

Abstract
Classical swine fever, caused by classical swine fever virus (CSFV), is a highly contagious disease that results in enormous economic losses in pig industries. The E2 protein is one of the main structural proteins of CSFV and is capable of inducing CSFV-neutralizing antibodies and cytotoxic T lymphocyte (CTL) activities in vivo. Thymosin α-1 (Tα1), an immune-modifier peptide, plays a very important role in the cellular immune response. In this study, genetically engineered Lactobacillus plantarum bacteria expressing CSFV E2 protein alone (L. plantarum/pYG-E2) and in combination with Tα1 (L. plantarum/pYG-E2-Tα1) were developed, and the immunogenicity of each as an oral vaccine to induce protective immunity against CSFV in pigs was evaluated. The results showed that recombinant L. plantarum/pYG-E2 and L. plantarum/pYG-E2-Tα1 were both able to effectively induce protective immune responses in pigs against CSFV infection by eliciting immunoglobulin A (IgA)-based mucosal, immunoglobulin G (IgG)-based humoral, and CTL-based cellular immune responses via oral vaccination. Significant differences (P < 0.05) in the levels of immune responses were observed between L. plantarum/pYG-E2-Tα1 and L. plantarum/pYG-E2, suggesting a better immunogenicity of L. plantarum/pYG-E2-Tα1 as a result of the Tα1 molecular adjuvant that can enhance immune responsiveness and augment specific lymphocyte functions. Our data suggest that the recombinant Lactobacillus microecological agent expressing CSFV E2 protein combined with Tα1 as an adjuvant provides a promising strategy for vaccine development against CSFV.
INTRODUCTION
Classical swine fever virus (CSFV), a member of the Pestivirus genus of the Flaviviridae family, is a small, enveloped, single-stranded RNA virus. The genome of CSFV consists of a positive-stranded RNA molecule of about 12.3 kb, encoding a single open reading frame that is translated into a 3,898-amino-acid polyprotein, giving rise to different CSFV proteins after coprocessing and after translational processing (1, 2). Of these, the E2 structural protein encompasses major antigenic domains and cytotoxic T lymphocyte (CTL) epitopes, suggesting a promising candidate for an immunogen with the capacity to induce neutralizing antibodies and CTL activities against CSFV (3–10). CSFV often causes severe and lethal disease in pigs, resulting in enormous economic losses in pig industries (11, 12). Therefore, the development of effective vaccines against CSFV infection is an issue of growing importance.
For the development of CSFV vaccines, the roles of neutralizing antibody-based humoral immune responses (13, 14) and CTL-based cellular immune responses (15–20) have been both highlighted. Furthermore, the invasion of CSFV often initiates at mucosal surfaces, particularly intestinal tissues, and thus, vaccination inducing IgA-based protective mucosal immunity via the mucosal approach could effectively prevent the virus from entering the body via mucosa and its further spread to the systemic circulation. Taken together, the design of new vaccines that could trigger protective antiviral mucosal and systemic immune responses will provide a promising strategy for the development of a vaccine against CSFV infection.
In this respect, mucosal immunization has been proven to be an effective approach (21, 22). The key here is to develop efficient vaccine carriers that can prevent immunogens from being inactivated by gastric acid or digested in the anterior part of the gut. One promising approach relies on the use of live vehicles (23). The potential of lactic acid bacteria to deliver heterologous antigens to the mucosal immune system has been investigated during the last decade (20, 24–29); this approach offers a number of advantages over the traditional parenteral vaccination, such as noninvasiveness and the possibility of eliciting both mucosal and systemic immune responses. Moreover, probiotic lactobacilli are well known for beneficial effects on the health of humans and animals, and they can survive transit through the upper gastrointestinal tract and colonize the intestinal tract (20, 30, 31), showing intrinsic adjuvant activity (32, 33).
Thymosin α-1 (Tα1) is a nontoxic immune-modifier peptide hormone secreted by the thymus and plays a very important role in cellular immune responses (34, 35), being able to trigger maturational events in lymphocytes, augment T-cell function and antibody production, and promote reconstitution of immune defects, which suggests that it may be an ideal molecular vaccine adjuvant. In the present study, a recombinant Lactobacillus strain expressing CSFV E2 protein combined with Tα1 was developed, and its immunogenicity as an oral vaccine to induce protective immune responses to CSFV in pigs was analyzed. Our data showed that oral vaccination with the recombinant strain was able to induce protective immunity against CSFV infection at the mucosal, humoral, and cellular levels, indicating a new vaccine strategy for the control of CSFV.
MATERIALS AND METHODS
Virus, bacterium, and plasmid.
Lactobacillus plantarum HA33-1, a naturally plasmid-free isolate from yogurt, and surface-expressed plasmid pYG301 (containing the bacteriophage T7 promoter, the cell wall anchor motif from Streptococcus pyogenes M6 protein, the chloramphenicol resistance determinant, and repA and repC replication elements) were used in this work. L. plantarum was grown anaerobically in MRS broth (Sigma, USA) at 37°C. CSFV strain Shimen was propagated in porcine kidney 15 (PK-15) cells in minimum essential medium (MEM; Life Technologies, USA), pH 7.0, plus 10% fetal bovine serum (FBS; Life Technologies, USA) at 37°C in a 5% CO2 incubator. Virus titers were determined by endpoint dilution titrations on PK-15 cells according to a previously described method (18, 36).
Construction of recombinant Lactobacillus strain expressing E2-Tα1.
