

Abstract
Background
HIV-infected individuals frequently exhibit brain dysfunction despite antiretroviral treatment. The neuropathological mechanisms underlying these abnormalities remain unclear, pointing to the importance of identifying biomarkers sensitive to brain dysfunction.
Methods
We examined 74 medically stable HIV-infected individuals using T1-weighted MRI. Volumes of the cortical grey matter (GM), white matter (WM), caudate, putamen, globus pallidus, thalamus, hippocampus, amygdala, and ventricles were derived using automated parcellation. A panel of plasma cytokines was measured using multiplexed bead array immunoassay. A model selection algorithm was used to select the combination of clinical and cytokine markers that best predicted each brain volumetric measure in a series of linear regression models.
Results
Higher CD4 nadir, shorter HIV infection duration, and antiretroviral treatment were significantly related to higher volumes of the putamen, thalamus, hippocampus, and WM. Older age was related to lower volumes in most brain regions and higher ventricular volume. Higher IFN-γ, MCP-1, and TNF-α were related to higher volumes of the putamen, pallidum, amygdala, GM, and WM. Higher IL-1β, IL-6, IL-16, IL-18, IP-10, MIP-1β, and SDF-1α were related to lower volumes of the putamen, pallidum, thalamus, hippocampus, amygdala, GM, and WM; and higher ventricular volume.
Conclusion
The current findings provide evidence linking smaller brain volumes to HIV disease history, antiretroviral treatment, and advanced age. Cytokine markers, especially IL-6 and IL-16, showed robust association with brain volumes even after accounting for other clinical variables, demonstrating their utility in examining the mechanisms of HIV-associated brain abnormalities.
Keywords: HIV infection, hepatitis C infection, antiretroviral treatment, MRI
Introduction
People living with HIV continue to exhibit brain dysfunction despite effective antiretroviral treatment. Although the prevalence of advanced dementia has dramatically declined since the advent of combination antiretroviral therapy (CART), the overall rates of neurocognitive dysfunction in people living with HIV remain strikingly similar to those observed pre-CART1, 2. Evidence from neuroimaging studies has corroborated this impression, demonstrating prevalent structural, metabolic, and cerebrovascular abnormalities even in medically stable HIV-infected individuals3-10.
The etiology of this persistence of HIV-associated brain dysfunction in the CART era remains unclear. Recent findings have linked neurocognitive function and structural and metabolic MRI markers to nadir CD4 levels5, 11, 12, suggesting the importance of distant history of immune suppression on current brain integrity. Hepatitis C (HCV) coinfection has recently emerged as an important contributor to both neurocognitive and structural brain abnormalities observed in HIV-infected individuals5, 13-17. In addition, HIV-associated neurocognitive impairment has been linked to chronic systemic immune activation and inflammation 18-21, pointing to the potential utility of peripheral cytokine markers both for probing the underlying neuropathological mechanisms and as biomarkers sensitive to brain dysfunction.
The current study examines the independent contributions and relative importance of these diverse clinical markers on brain volume alterations. Volumes of the cerebral cortex, cerebral white matter, subcortical structures, and ventricles were measured from automated panellation of high-resolution T1-weighted MRI images. Volumetric measures were examined in the context of markers of HIV disease, antiretroviral treatment, HCV coinfection, and a panel of thirteen plasma cytokine markers measured using a multiplexed bead array immunoassay. We utilized multivariate statistical models to examine the independent contributions of clinical and cytokine markers on brain volumetric measures, and a model selection algorithm was used to identify the subset of markers that best predict each brain volume. We hypothesized that advanced HIV disease, lack of antiretroviral treatment, HCV coinfection, older age, and abnormal cytokine markers would be related to lower volumes of brain structures and greater ventricular volume.
Methods
Participants
Participants include 74 HIV-infected (HIV+) individuals recruited as part of an NIH-sponsored study of HIV-associated brain dysfunction at The Miriam Hospital / Brown University. The study was approved by the institutional review board, and informed consent was obtained from each participant prior to enrollment. Exclusion criteria included 1) history of head injury with loss of consciousness > 10 minutes; 2) neurological conditions including dementia unrelated to HIV, seizure disorder, stroke, and opportunistic infection of the brain; 3) severe psychiatric illness that may impact brain function, e.g., schizophrenia; and 4) diagnosis of alcohol or substance abuse or dependence based on DSM-IV criteria within 6 months prior to neuroimaging. HIV serostatus was documented by ELISA and confirmed by Western blot test.
Participants were between 24 and 65 years of age (mean = 45, SD = 9.66). Average duration of HIV infection was about 13 years. Most participants (82%) received combination antiretroviral therapy (CART), and generally had well-controlled HIV viral load and intact immune function: 72% had undetectable plasma HIV RNA (<75 copies/ml), average CD4 count was 523. More than half (54%) of the participants reported nadir CD4 < 200, indicating history of significant immune suppression. A significant number (34%) of participants had active HCV infection documented by detectable serum HCV RNA by PCR. Table 1 shows relevant demographic and clinical information of participants.
Table 1.
Relevant demographic and clinical information of study participants.
| N | 74 |
| Age (years) | 45.28 ± 9.66 |
| Gender (% male) | 66.22% |
| Ethnicity (% Caucasian) | 50.00% |
| Current CD4 (cells/ml) | 523.32 ± 270.92 |
| Nadir CD4 (cells/ml) | 189.27 ± 159.10 |
| Plasma HIV RNA (% undetectable) | 71.62% |
| HIV infection duration (years) | 12.66 ± 7.01 |
| Receiving CART (%) | 82.43% |
| Current hepatitis C infection (%) | 33.78% |
| Lifetime Alcohol Dependence (%) | 51.35% |
| Lifetime Substance Dependence (%) | 54.05% |
Note: CART = combination antiretroviral treatment. Continuous variables are reported as mean±SD. Proportions are reported in %.
Cytokine Measurements
Blood samples acquired for each participant were separated, and plasma samples were immediately frozen and stored at -80°C. Aliquots were used to measure chemokine and cytokine levels using an xMAP multiplexed bead array immunoassay. This approach permits simultaneous quantification of multiple cytokines in solution by capturing them onto antibody coated spectrally distinct fluorescent microspheres, and measuring fluorescence intensity using the Luminex-100 system (LuminexCorp., Austin, TX). Levels of IFN-γ, IL-1β, IL-6, IL-8, IL-10, IL-16, IL-18, IP-10, MCP-1, MIP-1β, SDF-1α, TNF-α, and TRAIL were measured. These specific markers were selected for their sensitivity to different aspects of systemic immune activation and inflammatory processes relevant to neuroAIDS based on findings by our group and others18, 22, 23. All cytokines reached the prerequisite concentration levels, and measurements were verified to be reliable across samples. Brief descriptions of measured cytokines are provided in Table 2.
