

Abstract
It is believed that tumor development, outgrowth and metastasis are under the surveillance of the immune system. Although both innate and acquired immune systems play roles, innate immunity is the spearhead against tumors. Recent studies have revealed the critical role of natural killer (NK) cells in immune surveillance and that NK cell activity is considerably influenced by various agents, such as environmental factors, stress, foods and drugs. Some of these NK cell stimulants have been used in complementary and alternative medicine (CAM) since ancient times. Therefore, the value of CAM should be re-evaluated from this point of view. In this review, we overview the intimate correlation between NK cell functions and CAM agents, and discuss possible underlying mechanisms mediating this. In particular, neuro-immune crosstalk and receptors for CAM agents are the most important and interesting candidates for such mechanisms.
Keywords: β-glucan, lectin, NK cell receptor, nerve-immune crosstalk, tumor
Introduction
It has been well known since ancient times that complementary and alternative medicine (CAM), including exercise, provides a lot of benefit to health. Many CAM modalities are believed to prevent or even cure diseases, especially morbid ones such as cancer. However, until recently conventional medicine has largely rejected the use of CAM agents because little biological evidence has been provided for the functional mechanisms of many of them. For the past few decades, the value of CAM has been rediscovered by many modern scientific researchers. Modulation of immune functions by CAM agents is the mechanism most widely analyzed and has been suggested to provide some scientific evidence for the biological effects of various CAM agents. Specifically, evidence for up-regulation of natural killer (NK) cell numbers and/or NK cell cytotoxicity by these agents has been accumulating. NK cells have been well established as innate cytotoxic effector cells for self defense in both vertebrates and invertebrates, in both of which they may use similar mechanisms in the lysis of target cells (1). In this review we overview NK cell functions and their functional molecules, particularly in relation to cancer, and discuss the possible mechanisms of NK cell activation by various CAM agents.
Why are NK Cells Important?
Ever since Lewis Thomas and Macfarlane Burnet proposed the immune surveillance hypothesis against tumor development, the concept has been a hot focus of debate for more than 40 years (2,3). The original immune surveillance hypothesis was challenged because nude mice lacking T cells did not show a higher incidence of cancer than did syngenic immunocompetent mice (4). However, this contradictory conclusion is now taken as fine evidence that innate immunity is in the critical arms of immune surveillance against tumor development. Moreover, in the innate immune system NK cells, which do not express T-cell receptors that recognize specific peptides presented on the major histocompatibility complex (MHC), rather than T cells, seem well suited for this role. NK cells thus mediating natural cytotoxicity are composed predominantly of large granular lymphocytes (LGL) and some of small agranular lymphocytes, both of which express CD16 and CD56, but no T-cell receptor, on the cell surface (5,6). NK cells can induce cytolysis in the absence of MHC class I antigen expression on their target cells (5). This phenomenon is commonly understood according to the ‘missing self’ hypothesis (7). Culture of NK cells with some cytokines augments their cytotoxic activity so that they become able to induce cytolysis for a wide spectrum of cells, including tumor cells expressing autologous MHC class I. They are now called lymphokine-activated killer (LAK) cells, although LAK cells are also induced by T lymphocytes (8–10).
NK Cells and Cancer
Tumor Immunosurveillance by NK Cells in Animal Models
Animals with low levels of NK cell activity (beige, cyclophosphamide or anti-asialo GM1 antibody-treated mice) have been shown to develop an increased number of spontaneous and experimental tumors and their metastases (11–13). Conversely, animals with augmented NK cell activity (treated with corynebacterium parvum or IFN-inducer) display increased resistance to the development of metastasis (14). Adoptive transfer of splenocytes with high NK cell activity partially restored the anti-metastatic ability of anti-asialo GM1 and cyclophosphamide-treated animals (12,15). Direct evidence for the role of LGL in the prevention of blood-born metastases was demonstrated by the inhibition of experimental metastases in NK cell-deficient mice following adoptive transfer of LGL (16,17).
