

Abstract
The underlying factors related to the divergent findings of age-related fatigue for dynamic tasks are not well understood. The purpose here was to investigate age-related fatigability and recovery between a repeated constrained (isokinetic) and an unconstrained velocity (isotonic) task, in which participants performed fatiguing contractions at the velocity (isokinetic) or resistance (isotonic) corresponding with maximal power. To compare between tasks, isotonic torque–power relationships were constructed prior to and following both fatiguing tasks and during short-term recovery. Contractile properties were recorded from 9 old (~75 years) and 11 young (~25 years) men during three testing sessions. In the first session, maximal power was assessed, and sessions 2 and 3 involved an isokinetic or an isotonic concentric fatigue task performed until maximal power was reduced by 40 %. Compared with young, the older men performed the same number of contractions to task failure for the isokinetic task (~45 contractions), but 20 % fewer for the isotonic task (p < 0.05). Regardless of age and task, maximal voluntary isometric contraction strength, angular velocity, and power were reduced by ~30, ~13, and ~25 %, respectively, immediately following task failure, and only isometric torque was not recovered fully by 10 min. In conclusion, older men are more fatigable than the young when performing a repetitive maximal dynamic task at a relative resistance (isotonic) but not an absolute velocity (isokinetic), corresponding to maximal power.
Keywords: Isotonic, Isokinetic, Aging, Force–velocity, Sarcopenia, Recovery
Introduction
Natural adult aging is accompanied by various alterations within the neuromuscular system leading to reduced neural function, muscle mass, and quality, resulting in weakness, decreased maximal shortening velocity, and ultimately less power (Aagaard et al. 2010; Power et al. 2013). Despite the age-related decline in these various parameters, repeated muscular activation does not always lead to greater fatigability in older adults when compared with young adults, and seems to depend on the task. During sustained isometric tasks, older adults are usually less fatigable than young adults (Christie et al. 2011), until more advanced ages (Justice et al. 2014). However, there are far fewer studies focusing on age-related fatigability for dynamic as compared with isometric contractions. During moderate to fast isotonic concentric tasks, older adults are more fatigable than young (Christie et al. 2011; Power et al. 2013), but results are variable when reported for isokinetic tasks (Baudry et al. 2007; Rawson 2010; Callahan and Kent-Braun 2011; Dalton et al. 2012).
Comparisons across studies are hampered by the criterion measure used to assess fatigue (e.g., maximal voluntary isometric contraction (MVC), torque, and time to task failure) and differences in the dynamic protocol employed (isokinetic vs isotonic), which can influence the age-related fatigue differences (Dalton et al. 2010b; Dalton et al. 2012; Yoon et al. 2013). Both torque and velocity exhibit different fatigue-related mechanisms (Allen et al. 2008) and may be differentially impacted by age. The age-related loss of torque-generating capacity combined with slower contractile velocities results in a greater relative loss of power production than the percentage loss in either factor separately (Power et al. 2013). Maximal power production is achieved by an optimal combination of both submaximal torque and angular velocity and has emerged as a key component in quantifying physical function in older adults (Pojednic et al. 2012; Bean et al. 2013). However, decrements in contractile velocity rather than torque may be the more important contributing factor accounting for reductions in maximal power in older individuals (Dalton et al. 2010b; Dalton et al. 2012; Pojednic et al. 2012; Thompson et al. 2014). Thus, as a result of their slower contracting muscles, older adults may rely more on the torque component to generate maximal power than their younger counterparts (Dalton et al. 2014). Whether and how this age-related shift toward a greater reliance on torque alters the fatigue and recovery responses have not been explored.
Studies that used constrained velocity (i.e., isokinetic) tasks and thus defined fatigue as a loss of torque have reported the same (Laforest et al. 1990; Callahan et al. 2009; Dalton et al. 2012), less (Lanza et al. 2004; Rawson 2010), or more (Baudry et al. 2007; Callahan and Kent-Braun 2011) fatigue in older adults compared with young. Explaining the variability among these studies that used isokinetic tasks, and in comparison with those that have used isotonic tasks, is challenging due to the range of angular velocities tested. However, angular velocity and work performed during shortening contractions are likely key factors which may ultimately explain some of these discrepant findings. Furthermore, maximal velocity and rate of angular velocity production in older adults are diminished when compared with young (Thompson et al. 2014) and can potentially limit their ability to attain the pre-set (absolute) angular velocity imposed by an isokinetic dynamometer (Lanza et al. 2003; Callahan and Kent-Braun 2011). Failure to keep up with the isokinetic velocity hampers the development of maximal torque, thereby further confounding the assessment of fatigue and resulting in an overall limitation for meaningful comparisons among the limited studies using dynamic contractions with different conditions. Therefore, in order to better understand fatigue following high-intensity shortening contractions with adult aging, assessing performance in relation to each participant’s maximal power profile might be a better approach.
Fatigue resistance in older adults during isometric tasks appears to be due partly to a lower absolute energy cost of contraction (Tevald et al. 2010) owing to a greater reliance on oxidative phosphorylation compared with young adults (Lanza et al. 2005; Lanza et al. 2007). However, metabolic economy is the same during brief isometric MVCs (Christie et al. 2014), and the ATP cost per contraction is even greater during submaximal isotonic plantar flexions (Layec et al. 2014) compared with young. During shortening contractions in which joint angular velocity is unconstrained (i.e., isotonic), muscle power is substantially impaired in older individuals. This age-related impairment in power combined with a greater energetic cost during isotonic shortening contractions may explain the reduced fatigue resistance in older adults (Petrella et al. 2005; McNeil and Rice 2007; Dalton et al. 2010b; Dalton et al. 2012). Furthermore, assessing recovery of various parameters and multiple points of the torque–angular velocity–power relationship is helpful in understanding the factors of fatigue (Jones et al. 2006; Power et al. 2013), but this has not been explored following repeated shortening contractions in older adults.
Therefore, the current study was designed to assess the effects of the constraints imposed by a fixed angular velocity (isokinetic), or a fixed resistance or torque (isotonic) when comparing groups with significantly different neuromuscular capacities (old vs young). The aim was to assess and compare age-related fatigue and recovery between a repeated constrained (isokinetic) and an unconstrained velocity (isotonic) task, in which participants performed fatiguing contractions at the velocity (isokinetic) or resistance (isotonic) corresponding to their individual maximal power. The number of contractions to task failure was considered as the measure of fatigability between tasks and age groups. Furthermore, isotonic contractions over a range of resistances were used to assess velocity (time)-dependent mechanisms of the fatigue and recovery responses to the specific dynamic task. When matched for maximal power, we hypothesized that the older men would be more fatigable than the young for the isotonic task but that there would be no age-related difference for the isokinetic task. The contributing factors leading to task failure, however, would differ between young and old. The older men would be affected more by velocity-dependent factors, and the young would be impaired more by torque-dependent factors resulting in divergent shifts in the torque–power relationships. We also hypothesized that the older men would show a delayed recovery following task failure following both tasks, specifically a slower time course of recovery at lower loads (faster velocities) due to their slower contractile velocity.
Methods
Participants
Nine older men (74.9 ± 3.2 years; 172.2 ± 3.7 cm; 84.6 ± 10.5 kg) and 11 younger men (24.6 ± 4.0 years; 175.9 ± 7.8 cm; 77.7 ± 8.6 kg) were recruited from a local senior’s activity group, and the university student community, respectively. All participants were free from neuromuscular or musculoskeletal disorders and recreationally active, but not engaging in any systematic fitness training. Participants were instructed to refrain from strenuous exercise 1 day prior to each experimental session and to not consume caffeine 4 h before testing. Written informed consent was obtained from all participants prior to testing. The experimental protocol was approved by the local university’s ethics review board for experimentation on humans, and the study conformed to the Declaration of Helsinki.