Table 2.
Brief descriptions of measured cytokines.
| IFN-γ | Interferon-gamma | Produced by innate NK cells, acquired antigen-specific cytotoxic CD4+ and effector CD8+ T cells. Activates macrophages and critical for innate and adaptive immune responses to intracellular pathogens, tumor control, and inhibition of viral replication. |
| IL-1β | lnterleukin-1 beta | Produced by activated macrophages; mediates inflammatory responses, cell proliferation, apoptosis. Induces Cox-2 in CNS, causing inflammatory pain |
| IL-6 | lnterleukin-6 | Secreted by T cells and macrophages; triggers inflammation, acute phase response, fever. Anti-inflammatory effects include inhibiting TNF-α and IL-1, and activating IL-1ra and IL-10. |
| IL-8 | lnterleukin-8 | Made by macrophages and some epithelial and endothelial cells; Role in innate immune response. Major role in Chemotaxis of neutrophils. Also mediates inflammatory response and angiogenesis. |
| IL-10 | lnterleukin-10 | Produced by monocytes. Pleiotropic cytokine. As an anti-inflammatory cytokine, it inhibits macrophage and dendritic cell function, suppresses TNF-α. Acquires proinflammatory activity during immune response with IFN-α stimulation. |
| IL-16 | lnterleukin-16 | Secreted by lymphocytes. Pleiotropic cytokine. Functions as a chemoattractant (CD4+ cells), modulates T cell activation, and inhibits HIV replication. |
| IL-18 | lnterleukin-18 | Produced by macrophages and monocytes. Pro-inflammatory cytokine interacts with IL-12 to induce cell-mediated immune response with microbial infection and LPS, inducing severe inflammatory reactions. Stimulates NK and T cell release of IFN-γ, which activates macrophages. Inhibits IL4-dependent IgE, enhances B cell production. |
| IP-10 | Interferon-inducible protein 10 | Produced by various cell types including monocytes, endothelial cells, fibroblasts, keratinocytes. Induced by IFN-γ and TNF-α. Chemoattractant for activated T cells. |
| MCP-1 | Monocyte chemoattractant protein-1 | Expressed in monocytes, vascular endothelial cells, smooth muscle cells. CCL2 chemokine, induces monocyte attraction, and degranulation of basophils with histamine release. Induced by IL-1, TNF-α, PDGF, TGF-β, and LIF |
| MIP-1β | Macrophage inflammatory protein-1 beta | Produced by macrophages. CCL4 chemokine that generates local inflammatory responses, induces superoxide production by neutrophils. Chemotactic activity for lymphocytes, macrophages, NK cells, and monocytes with inflammation; down-regulates CCR5, inhibiting HIV-1 blocking. |
| SDF-1α | Stromal cell-derived factor-1 alpha | Expressed ubiquitously, except in blood cells. Small cytokine member of CXCL12 family of chemokines. Activates leukocytes due to strong chemotactic effects. Induced by pro-inflammatory stimuli, e.g. TNF-α and IL-1β. |
| TNF-α | Tumor necrosis factor-alpha | Secreted by macrophages, monocytes, neutrophils, T cells, NK cells after stimulation with LPS. CD4+ cells secrete TNF-α. Also made by astrocytes, microglial cells, smooth muscle cells, and fibroblasts. Mediates systemic inflammation, inhibits viral replication, and inhibits tumorigenesis. |
| TRAIL | TNF-related apoptosis-inducing ligand | Expressed broadly in tissues. Cytokine induces proapoptotic caspase activity by up-regulating pro-apoptotic Bel proteins. Causes apoptosis in hepatocytes, neural cells, and thymocytes |
MRI Data Acquisition and Analysis
All neuroimaging was performed on the same Siemens Tim Trio 3-tesla MRI scanner located at Brown University MRI Research Facility. High-resolution structural MRI of the whole brain was acquired in the sagittal plane using a T1-weighted MPRAGE pulse sequence with TE/TR = 3.06/2250ms, flip angle = 9°, FOV = 220mm, matrix = 256×256, slice thickness = 0.86mm. Head restraint was used during image acquisition and quality control of T1 images was performed immediately following acquisition to identify excessive movement and other artifacts. Repeated acquisitions were prescribed as necessary, and images with inadequate quality were excluded from analysis. Parcellation of brain regions was performed on each T1-weighted MRI image using automated algorithms implemented in Freesurfer, where anatomical labels are assigned to each voxel based on probabilistic estimates after a nonlinear registration to an atlas, and volumetric measures were derived for each brain24. Combined bilateral volumes of cortical grey matter (GM), cerebral white matter (WM), caudate, putamen, globus pallidus, thalamus, hippocampus, amygdala, and overall ventricles were thus derived (Figure 1). Intracranial volumes were also measured and included in all statistical models to control for variability in head size.
Figure 1.
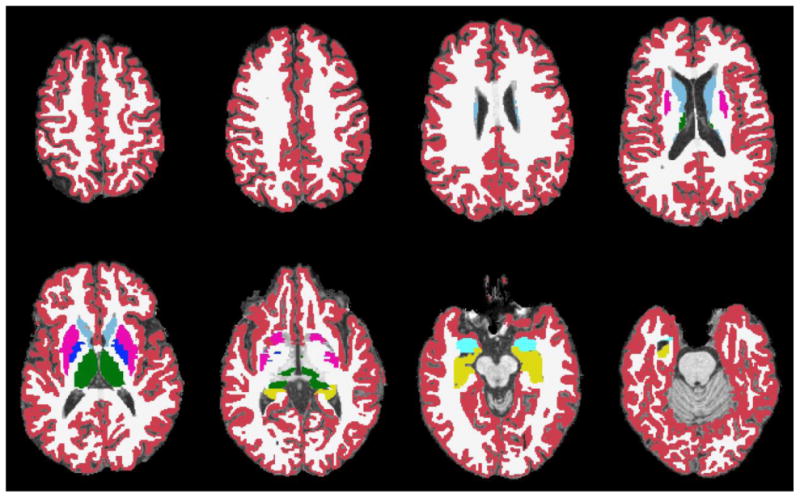
Segmented T1-weighted axial slices showing volumes of interest, including cortical grey matter (red), cerebral white matter (white), caudate (light blue), putamen (pink), globus pallidus (dark blue), thalamus (green), hippocampus (yellow), amygdala (teal), and overall ventricles (shown unmasked).