Genomic RNA of CSFV propagated in PK-15 cells was extracted using a MiniBEST universal RNA extraction kit (TaKaRa, Japan) according to the manufacturer's instructions, followed by reverse transcription to generate the first-strand cDNA using an Omniscript RT kit (Qiagen, Germany), following the manufacturer's instructions. Then, the CSFV E2 gene was synthesized by PCR with HiFi Taq DNA polymerase (TaKaRa, Japan), using the cDNA as the template, with cycling conditions of 35 cycles of 94°C for 45 s, 61.5°C for 45 s, 72°C for 60 s, and a final extension at 72°C for 10 min. The PCR primers were as follows: 5′-CTCGAGATGCGGCTAGCCTGCAAGGAAGATTACA-3′ (forward), containing an XhoI site (underlined), and 5′-GGATCCGG-AGGAGGAGGAGGAGGAACCAGCGGCGAGTTGTTC-3′ (reverse), containing a BamHI site and partial oligonucleotides encoding a linker (underlined). Then, the CSFV E2 gene (amplicon size, 1,155 bp) was inserted as an XhoI/BamHI fragment into plasmid pYG301, giving rise to pYG-E2. pET-Tα1 was constructed in our laboratory as follows: genomic DNA of calf thymus was extracted using a tissue/cell/blood genome DNA extraction kit (Qiagen, Germany), and the gene encoding Tα1 was amplified and inserted into plasmid pET-28a, generating pET-Tα1. The Tα1 gene was amplified from pET-Tα1 by PCR with the oligonucleotides 5′-GGATCCGGAGGAGGAGGAGGAGGAATGTCAGACGCAGCCGTA3′ (forward), containing a BamHI site and partial oligonucleotides encoding a linker (underlined), and 5′-GGTACCTTATCTTCCATTTTCTGCCTCTTCCACAACTTC-3′ (reverse), containing a KpnI site (underlined), with cycling conditions of 35 cycles of 94°C for 15 s, 57.5°C for 30 s, 72°C for 15 s, and a final extension at 72°C for 5 min. Then, the Tα1 gene (amplicon size of 102 bp) was cloned as a BamHI/KpnI fragment into pYG-E2, giving rise to recombinant plasmid pYG-E2-Tα1, where E2 and Tα1 were linked together by oligonucleotides encoding a flexible linker (GGGGGGGSGGGGGG). Then, pYG-E2-Tα1 was electroporated into competent cells of L. plantarum HA33-1 using a previously described method (29), generating a recombinant strain named L. plantarum/pYG-E2-Tα1.
Fusion protein expression.
L. plantarum/pYG-E2-Tα1 was cultured overnight in MRS medium plus 2% xylose at 37°C and then collected by centrifugation at 12,000 × g for 10 min. The cells were lysed by boiling for 10 min in 2× SDS-PAGE loading buffer (1 M Tris-HCl, pH 6.8, 7.0 mM SDS, 0.15 mM bromophenol blue, and 0.2% [vol/vol] glycerol), and the lysates were analyzed via 10% SDS-PAGE. After that, proteins were electrotransferred onto a nitrocellulose membrane (Invitrogen, USA), and the immunoblots were developed using mouse anti-CSFV E2 antiserum (to prepare mouse anti-CSFV E2 antiserum, the CSFV E2 protein was prokaryotically expressed by Escherichia coli, purified, and used to immunize BALB/c mice, and then the polyclonal antisera were collected and purified by IgG antibody screening spin plates [Pierce, USA]) and mouse anti-Tα1 antibody (Abcam, England). Horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG (Sigma, USA) diluted at 1:2,000 was used as the secondary antibody, and then immunolabeled bands were visualized with a chemiluminescent substrate reagent (Pierce, USA) according to the manufacturer's instructions. An immunofluorescence assay was also performed to determine the surface-displayed localization of the E2-Tα1 fusion protein from L. plantarum/pYG-E2-Tα1. In brief, L. plantarum/pYG-E2-Tα1 cells induced with xylose were collected by centrifugation, and then the cells were incubated with mouse anti-CSFV E2 antiserum (1:1,000) at 37°C for 2 h. After washing the cells thrice with sterile phosphate-buffered saline (PBS) to remove uncombined antibodies, the cells were incubated with fluorescein isothiocyanate-conjugated goat anti-mouse IgG (Sigma, USA) containing 1% Evans blue at 37°C for 2 h and then analyzed using confocal microscopy. Noninduced L. plantarum/pYG-E2-Tα1 was used as a control.
Oral immunization.
All animal experiments were approved by the Experimental Animal Care Ethics Committee of Northeast Forestry University. The L. plantarum/pYG-E2-Tα1 and L. plantarum/pYG-E2 cells that were induced by xylose overnight were centrifuged and resuspended in suspension buffer (0.2 mol/liter NaHCO3, 5% casein acid hydrolysate, 0.5% sucrose) to a concentration of ca. 1010 CFU/ml. Then, the recombinant strain suspensions were fully mixed with pig feed raw materials, and a manual extruder was used to cold extrude the mixtures into particles (0.5 cm by 1 cm on average) containing, on average, ca. 109 CFU/g of the recombinant strain. Experimental pigs (5 to 7 weeks old with no maternal antibody to CSFV) and miniature pigs of d/d haplotype were kept under specific-pathogen-free conditions. In order to test the immunogenicity of L. plantarum/pYG-E2-Tα1, vaccine groups of 15 pigs and 3 miniature pigs were vaccinated orally with 109 CFU/g L. plantarum/pYG-E2-Tα1 per kg of body weight, following a previously described immune protocol (20). Another group of 15 pigs and 3 miniature pigs orally immunized with L. plantarum/pYG-E2 were used as vaccine controls. Control groups of 15 pigs received pellet feed containing L. plantarum or PBS, respectively. On days 0, 7, 21, 35, 42, and 56 after the first immunization, blood samples were collected in vacuum tubes from each group and placed overnight at 4°C, after which the blood samples were centrifuged at 2,000 × g for 15 min and the sera were stored at −70°C until required. Moreover, intestinal lavages were collected using a previously described method (37). In brief, three pigs were randomly selected from each group, and after an overnight fast, lavage was performed using polyethylene glycol electrolyte lavage solution. The lavage solution was given at a rate of 250 ml every 15 min for a period of 2 h, and intestinal lavages were collected once the material passed from the rectum became liquid, clear, and free of fecal material. An approximately 10-ml sample was collected from each pig, filtered, centrifuged, treated with protease inhibitors, and then stored at −70°C.