Study design
Participants reported to the Neuromuscular Laboratory for three experimental testing days. Testing session 1 was structured to determine maximum power for the two distinct shortening contraction tasks (isokinetic and isotonic). For sessions 2 and 3, participants performed an isokinetic, or an isotonic fatiguing task based on the velocity or load, respectively, at which maximal power was achieved during testing session 1. The order of the dynamic fatigue task (session 2 or 3) was assigned pseudo-randomly. Testing days were separated by 2–5 days to avoid any residual effects of fatigue.
Experimental arrangement
A HUMAC NORM multi-joint dynamometer (CSMi Medical Solutions, Stoughton, MA) was used to measure torque and angular velocity produced by the knee extensors, and position of the knee joint in the isometric, isokinetic, and isotonic modes. Participants were seated comfortably in an upright position with the hip positioned at 100° and the knee of the dominant leg aligned with the dynamometer’s axis of rotation (Fig. 1a). An inelastic strap, 2 cm proximal to the tibial malleolus, secured the leg to the dynamometer knee attachment. The knee angle was placed to 80° (full knee extension = 180°) for all isometric measures and initial position for the dynamic contractions. The range of motion for all dynamic contractions was 70° (range 80° to 150°). Extraneous movements during knee extension efforts were prevented by inelastic straps fastened securely around the shoulders, hips, and thigh. All knee extensor torques, velocities, and angular positions were converted using a 12-bit analog-to-digital data acquisition board (Power 1401; Cambridge Electronic Design, Cambridge, UK) and sampled at 500 Hz using Spike2 software (version 7, Cambridge Electronic Design, Cambridge, UK).
Fig. 1.
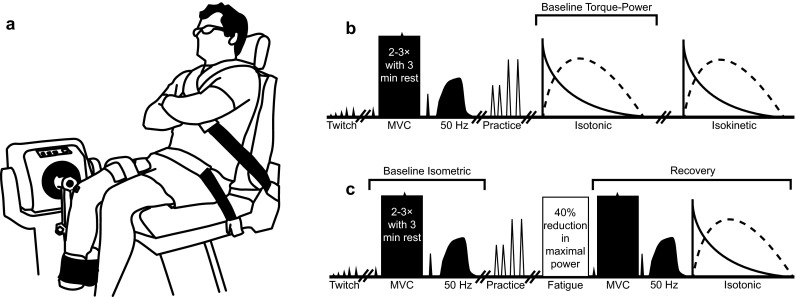
The experimental setup (a) and protocol timeline for testing session 1 (b) and testing sessions 2 and 3 (c). Procedures for testing sessions 2 and 3 were identical, except for the fatigue task in which the participants either performed maximal effort isokinetic or isotonic shortening contractions against a pre-set isovelocity or relative resistance (% MVC), respectively, corresponding to each participant’s maximal power attained during testing session 1. Shaded shapes represent isometric neuromuscular procedures and open shapes represent dynamic neuromuscular procedures. Double forward slashes represent rest periods of 5 min. The fatigue task consisted of maximal effort shortening contractions and was terminated when maximal power production was reduced by 40 % for three consecutive efforts. Twitch, peak twitch torque; MVC, maximal voluntary isometric contraction with electrically evoked contractile properties; 50 Hz, 50-Hz torque; Torque-Power, torque–angular velocity–power relationship contractions for the isotonic and isokinetic modes
To assess voluntary activation and neuromuscular properties of the knee extensors, electrically evoked contractions were delivered via two custom-made aluminum electrode pads (6–8 cm wide and 20–30 cm long) wrapped in a damp paper towel covered in conductive gel. The stimulating electrodes were placed transversally on the thigh, with the cathode ~6 cm proximal to the patella and the anode ~7 cm distal to the inguinal fold. Visual inspection and palpation were used to ensure only the knee extensors were activated during electrical stimulation. The square-wave electrical stimuli were delivered via a computer-controlled constant current stimulator (DS7AH, Digitimer Ltd, Welwyn Garden City, UK) with a 200-μs duration at 400 V.
Surface electromyography (EMG) of the knee extensors was assessed with self-adhering Ag-AgCl surface electrodes (H59P Repositionable Monitoring Electrodes; Kendall, Mansfield, MA) with the active electrode placed over the muscle belly of the rectus femoris, equidistant from the patella and greater trochanter. The reference and ground were placed over the patella. The skin was cleaned vigorously with an alcohol swab prior to electrode placement. The EMG signals were pre-amplified (×100), amplified (×2), bandwidth filtered (10–1000 Hz), converted by a 12-bit analog-to-digital system (Power 1401, Cambridge Electronic Design, Cambridge, UK), and sampled at 2500 Hz.
Experimental procedure
Testing session 1: baseline isometric measurements
Once the participant was oriented in the dynamometer, peak twitch torque (Pt) was determined. The stimulator current was increased progressively (15–20 mA per stimulation) until a plateau was achieved in Pt. The current was subsequently increased an additional 15 % to ensure consistent activation of the underlying muscle mass without a decrement in Pt. Next, two to three MVCs were performed for 3–5 s with 3 min of rest separating each effort. Participants were provided visual feedback of the torque on a computer monitor and encouraged strongly during all MVCs. Electrically evoked twitches (single stimuli) were delivered 2 s prior, during peak torque plateau (Ts), and 2 s following (Tr) each MVC. Three minutes later, the 50-Hz stimulation amplitude was determined by increasing the stimulator output (2–5 mA with a stimulation duration of 500 ms) progressively until peak evoked 50-Hz torque equated to ~40 % of MVC amplitude (Cheng and Rice 2005). The knee flexors were monitored visually and through palpation to ensure no spread of the electrical current to the antagonists. Five minutes of rest was given following the 50-Hz peak torque determination.
Testing session 1: baseline dynamic measurements
To improve the reliability of the isokinetic and isotonic (Power et al. 2011) contractions, participants were familiarized with four to six practice dynamic knee extensions, executed at slow (90° s−1) and fast (360° s−1) velocities for the isokinetic, and heavy (~35 % MVC) and moderate (~20 % MVC) resistances for the isotonic modes for a total of four practice conditions. The order and type of practice condition were pseudo-randomized among participants. Then, to determine maximal power, participants performed two sets of shortening contractions at eight velocities, or resistances for the isokinetic and isotonic modes, respectively, separated by 5 min of rest (see Fig. 1b). The order of these contraction modes was assigned pseudo-randomly among participants. The isokinetic contractions entailed two maximal effort knee extensions at velocities of 60°, 90°, 180°, 240°, 300°, 360°, 400°, and 500° s−1, whereas the isotonic contractions consisted of two maximal effort knee extensions at relative resistances of 10, 15, 20, 25, 30, 35, 40, and 45 % MVC (calculated from the baseline isometric MVC). At the end of the range of motion for each dynamic contraction (150°), the participant relaxed fully and the leg was returned passively to the start position (80°). Approximately 3 s separated each pair of maximal effort knee extensions. Isokinetic velocities and isotonic resistances were performed pseudo-randomly within sets, with the lowest load (isotonic) or fastest isokinetic angular velocity performed first to limit the influence of fatigue. Participants were instructed to contract as hard and as fast as possible, exhorted, and provided visual feedback of the power output during all maximal dynamic efforts.
