

To some in the signal transduction field, heterotrimeric guanine nucleotide–binding protein (G protein) signaling is a “mature” field from which no important new insights will be obtained. This naïve view is somewhat understandable. (How did you know that my grant renewal was not funded by that study section?) The basics of G protein signaling (1) are a staple of biology textbooks. In these pathways, a heptihelical G protein–coupled receptor (GPCR) forms a preactivation complex with an inactive heterotrimeric G protein, composed of Gα, Gβ, and Gγ subunits, in which Gα is bound to guanosine diphosphate (GDP). Upon binding an extracellular agonist (or detection of a photon), the GPCR causes Gα to release GDP, allowing it to bind GTP and undergo a conformational change. Gα •GTP then dissociates from both the GPCR and the Gβγ dimer, which retains the same conformation as in the heterotrimer. Once activated, Gα, Gβγ, or both bind downstream effectors, including adenylyl cyclases, components of mitogen-activated protein kinase (MAPK) pathways, and membrane channels to regulate cellular responses. The cycle is completed when the Gα subunit’s GTPase activity, which can be stimulated by regulator of G protein signaling proteins, hydrolyzes the gamma phosphate from the GTP molecule to which it is bound, thus restoring Gα to the GDP-bound, inactive conformation and allowing reassembly of the preactivation complex. Studies of proteins from Saccharomyces cerevisiae, Cryptococcus neoformans, and Schizosaccharomyces pombe have identified four “noncanonical” Gβ subunits that bind Gα subunits in place of traditional Gβ subunits and function in G protein signaling pathways. All four Gα-binding proteins are structurally similar to traditional Gβs and belie the opinion that we have figured out all there is to know about G protein signaling.
In yeast and fungal systems, receptor-mediated G protein signaling is associated with pheromone- and nutrient-sensing pathways (2–4) that control metabolism, sexual development, and dimorphic cell growth. The pheromone signaling pathway is best known for regulating a MAPK cascade, whereas the nutrient signaling pathway promotes adenylyl cyclase activation and adenosine 3′,5′-monophosphate (cAMP) signaling. In all yeast and fungi examined to date, there are two or three Gα-encoding genes, yet a single Gβ-encoding gene. Although the simplest explanation for fewer Gβ genes is that the one Gβ functionally interacts with multiple Gα subunits, this has yet to be demonstrated in any system and is clearly not true in S. cerevisiae and S. pombe. Thus, some fungal Gα subunits either use a noncanonical Gβ partner for signaling or act as monomeric G proteins. Because the latter model is difficult to prove, efforts have been made to identify noncanonical Gβ subunits in several organisms.
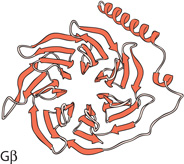
Before designating a protein as a noncanonical Gβ, we must define the criteria for this designation. The two most obvious criteria are that it binds a Gα and that it physically resembles a Gβ, although it must be sufficiently distinct from Gβ subunits or it would not be noncanonical. The seven-bladed propeller of the Gβ is one of the most easily recognized protein structures (5). Each blade is composed of an approximately 40-residue tryptophan–aspartic acid (WD) repeat, which folds into a four-stranded antiparallel β-sheet. WD repeat proteins make up a large protein superfamily that performs many distinct functions in eukaryotic cells. In fact, the WD repeat proteins are the largest protein family in S. pombe and the second-largest protein family in S. cerevisiae (6). Yet, although many fungal proteins contain WD repeats, there appears to be only one canonical Gβ in each fungal genome examined. Gβ subunits are 40 to 44% identical to each other, whereas only about 25% identical to other seven-bladed WD repeat proteins, and are thus readily distinguishable at the sequence level. It is less clear what additional criteria must be met for a protein to be viewed as a noncanonical Gβ, as discussed below.