Statistical Analysis
All statistical analyses were performed in R version 2.13.0 (http://www.r-project.org). A series of linear multiple regression models were used to examine the independent contributions of clinical variables and cytokine markers on each brain volumetric measure. Each full model consisted of clinical variables (age, current and nadir CD4 levels, detectable plasma HIV RNA, years of HIV infection, current CART regimen, and detectable serum HCV RNA), thirteen cytokine measures as noted above, and intracranial volumes to control for head size. Age, CD4 levels, and infection duration were included as continuous covariates, while the remaining clinical variables were included as dummy coded binary covariates. Cytokine and nadir CD4 levels were log-transformed to handle the skewed distributions of these variables. Due to the large number of variables in the full models under consideration, Akaike information criterion (AIC) was used to select the subset of markers that best predicted each brain volume25. Final linear regression models were selected by minimizing AIC, which takes into account both the goodness of fit and the model complexity.
Results
Table 3 shows the unstandardized regression coefficients and associated p-values for clinical and cytokine markers that were included in the final regression model following the model selection algorithm for each brain volumetric measure. Results are also presented graphically in Figure 2. As expected, intracranial volume was a highly significant covariate and was included in all final models to control for head size.
Table 3.
Unstandardized regression coefficients and associated p-values (in parentheses) for clinical and cytokine markers included in the final model selected by minimizing AIC for each brain volumetric measure. Rows indicate the predictors under consideration, and each final regression model is represented by one column of the table. Statistically significant covariates are highlighted in grey. AIC and adjusted R2 values of the final models are included in the final rows.
| Cortex | White Matter | Caudate | Putamen | Globus Pallidus | Thalamus | Hippo-campus | Amygdala | Ventricles | |
|---|---|---|---|---|---|---|---|---|---|
| Age | -1.95e+3 (<.0001) | -7.30e+2 (0.0635) | -1.62e+1 (0.0764) | -4.30e+1 (0.0001) | -1.18e+1 (0.0019) | -5.81e+1 (<.0001) | -2.10e+1 (0.0093) | -8.44e+0 (0.0428) | 4.41e+2 (<.0001) |
| CD4 current | -5.60e-1 (0.1468) | 2.78e-1 (0.0626) | 7.59e+0 (0.0353) | ||||||
| CD4 nadir | 1.75e+2 (0.0204) | 4.73e+1 (0.0795) | 1.97e+2 (0.0311) | 1.21e+2 (0.0445) | |||||
| HIV RNA | |||||||||
| HIV duration | -1.21e+3 (0.0355) | -3.18e+1 (0.0672) | |||||||
| CART | 2.58e+4 (0.0166) | 6.86e+2 (0.0342) | 6.35e+2 (0.0053) | ||||||
| HCV | 3.48e+2 (0.0964) | 4.96e+2 (0.0361) | 3.48e+2 (0.0009) | ||||||
| IFN-γ | 2.23e+4 (0.0114) | 2.41e+4 (0.0306) | 4.79e+2 (0.1370) | 4.24e+2 (0.0687) | |||||
| IL1-β | -1.22e+4 (0.1490) | -2.32e+2 (0.0069) | |||||||
| IL6 | -2.19e+4 (0.0095) | -6.21e+2 (0.0030) | -8.36e+2 (0.0021) | -4.82e+2 (0.0081) | 6.56e+3 (0.0003) | ||||
| IL8 | |||||||||
| IL10 | -2.53e+2 I (0.0526) | -9.03e+3 (0.0190) | |||||||
| IL16 | -2.48e+4 (0.0001) | -2.55e+2 (0.1253) | -6.63e+2 (0.0014) | -2.18e+2 (0.0038) | -2.37e+2 (0.1263) | -2.04e+2 (0.0091) | |||
| IL18 | -1.21e+4 (0.0023) | 2.37e+2 (0.1184) | 1.52e+2 (0.1436) | ||||||
| IP10 | 3.82e+3 (0.1153) | 4.99e+3 (0.1240) | -5.15e+1 (0.1450) | ||||||
| MCP1 | 7.44e+3 (0.0924) | 4.01e+2 (0.0127) | 1.38e+2 (0.0150) | 3.03e+2 (0.0691) | 1.29e+2 (0.0218) | ||||
| MIP-1β | 1.39e+2 (0.1325) | 2.47e+3 (0.0184) | |||||||
| SDF1-α | |||||||||
| TNF-α | |||||||||
| TRAIL | 4.46e+2 (0.0663) | ||||||||
| ICV | 2.31e-1 (<.0001) | 2.70e-1 (<0001) | 4.87e-3 (<.0001) | 4.51e-3 (<.0001) | 1.64e-3 (<.0001) | 7.57e-3 (<.0001) | 3.78e-3 (<0001) | 1.63e-3 (<.0001) | 2.17e-2 (0.0001) |
| Final AIC Adjusted R2 | 1494.88 0.8377 | 1528.72 0.7692 | 977.03 0.6283 | 999.36 0.6764 | 842.38 0.6289 | 1016.12 0.7893 | 959.31 0.6477 | 865.53 0.5357 | 1336.36 0.4142 |
Note: ICV = intracranial volume.
Figure 2.
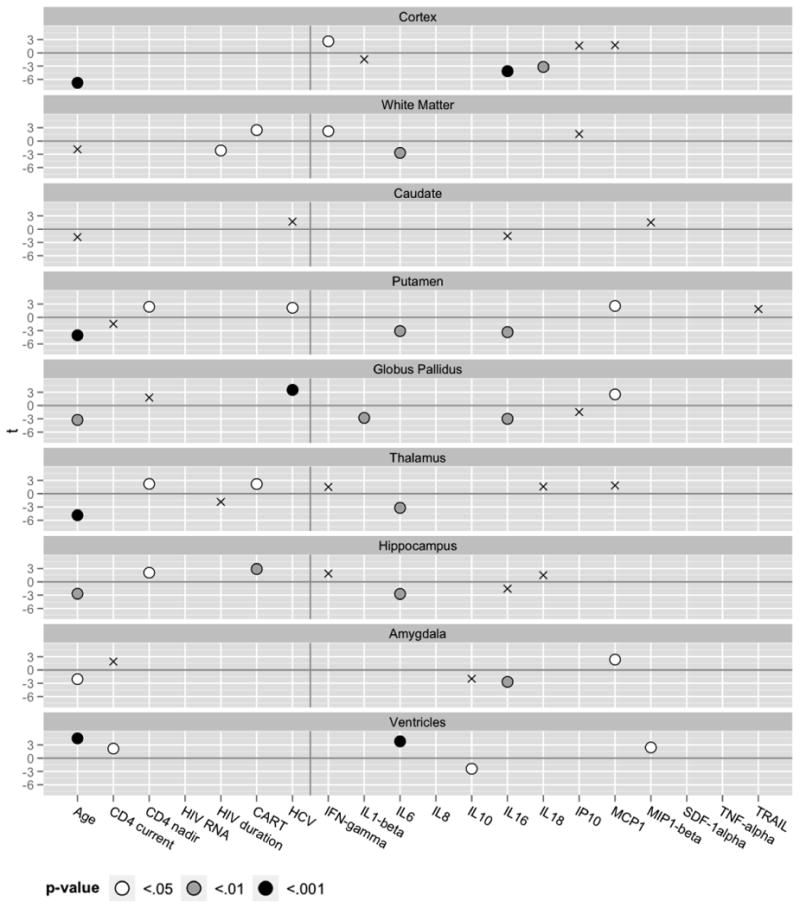
Final linear regression models selected by minimizing AIC. Each model includes the subset of clinical and cytokine markers that best predict each brain volumetric measure. Significant predictors are shaded to indicate p-value levels (see legend). Non-significant predictors retained in the models are marked with crosses (X). Regression coefficients were transformed to t-statistics to scale the plots for visualization. Refer to Table 3 for individual regression coefficient estimates and associated p-values of all covahates.