Testing sessions 2 and 3: fatigue tasks
Testing began with the same baseline isometric measurements and were performed in the same order as testing session 1. For the isotonic protocol, participants performed repeated maximal knee extensions against the resistance (% MVC) that produced maximum power as determined from testing session 1, whereas for the isokinetic protocol, participants performed repeated maximal knee extensions against an absolute velocity corresponding with maximum power determined from testing session 1. Five minutes following baseline testing, participants were familiarized with the fatigue task by performing ~5–10 dynamic contractions separated by brief rest intervals (~5–10 s). Once a constant peak power was attained (no change during five successive attempts), the participants rested for 5 min and then performed the respective fatigue protocol. Task failure was defined as a reduction of 40 % in maximum power for three consecutive contractions (Fig. 2).
Fig. 2.

Representative unprocessed data taken from one old and one young participant at the beginning and end (task failure) of the isokinetic (a) and isotonic (b) fatigue tasks. The black circles represent the torque and velocity values attained at peak power. The grey circles highlight the impact artifact upon completion of the 70° range of motion for the dynamic shortening knee extensions
Testing session 2 and 3: recovery
Immediately (~2 s following task termination), 2.5, 5, and 10 min following task failure for both dynamic fatigue tasks, isometric and dynamic neuromuscular properties were tested within a 90-s epoch and in the following order: twitch, MVC with superimposed twitch, post-MVC twitch, 50-Hz tetanus, and two maximal effort dynamic knee extensions at relative isotonic resistances of 10, 15, 20, 25, 30, 35, 40, and 45 % MVC (identical chronological order as experiment 1; Fig. 1c). Because of the limitations older adults may experience with higher isokinetic velocities (Lanza et al. 2003), to gain a better appreciation of the recovery of the velocity and torque components of power and to minimize the number of contractions during the brief time periods, isotonic testing across multiple resistances was used to assess fatigue and recovery.
Data analyses
The electrically evoked contractile properties were assessed by calculating Pt, time to peak twitch torque (TPT), normalized TPT to Pt, 50-Hz peak torque, 50-Hz half-relaxation time (HRT), and normalized 50-Hz HRT to 50-Hz peak torque. Isometric MVC was assessed by peak torque amplitude and voluntary activation using the interpolated twitch technique (% voluntary activation = [(1 − Ts / Tr) × 100]). The baseline isometric neuromuscular property values corresponding to the contraction with the highest torque amplitude were used for statistical analysis. Surface EMG activity during the MVCs following the fatigue tasks was analyzed by determining the root-mean-square (RMS) amplitude with a 500-ms epoch about the peak torque, and this value was normalized to the RMS amplitude of the baseline MVC for the respective testing session.
Baseline instantaneous peak power, at each pre-set velocity or resistance of the isokinetic and isotonic contractions, respectively, was measured from the contraction with the highest instantaneous value. Maximal power was determined as the highest peak power value across the range of velocities and loads for the isokinetic and isotonic contractions, respectively. Power (W) was calculated as the product of the instantaneous peak torque (N·m) and angular velocity (rad s−1), and maximal velocity values were taken from the 15 % MVC isotonic contraction. For the dynamic fatigue tasks, the number of knee extension contractions to task failure was taken as the criterion measure for fatigability, whereas a decline in range of motion (joint excursion) of the knee joint from the beginning to the end of the fatigue tasks was used as a complimentary measure. Range of motion (°) or knee joint excursion was averaged over the first five and last five knee extension contractions. Additionally, work was estimated for each contraction as the product of mean torque (N·m) and total range of knee joint excursion (rad; i.e., range of motion). Values for each contraction were summed to determine total work performed during each fatigue task.
Statistical analyses
Data were analyzed using SPSS version 20 (IBM, Chicago, IL). To ensure that baseline values of the isometric measures were not different from day-to-day, a one-way analysis of variance (ANOVA) was applied. Because there were no day-to-day differences, baseline values were pooled to test for age-related (group) differences. An unpaired t test was used to compare all isometric baseline values between the two age groups, except for voluntary activation for which a non-parametric Mann–Whitney U test was used.
For the baseline dynamic contractions, maximal velocity of the isotonic contractions at 15 % MVC, and the resistance (isotonic) or angular velocity (isokinetic) at which maximal power was attained were analyzed with an unpaired t test. Further, 95 % confidence intervals for the differences in means were calculated for baseline values, as appropriate. Maximal power (task × age), peak power for the isokinetic contractions (velocity × age), peak power for the isotonic contractions (resistance × age), and the angular velocity at which maximal power was attained for the dynamic tasks (velocity × age) were analyzed with a two-factor analysis of variance (ANOVA) with repeated measures.
For the fatigue tasks, total work performed, number of contractions to task failure, and absolute change in range of motion of the knee joint were analyzed using a two-way ANOVA (task × age) with repeated measures. Because we anticipated age-related differences in the isotonic, but not the isokinetic task, we used an a priori design to test the number of contractions to task failure per task with an unpaired t test. For fatigue and recovery, all isometric measures, and dynamic measures of maximal power, maximal velocity, and the resistance at which maximal power was achieved were analyzed using a three-factor ANOVA (task × time × age) with repeated measures. For relative change in peak power, a four-factor ANOVA (task × time × resistance × age) with repeated measures was used. The level of significance was set at p ≤ 0.05. When main effects or interactions were significant, post hoc analysis with a modified Bonferroni correction factor was performed. Descriptive data reported in the text and tables are reported as means ± standard deviations, and all data reported in figures are means ± standard errors of the mean.
Results
Isometric baseline
Baseline neuromuscular knee extensor values are reported in Table 1. Compared with young, the old men were 45, 38, and 45 % weaker for MVC (t (18) = 7.20, p < 0.01), Pt (t (18) = 5.42, p < 0.01), and 50-Hz (t (18) = 6.32, p < 0.01) knee extensor torque, but both age groups were capable of equally high voluntary activation (≥95 %; z = −0.96, p = 0.34). The 50-Hz HRT was 9 % slower (t (18) = −2.26, p < 0.05) for the old than young, but twitch TPT was not different (t (18) = −1.71, p = 0.10). When normalized to their respective peak torques, 50-Hz HRT and twitch TPT were 98 % (t (18) = −7.64, p < 0.01) and 60 % (t (18) = −6.16, p < 0.01) slower, respectively, for the old compared with young.
Table 1.