Tumor Immunosurveillance by NK Cells in Humans
Despite relevant evidence from animal studies, the role of NK cells in immunologic surveillance against cancer in humans is poorly defined. Circumstantial evidence for the crucial role of natural cytotoxicity in human cancer was obtained by studying patients with NK deficient states. Patients who have Chëdiak–Higashi syndrome (18), X-linked lymphoproliferative syndrome (19) or who have had renal transplants (20) all have reduced NK cell activity and an increased incidence of cancer. Various studies have shown peripheral blood NK cell activity to be significantly reduced in patients with cancer compared with non-cancer-bearing control subjects (21–23). Other researchers, however, have documented no reduction in NK cell activity in patients with some cancers, irrespective of the stage of the disease (24,25). Thus, the precise role of NK cell cytotoxicity in human cancer remains inconclusive. Overall, however, there appears to be a trend for NK cell activity to be reduced in certain tumors, possibly related to tumor volume or dissemination. Some studies reported an increased risk of developing cancer in individuals with low NK cell activity, and normal NK cell activity only in the early stage of cancer (21,26–28). Several groups have reported increased blood-born metastasis, significantly increased risk of death from uncontrolled regional and distant metastasis, the development of regional and distant metastasis and risk of local recurrence in patients with low NK cell activity (29–33). Moreover, one epidemiological study recently indicated that low NK cell activity is associated with increased cancer risk, although it is premature to conclude from this result that NK cell activation induces tumor prevention (34). However, even this report can not be taken as clear evidence for NK cell-dependent tumor surveillance in humans, since expression of MHC class I, the most critical molecules for susceptibility to NK cell activity, on developing tumors was not analyzed.
We suggest that natural cytotoxicity mediated by NK cells may have a role in the prevention of the development of cancer and in the establishment of metastasis in humans as well as mice, although more evidence at the molecular level needs to be accumulated to define the role of NK cells in tumor surveillance in humans. Moreover, further epidemiological studies are needed to more precisely define whether NK cell activity can predict further cancer development in patients at high risk, or if NK cell activity has prognostic value in cancer patients. An intriguing possibility is prevention and/or cure of cancer by augmenting NK cells by CAM agents.
Cytotoxic Molecules of NK Cells and Tumors
Several studies have demonstrated an increased tumor incidence in mice deficient in cytotoxic molecules. A higher incidence of tumors has been noted in mice deficient in perforin (35,36), which is the critical cytotoxic molecule of NK and cytotoxic T cells (37), and tumor necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL) (38,39). A higher spontaneous and methylcholanthrene (MCA)-induced tumor formation were also reported in STAT-1 [a signal molecule require for responsiveness to interferons (IFNs)], IFN-γ-, and IFN-γ receptor-deficient mice (40–43). These show that impairment of the receptor for cytokines activating NK cells results in defective immune surveillance against tumors. Although these studies provide clear evidence for immune surveillance, there have been few reports of exactly how the tumor cells are recognized as abnormal and then eliminated. Recent studies on NK cell receptors have indicated the possible receptors responsible for tumor recognition and thus provide new support for the hypothesis of NK cell-mediated tumor surveillance (Fig. 1).
Figure 1.

NK cells in tumor surveillance. NK cells are activated by some cytokines and direct recognition of ligands of their NK cell activating receptors, which result in direct tumor cell lysis through their cytotoxic molecules (peforin/granzyme, Fas ligand, TRAIL) and activation of other immune cells (macrophages and T cells) through cytokine production. IFN-γ produced by NK cells also plays a critical role in the inhibition of angiogenesis by tumor development.
NK Cell Receptors for Tumor Recognition
The first NK cell receptors clearly defined were shown to bind both classical and non-classical MHC class I molecules and block the killing of target cells by NK cells (44). This inhibition was associated with immunoreceptor tyrosine-based inhibitory motif (ITIM) domains in the cytoplasmic tails of these NK receptors. ITIMs provide docking sites for phosphatases that oppose the activity of tyrosine kinases, which are essential enzymes for NK cell activation. However, certain members of the NK receptor family, KIR2DS, Ly49D/H and CD94/NKG2C, do not possess ITIMs. Instead, they are associated with the adapter molecule DAP12, which has an immunoreceptor tyrosine-based activating motif (ITAM) domain capable of activating NK cells (44). Thus, the view is now emerging that the activity of NK cells is regulated by the balance between activating and inhibitory signals. However, other than the down regulation of MHC class I molecules, it has been unclear which other molecules are likely to tip the balance in favor of NK cell activation for tumor surveillance (Table 1).