Possibly the best-characterized G protein signaling pathway is the S. cerevisiae pheromone pathway, in which pheromone detection causes the Ste4–Ste18 Gβγ dimer to activate a MAPK pathway to control cell growth and gene expression (7, 8). The Gpa1 Gα is often overlooked, characterized as simply preventing inappropriate Gβγ signaling in the absence of pheromone detection. However, Slessareva et al. (9) showed that Gpa1 binds both the regulatory and catalytic subunits of phosphoinositol 3-kinase (PI3K): Vps15 and Vps34. This interaction takes place on the endosome rather than in the cell periphery, leads to Vps34 activation to raise PI3-phosphate levels, and is required for Gpa1Q323L-mediated signaling (Gpa1Q323L is a mutationally activated form of Gpa1), as well as contributing to normal pheromone signaling. Vps15, a 1454-residue protein of which the C-terminal 400 residues are predicted to form a seven-bladed WD repeat protein, appears to recruit Gpa1•GDP to the endosome, mimicking the role of Gβγ dimers in recruiting Gα subunits to the peripheral membrane. As such, a strong case can be made for designating Vps15 as a noncanonical Gβ. This also suggests two additional criteria for a noncanonical Gβ subunit: recruitment of Gα to the membrane in order to enhance signaling, and demonstration of a higher affinity for the GDP-bound Gα than for the GTP-bound form.
Because the S. cerevisiae Gpa2 Gα of the glucose-to-cAMP signaling pathway does not bind the Ste4p–Ste18p Gβγ dimer, efforts were made to identify Gpa2-binding proteins, leading to the identification of the kelch-repeat proteins Gpb1 (also known as Krh2) and Gpb2 (also known as Krh1) (10, 11). Crystal structure analysis of the Hypomyces rosellus galactose oxidase showed that kelch repeats fold into the same four-stranded antiparallel β-sheet as do WD repeats. Thus, Gpb1 and Gpb2, which contain seven kelch repeats, structurally resemble Gβ subunits. However, unlike Gβ subunits, they do not facilitate the interaction between the Gpa2 Gα and the Gpr1 GPCR (11). Instead, they negatively regulate Gpa2-mediated signaling. Two distinct mechanisms for this regulation have been reported. Epistasis analyses by Harashima et al. (12) indicated that Gpb1 and Gpb2 inhibit Ras2 activity [Ras2 binds and activates adenylyl cyclase (13)] by binding and stabilizing the Ira1 and Ira2 Ras GTPase-activating proteins (GAPs) (14). Gpb1 and Gpb2 are proposed to reduce Gpa2 activation before glucose detection and raise Ira1 and Ira2 activity to reduce Ras2 activity subsequent to glucose signaling.
However, this model cannot account for the observation by Lu and Hirsch (15) that cells lacking adenylyl cyclase require decreased concentrations of cAMP to remain viable if they also lack Gpb1 and Gpb2, suggesting that Gpb1 and Gpb2 act downstream of adenylyl cyclase to regulate protein kinase A (PKA) activity. In support of this, Peeters et al. (16) demonstrated that Gpb2 binds PKA catalytic subunits in the PKA holoenzyme, stabilizing the interaction between the catalytic and regulatory subunits, effectively raising the concentration of cAMP required to activate PKA. In this model, Gpa2 stimulates PKA activity by activating adenylyl cyclase to raise cAMP levels and by sequestering Gpb2 and Gpb1, which they showed preferentially bound Gpa2•GTP, to lower the concentration of cAMP required to activate PKA. Although Gpb2 and Gpb1 physically resemble Gβ subunits and bind Gpa2, they do not seem to perform any functional role normally assigned to a Gβ subunit. These proteins may be downstream effectors for Gpa2 rather than partners in signaling, suggesting a designation of Gβ mimic rather than noncanonical Gβ subunit. Additional studies to confirm whether the interaction between Gpa2 and these kelch repeat proteins occurs during glucose signaling or in the absence of glucose signaling will be important to understanding whether Gpb1 and Gpb2 regulate Gpa2 or vice versa.