Global Volumetric Measures
Cortical Grey Matter
Lower GM volume was significantly associated with older age, in addition to lower level of IFN-γ and higher levels of IL16 and IL18.
Cerebral White Matter
Lower WM volume was significantly associated with longer duration of HIV infection, lack of current CART, in addition to lower IFN-γ level and higher IL6 level.
Ventricles
Higher ventricular volume was significantly associated with age and higher current CD4 level, in addition to higher levels of IL6 and MIP-1β, and lower IL10 level.
Subcortical and Medial Temporal Structures
Caudate
Caudate nucleus volume was not significantly associated with any clinical or cytokine markers.
Putamen
Lower putamen volume was significantly associated with older age, lower CD4 nadir level, undetectable HCV RNA, in addition to higher levels of IL6 and IL16, and lower MCP1 level.
Globus Pallidus
Lower pallidum volume was significantly associated with older age, undetectable HCV RNA, in addition to higher levels of IL1-β and IL16, and lower MCP1 level.
Thalamus
Lower thalamus volume was significantly associated with older age, lower CD4 nadir level, and lack of current CART, in addition to higher IL6 level.
Hippocampus
Lower hippocampal volume was significantly associated with older age, lower CD4 nadir level, lack of current CART, in addition to higher IL6 level.
Amygdala
Lower amygdala volume was significantly associated with older age (p=0.0428), in addition to higher IL16 level and lower MCP1 level.
Impact of Lifetime Alcohol and Substance Use Disorders
To further examine the possible influence of more distant alcohol/substance use history, we repeated the regression analyses predicting each volumetric measure with an additional binary covariate indicating lifetime alcohol or substance dependence history based on DSM-IV criteria. Following the model selection algorithm, this variable was retained in models predicting volumes of the amygdala (β=135.97, p=0.1420), putamen (β=-430.05, p=0.0797), and thalamus (β=797.85, p=0.0018). The addition of this variable did not alter the patterns of the volumetric relationships with other predictors in these three models, while all other models remain identical to those reported above.
Discussion
Initial optimism that CART would lead to an eradication of HIV-associated brain dysfunction has been tempered by persistent neurocognitive impairment observed in people living with HIV despite virologic suppression. Chronic systemic immune activation and inflammation have been among the mechanisms proposed for these disturbances, though the underlying pathophysiology is still not fully understood. Using a multivariate statistical approach, we examined the independent contributions of age, clinical status, and an extensive panel of plasma cytokine markers on brain volumetric alterations in this cohort of 74 medically stable individuals with chronic HIV infection. Our findings corroborate existing evidence demonstrating pervasive brain atrophy in the context of HIV disease and older age5, 26. These structural alterations appear to be partially mitigated by antiretroviral treatment. Cytokine levels, most notably IL-6, IL-16, and MCP-1, were significantly associated with brain volumes even after accounting for the effects of the above clinical variables. The current findings demonstrated the potential utility of these markers both in elucidating the pathophysiological mechanisms underlying brain damage associated with HIV infection, and as systemic markers of HIV-associated brain dysfunction in the CART era.
A major strength of this study is the use of multiplexed assay to simultaneously investigate an extensive list of markers potentially relevant to brain structural abnormalities. Our findings extend those from previous studies in a number of ways. This is among the first studies to demonstrate associations between plasma cytokine and structural brain alterations, in addition to previously reported associations to neurocognitive dysfunction18, 20. To our knowledge, one published study has examined this link with brain volumetric alteration in a relatively small cohort of 10 HIV-infected individuals22. In addition to expanding the sample size, we also examined volumes of specific subcortical nuclei and medial temporal structures, along with the global volumetric measure of the grey/white matter and the ventricles as were reported in the previous study. The use of a multivariate model selection approach permitted the independent contributions and relative importance of each marker to be examined, while ensuring parsimony of the statistical models.
IL-6 and IL-16 emerged as the most robust cytokine predictors of smaller brain volumes in these models, together accounting for alterations in virtually all brain regions examined. These findings are consistent with expectation given the principal roles of these molecules in systemic immune responses and inflammatory processes. IL-16 in particular is an important chemoattractant cytokine for CD4+ T cells and contributes to the regulatory process of recruitment and activation of T cells at inflammation sites27. Corroborating the current findings, a previous study from our group showed IL-16 elevation to be among the strongest predictors of impairment in attention and executive and psychomotor functions18. IL-6 functions as both a pro- and anti-inflammatory cytokine and has been associated with HIV RNA level and implicated as a mediator of activation-induced CD4+ T cell losses28, 29. Although CSF IL-6 level has been linked to neurocognitive impairment in treatment-naive HIV-infected individuals30, to our knowledge, no previous studies have shown associations between systemic IL-6 level and structural brain measures in the context of HIV. Less widespread associations were found between higher IL1-β, IL18, and MIP-1β levels and smaller volumes of the globus pallidus and cerebral cortex, and larger ventricular volume. These latter findings are likewise novel and should be considered in the context of future replicating studies.
Higher MCP-1 levels were associated with greater basal ganglia and amygdala volumes. This is contrary to expectation given previous findings linking neurocognitive impairment and cerebral white matter and metabolite abnormalities to increased MCP-1 levels both in plasma18, 22, 31 and CSF 23, 32. Plasma level of MCP-1 has also been positively related to HIV RNA level33. MCP-1 is an important chemotactic factor for monocytes and CD4+ T cells, and serves to recruit immune cells to the site of inflammation34. Given its direct involvement with these immune cells, in addition to the fact that monocytes/macrophages are major producers of these molecules, MCP-1 is believed to have important roles in HIV pathogenesis35. While the basis for the positive relationships between MCP-1 and brain volumes cannot be ascertained from this study, it is possible that these findings reflect hypertrophy resulting from fluid accumulation secondary to inflammation of these brain structures. This possibility is further supported by findings suggesting that MCP-1 in plasma may largely be derived originally from within the CNS 36.
