.001 repeated-measures ANOVA and Bonferroni post hoc test.
To define the histamine receptors mediating the prodifferentiating properties of histamine, we first assessed the expression of H2Rs on monocytes and DCs. Human monocytes expressed H2Rs at high intensity. The expression was downregulated upon histamine stimulation and differentiation toward DCs (data not shown). Next, DCs were generated by histamine treatment in the presence or absence of ranitidine. The histamine-induced maturation of DCs, as reflected by induction of CD86 expression, was completely antagonized by ranitidine (Fig. 4B). AH20239AA did not antagonize histamine-induced DC maturation at concentrations equimolar to ranitidine (data not shown). H2Rs were expressed at similar levels by WT and NOX2-def PLB cells (data not shown). Similar to its action on DC maturation, ranitidine antagonized the histamine-induced differentiation of WT PLB cells (Fig. 4C).
Histamine promotes the accumulation of intratumoral DCs and reduces lymphoma growth in vivo
Treatment of naive C57BL/6 mice with histamine did not affect the number of DCs in spleen or in the peritoneal cavity. Also, histamine treatment did not alter the expression of MHC class II or CD86 by these DCs (data not shown). We reasoned that the prodifferentiating effects of histamine might be more pronounced in a malignant microenvironment, in which the redox balance is frequently shifted in favor of oxidation (23). We chose the EL-4 lymphoma model, in which tumor growth is reportedly controlled by DC function (24–26). EL-4 lymphoma cells were inoculated s.c. into C57BL/6 mice, followed by systemic treatment with histamine every second day during tumor progression. After 13 d, mice were sacrificed and analyzed for the presence and activation grade of splenic and intratumoral DCs and CD8+ T cells.
Treatment with histamine significantly reduced the growth rate of EL-4 tumors (Fig. 5A), along with a significant increase in the content of intratumoral CD11c+/MHCII+ DCs (Fig. 5B, 5C). There was no significant difference in HLA-DR or CD86 expression by the intratumoral DCs between treatment groups, although a trend toward increased expression of CD86 in histamine-treated animals was observed (Supplemental Fig. 3A). The number of splenic DCs in tumor-bearing mice was not markedly affected by histamine administration; however, treatment with histamine was associated with a significant induction of CD86 expression on splenic DCs, along with a significant increase in splenic CD8+ T cells (Fig. 5B, Supplemental Fig. 3).
FIGURE 5.

Histamine promotes the accumulation of intratumoral DCs and reduces EL-4 lymphoma growth in vivo in a NADPH oxidase/NOX2–dependent manner. (A) Growth of EL-4 lymphoma cells implanted s.c. into C57BL/6 mice treated with histamine (his; red bars) or NaCl (open bars). (B and E) Percentage of CD11c+IaIe+ splenic and intratumoral DCs after 13 d of tumor growth. (C) FACS plots of intratumoral DCs in a representative control (Ctl) mouse and corresponding results in a histamine-treated mouse (+his). Live (DAPI−) hematopoietic (CD45+) cells are displayed. (D) Tumor growth of EL-4 lymphoma in NOX2−/− C57BL/6 mice during treatment with histamine (red bars) or NaCl (open bars). In (A), (B), (D), and (E), the boxes represent the medians and quartiles, and the error bars show the minimum to maximum values. (F) Correlation between the percentage of CD11c+IaIe+ intratumoral DCs and tumor size after 13 d of growth for WT and NOX2−/− C57BL/6 mice. (G) Correlation between the percentage of CD3+CD8+ intratumoral T cells and tumor size after 13 d of growth for WT and NOX2−/− C57BL/6 mice. (H) Correlation between the percentage of CD11c+IaIe+ intratumoral DCs and the percentage of CD3+CD8+ intratumoral T cells. **p < 0.01, ***p < 0.001 Student t test.
Histamine-induced inhibition of lymphoma growth is NADPH oxidase/NOX2 dependent
Murine Gr1+ myeloid cells from WT, but not from NOX2-def (gp91phox−/−), C57BL/6 mice produced ROS in response to the hexapeptide Trp-Lys-Tyr-Met-Val-d-Met, which, in turn, was inhibited by histamine (Supplemental Fig. 2C, data not shown). In contrast to its effects in WT tumor-bearing mice, histamine did not affect the growth rate of s.c. EL-4 tumors in gp91phox−/− mice (Fig. 5D), and it did not alter the intratumoral infiltration of DCs in these animals (Fig. 5E). Also, the level of expression of CD86 on splenic and tumor DCs was unaffected by histamine treatment in NOX2-def mice, as was the percentage of CD8+ T cells (data not shown). In NOX2-sufficient and NOX2-def mice, a significant negative correlation was observed between the presence of intratumoral DCs and tumor size (Fig. 5E), as well as between intratumoral CD8+ T cells and tumor size (Fig. 5F), whereas a significant positive correlation was identified for the content of intratumoral DCs versus CD8+ T cells (Fig. 5G).