Table 1.
Summary of known NK cell receptors
| Species | Receptor | Ligands |
|---|---|---|
| Mice | ||
| Inhibitory | Ly49 | H-2K, H-2D |
| CD94/NKG2A | Qa-1b | |
| Activating | CD16 | IgG |
| NKp46 | Influenza haemagglutin, others? | |
| NKR-P1C | ? | |
| CD94/NKG2C | Qa-1b | |
| Ly49D | H-2Dd | |
| Ly49H | MCMV-induced? | |
| NKG2D | RAE1, H60, Mult1 | |
| CD244 | CD48 | |
| Human | ||
| Inhibitory | KIR2DL | |
| KIR3DL | HLA-C | |
| CD94/NKG2A(CD159a) | HLA-Bw4, HLA-A | |
| Activation | CD85j, CD85d | HLA-E |
| CD16 | HLA ClassI | |
| NKp30 | IgG | |
| NKp46 | ? | |
| KIR2DS | Influenza hemagglutin, others? | |
| KIR2DL | HLA-C, others | |
| CD94/NKG2C | HL-G | |
| NKp44 | HLA-E | |
| NKG2D | Influenza hemagglutin, others? | |
| CD244 | MICA/MICB, ULBP | |
| CD48 | ||
HLA, human leuocyte antigen; KIR, killer cell immunoglobulin-like receptors; MCMV, mouse cytomrgalovirus; MIC, MHC class I-chain related molecules; RAE-1, retinoic acid early inducible 1; ULBP, UL-16-binding protein.
The Role of NKG2D in Tumor Surveillance
NKG2D, encoded within the NK receptor gene complex, might be the activating receptor of NK cells critical for immune surveillance against tumors, since some NKG2D ligands (MICA and MICB in human, Rae1 and H60 in mice) can be induced by environmental stress and are expressed on many tumor cells (45–47). Moreover, not only NK cells but T cells expressing the γδ or αβ T cell receptor (TCR) express NKG2D, thus NKG2D turns out to be the missing link and unites innate and adoptive immunity in immune surveillance and anti-tumor immunity. Presumably, tumor cells, once they express NKG2D ligands, become able to induce immunological tolerance against attack from NK cells, as they are highly enhanced inhibitory ligands to be expressed.
CD94/NKG2A and CD94/NKG2C for Surveillance of ‘Modified Self’
CD94/NKG2 is the other crucial receptor for NK cell-mediated immune surveillance (44,48,49). HLA-E (Qa-1b in mice) is the relatively non-polymorphic ligand of inhibitory CD94/NKG2A and activating CD94/NKG2C. It has a preference for nonameric peptide, derived from the signal sequence of other class I molecules. Thus, CD94/NKG2A plays a critical role in blocking NK cell cytotoxicity by classical and non-classical MHC class I molecules. It might also be the receptor most likely to be responsible for the phenomenon explained by the ‘missing self’ hypothesis: namely, loss of MHC class I expression leads to sensitivity to lysis by NK cells (7). It was recently reported that a peptide from the signal sequence of stress protein hsp60 is loaded onto HLA-E, competing effectively with an MHC class I-derived peptide, and up-regulates surface expression of HLA-E (50). HLA-E loaded with the hsp-derived peptide does not bind to CD94/NKG2A. Therefore, stress-induced modification of HLA-E would activate CD94/NKG2A-expressing NK cells due to loss of inhibitory ligands, which would result in the augmentation of sensitivity to NK cells on the part of cells expressing hsp60. ‘Modified self’ (ligand modification of inhibitory receptor: CD94/NKG2A,HLA-E) is another mechanism by which susceptibility of tumor cells to NK cells is augmented, in addition to the ‘missing self (reduced class I expression)’ and ‘induced self’ (expression of NKG2D ligands) mechanisms (51).