Among the HIV disease markers, nadir CD4 suppression was most consistently associated with smaller brain volume. This is consistent with emerging literature indicating the effects of distant history of immune suppression on current brain integrity5, 11, 12. These findings relate to the possibility that viral replication and the resulting immune suppression occurring at an early stage of HIV disease have long term effects on the brain that may persist even after effective viral suppression and immune reconstitution. It is notable that the current findings contrast with recently published results from our group indicating a lack of association between nadir CD4 and diffusion tensor MRI measures of white matter integrity13. This dissociation may reflect differential effects of distant immunologic history on macroscopic (i.e., volumetric alteration) versus microstructural (i.e., cellular water diffusion) measures of brain integrity. Although it is possible that chronic measures such as nadir CD4 may also be a proxy for secondary factors such as poor access to health care, comorbid medical conditions, and socioeconomic status, these findings suggest the potentially beneficial role of early antiretroviral treatment to prevent significant immune suppression.
The finding related to duration of HIV infection is consistent with existing evidence linking general brain atrophy to infection chronicity9, 37. Specifically, our finding in this medically stable, primarily CART-treated cohort suggests particular susceptibility of cerebral white matter to chronic infection. The positive association between current CD4 and ventricular volume was contrary to expectation, and needs to be further examined in future studies. As expected, lack of antiretroviral treatment was associated with smaller brain volumes, including those of the white matter, thalamus, and hippocampus. These findings suggest a beneficial role of CART in mitigating brain atrophy in the context of HIV infection. It should be noted that the observed relationships with CART were not in the direction of antiretroviral toxicity as has been reported in recent studies38.
To further examine the impact of CART history in this chronically infected cohort, we classified participants based on estimated year of infection. Using 1995 as the cut point for CART availability, 41% of our sample was diagnosed with HIV pre-CART. As expected, individuals diagnosed prior to 1995 were significantly more likely to have history of immune suppression: 77% of pre-CART individuals vs. 41% of those diagnosed in the CART era presented with CD4 nadir < 200, χ2=7.8404, p=0.0051. It should be noted that while this difference in immune suppression history between the groups is likely related to CART availability at the time of diagnosis, it is likely also impacted by the longer duration of illness in the pre-CART group (t=16.08, p<.0001). We repeated the regression analyses reported above with an additional binary covariate indicating an HIV diagnosis before or after 1995. Interestingly, this additional predictor did not show significant relationships with any of the volumetric variables. While it is possible that these statistical outcomes were impacted by multicollinearity with HIV disease factors discussed above, it should be noted that our binary classifier based on diagnosis year likely failed to capture the complex and diverse antiretroviral treatment time course among these individuals, and future studies with more comprehensive assessment of ART history are needed to adequately examine this important question.
HCV coinfection was found to be associated with larger volumes of basal ganglia structures. Basal ganglia hypertrophy has been previously reported in the context of substance abuse 39, 40, and it is possible that the current findings reflect analogous neuroinflammatory processes specifically affecting these structures. HCV infection has been linked to microglial activation and neuroinflammation 41. The extent to which such events are contributed by direct viral infiltration of the CNS or indirectly through HCV-associated hepatic disturbances is unclear. However, multiple lines of evidence indicating direct HCV neuroinvasion have recently emerged 42, pointing to the importance of virologic control in preventing brain dysfunction. Regardless of the underlying mechanism, existing findings clearly indicate the presence of neuroinflammatory processes in HCV-infected individuals43-45. The pathophysiological basis of brain dysfunction associated with HCV infection, particularly in the context of HIV coinfection, remains to be fully delineated. Previous findings from our group and others have linked HIV/HCV coinfection to increased neurocognitive impairment14-17 and cerebral white matter abnormalities 5, 13. Additional studies are needed specifically examining neuroinflammation in the context of HIV/HCV coinfection.
While the current analyses excluded individuals with alcohol or substance use disorder within 6 months prior to the study, the possible influence of more distant alcohol/substance use history cannot be ruled out. To further examine this, we repeated the regression analyses predicting each volumetric measure with an additional binary covariate indicating lifetime alcohol or substance dependence history. Following the model selection algorithm, this variable was a significant predictor only of thalamic volume. While hypertrophy of the basal ganglia in the context of substance abuse has been reported 39, 40, the significant positive relationship between thalamic volume and lifetime alcohol/substance dependence in this cohort is a noteworthy and novel finding. Future studies that include more detailed assessment of distant alcohol and substance use are needed to further address this important question.
Not surprisingly, older age was among the most robust predictors of smaller volumes in virtually all measured brain structures. Although these findings are consistent with the substantial literature on age-related brain atrophy 46-48, it is possible that the robust associations observed here may reflect the augmented effects of aging in the context of HIV such as has been suggested49, 50. Future studies are needed that are adequately powered to examine the possibility of an HIV-associated accelerated aging phenomenon relevant to structural and functional brain changes.
In summary, the current findings provide novel evidence that cortical, subcortical and white matter volumes vary as a function of plasma cytokine levels among people with chronic HIV infection. These results from a cohort with an average infection duration of about 13 years corroborate an emerging body of evidence linking HIV-associated chronic systemic immune activation and inflammatory processes to compromises in brain integrity51-53. The fact that these cerebral correlates were observed for plasma cytokine levels is noteworthy, and indicates potential utility of these peripheral markers in predicting and monitoring brain alterations associated with chronic HIV infection. The relative importance of specific cytokines to brain structural integrity demonstrated here also contributes to the current understanding of mechanisms underlying HIV neuropathogenesis, which may aid in the development of beneficial therapies for HIV-associated neurocognitive dysfunction.
Inferences from this study are limited by its cross-sectional and observation nature. Future longitudinal studies are necessary to examine the causal relationships between the immunologic and inflammatory markers with structural and functional brain measures, especially in the context of antiretroviral treatment, aging, and comorbid conditions including HCV infection. Simultaneous examinations of the CNS and systemic trajectories of cytokine levels, in addition to those of other biomarkers, will also be important to further elucidate the underlying mechanisms of abnormal brain alterations in people living with HIV. Finally, novel neuroimaging markers are rapidly being developed that enable concurrent examination of multiple aspects of brain integrity, including those relevant to structural and functional connectivities and vascular integrity, which together with other biomarkers will help to extend the current understanding of the underlying pathophysiological mechanisms of HIV-associated brain dysfunction.
Acknowledgments
The research described was supported by NIH grants R00AA020235, R01MH074368, P01AA019072, and P30AI042853.
Footnotes
The authors declare that they have no conflict of interest.