ELISA of anti-CSFV E2 antibody levels.
A 96-well ELISA plate was coated overnight with whole CSFV viruses propagated in PK-15 cells at 4°C. Following saturation of the wells with PBS–10% bovine serum albumin (BSA) at 37°C for 2 h, sera or intestinal lavages were used as the primary antibody and incubated at 37°C for 1 h. After that, bound antibodies were detected using HRP-conjugated goat anti-pig IgG or IgA at a dilution of 1:2,000 (Sigma, USA) at 37°C for 1 h. Following washing the plate thrice with PBS–0.05% Tween 20, color development was performed using tetramethylbenzidine (Qiagen, Germany) as the colorimetric substrate, followed by measuring the absorbance at 450 nm.
Neutralization ability of antibodies obtained from vaccinated pigs.
A virus plaque reduction assay was performed to evaluate the neutralization ability of the serum IgG and mucosal IgA obtained from the vaccinated pigs. In brief, 50-μl amounts of undiluted samples were prepared in a 96-cell plate. CSFV was adjusted to a concentration of 200 50% tissue culture infective doses (TCID50) in 50 μl of virus diluent (10% concentrated Hanks balanced salt solution, 0.1% BSA, pH 7.4) and was added into the wells, mixed, and incubated at 37°C for 1 h. Then, 100 μl of PK-15 cells was added to the antibody-virus mixture and incubated in a 5% CO2 incubator at 37°C for 4 days. The overlay medium was discarded, and the wells were washed thrice with sterile PBS, pH 7.4, and stained with 1% crystal violet solution. Differences in the number of plaques formed between treatments were examined for the level of significance by Student's t test after analysis of variance.
Cellular immune responses.
To evaluate cellular immune responses induced in vaccinated pigs, the CTL activities and levels of lymphokines (gamma interferon [IFN-γ], interleukin 2 [IL-2], and tumor necrosis factor alpha [TNF-α]) were determined. The levels of IFN-γ, IL-2, and TNF-α in sera were determined by using the IFN-γ, IL-2, and TNF-α swine enzyme-linked immunosorbent assay (ELISA) kits (Invitrogen, USA) according to the manufacturer's instructions. For the determination of CTL activities, experimental miniature pigs of d/d haplotype were bled on day 42 after the first immunization to prepare peripheral blood mononuclear cells (PBMCs) by a previously described method (38), and intestinal tissue samples were obtained via laparotomy, from which lamina propria lymphocytes were prepared (39). These immune cells were seeded in a 96-well microplate to a concentration of 1 × 106/ml in RPMI 1640 (Sigma, USA) plus 10% FBS and then restimulated with 1 × 105 TCID50 CSFV per well for 5 days to generate effector cells. MAX cells (kindly provided by Y. Wang [NEFU, Harbin, China]) were infected with CSFV for 48 h, giving rise to target cells. In the cytotoxicity assay, 1 × 106/ml target cells were labeled with 100 μCi of Na251CrO4 (Amersham, USA) in a 5% CO2 incubator at 37°C for 1 h. After washing, the cells were resuspended in culture medium to a concentration of 1 × 103 cells per well. Then, the CSFV-infected target cells were added to various concentrations of effector cells (ranging from 1.25 × 104 to 5.0 × 104 cells per well) and incubated in a 5% CO2 incubator at 37°C for 4 h. The chromium levels in the supernatants were measured using a COBRA auto-gamma model 5003 gamma counter (Packard, USA). The percentage of specific cytolytic activity was calculated by a previously described method (20). Mock-infected target cells were used as a control, and all experiments were performed in triplicate.
Virus challenge experiment.
On day 35 after the first immunization, pigs that were orally vaccinated with L. plantarum/pYG-E2-Ta1, L. plantarum/pYG-E2, or PBS were challenged orally with 105 TCID50 of CSFV strain Shimen. These pigs were examined daily for clinical symptoms and fever, defined by the rectal temperature of the pig, and survival rates were recorded postchallenge.
Statistical analysis.
Data are presented as the mean results ± standard errors from three replicates per test in one experiment, repeated thrice. Student's t test was used to analyze differences between immunized and control groups. A P value of <0.05 was considered significant.
RESULTS
Expression of fusion protein.
The cell lysates of L. plantarum/pYG-E2-Tα1 were analyzed via SDS-PAGE and Western blot analysis. An appropriate-sized protein (about 46 kDa) was expressed in xylose-induced L. plantarum/pYG-E2-Tα1 cells (Fig. 1, lanes 1 and 2) but not in noninduced cells (Fig. 1, 3 and 4). Next, an immunoreactive band in a position similar to that observed via SDS-PAGE was developed by Western blotting using mouse anti-CSFV E2 antiserum (Fig. 1, lane a) and mouse anti-Tα1 antibody (Fig. 1, lane e); there was no corresponding band observed in noninduced L. plantarum/pYG-E2-Tα1 cells (Fig. 1, lanes b and f). Meanwhile, the E2 protein expressed by L. plantarum/pYG-E2 cells was analyzed by SDS-PAGE (Fig. 1, lane 5) and Western blotting (Fig. 1, lane c). Furthermore, an immunofluorescence assay showed that there was green-yellow fluorescence on the surface of L. plantarum/pYG-E2-Tα1 cells induced by xylose (Fig. 2A) but not on the surface of noninduced cells, which were dyed red by Evans blue (Fig. 2B). Our results indicated that the protein of interest can be efficiently expressed by L. plantarum/pYG-E2-Tα1 and is displayed on the surface of cells.
FIG 1.