Discussion
Ag-presenting DCs differentiate from various progenitors in the bone marrow or from monocyte precursors in the periphery (27, 28). This study aimed at defining the effects of histamine on myeloid cell differentiation, with a focus on its role in the development of monocyte-derived DCs. Earlier studies on the role of histamine for DC function and development yielded discordant results. Several groups reported that DCs matured by histamine in conjunction with a microbial maturation-promoting stimulus express similar levels of costimulatory molecules but produce lower levels of proinflammatory cytokines, such as IL-12, compared with DCs matured with the microbial stimulus alone (29–33). The altered cytokine expression was reportedly dependent, in part, on the agonistic action of histamine at H2Rs, and it favored the development of Th2-polarized (30, 32) or regulatory T cell-polarized (33) T cells in cultures of DCs and allogeneic T cells. These findings are challenged by results showing that H2R-KO mice are characterized by blunted Th1 responses and resistance to induction of Th1-dependent autoimmunity (34–36). In addition, the administration of histamine for therapeutic purposes in humans reportedly favors the maturation of Th1 cell, rather than Th2 or regulatory T cell, responses (37). Thus, when patients with malignant melanoma were treated with a histamine derivative (histamine dihydrochloride) in conjunction with IL-2, the frequencies of IFN-γ–producing CD4+ and CD8+ T cells (type 1 T cells) increased significantly in patients receiving histamine/IL-2, but not in those receiving IL-2 alone (37). The number of IL-13–producing T cells, reflecting Th2 polarization, was not altered in histamine-treated patients (37).
In the current study, DCs matured from human monocytes in the presence of histamine triggered significantly higher Th1 responses, as well as Th2 responses, compared with control DCs. These results were obtained in experiments in which histamine was present throughout the course of DC differentiation, without the addition of other maturation stimuli, such as LPS or other TLR ligands. In this setting, only low doses of proinflammatory cytokines were produced by the DCs, regardless of the presence or absence of histamine. Based on these observations, we propose that histamine polarizes DCs toward the induction of Th0-type responses and that additional stimuli are required to determine the polarity of the ensuing Th response.
The experiments using WT or NOX2-def variants of myelomonoblastic PLB cells suggested that the inhibitory action of histamine on NADPH oxidase–related functions is critical for the promotion of myeloid cell maturation. We observed that histamine inhibited ROS production from myeloid cells via H2Rs (Fig. 4A), which is in accordance with previous studies (8, 22). In analogy, the prodifferentiating effects of histamine on DCs and WT PLB cells were mediated by H2Rs, because they were completely blocked by the specific H2R antagonist ranitidine but not by an inert chemical control to this compound (Fig. 4B, 4C). Although further studies are required to define the precise mechanisms involved, a conceivable interpretation of these results is that NADPH oxidase–dependent ROS formation impedes the differentiation of myeloid cells and that histamine H2R-mediated inhibition of the NADPH oxidase facilitates myeloid cell differentiation, including the development of DCs from monocyte precursors. The finding that catalase, a potent scavenger of extracellular ROS, did not mimic the DC-promoting action of histamine implies that histamine acts mainly on the intracellular ROS compartment to facilitate DC maturation.
Treatment of naive mice with histamine in vivo did not alter the numbers of DCs or their expression of activation markers. However, mice treated with histamine during progression of EL-4 lymphomas showed a significantly enhanced accumulation of intratumoral DCs, which was paralleled by a reduction in tumor growth. The histamine-treated tumor-bearing mice also showed a higher expression of CD86 on splenic DCs and a higher content of splenic CD8+ T cells (Supplemental Fig. 3), further supporting that histamine also positively regulates DC function in vivo. There was a significant positive correlation between the number of splenic and tumor CD8+ T cells (R2 = 0.26, n = 26, p = 0.007) and a significant negative correlation between intratumoral CD8+ T cells and tumor size (Fig. 5G), as well as between splenic CD8+ T cells and tumor size (R2 = 0.27, n = 27, p = 0.005), which suggests that the immune phenotype in the spleen mirrors that of the effector cells at the tumor site. The reduction in tumor growth after treatment with histamine, as well as the induction of intratumoral DCs, was absent in genetically NOX2-def mice, suggesting that these effects of histamine resulted from regulation of NADPH oxidase/NOX2 function.