References
- 1.Tozzi V, Balestra P, Bellagamba R, Corpolongo A, Salvatori MF, Visco-Comandini U, Vlassi C, Giulianelli M, Galgani S, Antinori A, Narciso P. Persistence of neuropsychologic deficits despite long-term highly active antiretroviral therapy in patients with HIV-related neurocognitive impairment: prevalence and risk factors. J Acquir Immune Defic Syndr. 2007 Jun 1;45(2):174–182. doi: 10.1097/QAI.0b013e318042e1ee. [DOI] [PubMed] [Google Scholar]
- 2.Heaton RK, Clifford DB, Franklin DR, Jr, Woods SP, Ake C, Vaida F, Ellis RJ, Letendre SL, Marcotte TD, Atkinson JH, Rivera-Mindt M, Vigil OR, Taylor MJ, Collier AC, Marra CM, Gelman BB, McArthur JC, Morgello S, Simpson DM, McCutchan JA, Abramson I, Gamst A, Fennema-Notestine C, Jernigan TL, Wong J, Grant I. HIV-associated neurocognitive disorders persist in the era of potent antiretroviral therapy: CHARTER Study. Neurology. 2010 Dec 7;75(23):2087–2096. doi: 10.1212/WNL.0b013e318200d727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Cardenas VA, Meyerhoff DJ, Studholme C, Kornak J, Rothlind J, Lampiris H, Neuhaus J, Grant RM, Chao LL, Truran D, Weiner MW. Evidence for ongoing brain injury in human immunodeficiency virus-positive patients treated with antiretroviral therapy. Journal of neurovirology. 2009 Jul;15(4):324–333. doi: 10.1080/13550280902973960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Harezlak J, Buchthal S, Taylor M, Schifitto G, Zhong J, Daar E, Alger J, Singer E, Campbell T, Yiannoutsos C, Cohen R, Navia B. Persistence of HIV-associated cognitive impairment, inflammation, and neuronal injury in era of highly active antiretroviral treatment. AIDS. 2011 Mar 13;25(5):625–633. doi: 10.1097/QAD.0b013e3283427da7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Jernigan TL, Archibald SL, Fennema-Notestine C, Taylor MJ, Theilmann RJ, Julaton MD, Notestine RJ, Wolfson T, Letendre SL, Ellis RJ, Heaton RK, Gamst AC, Franklin DR, Jr, Clifford DB, Collier AC, Gelman BB, Marra C, McArthur JC, McCutchan JA, Morgello S, Simpson DM, Grant I. Clinical factors related to brain structure in HIV: the CHARTER study. Journal of neurovirology. 2011 Jun;17(3):248–257. doi: 10.1007/s13365-011-0032-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gongvatana A, Schweinsburg BC, Taylor MJ, Theilmann RJ, Letendre SL, Alhassoon OM, Jacobus J, Woods SP, Jernigan TL, Ellis RJ, Frank LR, Grant I. White matter tract injury and cognitive impairment in human immunodeficiency virus-infected individuals. Journal of neurovirology. 2009 Apr;15(2):187–195. doi: 10.1080/13550280902769756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Roc AC, Anees BM, Chawla S, Korczykowski M, Wolf RL, Kolson DL, Detre JA, Poptani H. Detection of human immunodeficiency virus induced inflammation and oxidative stress in lenticular nuclei with magnetic resonance spectroscopy despite antiretroviral therapy. Archives of neurology. 2007 Sep;64(9):1249–1257. doi: 10.1001/archneur.64.9.noc60125. [DOI] [PubMed] [Google Scholar]
- 8.Anees BM, Sisti D, Vaida F, Liang CL, Leontiev O, Perthen JE, Buxton RB, Benson D, Smith DM, Little SJ, Richman DD, Moore DJ, Ellis RJ. Resting cerebral blood flow: a potential biomarker of the effects of HIV in the brain. Neurology. 2009 Sep 1;73(9):702–708. doi: 10.1212/WNL.0b013e3181b59a97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Becker JT, Sanders J, Madsen SK, Ragin A, Kingsley L, Maruca V, Cohen B, Goodkin K, Martin E, Miller EN, Sacktor N, Alger JR, Barker PB, Saharan P, Carmichael OT, Thompson PM. Subcortical brain atrophy persists even in HAART-regulated HIV disease. Brain imaging and behavior. 2011 Jun;5(2):77–85. doi: 10.1007/s11682-011-9113-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lepore N, Brun C, Chou YY, Chiang MC, Dutton RA, Hayashi KM, Luders E, Lopez OL, Aizenstein HJ, Toga AW, Becker JT, Thompson PM. Generalized tensor-based morphometry of HIV/AIDS using multivariate statistics on deformation tensors. IEEE transactions on medical imaging. 2008 Jan;27(1):129–141. doi: 10.1109/TMI.2007.906091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cohen RA, Harezlak J, Schifitto G, Hana G, Clark U, Gongvatana A, Paul R, Taylor M, Thompson P, Alger J, Brown M, Zhong J, Campbell T, Singer E, Daar E, McMahon D, Tso Y, Yiannoutsos CT, Navia B. Effects of nadir CD4 count and duration of human immunodeficiency virus infection on brain volumes in the highly active antiretroviral therapy era. Journal of neurovirology. 2010 Feb;16(1):25–32. doi: 10.3109/13550280903552420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ellis RJ, Badiee J, Vaida F, Letendre S, Heaton RK, Clifford D, Collier AC, Gelman B, McArthur J, Morgello S, McCutchan JA, Grant I. CD4 nadir is a predictor of HIV neurocognitive impairment in the era of combination antiretroviral therapy. AIDS. 2011 Sep 10;25(14):1747–1751. doi: 10.1097/QAD.0b013e32834a40cd. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gongvatana A, Cohen RA, Correia S, Devlin KN, Miles J, Kang H, Ombao H, Navia B, Laidlaw DH, Tashima KT. Clinical contributors to cerebral white matter integrity in HIV-infected individuals. Journal of neurovirology. 2011 Oct;17(5):477–486. doi: 10.1007/s13365-011-0055-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Cherner M, Letendre S, Heaton RK, Durelle J, Marquie-Beck J, Gragg B, Grant I. Hepatitis C augments cognitive deficits associated with HIV infection and methamphetamine. Neurology. 2005 Apr 26;64(8):1343–1347. doi: 10.1212/01.WNL.0000158328.26897.0D. [DOI] [PubMed] [Google Scholar]
- 15.Clifford DB, Evans SR, Yang Y, Gulick RM. The neuropsychological and neurological impact of hepatitis C virus co-infection in HIV-infected subjects. AIDS. 2005 Oct;19(Suppl 3):S64–71. doi: 10.1097/01.aids.0000192072.80572.43. [DOI] [PubMed] [Google Scholar]