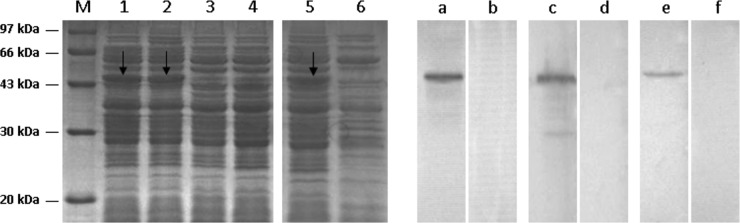
Expression of the E2-Tα1 fusion protein from L. plantarum/pYG-E2-Tα1 was identified by SDS-PAGE and Western blotting. M, molecular mass marker. An ∼46-kDa protein was expressed in L. plantarum/pYG-E2-Tα1 cells induced by xylose (lanes 1 and 2) but not in noninduced cells (lanes 3 and 4). Western blotting showed that an immunoreactive band was developed in a position similar to that observed by SDS-PAGE using mouse anti-CSFV E2 antibody (lane a) and mouse anti-Tα1 antibody (lane e), respectively. However, there was no corresponding band observed in noninduced L. plantarum/pYG-E2-Tα1 (lanes b and f). The E2 protein expressed by L. plantarum/pYG-E2 was identified via SDS-PAGE (lane 5) and Western blotting (lane c), and there was no corresponding band observed in the noninduced cells (lanes 6 and d).
FIG 3.
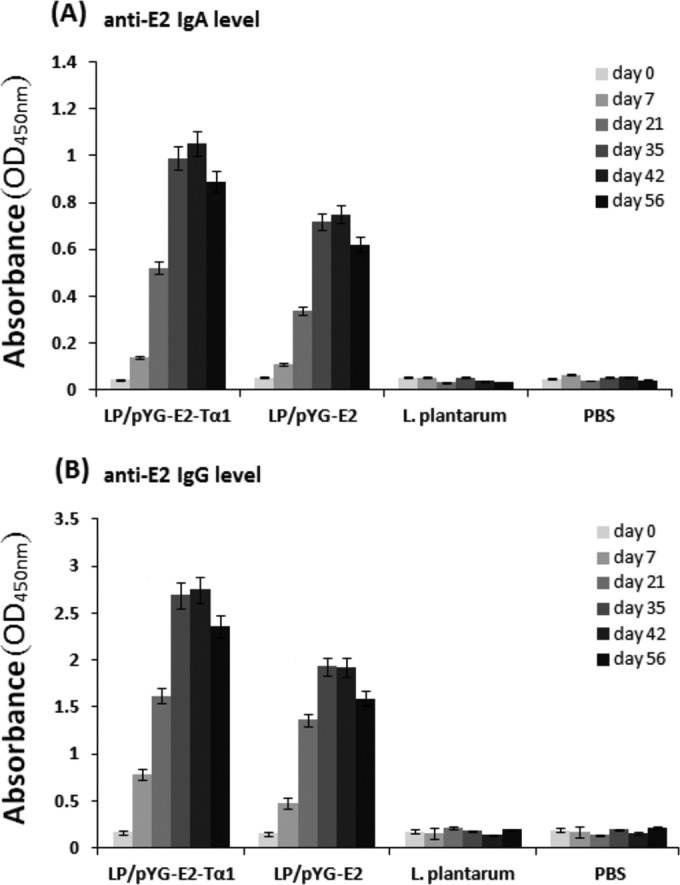
Anti-CSFV E2 IgA levels in intestinal lavage fluid (A), and anti-CSFV E2 IgG levels in serum (B). Pigs were orally immunized with L. plantarum/pYG-E2-Tα1 or L. plantarum/pYG-E2. Control groups of pigs received L. plantarum and PBS, respectively. On days 0, 7, 21, 35, 42, and 56 after the first immunization, the IgA and IgG levels were determined by ELISA. The data show that both mucosal and systemic immune responses against CSFV can be efficiently elicited by L. plantarum/pYG-E2-Tα1 and L. plantarum/pYG-E2 via oral administration and that L. plantarum/pYG-E2-Tα1 showed a better immunogenicity than L. plantarum/pYG-E2 as a result of the Tα1 molecular adjuvant. Compared to the results for control groups, significant levels of IgA and IgG were observed both in the L. plantarum/pYG-E2-Tα1 group and the L. plantarum/pYG-E2 group (P < 0.01) after the second booster (on day 30). Moreover, significant levels (P < 0.05) of IgA and IgG were elicited in the L. plantarum/pYG-E2-Tα1 group compared to the levels in the L. plantarum/pYG-E2 group after the second booster. Bars represent the mean results ± standard errors of the means for each group.
FIG 4.
Inhibition of virus plaque formation by anti-CSFV E2 IgA (A) and anti-CSFV E2 IgG (B) collected from pigs orally immunized with L. plantarum/pYG-E2-Tα1 and L. plantarum/pYG-E2, respectively. The antibodies were collected on days 0, 7, 21, 35, 42, and 56 after the first immunization. The inhibition value was expressed as the maximum reduction in the number of plaques. Results are the means ± standard errors of the means.
FIG 2.

Immunofluorescence reaction of CSFV E2 protein on the cell surface of L. plantarum/pYG-E2-Tα1. The immunofluorescence assay showed green-yellow fluorescence on the surface of L. plantarum/pYG-E2-Tα1 cells induced by xylose (A), while no fluorescence appeared on the surface of the same cells without induction, which were dyed red by Evans blue (B).
Immune responses to CSFV induced in pigs.
The capacity of L. plantarum/pYG-E2-Tα1 to induce mucosal and systemic immune responses in pigs via oral administration was evaluated by determining the presence of anti-E2 IgG and IgA antibodies, respectively, via ELISA. As shown by the data in Fig. 3, compared to the results for the control groups, significant levels (P < 0.01) of antigen-specific mucosal IgA (Fig. 3A) and antigen-specific systemic IgG (Fig. 3B) were elicited in pigs orally vaccinated with L. plantarum/pYG-E2-Tα1 or L. plantarum/pYG-E2 after the second booster, while there was no substantial difference (P > 0.05) observed among the negative-control groups before and after immunization. Compared to the immunogenicity of the L. plantarum/pYG-E2 strain, which expresses only E2 protein, the L. plantarum/pYG-E2-Tα1 strain, which expresses E2 protein in combination with Tα-1, promoted stronger antigen-specific immune responses at both the mucosal and the systemic level (P < 0.05), indicating an effective vaccine molecular adjuvant feature of Tα-1.