NK Cells and CAM
Activation of NK Cells by CAM Agents
For the past few decades, scientific investigations on CAM have remarkably advanced and partly supported their medical efficacy through preclinical and clinical experiments. Such CAM modalities include: Oriental medicine, especially traditional Chinese medicine (including Kampo and acupuncture), extract products from natural plants, animal molecules and live lactic acid bacteria. In particular, many investigators have suggested that NK cell activation is one of the critical mechanisms for the biological effects induced by various CAM agents. For example, intake of green tea and some kind of live lactic acid bacteria enhanced NK cell activity (52–54). Administration of extracts from ginseng (52), aged garlic (55), Viscum album (mistletoe) (56,57), Cichorium intybus (58), Echinacea purpurea root (59), Derris scandens hydroalcholic (60), some wild plants (61), Chinese herbs (62) and some kinds of mushrooms (63) significantly augmented NK cytotoxicity or restored NK cell activity in some immune-suppressive conditions. Some of these agents showed inhibition of experimental metastasis of cancer. Oral administration of Phyllanthus emblica, which is known as an excellent source of vitamin C, enhanced NK cell activity and antibody-dependent cellular cytotoxicity (ADCC) (64), thus, supplemental nutriments might enhance NK cell activity. Moreover, acupuncture (65,66), skin rubdown (67), relaxation (68,69), message therapy (70), music therapy (71), mirthful laughter (72,73) and hypnotherapy (74) enhanced NK cell activity and/or NK cell numbers. There are a considerable numbers of studies reporting that acute and chronic exercise and long-term repeated exercise (training effects) elevated NK cell activity in peripheral blood (75–79) (Table 2).
Table 2.
Summary of NK cell activation-inducing CAM
| CAM | Immunological effects |
|---|---|
| Green tea | Humoral and cell-mediated immunity including NK cell activation |
| Ginseng | Increase in macrophage, NK, T and B cells number |
| Vitamin supplementation | Increase in antibody titer |
| Honey | Increase in antibody titer |
| Lactobacillus casei strain Shirota | Increased NK cell activity |
| Extract | |
| Aged garlic | Prevent a reduction of NK cell activity by psychological stress; prevent the decrease of spleen weight by psychological stress |
| Viscum album (mistletoe) | Increased NK cell number and NK cell activity |
| Cichorium intybus | Increase in circulating leukocytes; increase in phagocytic activity, NK cell activity, proliferation and IFN-α production |
| Echinaces purpurae root | Increased NK cell activity |
| Derris scandens | Increased NK cell activity in HIV-infected individuals |
| Nigella sativum seeds | Increased NK cell activity |
| Allium sativum bulb | Increased NK cell activity |
| Onopordum acanthium stem and leaves | Increased NK cell activity |
| Allium cepa bulbs | Increased NK cell activity |
| Chinese herb (e.g. Shikaron) | Increased NK cell activity |
| Phyllanthus emblica | Increased NK cell activity and antibody dependent cellular cytotoxicity (ADCC) |
| Mushrooms (e.g. Lentinus edodes and Agaricus blazei) | Increased NK cell activity |
| Physiological | |
| Acupuncture | Increase in T cell and NK cell numbers; increase in monocyte phagocytosis |
| Electroacupancture | Increased NK cell activity and IFN-α and IL-2 production |
| Skin rubdown | Increased NK cell activity |
| Exercise (dependent on time and frequency) | Increased NK cell activity and NK cell numbers |
| Psychological | |
| Relaxation | Increased NK cell activity and T cell response; increase in the number of peripheral blood lymphocytes |
| Message therapy | Increase in NK cell number in HIV-infected individuals |
| Music therapy | Increase in NK cell number and NK cell activity |
| Mirthful laughter | Prevents a reduction of NK cell activity by psychological stress; increased NK cell activity; increase in NK cells, T cells, immunoglobulins and IFN-α production |
Thus, numerous studies have reported the augmentation of NK cell number and/or NK cell cytotoxicity by CAM agents as described above. In order to examine this important issue further, however, criteria for the quality of evidence need to be established. It would not mean much, for example, if a report only showed that a certain CAM agent increased the number of NK cells in mice or humans.