- 16.Hinkin CH, Castellon SA, Levine AJ, Barclay TR, Singer EJ. Neurocognition in individuals co-infected with HIV and hepatitis C. J Addict Dis. 2008;27(2):11–17. doi: 10.1300/j069v27n02_02. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Devlin KN, Gongvatana A, Clark US, Chasman JD, Westbrook ML, Tashima KT, Navia B, Cohen RA. Neurocognitive effects of HIV, hepatitis C, and substance use history. Journal of the International Neuropsychological Society : JINS. 2012 Jan;18(1):68–78. doi: 10.1017/S1355617711001408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cohen RA, de la Monte S, Gongvatana A, Ombao H, Gonzalez B, Devlin KN, Navia B, Tashima KT. Plasma cytokine concentrations associated with HIV/hepatitis C coinfection are related to attention, executive and psychomotor functioning. Journal of neuroimmunology. 2011 Apr;233(1-2):204–210. doi: 10.1016/j.jneuroim.2010.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Burdo TH, Weiffenbach A, Woods SP, Letendre S, Ellis RJ, Williams KC. Elevated sCD163 is a marker of neurocognitive impairment in HIV infection. AIDS. 2013 Feb 21; doi: 10.1097/QAD.0b013e32836010bd. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ryan LA, Zheng J, Brester M, Bohac D, Hahn F, Anderson J, Ratanasuwan W, Gendelman HE, Swindells S. Plasma levels of soluble CD14 and tumor necrosis factor-alpha type II receptor correlate with cognitive dysfunction during human immunodeficiency virus type 1 infection. J Infect Dis. 2001 Sep 15;184(6):699–706. doi: 10.1086/323036. [DOI] [PubMed] [Google Scholar]
- 21.Sevigny JJ, Albert SM, McDermott MP, McArthur JC, Sacktor N, Conant K, Schifitto G, Seines OA, Stern Y, McClernon DR, Palumbo D, Kieburtz K, Riggs G, Cohen B, Epstein LG, Marder K. Evaluation of HIV RNA and markers of immune activation as predictors of HIV-associated dementia. Neurology. 2004 Dec 14;63(11):2084–2090. doi: 10.1212/01.wnl.0000145763.68284.15. [DOI] [PubMed] [Google Scholar]
- 22.Ragin AB, Wu Y, Ochs R, Scheidegger R, Cohen BA, Edelman RR, Epstein LG, McArthur J. Biomarkers of neurological status in HIV infection: a 3-year study. Proteomics Clinical applications. 2010 Mar;4(3):295–303. doi: 10.1002/prca.200900083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Letendre SL, Zheng JC, Kaul M, Yiannoutsos CT, Ellis RJ, Taylor MJ, Marquie-Beck J, Navia B. Chemokines in cerebrospinal fluid correlate with cerebral metabolite patterns in HIV-infected individuals. Journal of neurovirology. 2011 Feb;17(1):63–69. doi: 10.1007/s13365-010-0013-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Fischl B, Salat DH, van der Kouwe AJ, Makris N, Segonne F, Quinn BT, Dale AM. Sequence-independent segmentation of magnetic resonance images. Neuroimage. 2004;23(Suppl 1):S69–84. doi: 10.1016/j.neuroimage.2004.07.016. [DOI] [PubMed] [Google Scholar]
- 25.Akaike H. A new look at the statistical model identification. IEEE Transactions on Automatic Control. 1974;19(6):716–723. [Google Scholar]
- 26.Lockhart SN, DeCarli C. Structural Imaging Measures of Brain Aging. Neuropsychology review. 2014 Aug 22; doi: 10.1007/s11065-014-9268-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Cruikshank WW, Kornfeld H, Center DM. lnterleukin-16. Journal of leukocyte biology. 2000 Jun;67(6):757–766. doi: 10.1002/jlb.67.6.757. [DOI] [PubMed] [Google Scholar]
- 28.Lederman MM, Kalish LA, Asmuth D, Fiebig E, Mileno M, Busch MP. 'Modeling' relationships among HIV-1 replication, immune activation and CD4+ T-cell losses using adjusted correlative analyses. AIDS. 2000 May 26;14(8):951–958. doi: 10.1097/00002030-200005260-00006. [DOI] [PubMed] [Google Scholar]
- 29.Serieller J, Chalaris A, Schmidt-Arras D, Rose-John S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochimica et biophysica acta. 2011 May;1813(5):878–888. doi: 10.1016/j.bbamcr.2011.01.034. [DOI] [PubMed] [Google Scholar]
- 30.Airoldi M, Bandera A, Trabattoni D, Tagliabue B, Arosio B, Soria A, Rainone V, Lapadula G, Annoni G, Clerici M, Gori A. Neurocognitive impairment in HIV-infected naive patients with advanced disease: the role of virus and intrathecal immune activation. Clinical & developmental immunology. 2012;2012:467154. doi: 10.1155/2012/467154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Woods SP, Morgan EE, Marquie-Beck J, Carey CL, Grant I, Letendre SL. Markers of macrophage activation and axonal injury are associated with prospective memory in HIV-1 disease. Cognitive and behavioral neurology : official journal of the Society for Behavioral and Cognitive Neurology. 2006 Dec;19(4):217–221. doi: 10.1097/01.wnn.0000213916.10514.57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Yuan L, Qiao L, Wei F, Yin J, Liu L, Ji Y, Smith D, Li N, Chen D. Cytokines in CSF correlate with HIV-associated neurocognitive disorders in the post-HAART era in China. Journal of neurovirology. 2013 Apr;19(2):144–149. doi: 10.1007/s13365-013-0150-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Weiss L, Si-Mohamed A, Giral P, Castiel P, Ledur A, Blondin C, Kazatchkine MD, Haeffner-Cavaillon N. Plasma levels of monocyte chemoattractant protein-1 but not those of macrophage inhibitory protein-1 alpha and RANTES correlate with virus load in human immunodeficiency virus infection. J Infect Dis. 1997 Dec;176(6):1621–1624. doi: 10.1086/517341. [DOI] [PubMed] [Google Scholar]
- 34.Ajuebor MN, Flower RJ, Hannon R, Christie M, Bowers K, Verity A, Perretti M. Endogenous monocyte chemoattractant protein-1 recruits monocytes in the zymosan peritonitis model. Journal of leukocyte biology. 1998 Jan;63(1):108–116. doi: 10.1002/jlb.63.1.108. [DOI] [PubMed] [Google Scholar]