CSFV-neutralizing ability of antibodies.
Virus plaque reduction assays were carried out to determine the neutralizing ability of antibodies obtained from pigs orally immunized with L. plantarum/pYG-E2-Tα1 on days 0, 7, 21, 35, 42, and 56 after the first immunization. The results showed that the presence of anti-CSFV E2 IgA (Fig. 4A) or IgG (Fig. 4B) antibody in the culture medium conferred significant neutralizing ability against CSFV infection (P < 0.05), and the antibody samples obtained from the pigs orally vaccinated with L. plantarum/pYG-E2-Tα1 showed better neutralizing ability than those obtained from the pigs orally immunized with L. plantarum/pYG-E2.
Specific cytotoxic responses induced by L. plantarum/pYG-E2-Tα1 in vivo.
The capacity of L. plantarum/pYG-E2-Tα1 to elicit specific cytotoxic responses against CSFV in pigs at mucosal and systemic levels was analyzed in vivo. PBMCs and lamina propria lymphocytes obtained from the miniature pigs orally vaccinated with L. plantarum/pYG-E2-Tα1 on day 42 after the first immunization were restimulated with CSFV in vitro, and their cytotoxic activities were determined based on the lysis of CSFV-infected target cells labeled with chromium. Mock-infected MAX cells were used as a control. The results showed that specific CTL activity against CSFV-infected MAX cells was observed at both the mucosal level (Fig. 5A) and the systemic level (Fig. 5B) when the CSFV-restimulated PBMCs and lamina propria lymphocytes, respectively, were used as effector cells. Moreover, the levels of lymphokines IFN-γ (Fig. 6), IL-2 (Fig. 7), and TNF-α (Fig. 8) induced in pigs were determined. L. plantarum/pYG-E2-Tα1 triggered a more effective cellular immune responses than L. plantarum/pYG-E2 due to the adjuvant effect of Tα1.
FIG 5.

CSFV-specific CTL activities of effector cells restimulated with CSFV against CSFV-infected target cells. Lamina propria lymphocytes (A) and PBMCs (B) obtained from pigs orally vaccinated with L. plantarum/pYG-E2-Tα1 (●) and L. plantarum/pYG-E2 (▲) were restimulated with CSFV in vitro to be used as effector cells. The cytotoxicity of the T cells was determined in a chromium release assay by lysis of virus-sensitized MAX cells as targets. Mock-infected cells (○) were used as a control.
FIG 6.
Levels of IFN-γ induced in pigs after oral immunization as determined by ELISA. Bars represent the mean results ± standard errors of the means in each group.
FIG 7.
Levels of IL-2 induced in pigs after oral immunization as determined by ELISA. Bars represent the mean results ± standard errors of the means in each group.
FIG 8.
Levels of TNF-α induced in pigs after oral immunization as determined by ELISA. Bars represent the mean results ± standard errors of the means in each group.
CSFV challenge against pigs vaccinated with L. plantarum/pYG-E2-Tα1.
A virus challenge experiment was performed to evaluate the protective effect of L. plantarum/pYG-E2-Tα1 vaccination of pigs against CSFV infection postvaccination. The results showed an effective protection rate of 80% against CSFV challenge in pigs vaccinated with L. plantarum/pYG-E2-Tα1, which was higher than that (53.3%) in pigs vaccinated with L. plantarum/pYG-E2, while the control group of pigs that received PBS only developed severe clinical symptoms of classical swine fever (CSF) after virus challenge and were euthanized when moribund. The pigs orally vaccinated with L. plantarum/pYG-E2-Tα1 or L. plantarum/pYG-E2 showed a virus challenge-induced peak fever (means of 39.7°C for the L. plantarum/pYG-E2-Tα1 group and 40.9°C for the L. plantarum/pYG-E2 group) from day 4 to 6 postchallenge, and on days 7 to 9 after challenge, the surviving pigs recovered gradually from clinical signs of CSFV, while the PBS group of pigs showed a longer period of high fever (mean of 42.1°C) after virus infection, until they were euthanized. Moreover, the vaccine group of pigs orally immunized with L. plantarum/pYG-E2-Tα1 showed a reduced mortality pattern and significantly improved survival relative to the outcomes for the vaccine control group of pigs that received L. plantarum/pYG-E2, indicating that a better protective effect was obtained from L. plantarum/pYG-E2-Tα1 via oral immunization.
DISCUSSION
Although CSF has been efficiently controlled by stamping-out methods and vaccination or a combination of the two, the eradication strategy has inevitably caused enormous economic losses worldwide. Moreover, the European Union has banned vaccination using traditional CSFV avirulent vaccines against CSFV since 1990 because infections induced by vaccines or field virus cannot be distinguished serologically (17). Therefore, the development of new efficacious vaccines to protect animals against CSFV is being pursued. For vaccines against CSFV such as subunit, DNA, or vector vaccines, the vaccine delivery strategies all focus on the intramuscular injection route. The initial infection of CSFV mainly occurs at the mucosal tissues, particularly in the intestine. Therefore, it is important to develop vaccines that not only induce effective mucosal immune responses to prevent the virus from invading at the mucosa but also elicit specific immunity in systemic lymphoid tissues once viral infections that have gained entrance through the mucosal surfaces become systemic. Thus, the design of vaccines delivered via the mucosal route to induce protective immunity against CSFV will be an effective avenue. The key for vaccination via the mucosal route is that the immunogen must be protected from premature disintegration and degradation due to low pH and proteases in the gastrointestinal tract. In this study, a strain of L. plantarum expressing CSFV E2 protein in combination with Tα1 as an adjuvant was developed, and its immunogenicity as an oral vaccine to induce protective immune responses against CSFV in pigs was investigated.