A considerable number of reports have presented clinical observations. However, large-scale epidemiological studies to establish intimate correlations of any single CAM agent with NK cells in any specific disease conditions (e.g. cancer) are still lacking. Future epidemiological studies should be based on sound biological experiments both in vitro and in vivo. In selecting parameters for prospective epidemiological studies, we would like to draw attention to the next point below, and also the problem of individual differences between possible ‘CAM-agent receptors’ as shown in the following sections. Without such biological considerations, epidemiological analysis will be pointless.
It is now well known that NK cell activity is regulated by expression levels of cytotoxic molecules, activating receptors and/or inhibitory receptors. Thus, decrease or increase in NK cell number in peripheral blood mononuclear cell (PBMC) does not necessarily result in a decrease or increase in NK cell activity, respectively. Therefore, in designing any further experiments in this field, NK cell numbers and NK cell cytotoxicity should be analyzed simultaneously, and responsible molecules or mechanisms for CAM effects should be clearly defined.
Cytotoxic and activation mechanisms of NK cells have been demonstrated in experimental models using various gene-modified mice (transgenic and gene-knock-out mice). Therefore, we recommend researchers to design experiments using such laboratory mice. It should become the standard preclinical system in which mechanisms of the effects of any CAM agents in the activation of NK cells are to be clearly demonstrated in vivo.
Example: NK Cell Activation by Lactobacillus casei Strain Shirota and Extract of Mushrooms
We have analyzed the effect on immunological function of a fermented milk drink containing Lactobacillus casei strain Shirota (LcS) (53). The frequency and the number of NK cells, CD4+ T cells or CD8+ T cells were not significantly changed, and T cell responsiveness to mitogens (Con A and PHA) and allantigen stimulation were unchanged. Moreover, significant increase of IFN-α and IFN-γ could not be detected in serum at all. However, NK cell cytotoxicity was significantly increased, and the enhancement of NK cell activity was particularly prominent in the low NK activity individuals (Fig. 2). We have also analyzed NK cell activity after the intake of the extract of mushrooms, Agaricus blazei and Lentinus edodes, which are commonly used CAM agents in Japan. Although intake of these extracts did not induce significant change in lymphocyte populations and T cell responses, NK cell activity was augmented. Notably, susceptibility to extract of Agaricus blazei was varied among individuals; however, it was closely correlated to the responsiveness to the extract of Lentinus edodes but not LcS. This suggests that these individuals are selectively susceptible to some general components of the mushroom extracts (Table 3). Conversely, NK cell activity was highly augmented by LcS but not the mushroom extracts in some volunteers, and only slight augmentation of NK cell activity was observed by either LcS or mushrooms extract in some others, even if their NK cell activity was relatively low.
Figure 2.
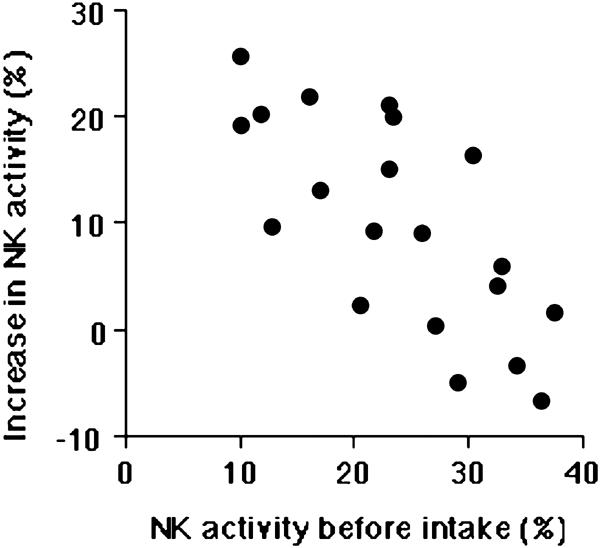
Intimate correlation between the magnitude of the increase in NK cell activity and the level of NK cell activity. The increase in NK cell activity after the intake of LcS drink is prominent in individuals with low NK cell activity. The correlation was determined by Pearson's methods; statistical significance was P < 0.05.
Table 3.