- 35.Deshmane SL, Kremlev S, Amini S, Sawaya BE. Monocyte chemoattractant protein-1 (MCP-1): an overview. Journal of interferon & cytokine research : the official journal of the International Society for Interferon and Cytokine Research. 2009 Jun;29(6):313–326. doi: 10.1089/jir.2008.0027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Monteiro de Almeida S, Letendre S, Zimmerman J, Lazzaretto D, McCutchan A, Ellis R. Dynamics of monocyte chemoattractant protein type one (MCP-1) and HIV viral load in human cerebrospinal fluid and plasma. Journal of neuroimmunology. 2005 Dec;169(1-2):144–152. doi: 10.1016/j.jneuroim.2005.07.012. [DOI] [PubMed] [Google Scholar]
- 37.Elovaara I, Poutiainen E, Raininko R, Valanne L, Virta A, Valle SL, Lahdevirta J, livanainen M. Mild brain atrophy in early HIV infection: the lack of association with cognitive deficits and HIV-specific intrathecal immune response. Journal of the neurological sciences. 1990 Nov;99(2-3):121–136. doi: 10.1016/0022-510x(90)90149-h. [DOI] [PubMed] [Google Scholar]
- 38.Ciccarelli N, Fabbiani M, Di Giambenedetto S, Fanti I, Baldonero E, Bracciale L, Tamburrini E, Cauda R, De Luca A, Silveri MC. Efavirenz associated with cognitive disorders in otherwise asymptomatic HIV-infected patients. Neurology. 2011 Apr 19;76(16):1403–1409. doi: 10.1212/WNL.0b013e31821670fb. [DOI] [PubMed] [Google Scholar]
- 39.Jernigan TL, Gamst AC, Archibald SL, Fennema-Notestine C, Mindt MR, Marcotte TD, Heaton RK, Ellis RJ, Grant I. Effects of methamphetamine dependence and HIV infection on cerebral morphology. The American journal of psychiatry. 2005 Aug;162(8):1461–1472. doi: 10.1176/appi.ajp.162.8.1461. [DOI] [PubMed] [Google Scholar]
- 40.Jacobsen LK, Giedd JN, Gottschalk C, Kosten TR, Krystal JH. Quantitative morphology of the caudate and putamen in patients with cocaine dependence. The American journal of psychiatry. 2001 Mar;158(3):486–489. doi: 10.1176/appi.ajp.158.3.486. [DOI] [PubMed] [Google Scholar]
- 41.Grover VP, Pavese N, Koh SB, Wylezinska M, Saxby BK, Gerhard A, Forton DM, Brooks DJ, Thomas HC, Taylor-Robinson SD. Cerebral microglial activation in patients with hepatitis C: in vivo evidence of neuroinflammation. J Viral Hepat. 2012 Feb;19(2):e89–96. doi: 10.1111/j.1365-2893.2011.01510.x. [DOI] [PubMed] [Google Scholar]
- 42.Laskus T, Radkowski M, Adair DM, Wilkinson J, Scheck AC, Rakela J. Emerging evidence of hepatitis C virus neuroinvasion. AIDS. 2005 Oct;19(Suppl 3):S140–144. doi: 10.1097/01.aids.0000192083.41561.00. [DOI] [PubMed] [Google Scholar]
- 43.Bokemeyer M, Ding XQ, Goldbecker A, Raab P, Heeren M, Arvanitis D, Tillmann HL, Lanfermann H, Weissenborn K. Evidence for neuroinflammation and neuroprotection in HCV infection-associated encephalopathy. Gut. 2011 Mar;60(3):370–377. doi: 10.1136/gut.2010.217976. [DOI] [PubMed] [Google Scholar]
- 44.Forton D, Allsop J, Main J, Foster G, Thomas H, Taylor-Robins S. Evidence for a cerebral effect of the hepatitis C virus. Lancet. 2001:38–39. doi: 10.1016/S0140-6736(00)05270-3. [DOI] [PubMed] [Google Scholar]
- 45.Forton DM, Hamilton G, Allsop JM, Grover VP, Wesnes K, O'Sullivan C, Thomas HC, Taylor-Robinson SD. Cerebral immune activation in chronic hepatitis C infection: a magnetic resonance spectroscopy study. Journal of hepatology. 2008 Sep;49(3):316–322. doi: 10.1016/j.jhep.2008.03.022. [DOI] [PubMed] [Google Scholar]
- 46.Zimmerman ME, Brickman AM, Paul RH, Grieve SM, Tate DF, Gunstad J, Cohen RA, Aloia MS, Williams LM, Clark CR, Whitford TJ, Gordon E. The relationship between frontal gray matter volume and cognition varies across the healthy adult lifespan. Am J Geriatr Psychiatry. 2006 Oct;14(10):823–833. doi: 10.1097/01.JGP.0000238502.40963.ac. [DOI] [PubMed] [Google Scholar]
- 47.Brickman AM, Zimmerman ME, Paul RH, Grieve SM, Tate DF, Cohen RA, Williams LM, Clark CR, Gordon E. Regional white matter and neuropsychological functioning across the adult lifespan. Biol Psychiatry. 2006 Sep 1;60(5):444–453. doi: 10.1016/j.biopsych.2006.01.011. [DOI] [PubMed] [Google Scholar]
- 48.Raz N, Ghisletta P, Rodrigue KM, Kennedy KM, Lindenberger U. Trajectories of brain aging in middle-aged and older adults: regional and individual differences. Neuroimage. 2010 Jun;51(2):501–511. doi: 10.1016/j.neuroimage.2010.03.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Valcour V, Shikuma C, Shiramizu B, Waiters M, Poff P, Seines O, Hoick P, Grove J, Sacktor N. Higher frequency of dementia in older HIV-1 individuals: the Hawaii Aging with HIV-1 Cohort. Neurology. 2004 Sep 14;63(5):822–827. doi: 10.1212/01.wnl.0000134665.58343.8d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kirk JB, Goetz MB. Human immunodeficiency virus in an aging population, a complication of success. Journal of the American Geriatrics Society. 2009 Nov;57(11):2129–2138. doi: 10.1111/j.1532-5415.2009.02494.x. [DOI] [PubMed] [Google Scholar]
- 51.Williams K, Westmoreland S, Greco J, Ratai E, Lentz M, Kim WK, Fuller RA, Kim JP, Autissier P, Sehgal PK, Schinazi RF, Bischofberger N, Piatak M, Lifson JD, Masliah E, Gonzalez RG. Magnetic resonance spectroscopy reveals that activated monocytes contribute to neuronal injury in SIV neuroAIDS. The Journal of clinical investigation. 2005 Sep;115(9):2534–2545. doi: 10.1172/JCI22953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Lentz MR, Kim WK, Lee V, Bazner S, Halpern EF, Venna N, Williams K, Rosenberg ES, Gonzalez RG. Changes in MRS neuronal markers and T cell phenotypes observed during early HIV infection. Neurology. 2009 Apr 28;72(17):1465–1472. doi: 10.1212/WNL.0b013e3181a2e90a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Pulliam L, Rempel H, Sun B, Abadjian L, Calosing C, Meyerhoff DJ. A peripheral monocyte interferon phenotype in HIV infection correlates with a decrease in magnetic resonance spectroscopy metabolite concentrations. AIDS. 2011 Sep 10;25(14):1721–1726. doi: 10.1097/QAD.0b013e328349f022. [DOI] [PMC free article] [PubMed] [Google Scholar]