For vaccine research, one important step is to identify the immune responses and immune defenses stimulated by an immunogen against infection in the host. The role of humoral immunity in CSFV-infected animals has been highlighted (13, 14). It is widely known that the E2 glycoprotein of CSFV is the principal target for inducing neutralizing antibodies against CSFV infection (6–10). Our results showed that the recombinant L. plantarum strain L. plantarum/pYG-E2-Tα1 was able to elicit significant levels of systemic immune response, producing anti-CSFV E2 IgG with high virus-neutralizing activity in pigs via oral vaccination. Recently, several experimental studies of CSFV vaccines have reported that humoral immune responses to CSFV E2 induced via intramuscular injection conferred effective protection against CSFV, such as E2 protein expressed in orf virus (6), in yeast (9), and in baculovirus (10), as well as milk-derived E2 protein (8). Compared to the results for these E2 subunit vaccines, the Lactobacillus-based E2 live vector vaccine developed in this study could induce not only an IgG-based humoral immune response but also an IgA-based mucosal immune response via oral vaccination. IgA is the predominant antibody at the mucosal surface, produced locally at a level that exceeds those of other immunoglobulins and playing an important role in preventing the invasion of pathogens (40). Although E2 protein loaded on poly(d,l-lactide-co-glycolide) (PLGA) microspheres could elicit mucosal and systemic immune responses via the gastrointestinal route (7), the Lactobacillus-based live vector used in this work offers a number of advantages over the PLGA microsphere vector, such as colonization ability in the intestinal tract and probiotic and intrinsic adjuvant activities (30, 32).
Furthermore, cellular immune responses, especially the production of CSFV-specific CD8+ CTLs, represent an important defense mechanism in the elimination of cells infected by CSFV (5, 17, 18). We evaluated the capacity of L. plantarum/pYG-E2-Tα1 to stimulate CSFV-specific CTL responses in vivo and in vitro. Our findings showed that a CSFV-specific CTL response was determined in the intestinal mucosal tissues of miniature pigs vaccinated orally with L. plantarum/pYG-E2-Tα1 but not in unimmunized animals, which indicated that the effector cells obtained by CSFV restimulation of lamina propria lymphocytes obtained from vaccinated pigs were able to recognize naturally CSFV-infected target cells. The result indicates that CSFV replication within local tissues could be effectively controlled following the induction of a local CD8+ CTL response coupled with the mucosal antibody IgA prior to systemic dissemination. Moreover, CSFV-specific CD8+ CTL were also induced systemically in pigs by L. plantarum/pYG-E2-Tα1 via oral vaccination, and significant levels of the lymphokines IFN-γ, IL-2, and TNF-α were determined to have been induced in pigs. Subsequently, we tested the protective capacity of L. plantarum/pYG-E2-Tα1 against CSFV challenge of pigs, and the pigs immunized with L. plantarum/pYG-E2-Tα1 had obtained effective protection against CSFV challenge, while control groups of pigs developed severe clinical symptoms of CSF with a high fever after virus challenge.
In order to evaluate the adjuvant effect of Tα1, we constructed two recombinant strains of L. plantarum, strain L. plantarum/pYG-E2, expressing CSFV E2 protein only, and strain L. plantarum/pYG-E2-Tα1, expressing CSFV E2 protein in combination with Tα1 polypeptide, and analyzed their immunogenicities. Our data showed that both mucosal and systemic immune responses against CSFV could be efficiently elicited by these two recombinant strains via oral administration. In contrast, L. plantarum/pYG-E2-Tα1 showed better immunogenicity than L. plantarum/pYG-E2, as a result of the Tα1 molecular adjuvant. Significant differences in the IgA and IgG levels (P < 0.05) induced were observed in pigs vaccinated with L. plantarum/pYG-E2-Tα1 compared to their levels in pigs vaccinated with L. plantarum/pYG-E2, particularly for cellular immune responses. Tα1 can affect immune-regulatory T-cell function, enhance immune responsiveness, augment specific lymphocyte functions, including T-cell maturation, antibody production, and T-cell-mediated cytotoxicity, and promote the expression of lymphokines such as TNF-α, IFN-γ, and IL-2, suggesting a safe and effective vaccine molecular adjuvant.
In conclusion, our results clearly demonstrate that the recombinant L. plantarum strain expressing CSFV E2 protein in combination with Tα1 polypeptide developed in this study provides a safe and effective oral vaccine to induce protective immunity in pigs against CSFV infection, suggesting a promising vaccine formulation for the development of a CSF vaccine.
ACKNOWLEDGMENTS
This work was supported by Natural Science Foundation of Heilongjiang Province (grant C201349) and Special Fund of Harbin Science and Technology Innovation Talent (grant 2012RFQYN032).