The human fungal pathogen Cryptococcus neoformans appears to encode three Gα subunits but only one Gβ that functions in pheromone signaling. In a two-hybrid screen for proteins that bind the Gpa1 Gα of the cAMP pathway, Palmer et al. (17) identified a RACK1 (receptor for activated C kinase 1) homolog, Gib2, as a noncanonical Gβ subunit. RACK1 proteins contain seven WD repeats but are clearly not members of the Gβ protein family (18). Additional in vitro co-immunoprecipitations confirmed this interaction, and two-hybrid assays demonstrated an interaction between Gib2 and two likely Gγ subunits, Gpg1 and Gpg2. Antisense inhibition of Gib2 expression reduced cAMP levels, the most direct evidence that Gib2 positively regulates the cAMP pathway, although it remains to be seen whether this is a Gβ-like, Gpa1-dependent effect or a RACK1-like effect. Mammalian RACK1 binds the cAMP phosphodiesterase PDE4D5 (19), suggesting that Gib2 could bind and inhibit phosphodiesterase to reduce cAMP turnover. Gib2 also appears to act downstream of adenylyl cyclase by binding Smg1, a multicopy suppressor of a gpa1 mutation, the transcription of which is regulated by Gpa1 activity (20). This may explain the ability of Gib2 overexpression to partially suppress the loss of Gpa1 (as would phosphodiesterase inhibition), although much remains to be done to tease apart the role of Gib2 as a RACK1 homolog relative to its function as a Gβ-like protein. The ability to bind candidate Gγ subunits [although RACK1 proteins also bind Gβγ dimers (18)], along with the positive role in cAMP signaling, support a noncanonical Gβ designation for Gib2. However, evidence that Gib2 couples Gpa1 to a GPCR is still needed.
Unlike in S. cerevisiae and C. neoformans, the S. pombe glucose-to-cAMP pathway possesses an authentic heterotrimeric G protein (2, 21–23), whereas pheromone signaling is mediated by the Gpa1 Gα, which functions in the absence of a traditional Gβγ partner (24, 25). Using a two-hybrid screen, Goddard et al. (26) identified the seven-WD-repeat protein Gnr1 (systematic name SPCC1020.09) as binding Gpa1. Deletion and overexpression analyses of Gnr1 suggest that it negatively regulates pheromone signaling; however, the deletion result suggests that Gnr1 is not acting as a traditional Gβ to promote coupling of Gα to the GPCR. More remarkably, overexpression of mammalian Gβ subunits reduced the elevated pheromone signaling caused by deleting gnr1, although these Gβ subunits do not interact with Gpa1 in two-hybrid experiments. The lack of this twohybrid interaction does not preclude the possibility that these Gβ subunits bind Gpa1 in S. pombe or that they somehow prevent Gpa1 from functioning; however, the effect on signaling may be Gpa1-independent. Thus, it is too early to judge whether Gnr1 is a noncanonical Gβ, because the link between its interaction with Gpa1 and its role in pheromone signaling remain unclear.
The four proteins discussed here have claims of various strengths for consideration as noncanonical Gβ subunits (see Table 1 for a compilation of characteristics). All bind Gα subunits and physically resemble Gβ subunits. However, only the S. cerevisiae Vps15 protein can be said to carry out a true Gβ-like role, which I would define as facilitating signaling by its Gα partner. Although C. neoformans Gib2 functions in a positive manner in the same pathway as Gpa1, it remains to be determined whether it is doing so by a Gβ-like mechanism. Meanwhile, S. cerevisiae Gbp1 and Gbp2, as well as S. pombe Gnr1, are not required for the coupling of their respective Gα subunits to the GPCRs of these pathways, arguing against a designation of noncanonical Gβ subunit. What we clearly see is that Gα subunits bind authentic Gβ subunits and Gβ-like proteins, although it remains to be seen whether the nature of the interaction is similar. It is worth remembering that our appreciation of the signaling roles for authentic G protein subunits has evolved over the years; there was a time when the idea of signaling by Gβγ dimers was heretical. We should be wary that the term “noncanonical Gβ” may constrain our thinking about the roles of these proteins in their respective signaling pathways and our interpretation of the studies into their functions. As mentioned above, the S. cerevisiae Gpb1 and Gpb2 proteins, might be more properly viewed as downstream effectors in Gα-mediated signaling pathways.
Table 1.