Baseline neuromuscular properties of the knee extensors
| Young | Old | % CI | |
|---|---|---|---|
| MVC (N·m) | 269.2 ± 45.3* | 147.9 ± 24.9 | 85.9, 156.7 |
| Voluntary activation (%) | 96.4 ± 2.1 | 94.9 ± 3.6 | −1.0, 4.3 |
| P t (N·m) | 48.4 ± 9.4* | 30.3 ± 3.8 | 11.2, 25.3 |
| 50 Hz (N·m) | 104.9 ± 20.7* | 59.1 ± 10.3 | 31.6, 63.2 |
| TPT (ms) | 95.9 ± 3.9 | 97.9 ± 2.8 | −5.6, 0.6 |
| HRT (ms) | 124.6 ± 13.8* | 136.9 ± 7.6 | −22.8, −0.8 |
| Normalized TPT (ms N·m−1) | 2.1 ± 0.5* | 3.3 ± 0.4 | −1.7, −0.8 |
| Normalized HRT (ms N·m−1) | 1.2 ± 0.3* | 2.4 ± 0.4 | −1.5, −0.9 |
| Velocity (° s−1) | 417.7 ± 29.4* | 343.1 ± 29.5 | 47.0, 102.4 |
| Isotonic maximal power (W) | 623.1 ± 122.7* | 275.8 ± 29.8 | 258.9, 435.7 |
| Isokinetic maximal power (W) | 677.1 ± 117.1* | 294.1 ± 42.3 | 296.3, 469.5 |
| Maximal power load (% MVC) | 27.7 ± 6.1* | 33.3 ± 5.0 | −10.9, −0.3 |
| Maximal power velocity (° s−1) | 376.4 ± 42.7* | 313.3 ± 26.5 | 28.6, 97.4 |
Values are means ± standard deviations with 95 % confidence intervals (CIs) for the difference in means
MVC maximal voluntary isometric contraction, P t peak twitch torque, 50 Hz 50-Hz torque, TPT time to peak twitch torque, HRT 50-Hz half-relaxation time, Velocity maximal velocity at 15 % MVC
*Significant difference between young and old men (p < 0.05)
Dynamic baseline
Maximal power was not different between tasks (task, F1,18 = 2.55, p = 0.13; interaction, F1,18 = 0.62, p = 0.44), but the older men were ~56 % less powerful than the young for both dynamic contractions (age, F1,18 = 109.16, p < 0.01). The older men achieved an 18 % lower maximal velocity at 15 % MVC than the young (t (18) = 5.66, p < 0.01). Hence, the old achieved maximal power at a 17 % lower absolute velocity (t (18) = 3.85, p < 0.01) and 20 % heavier relative load (t (18) = −2.22, p < 0.05) than the young for the isokinetic and isotonic tasks, respectively (Table 1). Furthermore, the older (isokinetic, 313.3 ± 26.5° s−1; isotonic, 232.3 ± 32.4° s−1) and the younger (isokinetic, 376.4 ± 42.7° s−1; isotonic, 330.4 ± 45.0° s−1) men achieved maximal power with 26 and 12 % faster angular velocities for the isokinetic than the isotonic task, respectively (age, F1,18 = 36.68, p < 0.01; task, F1,18 = 34.28, p < 0.01; interaction, F1,18 = 2.63, p = 0.12).
Isokinetic
All older men were unable to keep up with the dynamometer to produce reliable torque at 500° s−1, and two subjects were unable to achieve isokinetic knee extensions at 400° s−1. Thus, values taken from the 500° s−1 were not utilized for further statistical analysis. The older men were 45–62 % less powerful than the young at all absolute velocities (Fig. 3a, b) with the difference between age groups increasing as velocity increased (age, F1,18 = 83.84, p < 0.01; velocity, F6,108 = 271.35, p < 0.01; and interaction, F6,108 = 63.79, p < 0.01). Peak power increased successively up to 360° s−1 for the young men, whereas peak power plateaued at 180° s−1 and remained flat thereafter for the older men (Fig. 3a).
Fig. 3.
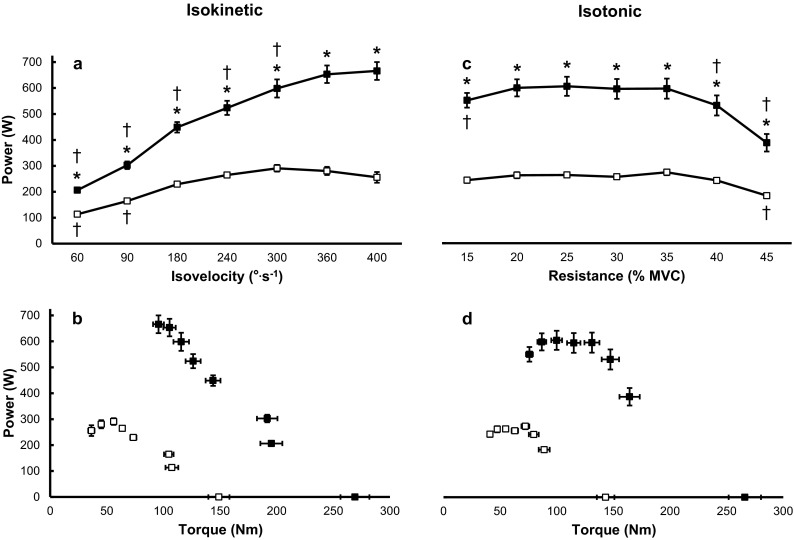
Baseline knee extension peak power over varying absolute isokinetic velocities (a) and relative isotonic resistances (c) for the old (open squares) and young (filled squares) men. The old men were less powerful than the young (*p < 0.05) for all absolute velocities and relative resistances. Isokinetic peak power achieved at 60° and 90° s−1 and isotonic peak power at 45 % maximal voluntary isometric contraction (MVC) were lower than all other velocities and relative resistances, respectively, for the old men († p < 0.05). For the young men, isokinetic peak power progressively increased up to 360° s−1 and all values ≤300° s−1 were lower than values attained at 360°s−1 and 400° s−1 († p < 0.05), whereas peak power was lower at 15, 40, and 45 % MVC than all other isotonic resistances († p < 0.05). Panels (b) and (d) highlight a shift down and to the left in the torque–power relationship for the old compared with young men. Data equivalent to 0 W in panels (b) and (d) indicate MVC values. All values are means ± standard error of the mean
Isotonic
All participants were capable of performing the knee extension requirements throughout the range of relative resistances. However, the young men exceeded the velocity limits of the dynamometer (500° s−1) at 10 % MVC; thus, values taken from this relative resistance were not used for statistical analysis. Across all relative resistances, the older men were 53–57 % less powerful than the young (age, F1,18 = 63.12, p < 0.01; resistance, F6,108 = 64.88, p < 0.01; interaction, F6,108 = 13.37, p < 0.01) (Fig. 3c, d). The torque-power relationship was relatively flat across resistances for the old men compared with the young, such that the only detectable differences in peak power for the old were between 45 % MVC and all other relative resistances. For the young men, peak power was greatest for relative resistances of 20–35 % MVC, whereby these values were larger than 15, 40, and 45 % MVC (Fig. 3c).
Fatigue task
The isokinetic task was less fatigable than the isotonic task such that the old and young men performed 38 and 33 % more isokinetic than isotonic contractions (task, F1,18 = 70.61, p < 0.01), respectively (Table 2). With respect to age, the old men completed 20 % fewer contractions for the isotonic task than their younger counterparts (t (18) = 2.24, p < 0.05), but no age-related difference was detected for the isokinetic task (t (18) = 1.33, p = 0.18). Throughout the fatigue tasks, the older men performed 53 and 56 % less total work than the young for the isokinetic and isotonic contractions, respectively (age, F1,18 = 42.34, p < 0.01). There was no detectable difference in total amount of work between isokinetic and isotonic tasks (task, F1,18 = 0.56, p = 0.47; interaction, F1,18 = 0.27, p = 0.61). By the end of the isokinetic fatigue task, range of motion of the knee joint did not decrease for either age group, but range of motion was reduced by ~21° at task failure in the isotonic fatigue task for both the old and young men (Table 2; task, F1,18 = 124.53, p < 0.01; age, F1,18 = 0.39, p = 0.54; interaction, F1,18 = 0.57, p = 0.46).
Table 2.