REFERENCES
- 1.Meyers G, Rümenapf T, Thiel H-J. 1989. Molecular cloning and nucleotide sequence of the genome of hog cholera virus. Virology 171:555–567. doi: 10.1016/0042-6822(89)90625-9. [DOI] [PubMed] [Google Scholar]
- 2.Meyers G, Thiel H-J. 1996. Molecular characterization of pestiviruses. Adv Virus Res 47:53–118. doi: 10.1016/S0065-3527(08)60734-4. [DOI] [PubMed] [Google Scholar]
- 3.Tarradas J, Argilaguet JM, Rosell R, Nofrarías M, Crisci E, Córdoba L, Pérez-Martín E, Díaz I, Rodríguez F, Domingo M, Montoya M, Ganges L. 2010. Interferon-gamma induction correlates with protection by DNA vaccine expressing E2 glycoprotein against classical swine fever virus infection in domestic pigs. Vet Microbiol 142:51–58. doi: 10.1016/j.vetmic.2009.09.043. [DOI] [PubMed] [Google Scholar]
- 4.Rűmenapf T, Stark R, Meyers G, Thiel HJ. 1991. Structural proteins of hog cholera virus expressed by vaccinia virus: further characterization and induction of protective immunity. J Virol 65:589–597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ceppi M, de Bruin MGM, Seuberlich T, Balmelli C, Pascolo S, Ruggli N, Wienhold D, Tratschin JD, McCullough KC, Summerfield A. 2005. Identification of classical swine fever virus protein E2 as a target for cytotoxic T cells by using mRNA-transfected antigen-presenting cells. J Gen Virol 86:2525–2534. doi: 10.1099/vir.0.80907-0. [DOI] [PubMed] [Google Scholar]
- 6.Voigt H, Merant C, Wienhold D, Braun A, Hutet E, Le Potier MF, Saalmüller A, Pfaff E, Büttner M. 2007. Efficient priming against classical swine fever with a safe glycoprotein E2 expressing Orf virus recombinant (ORFV VrV-E2). Vaccine 25:5915–5926. doi: 10.1016/j.vaccine.2007.05.035. [DOI] [PubMed] [Google Scholar]
- 7.Brandhonneur N, Loizel C, Chevanne F, Wakeley P, Jestin A, Le Potier MF, Le Corre P. 2009. Mucosal or systemic administration of rE2 glycoprotein antigen loaded PLGA microspheres. Int J Pharm 373:16–23. doi: 10.1016/j.ijpharm.2009.01.020. [DOI] [PubMed] [Google Scholar]
- 8.Toledo JR, Barrera M, Farnós O, Gómez S, Rodríguez MP, Aguero F, Ormazabal V, Parra NC, Suárez L, Sánchez O. 2010. Human αIFN co-formulated with milk derived E2-CSFV protein induce early full protection in vaccinated pigs. Vaccine 28:7907–7914. doi: 10.1016/j.vaccine.2010.09.073. [DOI] [PubMed] [Google Scholar]
- 9.Lin GJ, Deng MC, Chen ZW, Liu TY, Wu CW, Cheng CY, Chien MS, Huang C. 2012. Yeast expressed classical swine fever E2 subunit vaccine candidate provides complete protection against lethal challenge infection and prevents horizontal virus transmission. Vaccine 30:2336–2341. doi: 10.1016/j.vaccine.2012.01.051. [DOI] [PubMed] [Google Scholar]
- 10.Zhang H, Li X, Peng G, Tang C, Zhu S, Qian S, Xu J, Qian P. 2014. Glycoprotein E2 of classical swine fever virus expressed by baculovirus induces the protective immune responses in rabbits. Vaccine 32:6607–6613. doi: 10.1016/j.vaccine.2014.10.003. [DOI] [PubMed] [Google Scholar]
- 11.Edwards S, Fukusho A, Lefèvre PC, Lipowski A, Pejsak Z, Roehe P, Westergaard J. 2000. Classical swine fever: the global situation. Vet Microbiol 73:103–119. doi: 10.1016/S0378-1135(00)00138-3. [DOI] [PubMed] [Google Scholar]
- 12.Moennig V. 2000. Introduction to classical swine fever: virus, disease and control policy. Vet Microbiol 73:93–102. doi: 10.1016/S0378-1135(00)00137-1. [DOI] [PubMed] [Google Scholar]
- 13.Terpstra C, Wensvoort G. 1988. The protective value of vaccine induced neutralizing antibody titres in swine fever. Vet Microbiol 16:123–128. doi: 10.1016/0378-1135(88)90036-3. [DOI] [PubMed] [Google Scholar]
- 14.van Gennip HGP, Bouma A, van Rijn PA, Widjojoatmodjo MN, Moormann RJM. 2002. Experimental non-transmissible marker vaccines for classical swine fever (CSF) by transcomplementation of Erns or E2 of CSFV. Vaccine 20:1544–1556. doi: 10.1016/S0264-410X(01)00497-2. [DOI] [PubMed] [Google Scholar]
- 15.Armengol E, Wiesmüller K-H, Wienhold D, Büttner M, Pfaff E, Jung G, Saalmüller A. 2002. Identification of T-cell epitopes in the structural and non-structural proteins of classical swine fever virus. J Gen Virol 83(Pt 3):551–560. [DOI] [PubMed] [Google Scholar]
- 16.Kimman TG, Bianchi ATJ, Wensvoort G, de Bruin TGM, Meliefste C. 1993. Cellular immune response to hog cholera virus (HCV): T cells of immune pigs proliferate in vitro upon stimulation with live HCV, but the E1 envelope glycoprotein is not a major T-cell antigen. J Virol 67:2922–2927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Pauly T, Elbers K, König M, Lengsfeld T, Saalmüller A, Thiel HJ. 1995. Classical swine fever virus-specific cytotoxic T lymphocytes and identification of a T cell epitope. J Gen Virol 76:3039–3049. doi: 10.1099/0022-1317-76-12-3039. [DOI] [PubMed] [Google Scholar]
- 18.Rau H, Revets H, Balmelli C, McCullough KC, Summerfield A. 2006. Immunological properties of recombinant classical swine fever virus NS3 protein in vitro and in vivo. Vet Res 37:155–168. doi: 10.1051/vetres:2005049. [DOI] [PubMed] [Google Scholar]
- 19.van Zijl M, Wensvoort G, De Kluyver E, Hulst M, Van Der Gulden H, Gielkens A, Berns A, Moormann R. 1991. Live attenuated pseudorabies virus expressing envelope glycoprotein E1 of hog cholera virus protects swine against both pseudorabies and hog cholera. J Virol 65:2761–2765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Xu YG, Cui LC, Tian CY, Zhang GC, Huo GC, Tang LJ, Li YJ. 2011. Immunogenicity of recombinant classic swine fever virus CD8+ T lymphocyte epitope and porcine parvovirus VP2 antigen coexpressed by Lactobacillus casei in swine via oral vaccination. Clin Vaccine Immunol 18:1979–1986. doi: 10.1128/CVI.05204-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Chen H. 2000. Recent advances in mucosal vaccine development. J Control Release 67:117–128. doi: 10.1016/S0168-3659(00)00199-1. [DOI] [PubMed] [Google Scholar]