In conjunction with previous reports demonstrating a role for DCs in controlling tumor growth in the EL-4 lymphoma model (24–26), our results support a hypothesis that improved DC function contribute to a reduction in lymphoma growth after treatment with histamine in vivo. The findings of a negative correlation between intratumoral DC accumulation and EL-4 tumor size and a positive correlation between intratumoral DC accumulation and the presence of CD8+ T cells in tumors are in line with this hypothesis. A negative correlation also was observed between the presence of intratumoral CD8+ T cells and tumor size. However, these results do not preclude the possibility that contributions by supplementary or alternative mechanisms may explain the reduction in tumor growth by histamine treatment, indicating that further mechanistic studies are warranted.
Supplementary Material
Acknowledgments
We thank Dr. Mary Dinauer for providing the PLB WT and NOX2-KO cell lines and Dr. Ingo Schmitz for providing the EL-4 thymoma cell line.
This work was supported by Swedish Research Council Grants 2012-2047, 2012-3205, and 2011-3003, the Swedish Society for Medical Research, Swedish Society of Medicine Grant SLS-406151, Swedish Cancer Society Grants CAN 212/595 and CAN 213/550, the Swedish State via Agreements for Medical Education ALFGBG-151441 and ALFGBG-292701, the Erna and Victor Hasselblad Foundation, the Torsten and Ragnar Söderberg Foundation, the BioCARE National Strategic Research Program at the University of Gothenburg, and the Sahlgrenska Academy at the University of Gothenburg.
The online version of this article contains supplemental material.
- DC
- dendritic cell
- H2R
- H2-type receptor
- KO
- knockout
- MFI
- median fluorescence intensity
- NOX2-def
- NOX2 deficient
- PLB
- PLB-985
- ROS
- reactive oxygen species
- WT
- wild-type.
Disclosures
K.H. holds patents on the use of histamine in cancer treatment. The other authors have no financial conflicts of interest.
References
- 1.Gabrilovich D. 2004. Mechanisms and functional significance of tumour-induced dendritic-cell defects. Nat. Rev. Immunol. 4: 941–952. [DOI] [PubMed] [Google Scholar]
- 2.Gabrilovich D. I., Nagaraj S. 2009. Myeloid-derived suppressor cells as regulators of the immune system. Nat. Rev. Immunol. 9: 162–174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Gabrilovich D. I., Ostrand-Rosenberg S., Bronte V. 2012. Coordinated regulation of myeloid cells by tumours. Nat. Rev. Immunol. 12: 253–268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lee B. N., Follen M., Rodriquez G., Shen D. Y., Malpica A., Shearer W. T., Reuben J. M. 2006. Deficiencies in myeloid antigen-presenting cells in women with cervical squamous intraepithelial lesions. Cancer 107: 999–1007. [DOI] [PubMed] [Google Scholar]
- 5.Pinzon-Charry A., Ho C. S., Maxwell T., McGuckin M. A., Schmidt C., Furnival C., Pyke C. M., López J. A. 2007. Numerical and functional defects of blood dendritic cells in early- and late-stage breast cancer. Br. J. Cancer 97: 1251–1259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Pinzon-Charry A., Maxwell T., López J. A. 2005. Dendritic cell dysfunction in cancer: a mechanism for immunosuppression. Immunol. Cell Biol. 83: 451–461. [DOI] [PubMed] [Google Scholar]
- 7.Yang X. D., Ai W., Asfaha S., Bhagat G., Friedman R. A., Jin G., Park H., Shykind B., Diacovo T. G., Falus A., Wang T. C. 2011. Histamine deficiency promotes inflammation-associated carcinogenesis through reduced myeloid maturation and accumulation of CD11b+Ly6G+ immature myeloid cells. Nat. Med. 17: 87–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hellstrand K., Asea A., Dahlgren C., Hermodsson S. 1994. Histaminergic regulation of NK cells. Role of monocyte-derived reactive oxygen metabolites. J. Immunol. 153: 4940–4947. [PubMed] [Google Scholar]