Neuromuscular properties during the fatigue task
| Resistance | Isokinetic | Isotonic | ||||
|---|---|---|---|---|---|---|
| Young | Old | 95 % CI | Young | Old | 95 % CI | |
| Contractions (#) | 47.3 ± 8.8† | 41.0 ± 10.6† | −3.4, 16.0 | 31.8 ± 5.0* | 25.4 ± 7.7 | 0.4, 12.4 |
| Work (J) | 3648.9 ± 922.0* | 1714.3 ± 526.6 | 1156.1, 2713.1 | 3611.4 ± 790.2* | 1589.1 ± 548.1 | 1367.8, 2676.7 |
| ROM (°Δ) | 0.4 ± 1.0† | 0.6 ± 1.7† | −1.6, 1.0 | 22.8 ± 5.8 | 18.6 ± 6.4 | −4.7, 9.3 |
Both age groups performed more contractions and maintained a larger range of motion (ROM) during the isokinetic fatigue task compared with the isotonic (†p < 0.05). The older men performed fewer contractions than the young for the isotonic task and less absolute total work for both fatigue tasks (*p < 0.05). Values are means ± standard deviations with 95 % confidence intervals (CIs) for the difference in means
Fatigue and recovery: isometric measures
Immediately following task failure of both tasks, MVC torque was reduced by ~30 % (time, F4,72 = 86.48, p < 0.01) and did not recover fully by 10 min for either age group (Fig. 4a; age, F1,18 = 3.84, p = 0.07; task, F1,18 = 1.34, p = 0.26). For both fatiguing tasks, immediately following task failure, voluntary activation was reduced to 93.0 ± 5.6 % with no group difference and did not recover fully (age, F1,18 = 0.95, p = 0.34; time, F4,72 = 8.58, p < 0.01; task, F1,18 = 1.55, p = 0.23), which likely contributed partly to the reduced MVC amplitude. In support, surface EMG amplitude during the MVC was reduced to 75.6 ± 22.1 % immediately following task failure for both tasks and age groups and by 10 min only recovered to 84.5 ± 11.3 % (time, F4,72 = 14.96, p < 0.01; task, F1,18 = 1.75, p = 0.20; age, F1,18 = 0.02, p = 0.91).
Fig. 4.

Maximal voluntary isometric contraction (MVC; a), maximal velocity at 15 % MVC (b), and maximal power (c) were reduced immediately following task failure of the isokinetic and isotonic contractions for the old (open squares) and young (filled squares) men († p < 0.05). The MVC did not recover fully by 10 min, whereas maximal velocity recovered within this time frame and maximal power recovered fully by 2.5 min for both fatigue tasks and age groups. Dashed lines indicate when the fatigue task was performed. Immediately following task failure, immediate; and recovery at 2.5, 5, and 10 min, R2.5, R5, and R10, respectively. Values are means ± standard error of the mean
The Pt was reduced to 64.1 ± 20.3 % in the young men following task failure for both tasks and recovered within 2.5 min. The old men, however, exhibited no changes in Pt immediately following task failure for both tasks (age, F1,18 = 10.17, p < 0.01; time, F4,72 = 18.56, p < 0.01; time × age, F4,72 = 10.17, p < 0.01; task × time, F4,72 = 4.67, p < 0.05). For the 50-Hz contraction, torque was reduced to 76.7 ± 8.7 and 63.6 ± 8.9 % of baseline for the old and young men, respectively, immediately following task failure of both tasks, but the depression was less for the old men, and neither group recovered fully within 10 min (time, F4,72 = 14.96, p < 0.01; time × age, F4,72 = 10.17, p < 0.01; task × time, F4,72 = 4.67, p < 0.05). Half-relaxation time of the 50 Hz increased to 154.4 ± 26.9 % immediately following task failure and recovered fully by 5 min (time, F4,72 = 73.75, p < 0.01), but twitch TPT was unaltered following task failure for both fatigue tasks and age groups.
Fatigue and recovery: dynamic measures
Maximal velocity was reduced by 13 % immediately following task failure for both groups and tasks and recovered fully by 10 min (Fig. 4b; time, F4,72 = 32.67, p < 0.01; task, F1,18 = 0.42, p = 0.53; age, F1,18 = 0.07, p = 0.79), whereas maximal power was decreased by 25 % immediately following task failure and recovered within 2.5 min (Fig. 4c; time, F4,72 = 27.63, p < 0.01; task, F1,18 = 1.36, p = 0.26; age, F1,18 = 0.01, p = 0.96). Maximal power production was moderately and negatively correlated with HRT immediately following task failure and throughout recovery (r = −0.48; p < 0.01). The relative isotonic resistance at which maximum power was achieved shifted similarly toward lighter resistances (old, 20.8 ± 7.5 % MVC; young, 21.9 ± 5.1 % MVC) for both age groups immediately following task failure for either fatigue task, and recovered to baseline values by 2.5 min of recovery (old, 31.7 ± 6.9 % MVC; young, 29.3 ± 7.1 % MVC; time, F4,72 = 9.77, p < 0.01; task, F1,18 = 2.60, p = 0.12; age, F1,18 = 2.53, p = 0.13).
Peak power was not altered differently when comparing fatigue task or age group; thus, these data were collapsed across age and task for further analysis. Peak power was reduced immediately following task failure and, except for 25 % MVC, recovered fully by 10 min for all other relative isotonic resistances (Fig. 5a; time, F4,72 = 37.38, p < 0.01). Immediately and 2.5 min following task failure, the 15 % MVC resistance exhibited the least relative fatigue, and peak power impairment was exacerbated with an increase in isotonic resistance (Fig. 5a; resistance, F6,108 = 21.27, p < 0.01; time × resistance, F24,432 = 17.76, p < 0.01). Thus, both age groups exhibited a shift downward and leftward in the voluntary torque–power relationship immediately following task failure (Fig. 5b).
Fig. 5.

Peak power across a range of relative isotonic resistances immediately (Post), 2.5 (R2.5), 5 (R5), and 10 min (R10) following task failure for the old (open squares) and young (filled squares) men (Column a). Because there was no main effect for task, data were collapsed across the isokinetic and isotonic fatigue tasks for presentation. † denotes a significant difference from baseline (p < 0.05). ‡ indicates a significant difference than all other resistances during the same recovery time point. The dashed line represents baseline values. Column b highlights the alteration of the absolute torque–power relationship for the old (open circles) and young (filled circles) men at each recovery time point compared with baseline (old, open squares; young, filled squares). Data equivalent to 0 W represent values taken from the maximum voluntary isometric contraction. Values are means ± standard error of the mean
Discussion
The present study addresses important issues in furthering our understanding of how age-related alterations in the neuromuscular system affect fatigability during dynamic tasks. Uniquely, both isokinetic and isotonic fatigue tasks were matched to correspond with each participant’s maximal power, and following task failure, isotonic torque–power relationships were used to compare factors of fatigability and short-term recovery (up to 10 min) in older and younger men. In agreement with our main hypothesis, the older adults performed 20 % fewer isotonic knee extensions to task failure than the young, and there were no detectable age-related differences for the isokinetic contractions. Thus, the old men were more fatigable when executing the isotonic task but exhibited similar fatigability as the young during the isokinetic task. However, contrary to our secondary hypothesis, both age groups exhibited a similar degree of a downward and leftward shift in the torque-power relationship and had similar alterations in isometric contractile properties immediately following failure of both tasks. In addition, the angular velocities needed to achieve maximal power were slower for the isotonic contractions than the isokinetic task in both age groups. These findings indicate that factors other than age-related impairments in velocity production influence the greater fatigability of older men during isotonic contractions.