- 22.McGhee JR, Mestecky J, Dertzbaugh MT, Eldridge JK, Hirasawa M, Kiyono H. 1992. The mucosal immune system: from fundamental concepts to vaccine development. Vaccine 10:75–88. doi: 10.1016/0264-410X(92)90021-B. [DOI] [PubMed] [Google Scholar]
- 23.Medina E, Guzman CA. 2001. Use of live bacterial vaccine vectors for antigen delivery: potential and limitation. Vaccine 19:1573–1580. doi: 10.1016/S0264-410X(00)00354-6. [DOI] [PubMed] [Google Scholar]
- 24.Hou XL, Yu LY, Liu J, Wang GH. 2007. Surface-displayed porcine epidemic diarrhea viral (PEDV) antigens on lactic acid bacteria. Vaccine 26:24–31. doi: 10.1016/j.vaccine.2007.10.065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Qiao X, Li G, Wang X, Li X, Liu M, Li Y. 2009. Recombinant porcine rotavirus VP4 and VP4-LTB expressed in Lactobacillus casei induced mucosal and systemic antibody responses in mice. BMC Microbiol 9:249. doi: 10.1186/1471-2180-9-249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Villena J, Salva S, Agüero G, Alvarez S. 2011. Immunomodulatory and protective effect of probiotic Lactobacillus casei against Candida albicans infection in malnourished mice. Microbiol Immunol 55:434–445. doi: 10.1111/j.1348-0421.2011.00334.x. [DOI] [PubMed] [Google Scholar]
- 27.Wei CH, Liu JK, Hou XL, Yu LY, Lee JS, Kim CJ. 2010. Immunogenicity and protective efficacy of orally or intranasally administered recombinant Lactobacillus casei expressing ETEC K99. Vaccine 28:4113–4118. doi: 10.1016/j.vaccine.2009.05.088. [DOI] [PubMed] [Google Scholar]
- 28.Xu YG, Li YJ. 2007. Induction of immune responses in mice after intragastric administration of Lactobacillus casei producing porcine parvovirus VP2 protein. Appl Environ Microbiol 73:7041–7047. doi: 10.1128/AEM.00436-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zhao LL, Liu M, Ge JW, Qiao XY, Li YJ, Liu DQ. 2012. Expression of infectious pancreatic necrosis virus (IPNV) VP2-VP3 fusion protein in Lactobacillus casei and immunogenicity in rainbow trouts. Vaccine 30:1823–1829. doi: 10.1016/j.vaccine.2011.12.132. [DOI] [PubMed] [Google Scholar]
- 30.Alander M, Satokari R, Korpela R, Saxelin M, Vilpponen-Salmela T, Mattila-Sandholm T. 1999. Persistence of colonization of human colonic mucosa by a probiotic strain, Lactobacillus rhamnosus GG, after oral consumption. Appl Environ Microbiol 65:351–354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lee YK, Ho PS, Low CS, Arvilommi H, Salminen S. 2004. Permanent colonization by Lactobacillus casei is hindered by the low rate of cell division in mouse gut. Appl Environ Microbiol 70:670–674. doi: 10.1128/AEM.70.2.670-674.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ogawa T, Asai Y, Yasuda K, Sakamoto H. 2005. Oral immunoadjuvant activity of a new symbiotic Lactobacillus casei subsp casei in conjunction with dextran in BALB/c mice. Nutr Res 25:295–304. doi: 10.1016/j.nutres.2004.10.012. [DOI] [Google Scholar]
- 33.Perdigon G, Fuller R, Raya R. 2001. Lactic acid bacteria and their effect on the immune system. Curr Issues Intest Microbiol 1:27–42. [PubMed] [Google Scholar]
- 34.Jiang YF, Ma ZH, Zhao PW, Pan Y, Liu YY, Feng JY, Niu JQ. 2010. Effect of thymosin-α1 on T-helper 1 cell and T-helper 2 cell cytokine synthesis in patients with hepatitis B virus e antigen positive chronic hepatitis B. J Int Med Res 38:2053–2062. doi: 10.1177/147323001003800620. [DOI] [PubMed] [Google Scholar]
- 35.Li CL, Zhang T, Saibara T, Nemoto Y, Ono M, Akisawa N, Iwasaki S, Maeda T, Onishi S. 2002. Thymosin alpha1 accelerates restoration of T cell-mediated neutralizing antibody response in immunocompromised hosts. Int Immunopharmacol 2:39–46. doi: 10.1016/S1567-5769(01)00136-9. [DOI] [PubMed] [Google Scholar]
- 36.Knoetig SM, Summerfield A, Spagnuolo-Weaver M, McCullough KC. 1999. Immuno-pathogenesis of classical swine fever: role of monocytic cells. Immunology 97:359–366. doi: 10.1046/j.1365-2567.1999.00775.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.O'Mahony S, Barton JR, Crichton S, Ferguson A. 1990. Appraisal of gut lavage in the study of intestinal humoral immunity. Gut 31:1341–1344. doi: 10.1136/gut.31.12.1341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Belyakov IM, Hel Z, Kelsall B, Kuznetsov VA, Ahlers JD, Nacsa J, Watkins DI, Allen TM, Sette A, Altman J, Woodward R, Markham PD, Clements JD, Franchini G, Strober W, Berzofsky JA. 2001. Mucosal AIDS vaccine reduces disease and viral load in gut reservoir and blood after mucosal infection of macaques. Nat Med 7:1320–1326. doi: 10.1038/nm1201-1320. [DOI] [PubMed] [Google Scholar]
- 39.Summerfield A, Rziha HJ, Saalmüller A. 1996. Functional characterization of porcine CD4+CD8+ extrathymic T lymphocytes. Cell Immunol 168:291–296. doi: 10.1006/cimm.1996.0078. [DOI] [PubMed] [Google Scholar]
- 40.Brandtzaeg P. 1994. Distribution and characteristics of mucosal immunoglobulin producing cells, p 251–279. In Strober W, Mestecky J, Lamm ME, Bienenstock J, Ogra PL (ed), Handbook of mucosal immunology. Academic Press, San Diego, CA. [Google Scholar]