A comparison of the fungal Gβ-like proteins.
 |
Species | Cellular process | Molecular character |
GPCR | Gα partner | βγ dimer | Downstream effectors |
|---|---|---|---|---|---|---|---|
 |
S. cerevisiae | Pheromone signaling | Seven-bladed WD repeat predicted for the C-terminal 400 amino acids (structure based on PDB: 2CNX) | Unknown | Gpa1•GDP | No | Catalytic subunit of PI3K |
 |
S. cerevisiae | Glucose to cAMP signaling | Seven kelch repeats forming Gβ-like β-propeller (structure based on residues WGP through FTPN of PDB: 1GOF) | Gpr1 | Gpa2•GDP Gpa2•GTP | Yes: Noncanonical Gγ Gpg1 | PKA, Ira1, Ira2 |
 |
C. neoformans | Glucose to cAMP signaling | Seven-bladed WD repeats predicted based on homology with RACK1 (structure based on chain A from PDB: 1TRJ) | Gpr4 | Gpa1 | Yes: γ subunits are Gpg1 Gpg2 | Smg1 |
 |
S. pombe | Pheromone signalling | Seven-bladed WD repeat protein (structure based on PDB: 2CNX) | Mam2, Map3 | Gpa1 | Unknown | Unknown |
These studies may say as much about Gα subunits that lack traditional Gβ partners as they do about the Gβ-like partners. As suggested by the Gnr1 study (26), Gα subunits such as S. pombe Gpa1 may be fairly promiscuous in their interactions with Gβ and Gβ-like proteins. This promiscuity could lead to the loss of their traditional Gβ partner. Selective pressure for the loss of the Gβ would occur if Gα binding to Gβ-like proteins created a pool of dissociated, and thus activated, Gβγ dimers that inappropriately activated downstream effectors. As in these fungal systems, a similar link between promiscuity and the loss of a partner has been observed in humans, as shown by a study examining the effect of adultery on the stability of marriages (27).
References
- 1.Sprang SR. G protein mechanisms: Insights from structural analysis. Annu. Rev. Biochem. 1997;66:639–678. doi: 10.1146/annurev.biochem.66.1.639. [DOI] [PubMed] [Google Scholar]
- 2.Hoffman CS. Except in every detail: Comparing and contrasting G-protein signaling in Saccharomyces cerevisiae and Schizosaccharomyces pombe. Eukaryot. Cell. 2005;4:495–503. doi: 10.1128/EC.4.3.495-503.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kays AM, Borkovich KA. In: The Mycota III. Brambl R, Marzluf GA, editors. Berlin-Heidelberg: Springer-Verlag; 2004. pp. 175–207. [Google Scholar]
- 4.Lengeler KB, Davidson RC, D’Souza C, Harashima T, Shen WC, Wang P, Pan X, Waugh M, Heitman J. Signal transduction cascades regulating fungal development and virulence. Microbiol. Mol. Biol. Rev. 2000;64:746–785. doi: 10.1128/mmbr.64.4.746-785.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Smith TF, Gaitatzes C, Saxena K, Neer EJ. The WD repeat: A common architecture for diverse functions. Trends Biochem. Sci. 1999;24:181–185. doi: 10.1016/s0968-0004(99)01384-5. [DOI] [PubMed] [Google Scholar]