- 9.Brune M., Hansson M., Mellqvist U. H., Hermodsson S., Hellstrand K. 1996. NK cell-mediated killing of AML blasts: role of histamine, monocytes and reactive oxygen metabolites. Eur. J. Haematol. 57: 312–319. [DOI] [PubMed] [Google Scholar]
- 10.Aurelius J., Martner A., Brune M., Palmqvist L., Hansson M., Hellstrand K., Thoren F. B. 2012. Remission maintenance in acute myeloid leukemia: impact of functional histamine H2 receptors expressed by leukemic cells. Haematologica 97: 1904–1908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mellqvist U. H., Hansson M., Brune M., Dahlgren C., Hermodsson S., Hellstrand K. 2000. Natural killer cell dysfunction and apoptosis induced by chronic myelogenous leukemia cells: role of reactive oxygen species and regulation by histamine. Blood 96: 1961–1968. [PubMed] [Google Scholar]
- 12.Rada B., Leto T. L. 2008. Oxidative innate immune defenses by Nox/Duox family NADPH oxidases. Contrib. Microbiol. 15: 164–187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hellstrand K., Hansson M., Hermodsson S. 2000. Adjuvant histamine in cancer immunotherapy. Semin. Cancer Biol. 10: 29–39. [DOI] [PubMed] [Google Scholar]
- 14.Brune M., Castaigne S., Catalano J., Gehlsen K., Ho A. D., Hofmann W. K., Hogge D. E., Nilsson B., Or R., Romero A. I., et al. 2006. Improved leukemia-free survival after postconsolidation immunotherapy with histamine dihydrochloride and interleukin-2 in acute myeloid leukemia: results of a randomized phase 3 trial. Blood 108: 88–96. [DOI] [PubMed] [Google Scholar]
- 15.Martner A., Thorén F. B., Aurelius J., Hellstrand K. 2013. Immunotherapeutic strategies for relapse control in acute myeloid leukemia. Blood Rev. 27: 209–216. [DOI] [PubMed] [Google Scholar]
- 16.Kusmartsev S., Gabrilovich D. I. 2003. Inhibition of myeloid cell differentiation in cancer: the role of reactive oxygen species. J. Leukoc. Biol. 74: 186–196. [DOI] [PubMed] [Google Scholar]
- 17.Tucker K. A., Lilly M. B., Heck L., Jr., Rado T. A. 1987. Characterization of a new human diploid myeloid leukemia cell line (PLB-985) with granulocytic and monocytic differentiating capacity. Blood 70: 372–378. [PubMed] [Google Scholar]
- 18.Zhen L., King A. A., Xiao Y., Chanock S. J., Orkin S. H., Dinauer M. C. 1993. Gene targeting of X chromosome-linked chronic granulomatous disease locus in a human myeloid leukemia cell line and rescue by expression of recombinant gp91phox. Proc. Natl. Acad. Sci. USA 90: 9832–9836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Svec, D., D. Andersson, M. Pekny, R. Sjoback, M. Kubista, and A. Stahlberg. 2013. Direct cell lysis for single-cell gene expression profiling. Front. Oncol. 3: 274. Available at: http://journal.frontiersin.org/article/10.3389/fonc.2013.00274/full. [DOI] [PMC free article] [PubMed]
- 20.Ståhlberg A., Elbing K., Andrade-Garda J. M., Sjögreen B., Forootan A., Kubista M. 2008. Multiway real-time PCR gene expression profiling in yeast Saccharomyces cerevisiae reveals altered transcriptional response of ADH-genes to glucose stimuli. BMC Genomics 9: 170 Available at: http://www.biomedcentral.com/1471-2164/9/170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. doi: 10.1016/s0022-1759(99)00146-5. Dahlgren, C., and A. Karlsson. 1999. Respiratory burst in human neutrophils. J. Immunol. Meth. 232: 3–14. [DOI] [PubMed] [Google Scholar]
- 22.Vasicek O., Lojek A., Jancinova V., Nosal R., Ciz M. 2014. Role of histamine receptors in the effects of histamine on the production of reactive oxygen species by whole blood phagocytes. Life Sci. 100: 67–72. [DOI] [PubMed] [Google Scholar]
- 23.Cairns R. A., Harris I. S., Mak T. W. 2011. Regulation of cancer cell metabolism. Nat. Rev. Cancer 11: 85–95. [DOI] [PubMed] [Google Scholar]