Baseline
The older men produced 38–62 % less torque than the young for the voluntary and electrically evoked isometric and voluntary isokinetic knee extensions, despite similar high voluntary activation levels (≥95 %) during the MVCs. The lower knee extensor strength in the old than young is comparable to previous findings (Roos et al. 1999; Lanza et al. 2003; Petrella et al. 2005; Callahan et al. 2009; Dalton et al. 2012). Additionally, normalized contractile properties were 60–98 % slower in the old than young, which was accompanied by 16–20 % lower peak angular velocities at isotonic resistances <40 % MVC. The age-related slowing in contractile function is in agreement with previous reports in this (Roos et al. 1999; Petrella et al. 2005; Dalton et al. 2012; Thompson et al. 2014) and other muscle groups (Valour et al. 2003; McNeil and Rice 2007; Dalton et al. 2010a, b, 2014; Power et al. 2014). Consequently, the slower and weaker knee extensors resulted in a ~56 % lower maximal power capacity in the older men than young, which was accompanied by a shift in maximum power achievement to slower absolute velocities and higher relative torques for the isokinetic and isotonic tasks, respectively. Similar to a previous report (Lanza et al. 2003) testing faster isokinetic velocities, all older men in the present study could not keep up with the dynamometer velocity to produce reliable torque at 500° s−1, and two subjects were unable to perform the isokinetic knee extensions at 400° s−1. Unlike the isokinetic task, each participant was capable of performing the knee extension requirements for the relative resistances in the isotonic mode, suggesting that isotonic tasks are likely a better overall measure to assess power when comparing among groups with different neuromuscular capacities (i.e., older vs young adults). Additionally, isotonic actions may reflect more closely normal function (fixed resistances with varying velocities) than when joint angular velocity is constrained and resistance can vary (isokinetic).
Task failure
When matched for relative maximal power, the older and younger men performed a similar number of isokinetic contractions (~45) to achieve a 40 % reduction in power (task failure). These findings of no age-related difference in fatigability of the knee extensors resemble most (Laforest et al. 1990; Lindstrom et al. 1997; Deschenes et al. 2008; Callahan et al. 2009; Callahan and Kent-Braun 2011; Dalton et al. 2012), but not all previous studies in this muscle group (Rawson 2010; Callahan and Kent-Braun 2011). Greater age-related differences emerged only at the faster velocities (Callahan and Kent-Braun 2011), but these velocities of typically between 90° and 270° s−1 are not fast for angular knee extensions (Dalton et al. 2012; Thompson et al. 2014). In the present study, we tested velocities greater than 300° s−1. Furthermore and importantly, unlike previous reports in which subjects were likely operating on different portions of their respective torque–power relationships, both the old and young participants were functioning at their maximal power or optimal torque–angular velocity relationship for the isokinetic task. Thus, the dynamic tasks performed here better reflect the optimal and maximal capacity of each individual tested, and the results support the studies that indicate no age-related differences in fatigability during isokinetic tasks (Laforest et al. 1990; Lindstrom et al. 1997; Deschenes et al. 2008; Callahan et al. 2009; Callahan and Kent-Braun 2011; Dalton et al. 2012). Whether this is due to a limitation with this testing modality or to physiological factors (see below) is not entirely clear.
For the isotonic task, however, when matched for maximal power, we found that older adults were more fatigable than their younger counterparts. The older men required ~25 contractions to reach task failure (40 % reduction in maximal power), whereas the younger men performed ~32 contractions, a difference of ~20 %. This greater fatigability in the old is similar to other reports on the knee extensors using maximal effort isotonic shortening contractions (Petrella et al. 2005; Dalton et al. 2012) and solidifies those findings now when comparisons are matched for maximal power. Previously, it was suggested that the high velocity demand placed upon the older adults during shortening actions is a critical determinant of the greater fatigue-related reductions in power for older adults than young (Callahan and Kent-Braun 2011; Dalton et al. 2012). However, in the present investigation, when matched for power, the older adults performed the maximal isotonic efforts at ~68 % of their maximal loaded angular velocity capacity, whereas the young men performed the task at ~80 %. Thus, the older men performed the isotonic fatigue task at a slower relative velocity, yet exhibited greater fatigability (i.e., performed fewer contractions) than the young. Despite the importance of contractile velocity per se for dynamic contractions (Dalton et al. 2012; Pojednic et al. 2012; Bean et al. 2013; Thompson et al. 2014), it is clear that by accounting for the different age-related inherent capabilities of the neuromuscular system (equating for maximal power) that factors other than only velocity are involved in the greater fatigability of older men compared with young.
Typically for sustained isometric tasks, older adults are less fatigable than younger adults (Christie et al. 2011) because perhaps older individuals exhibit a greater reliance on oxidative phosphorylation and less on glycolytic pathways compared with the young (Lanza et al. 2007; Christie et al. 2014), leading to less acidosis and subsequently less H2PO4 (Kent-Braun 2009). However, the metabolic demand of shortening contractions is greater than that of sustained isometric contractions (Newham et al. 1995), and the enhanced energetic cost likely mitigates any age-related metabolic advantage. For example, during constant load sub-maximal isotonic plantar flexions at an effort equivalent to ~40 % maximal power output, Layec et al. (2014) found that the ATP cost per contraction was remarkably higher for older adults than young. Moreover, this increased energy usage was correlated with age-related impairments in plantar flexor power. Therefore, the age-related metabolic advantage reported for isometric tasks seems to be mitigated during brief and intermittent isometric (Christie et al. 2014; Layec et al. 2015) and possibly isokinetic contractions and even reversed during repetitive isotonic shortening contractions as demonstrated when the task is matched for maximal power (as shown here). In support of this idea, the total amount of work performed in the present study was not different when comparing the type of task. Nevertheless, the isokinetic task required a greater number contractions and a smaller reduction in range of motion at task failure for both age groups than the isotonic task (Table 2). This likely indicates that the isokinetic contractions were less energetically demanding than the isotonic contractions for both age groups. Thus, based on previous studies (Layec et al. 2014), it seems that the greater fatigability reported here for the isotonic contractions could reflect a larger metabolic cost per contraction during the isotonic task for the older men than young.
Fatigue and recovery
Immediately following task failure for both dynamic fatigue tasks in the present study, voluntary and electrically evoked isometric torques were reduced to similar levels for the old and young men. This finding is in agreement with other intermittent knee extensor fatigue tasks (Callahan et al. 2009; Dalton et al. 2012; Mcphee et al. 2014) and extends these observations to include dynamic shortening tasks matched for maximal power. This reduction in MVC torque can be partially explained by centrally mediated mechanisms, which does not appear to be age-dependent when testing healthy older men, at least for the knee extensors. For instance, during the MVCs tested immediately following task failure, voluntary activation was reduced to ~93 % and did not recover fully. In support, surface EMG amplitude during the MVC was reduced and only recovered to ~85 % baseline values by 10 min. Neuromuscular propagation is likely not a contributing factor as we reported elsewhere that the M-wave response was not different in young and old men following three types of maximal effort concentric knee extension tasks (Dalton et al 2012). The fatigue-induced reductions could partially arise from supraspinal and spinal factors and seem to recover similarly in older and young men within 10 min following task failure for the knee extensors.
Indeed, normalizing the dynamic fatigue task to an absolute velocity or percentage of an isometric MVC imposes a high speed upon older participants and was beneficial for highlighting the limitation older adults have with contractile velocity during repetitive tasks (Petrella et al. 2005; McNeil and Rice 2007; Dalton et al. 2010b; Callahan and Kent-Braun 2011; Dalton et al. 2012). However, this procedure may not be the best approach when comparing populations with differing contractile properties. For example, the older men in the current study exhibited slower contractile properties and maximal isotonic voluntary velocities with a leftward and downward shift in the torque–power relationship at baseline. Thus, if we matched for a measure other than maximal power, each age group would likely be operating on different portions of their torque–angular velocity relationships (Raj et al. 2010). To account for this limitation, here we matched for maximal power, and although the rate of fatigue was quicker in the old during the isotonic task than young, there was no difference in recovery of maximal power for either age group or task.