- 6.Wood V, Gwilliam R, Rajandream MA, Lyne M, Lyne R, Stewart A, Sgouros J, Peat N, Hayles J, Baker S, Basham D, Bowman S, Brooks K, Brown D, Brown S, Chillingworth T, Churcher C, Collins M, Connor R, Cronin A, Davis P, Feltwell T, Fraser A, Gentles S, Goble A, Hamlin N, Harris D, Hidalgo J, Hodgson G, Holroyd S, Hornsby T, Howarth S, Huckle EJ, Hunt S, Jagels K, James K, Jones L, Jones M, Leather S, McDonald S, McLean J, Mooney P, Moule S, Mungall K, Murphy L, Niblett D, Odell C, Oliver K, O’Neil S, Pearson D, Quail MA, Rabbinowitsch E, Rutherford K, Rutter S, Saunders D, Seeger K, Sharp S, Skelton J, Simmonds M, Squares R, Squares S, Stevens K, Taylor K, Taylor RG, Tivey A, Walsh S, Warren T, Whitehead S, Woodward J, Volckaert G, Aert R, Robben J, Grymonprez B, Weltjens I, Vanstreels E, Rieger M, Schafer M, Muller-Auer S, Gabel C, Fuchs M, Düsterhöft A, Fritzc C, Holzer E, Moestl D, Hilbert H, Borzym K, Langer I, Beck A, Lehrach H, Reinhardt R, Pohl TM, Eger P, Zimmermann W, Wedler H, Wambutt R, Purnelle B, Goffeau A, Cadieu E, Dreano S, Gloux S, Lelaure V, Mottier S, Galibert F, Aves SJ, Xiang Z, Hunt C, Moore K, Hurst SM, Lucas M, Rochet M, Gaillardin C, Tallada VA, Garzon A, Thode G, Daga RR, Cruzado L, Jimenez J, Sanchez M, del Rey F, Benito J, Dominguez A, Revuelta JL, Moreno S, Armstrong J, Forsburg SL, Cerutti L, Lowe T, McCombie WR, Paulsen I, Potashkin J, Shpakovski GV, Ussery D, Barrell BG, Nurse P. The genome sequence of Schizosaccharomyces pombe. Nature. 2002;415:871–880. doi: 10.1038/nature724. [DOI] [PubMed] [Google Scholar]
- 7.Dohlman HG. G proteins and pheromone signaling. Annu. Rev. Physiol. 2002;64:129–152. doi: 10.1146/annurev.physiol.64.081701.133448. [DOI] [PubMed] [Google Scholar]
- 8.Dohlman HG, Thorner JW. Regulation of G protein-initiated signal transduction in yeast: Paradigms and principles. Annu. Rev. Biochem. 2001;70:703–754. doi: 10.1146/annurev.biochem.70.1.703. [DOI] [PubMed] [Google Scholar]
- 9.Slessareva JE, Routt SM, Temple B, Bankaitis VA, Dohlman HG. Activation of the phosphatidylinositol 3-kinase Vps34 by a G protein alpha subunit at the endosome. Cell. 2006;126:191–203. doi: 10.1016/j.cell.2006.04.045. [DOI] [PubMed] [Google Scholar]
- 10.Batlle M, Lu A, Green DA, Xue Y, Hirsch JP. Krh1p and Krh2p act downstream of the Gpa2p Gα subunit to negatively regulate haploid invasive growth. J. Cell Sci. 2003;116:701–710. doi: 10.1242/jcs.00266. [DOI] [PubMed] [Google Scholar]
- 11.Harashima T, Heitman J. The Gα protein Gpa2 controls yeast differentiation by interacting with kelch repeat proteins that mimic Gβ subunits. Mol. Cell. 2002;10:163–173. doi: 10.1016/s1097-2765(02)00569-5. [DOI] [PubMed] [Google Scholar]
- 12.Harashima T, Anderson S, Yates JR, 3rd, Heitman J. The kelch proteins Gpb1 and Gpb2 inhibit Ras activity via association with the yeast RasGAP neurofibromin homologs Ira1 and Ira2. Mol. Cell. 2006;22:819–830. doi: 10.1016/j.molcel.2006.05.011. [DOI] [PubMed] [Google Scholar]
- 13.Toda T, Uno I, Ishikawa T, Powers S, Kataoka T, Broek D, Cameron S, Broach J, Matsumoto K, Wigler M. In yeast, RAS proteins are controlling elements of adenylate cyclase. Cell. 1985;40:27–36. doi: 10.1016/0092-8674(85)90305-8. [DOI] [PubMed] [Google Scholar]
- 14.Tanaka K, Nakafuku M, Satoh T, Marshall MS, Gibbs JB, Matsumoto K, Kaziro Y, Toh-e A. S. cerevisiae genes IRA1 and IRA2 encode proteins that may be functionally equivalent to mammalian ras GTPase activating protein. Cell. 1990;60:803–807. doi: 10.1016/0092-8674(90)90094-u. [DOI] [PubMed] [Google Scholar]