- 24.Herber D. L., Cao W., Nefedova Y., Novitskiy S. V., Nagaraj S., Tyurin V. A., Corzo A., Cho H. I., Celis E., Lennox B., et al. 2010. Lipid accumulation and dendritic cell dysfunction in cancer. Nat. Med. 16: 880–886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Nagaraj S., Pisarev V., Kinarsky L., Sherman S., Muro-Cacho C., Altieri D. C., Gabrilovich D. I. 2007. Dendritic cell-based full-length survivin vaccine in treatment of experimental tumors. J. Immunother. 30: 169–179. [DOI] [PubMed] [Google Scholar]
- 26.Esche C., Subbotin V. M., Maliszewski C., Lotze M. T., Shurin M. R. 1998. FLT3 ligand administration inhibits tumor growth in murine melanoma and lymphoma. Cancer Res. 58: 380–383. [PubMed] [Google Scholar]
- 27.Satpathy A. T., Wu X., Albring J. C., Murphy K. M. 2012. Re(de)fining the dendritic cell lineage. Nat. Immunol. 13: 1145–1154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Geissmann F., Manz M. G., Jung S., Sieweke M. H., Merad M., Ley K. 2010. Development of monocytes, macrophages, and dendritic cells. Science 327: 656–661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Frei R., Ferstl R., Konieczna P., Ziegler M., Simon T., Rugeles T. M., Mailand S., Watanabe T., Lauener R., Akdis C. A., O’Mahony L. 2013. Histamine receptor 2 modifies dendritic cell responses to microbial ligands. J. Allergy Clin. Immunol. 132: 194–204. [DOI] [PubMed] [Google Scholar]
- 30.Caron G., Delneste Y., Roelandts E., Duez C., Bonnefoy J. Y., Pestel J., Jeannin P. 2001. Histamine polarizes human dendritic cells into Th2 cell-promoting effector dendritic cells. J. Immunol. 167: 3682–3686. [DOI] [PubMed] [Google Scholar]
- 31.Gutzmer R., Langer K., Lisewski M., Mommert S., Rieckborn D., Kapp A., Werfel T. 2002. Expression and function of histamine receptors 1 and 2 on human monocyte-derived dendritic cells. J. Allergy Clin. Immunol. 109: 524–531. [DOI] [PubMed] [Google Scholar]
- 32.Mazzoni A., Young H. A., Spitzer J. H., Visintin A., Segal D. M. 2001. Histamine regulates cytokine production in maturing dendritic cells, resulting in altered T cell polarization. J. Clin. Invest. 108: 1865–1873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Zhai R. R., Jiang A. P., Wang H. B., Ma L., Ren X. X., Jiang J. F., Wu L., Wei J. F., Wang J. H. 2013. Histamine enhances HIV-1-induced modulation of dendritic cells to skew naïve T cell differentiation toward regulatory T cells. Virology 442: 163–172. [DOI] [PubMed] [Google Scholar]
- 34.Teuscher C., Poynter M. E., Offner H., Zamora A., Watanabe T., Fillmore P. D., Zachary J. F., Blankenhorn E. P. 2004. Attenuation of Th1 effector cell responses and susceptibility to experimental allergic encephalomyelitis in histamine H2 receptor knockout mice is due to dysregulation of cytokine production by antigen-presenting cells. Am. J. Pathol. 164: 883–892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Saligrama N., Case L. K., del Rio R., Noubade R., Teuscher C. 2013. Systemic lack of canonical histamine receptor signaling results in increased resistance to autoimmune encephalomyelitis. J. Immunol. 191: 614–622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Saligrama N., Noubade R., Case L. K., del Rio R., Teuscher C. 2012. Combinatorial roles for histamine H1-H2 and H3-H4 receptors in autoimmune inflammatory disease of the central nervous system. Eur. J. Immunol. 42: 1536–1546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Asemissen, A. M., C. Scheibenbogen, A. Letsch, K. Hellstrand, F. Thoren, K. Gehlsen, A. Schmittel, E. Thiel, and U. Keilholz. 2005. Addition of histamine to interleukin 2 treatment augments type 1 T-cell responses in patients with melanoma in vivo: immunologic results from a randomized clinical trial of interleukin 2 with or without histamine (MP 104). Clin. Cancer Res. 11: 290–297. [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.