The fatigue-related reduction in power generating capacity is supported by the 50-Hz HRT slowing to the same extent immediately following task failure for both age groups and tasks. This measure was ~54 % slower immediately following task failure and recovered fully within 5 min. Previous studies have reported a negative moderate relationship between loss in peak power and slowing in HRT at task failure following fast unconstrained shortening contractions in young (Cheng and Rice 2005) and older men (Dalton et al. 2010b; Dalton et al. 2012). When the task was matched for maximal power, we found a moderate correlation (r = −0.48) between changes in maximal power and HRT following task failure and throughout recovery irrespective of contraction type (i.e., isokinetic or isotonic shortening) and age. The addition of these results with previous work (Cheng and Rice 2005; Dalton et al. 2010b; Dalton et al. 2012) further solidifies that the reductions in maximal power may be limited partially by factors related to prolongation of muscle relaxation time (Jones et al. 2006) in both age groups.
Surprisingly, maximal power generation was attained at a lower percent of MVC immediately following task failure equally for both the old and young men. Thus, the reduction in peak power was exacerbated at the higher isotonic resistances. Due to a greater transient reduction in maximal torque-generating capacity than maximal velocity (i.e., greater reductions in peak power at higher vs lighter resistances), the torque–power relationship was shifted downward and leftward immediately following task failure for both age groups (Fig. 5). The dynamic torque-generating capacity at each isotonic resistance following both fatigue tasks represented a greater relative percentage of initial MVC, as illustrated by the decrease in isometric MVC amplitude (Fig. 4) and thus required a greater emphasis on torque generation compared with baseline values in order for all participants to complete the task successfully. This finding is in contrast to our hypothesis as the heavier isotonic resistances exhibited the greater reductions in peak power for both age groups (Fig. 5). The leftward shift in the torque–power relationship (Fig. 5b) whereby maximal power was produced at lighter resistances for both age groups and tasks afforded a compensatory strategy to maintain minimal reductions in maximal power (only decreased by ~25 %) despite large impairments in peak power at the higher resistances (40–70 % for loads ≥30 % MVC). Thus, following task failure, both groups relied more on the velocity component for power generation with less reliance on torque. However, this compensation was reversed by 2.5 min of recovery as torque-generating capacity began to recover (Figs. 4 and 5), albeit not fully. Therefore, when fatigued to the same relative maximal power for both isokinetic and isotonic tasks, the recovery of neuromuscular function is similar between older and young men, despite the older adults exhibiting greater fatigability during the isotonic task.
In summary, the older and young men performed an isokinetic and isotonic fatigue task at a load or resistance, respectively, that corresponded with each individual’s maximal knee extensor power. Because both age groups were operating at their optimal portion of the torque–angular velocity relationship (i.e., maximal power) and were fatigued to the same reduction in power, we may have minimized the ability to assess any differences between groups throughout recovery. Nevertheless, the older men performed fewer isotonic knee extensions than the young, but a similar number of isokinetic contractions by task failure, despite the older men performing only ~45 % of the total work as that produced by the young. Hence, when compared with the young, the older men were more fatigable for the isotonic task but exhibited similar fatigue during the isokinetic task. Because the torque–power relationship shifted downward and to the left similarly in both tasks and age groups, it seems that the velocity component of power was not a contributing factor to the greater fatigue exhibited by the older men than young when both age groups were matched for maximal power. Although age-related velocity impairments are critical during certain dynamic tasks (Petrella et al. 2005; McNeil and Rice 2007; Dalton et al. 2010b; Callahan and Kent-Braun 2011; Dalton et al. 2012), other age-related factors, such as reduced metabolic economy or increased ATP cost per contraction (Layec et al. 2014; Layec et al. 2015), are likely limiting mechanisms of contractile performance during repetitive isotonic contractions when age groups are matched for maximal power.
Acknowledgments
The authors would like to thank all participants of the study, and to acknowledge Demetri Makrakos and Amelia Lanning for their help with data collection and analysis. The authors are grateful to Dr. Anita Christie for comments on a previous version of this manuscript. This work was supported by the Natural Sciences and Engineering Research Council (NSERC) of Canada. G.A. Power was supported by a Banting postdoctoral fellowship (Canadian Institutes for Health Research; CIHR) and Alberta Innovates Health Solutions (AIHS).
References
- Aagaard P, Suetta C, Caserotti P, Magnusson SP, Kjaer M. Role of the nervous system in sarcopenia and muscle atrophy with aging: strength training as a countermeasure. Scand J Med Sci Sports. 2010;20:49–64. doi: 10.1111/j.1600-0838.2009.01084.x. [DOI] [PubMed] [Google Scholar]
- Allen DG, Lamb GD, Westerblad H. Skeletal muscle fatigue: cellular mechanisms. Physiol Rev. 2008;88:287–332. doi: 10.1152/physrev.00015.2007. [DOI] [PubMed] [Google Scholar]
- Baudry S, Klass M, Pasquet B, Duchateau J. Age-related fatigability of the ankle dorsiflexor muscles during concentric and eccentric contractions. Eur J Appl Physiol. 2007;100:515–525. doi: 10.1007/s00421-006-0206-9. [DOI] [PubMed] [Google Scholar]
- Bean JF, Latham NK, Holt N, Kurlinksi L, Ni P, Leveille S, Percac-Lima S, Jette A. Which neuromuscular attributes are most associated with mobility among older primary care patients? Arch Phys Med Rehabil. 2013;94:2381–2388. doi: 10.1016/j.apmr.2013.07.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Callahan DM, Kent-Braun JA. Effect of old age on human skeletal muscle force-velocity and fatigue properties. J Appl Physiol. 2011;111:1345–1352. doi: 10.1152/japplphysiol.00367.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Callahan DM, Foulis SA, Kent-Braun JA. Age-related fatigue resistance in the knee extensor muscles is specific to contraction mode. Muscle Nerve. 2009;39:692–702. doi: 10.1002/mus.21278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng AJ, Rice CL. Fatigue and recovery of power and isometric torque following isotonic knee extensions. J Appl Physiol. 2005;99:1446–1452. doi: 10.1152/japplphysiol.00452.2005. [DOI] [PubMed] [Google Scholar]
- Christie AD, Snook EM, Kent-Braun JA. Systematic review and meta-analysis of skeletal muscle fatigue and old age. Med Sci Sport Exerc. 2011;43:568–577. doi: 10.1249/MSS.0b013e3181f9b1c4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christie AD, Tonson A, Larsen RG, DeBlois JP, Kent JA. Human skeletal muscle metabolic economy in vivo: effects of contraction intensity, age, and mobility impairment. Am J Physiol Regul Integr Comp Physiol. 2014;307:R1124–R1135. doi: 10.1152/ajpregu.00083.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dalton BH, Harwood B, Davidson AW, Rice CL. Recovery of motoneuron output is delayed in old men following high-intensity fatigue. J Neurophysiol. 2010;103:977–985. doi: 10.1152/jn.00908.2009. [DOI] [PubMed] [Google Scholar]