- 15.Lu A, Hirsch JP. Cyclic AMP-independent regulation of protein kinase A substrate phosphorylation by Kelch repeat proteins. Eukaryot. Cell. 2005;4:1794–1800. doi: 10.1128/EC.4.11.1794-1800.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Peeters T, Louwet W, Gelade R, Nauwelaers D, Thevelein JM, Versele M. Kelch-repeat proteins interacting with the Galpha protein Gpa2 bypass adenylate cyclase for direct regulation of protein kinase A in yeast. Proc. Natl. Acad. Sci. U.S.A. 2006;103:13034–13039. doi: 10.1073/pnas.0509644103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Palmer DA, Thompson JK, Li L, Prat A, Wang P. Gib2, a novel Gbeta-like/ RACK1 homolog, functions as a Gbeta subunit in cAMP signaling and is essential in Cryptococcus neoformans. J. Biol. Chem. 2006;281:32596–32605. doi: 10.1074/jbc.M602768200. [DOI] [PubMed] [Google Scholar]
- 18.McCahill A, Warwicker J, Bolger GB, Houslay MD, Yarwood SJ. The RACK1 scaffold protein: A dynamic cog in cell response mechanisms. Mol. Pharmacol. 2002;62:1261–1273. doi: 10.1124/mol.62.6.1261. [DOI] [PubMed] [Google Scholar]
- 19.Yarwood SJ, Steele MR, Scotland G, Houslay MD, Bolger GB. The RACK1 signaling scaffold protein selectively interacts with the cAMP-specific phosphodiesterase PDE4D5 isoform. J. Biol. Chem. 1999;274:14909–14917. doi: 10.1074/jbc.274.21.14909. [DOI] [PubMed] [Google Scholar]
- 20.Pukkila-Worley R, Gerrald QD, Kraus PR, Boily MJ, Davis MJ, Giles SS, Cox GM, Heitman J, Alspaugh JA. Transcriptional network of multiple capsule and melanin genes governed by the Cryptococcus neoformans cyclic AMP cascade. Eukaryot. Cell. 2005;4:190–201. doi: 10.1128/EC.4.1.190-201.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hoffman CS. Glucose sensing via the protein kinase A pathway in Schizosaccharomyces pombe. Biochem. Soc. Trans. 2005;33:257–260. doi: 10.1042/BST0330257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Landry S, Hoffman CS. The git5 Gβ and git11 Gγ form an atypical Gβγ dimer acting in the fission teast glucose/cAMP pathway. Genetics. 2001;157:1159–1168. doi: 10.1093/genetics/157.3.1159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Landry S, Pettit MT, Apolinario E, Hoffman CS. The fission yeast git5 gene encodes a Gβ subunit required for glucose-triggered adenylate cyclase activation. Genetics. 2000;154:1463–1471. doi: 10.1093/genetics/154.4.1463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ladds G, Davis K, Das A, Davey J. A constitutively active GPCR retains its G protein specificity and the ability to form dimers. Mol. Microbiol. 2005;55:482–497. doi: 10.1111/j.1365-2958.2004.04394.x. [DOI] [PubMed] [Google Scholar]
- 25.Obara T, Nakafuku M, Yamamoto M, Kaziro Y. Isolation and characterization of a gene encoding a G-protein α subunit from Schizosaccharomyces pombe: Involvement in mating and sporulation pathways. Proc. Natl. Acad. Sci. U.S.A. 1991;88:5877–5881. doi: 10.1073/pnas.88.13.5877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Goddard A, Ladds G, Forfar R, Davey J. Identification of Gnr1p, a negative regulator of Galpha signalling in Schizosaccharomyces pombe, and its complementation by human Gbeta subunits. Fungal Genet. Biol. 2006;43:840–851. doi: 10.1016/j.fgb.2006.06.005. [DOI] [PubMed] [Google Scholar]
- 27.Charny IW, Parnass S. The impact of extramarital relationships on the continuation of marriages. J. Sex Marital Ther. 1995;21:100–115. doi: 10.1080/00926239508404389. [DOI] [PubMed] [Google Scholar]