Intimate correlation of susceptibility to extracts of mushrooms
| Agaricus blazei* | Lentinus edodes* | Lactobacillus casei strain Shirota | Population | |
|---|---|---|---|---|
| Group A | ++ | ++ | various | 25% |
| Group B | + | + | various | 50% |
| Group C | − | − | various | 25% |
| Group D | ++ or + | − | 0% | |
| Group E | − | ++ or + | 0% |
* ++, 20% or more increase of NK cell cytotoxicity at E/T = 20; +, around 10% increase in NK cell cytotoxicity at E/T = 20.
It has often been suggested that effects of some CAM modalities are varied among individuals. Our examples presented above suggest that the individual differences in the responsiveness to CAM agents may be due to the individual difference of expression of various CAM receptors. If it really is the case, it supports the concept that proper selection of CAM agents for each individual is needed in order for their effects to be manifested.
How do CAM Agents Activate NK Cells?
The precise mechanisms of NK cell activation by CAM agents have not been clearly defined. The two main possibilities are: (i) augmentation of cytotoxic molecules in NK cells and/or (ii) up-regulation of activating NK cell receptors and/or down-regulation of inhibitory NK cell receptors. On the other hand, improvement of general ‘health’ conditions by some CAM agents, e.g. exercise, may result in NK cell activation, although precise pathways are still unclear. We shall discuss in this section two interesting mechanisms, one indirect and the other direct, which seem to be reasonable explanations for the NK cell activation induced by CAM agents.
Neuro-immune Crosstalk
A growing body of evidence shows a crucial role of the autonomic nervous system in modulating the immune response in stress (75). For example, the autonomic nervous system might play an important role in the modulation of the immune response after both acute and chronic exercise.
Intimate crosstalk between the central nerve systems and the immune systems has been demonstrated by experiments using electrical stimulation of the ventromedial horn of the hypothalamus, which is the site in the central nervous system responsible for delivering sympathetic stimulus to spleen (80,81). There is also evidence to show a crucial role of the autonomic nervous system in modulation of the immune response, especially under the influence of stressors and exercise (82,83), although little is known about the role of parasympathetic nerves compared to the large amount of information available on sympathetic nerves in immune regulation.
Modulation of the immune response can be mediated either through innervation by sympathetic nerves or through the receptors for their chemical transmitters on immune cells in these organs, as described below.
Regulation of Blood Flow by the Sympathetic Nervous System
An important role of autonomic nerves in both primary and secondary lymphoid organs was reported for the regulation of blood flow (81,84,85), which plays a major role in immune regulation by facilitating the right cells to lodge or move to where they can efficiently accomplish their task (86,87).
Direct Effects of the Sympathetic Nervous System on Immune Cells
A growing number of studies suggests that sympathetic nerves can influence immune cells more directly than just by regulating the blood flow. Catecholamines released from sympathetic terminals are major mediators responsible for immune regulation when they bind to their specific receptors (88). Their suppressive nature on NK cell cytotoxicity has been reported (89,90). In striking contrast, acute deprivation of sympathetic innervation causes enhancement, while its chronic deprivation seems to cause suppression, of NK cell activity, and diminished cell-mediated immunity (91,92).
Impact of Sympathetic Modulation on the Distribution of Leukocytes
An active involvement of autonomic regulation in leukocyte distribution is suggested by the following fact. The increase in circulating NK cells and decreased NK cell attachment to endothelial cells induced by catecholamines but independent of adhesion-molecules, was inhibited by β2-adrenergic antagonist (93–95). NK cells have higher expression of β-adrenergic receptors and lower expression of cholinergic receptors, and are thereby influenced by sympathetic and parasympathetic activities (96). Mental stress accompanied by sympathetic activation increases the number of NK cells in circulation, which is blocked by β-adrenergic antagonist (93,97,98). In the same study it was demonstrated that spleen does not serve as a pool for NK cells (93). On the other hand, an increased accumulation of catecholamine-stimulated lymphocytes to secondary lymphoid organs might account for the transient decrease in lymphocytes from circulation after catecholamine injection (99). Thus, the origin of NK cells released into circulation after β-adrenergic stimulation is an open question yet to be answered.