- Dalton BH, Power GA, Vandervoort AA, Rice CL. Power loss is greater in old men than young men during fast plantar flexion contractions. J Appl Physiol. 2010;109:1441–1447. doi: 10.1152/japplphysiol.00335.2010. [DOI] [PubMed] [Google Scholar]
- Dalton BH, Power GA, Vandervoort AA, Rice CL. The age-related slowing of voluntary shortening velocity exacerbates power loss during repeated fast knee extensions. Exp Gerontol. 2012;47:85–92. doi: 10.1016/j.exger.2011.10.010. [DOI] [PubMed] [Google Scholar]
- Dalton BH, Allen MD, Power GA, Vandervoort AA, Rice CL. The effect of knee joint angle on plantar flexor power in young and old men. Exp Gerontol. 2014;52:70–76. doi: 10.1016/j.exger.2014.01.011. [DOI] [PubMed] [Google Scholar]
- Deschenes MR, Holdren AN, McCoy RW. Adaptations to short-term muscle unloading in young and aged men. Med Sci Sports Exerc. 2008;40:856–863. doi: 10.1249/MSS.0b013e318164f4b6. [DOI] [PubMed] [Google Scholar]
- Jones DA, de Ruiter CJ, de Haan A. Change in contractile properties of human muscle in relationship to the loss of power and slowing of relaxation seen with fatigue. J Physiol. 2006;576:913–922. doi: 10.1113/jphysiol.2006.116343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Justice JN, Mani D, Pierpoint LA, Enoka RM. Fatigability of the dorsiflexors and associations among multiple domains of motor function in young and old adults. Exp Gerontol. 2014;55:92–101. doi: 10.1016/j.exger.2014.03.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kent-Braun JA. Skeletal muscle fatigue in old age: whose advantage? Exerc Sport Sci Rev. 2009;37:3–9. doi: 10.1097/JES.0b013e318190ea2e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laforest S, St-Pierre DMM, Cyr J, Gayton D. Effects of age and regular exercise on muscle strength and endurance. Eur J Appl Physiol Occup Physiol. 1990;60:104–111. doi: 10.1007/BF00846029. [DOI] [PubMed] [Google Scholar]
- Lanza IR, Towse TF, Caldwell GE, Wigmore DM, Kent-Braun JA. Effects of age on human muscle torque, velocity, and power in two muscle groups. J Appl Physiol. 2003;95:2361–2369. doi: 10.1152/japplphysiol.00724.2002. [DOI] [PubMed] [Google Scholar]
- Lanza IR, Russ DW, Kent-Braun JA. Age-related enhancement of fatigue resistance is evident in men during both isometric and dynamic tasks. J Appl Physiol. 2004;97:967–975. doi: 10.1152/japplphysiol.01351.2003. [DOI] [PubMed] [Google Scholar]
- Lanza IR, Befroy DE, Kent-Braun JA. Age-related changes in ATP-producing pathways in human skeletal muscle in vivo. J Appl Physiol. 2005;99:1736–1744. doi: 10.1152/japplphysiol.00566.2005. [DOI] [PubMed] [Google Scholar]
- Lanza IR, Larsen RG, Kent-Braun JA. Effects of old age on human skeletal muscle energetics during fatiguing contractions with and without blood flow. J Physiol. 2007;583:1093–1105. doi: 10.1113/jphysiol.2007.138362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Layec G, Trinity JD, Hart CR, Kim S-E, Groot HJ, Le Fur Y, Sorensen JR, Jeong E-K, Richardson RS. In vivo evidence of an age-related increase in ATP cost of contraction in the plantar flexor muscles. Clin Sci (Lond) 2014;126:581–592. doi: 10.1042/CS20130442. [DOI] [PubMed] [Google Scholar]
- Layec G, Hart CR, Trinity JD, Le Fur Y, Jeong E-K, Richardson RS. Skeletal muscle work effigiency with age: the role of non-contractile processes. Clin Sci. 2015;128:213–223. doi: 10.1042/CS20140274. [DOI] [PubMed] [Google Scholar]
- Lindstrom B, Lexell J, Gerdle B, Downham D. Skeletal muscle fatigue and endurance in young and old men and women. J Gerontol Biol Sci. 1997;52:59–66. doi: 10.1093/gerona/52A.1.B59. [DOI] [PubMed] [Google Scholar]
- McNeil CJ, Rice CL. Fatigability is increased with age during velocity-dependent contractions of the dorsiflexors. J Gerontol Ser A Biol Sci Med Sci. 2007;62:624–629. doi: 10.1093/gerona/62.6.624. [DOI] [PubMed] [Google Scholar]
- Mcphee JS, Maden-Wilkinson TM, Narici MV, Jones DA, Degens H. Knee extensor fatigue resistance of young and older men and women performing sustained and brief intermittent isometric contractions. Muscle Nerve. 2014;50:393–400. doi: 10.1002/mus.24174. [DOI] [PubMed] [Google Scholar]
- Newham DJ, Jones DA, Turner DL, McIntyre D. The metabolic costs of different types of contractile activity of the human adductor pollicis muscle. J Physiol. 1995;488:815–819. doi: 10.1113/jphysiol.1995.sp021013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petrella JK, Kim J, Tuggle SC, Hall SR, Bamman MM. Age differences in knee extension power, contractile velocity, and fatigability. J Appl Physiol. 2005;98:211–220. doi: 10.1152/japplphysiol.00294.2004. [DOI] [PubMed] [Google Scholar]
- Pojednic RM, Clark DJ, Patten C, Reid K, Phillips EM, Fielding RA. The specific contributions of force and velocity to muscle power in older adults. Exp Gerontol. 2012;47:608–613. doi: 10.1016/j.exger.2012.05.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Power GA, Dalton BH, Rice CL, Vandervoort AA. Reproducibility of velocity-dependent power: before and after lengthening contractions. Appl Physiol Nutr Metab. 2011;36:626–633. doi: 10.1139/h11-068. [DOI] [PubMed] [Google Scholar]
- Power GA, Dalton BH, Rice CL. Human neuromuscular structure and function in old age: a brief review. J Sport Heal Sci. 2013;2:215–226. doi: 10.1016/j.jshs.2013.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Power GA, Allen MD, Booth WJ, Thompson RT, Marsh GD, Rice CL. The influence on sarcopenia of muscle quality and quantity derived from magnetic resonance imaging and neuromuscular properties. Age (Dordr) 2014;36:9642. doi: 10.1007/s11357-014-9642-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raj IS, Bird SR, Shield AJ. Aging and the force-velocity relationship of muscles. Exp Gerontol. 2010;45:81–90. doi: 10.1016/j.exger.2009.10.013. [DOI] [PubMed] [Google Scholar]
- Rawson ES. Enhanced fatigue resistance in older adults during repeated sets of intermittent contractions. J Strength Cond Res. 2010;24:251–256. doi: 10.1519/JSC.0b013e3181a8f7cf. [DOI] [PubMed] [Google Scholar]
- Roos MR, Rice CL, Connelly DM, Vandervoort AA. Quadriceps Muscle strength, contractile properties, and motor unit firing rates in young and old men. Muscle Nerve. 1999;22:1094–1103. doi: 10.1002/(SICI)1097-4598(199908)22:8<1094::AID-MUS14>3.0.CO;2-G. [DOI] [PubMed] [Google Scholar]
- Tevald MA, Foulis SA, Lanza IR, Kent-Braun JA. Lower energy cost of skeletal muscle contractions in older humans. Am J Physiol Regul Integr Comp Physiol. 2010;298:R729–R739. doi: 10.1152/ajpregu.00713.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson BJ, Conchola EC, Palmer TB, Stock MS. Effects of aging on maximal and rapid velocity capacities of leg extensors. Exp Gerontol. 2014;58:128–131. doi: 10.1016/j.exger.2014.07.019. [DOI] [PubMed] [Google Scholar]
- Valour D, Ochala J, Ballay Y, Pousson M. The influence of ageing on the force—velocity—power characteristics of human elbow flexor muscles. Exp Gerontol. 2003;38:387–395. doi: 10.1016/S0531-5565(02)00265-6. [DOI] [PubMed] [Google Scholar]
- Yoon T, Schlinder-Delap B, Hunter SK. Fatigability and recovery of arm muscles with advanced age for dynamic and isometric contractions. Exp Gerontol. 2013;48:259–268. doi: 10.1016/j.exger.2012.10.006. [DOI] [PMC free article] [PubMed] [Google Scholar]