Taken together, the sympathetic nervous system does not simply suppress the immune system but might help in organizing the immune response sequentially and spatially by modulating the distribution of immune cells. Modification of blood flow and cell adhesion must be the crucial event for increasing NK cells in peripheral blood by CAM agents, since increase of NK cell numbers does not always link to augmentation of NK cell cytotoxicity (Fig. 3).
Figure 3.

Summary of immune system control by autonomic nervous system. The central nervous and immune systems intimately crosstalk in the live body. In particular, sympathetic nerves mediate inhibitory effects on NK cells. NK cell distribution and NK cell activity are controlled by autonomic nervous systems directly and indirectly.
Receptors for CAM Agents on NK Cells?
Control of the immune functions mediated by the nervous system would be the major mechanism underlying NK cell activation by physical CAM modalities (e.g. exercise, skin rubdown and acupuncture) and psychological CAM ones (e.g. relaxation, message therapy, music therapy and mirthful laughter). On the other hand, numerous CAM agents have also been reported to activate NK cell cytotoxity, improve general health and reduce the risk of cancer development. This type of phenomena, if real, should be understood as reasonable if NK cells and/or some innate immune cells expressed receptors for some components of various types of CAM agents (e.g. liquid and food). Toll-like receptors and receptors to lectins and β-glucan polysaccharides are the most likely candidates responsible for NK cell activation induced by intake-type CAM agents (100). These CAM-agent receptors may well be preferentially expressed on macrophages or dendritic cells. In such cases, these CAM-agent-responding cells produce cytokines (e.g. type-I IFN, IL-12 and IL-18), which induce NK cell activation.
Toll-like Receptors Involved?
Toll-like receptors (TLRs) function as the pattern-recognition receptors in mammals and play an essential role in their recognition of microbial and fungal components (101–104). Ten members of the TLR family have been identified in humans. TLRs activate NK-κB and other signaling pathways, which results in the secretion of various inflammatory cytokines (101–105). It has been reported that TLR-2/6 recognize some components of zymosan, but not β-glucan, which result in production of cytokines and chemokines (106,107). Thus, TLRs might play important roles in the biological effects of some intake-type CAM agents. This is a very interesting subject for further studies.
Receptors for Lectin Involved?
It has been established that various members of the lectin family modify immunological functions (108,109). Wild plants and fungi have traditionally been the single largest source of lead lectin compounds for the development of therapeutics agents by the pharmaceutical industry. Currently, mushroom and plant polysaccharides are the focus of attention in relation to CAM, stimulating scientific analysis and drug development to prevent and treat cancer. Thus, some unknown C-type lectin-like receptors for certain components of CAM agents and/or lectin, and lectin interaction, might play roles in CAM effects.
Receptors for β-Glucan Involved?
The in vivo administration of β-glucan has been shown to potentiate host responses against a variety of conditions, including tumor development and infection (100,110,111). This has led to a number of clinical trials using β-glucans for tumor immunotherapy with some promising results. Thus, β-glucan receptors are one likely class of receptors responsible for NK cell activation by CAM agents.
The β-glucans are a heterogeneous group of glucose polymers, consisting of a backbone of β(1→3)-linked β-D-glucopyranosyl units with β(1→6)-linked side chains of varying distribution and length. These polysaccharides are major cell wall structural components in fungi, mushrooms, plants and some bacteria. As they are not found in animals, these carbohydrates are considered to be classic pattern-recognition molecules (101) and are recognized by the innate immune system. Vertebrate recognition of β-glucans appears to occur exclusively via several cell surface receptors, and although complement opsonization does contribute to the recognition of particulate glucans, no plasma molecules recognizing this carbohydrate structure have been identified. β-Glucan recognition systems in invertebrates are completely different from vertebrates (112); however, the recognition of β-glucan by both systems results in the triggering of innate immunity (Table 4).
Table 4.
Summary of immunostimulating polysaccharides of higher basidomycetes
| Polysaccharide | Species |
|---|---|
| Glucans | |
| α-(1→6)-; α-(1→4)-glucan | Agaricus blazei |
| α-(1→4)-; β-(1→6)-glucan | Agaricus blazei |
| β-(1→6)-; α-(1→3)-glucan | Agaricus blazei |
| β-(1→6)-; β-(1→3)-glucan | Agaricus blazei |
| Galactomannoglucan | Ganoderma tsugae |
| Galactoxyloglucan | Hericium erninaceus |
| Glucan phosphate | Saccharomyces cervisiae |
| Grifolan | Grifola frondosa (Maitake mushroom) |
| Lentinan | Lentinus edodes (Shiitake mushroom) |
| Mannnogalactoglucan | Pleurotus pulmonarius |
| Agaricus blazei | |
| PGG-Glucan | Saccharomyces cervisiae |
| Riboglucan | Agaricus blazei |
| Schizophyllan | Schizophyllum commune |
| Scleroglucan | Sclerotium glucanicum |
| SSG-Glucan | Sclerotinia sclerotiorum |
| Xyloglucan | Grifola frondosa |
| Pleurotus pulmonarius | |
| Zymogen | Saccharomyces cervisiae |
| Glycan | |
| β-(1→2)-; β-(1→3)-glucomannan | Agaricus blazei |
| Galactoglucomannan | Lentinus edodes (Shiitake mushroom) |
| Glucogalactan | Ganoderma tsugae |
| Glucomannan | Agaricus blazei |
| Glucoxylan | Hericium erninaceus |
| Pleurotus pulmonarius | |
| Mannogalactan | Pleurotus pulmonarius |
| Mannogalactofucan | Grifola frondosa |
| Mannnoglucoxylan | Hericium erninaceus |
| Xylan | Hericium erninaceus |
β-Glucan receptor activity has subsequently been reported on a variety of leukocytes, including macrophages, neutrophils, eosinophils and NK cells, as well as on non-immune cells including endotherial cells, alveolar epithelial cells and fibroblasts. Non-opsonic recognition of β-glucan by these cells has been ascribed to multiple receptors (113), and indeed a number of β-glucan receptors have been identified, including CR3 (110), lactosylceramide (CDw17) (114), scavenger receptors (115) and dectin-1 (116–118). Of these receptors, however, dectin-1 has been clearly shown to play a role in mediating the biological response to β-glucan (119). Although the mechanisms are not yet fully understood, the generation of the response starts at the cell surface and requires a functional ITAM domain in cytoplasmic tails of dectin-1. The ITAM motif becomes phosphorylated after β-glucan binding, suggesting that signaling might occur in a similar fashion to that of other ITAM-containing receptors, such as NK-activating receptors and Fc receptors.
Not only β-glucan polysaccharides, but also oligosaccharides have been reported as the effector components of some CAM agents, which down-regulate production of immunosuppressive cytokines (100). It has been suggested that glycochain, glycopeptide and glycolipid play a critical role in numerous biological events, including immunological responses (120–125). However, glycobiology still cannot clearly reveal the molecular mechanisms for their biological functions. Further analysis in glycobiology will identify the crucial receptors for CAM on immune cells and elucidate the essential molecules and mechanisms. As this field is rapidly progressing, our perspective may be drastically changed in several years. It will be interesting for us to present a revised and expanded version of this review in a couple of years.
Acknowledgments
This work was supported by the Ministry of Education, Science, and Culture, Japan and Human Frontier Science Program.
References
- 1.Cooper EL, Kauscheke E, Cossarizza A. Digging for natural immunity since Darwin and Metchnikoff. BioEssays. 2002;24:319–333. doi: 10.1002/bies.10077. [DOI] [PubMed] [Google Scholar]
- 2.Burnet FM. Cancer: a biological approach. Br Med J. 1957;1:779–785. doi: 10.1136/bmj.1.5022.779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Burnet FM. Immunological aspects of malignant disease. Lancet. 1967;1:1171–1174. doi: 10.1016/s0140-6736(67)92837-1. [DOI] [PubMed] [Google Scholar]
- 4.Stutman O. Tumor development after 3-methylcholanthrene in immunologically deficient athymic-nude mice. Science. 1974;183:534–536. doi: 10.1126/science.183.4124.534. [DOI] [PubMed] [Google Scholar]
- 5.Trinchieri G. Biology of natural killer cells. Adv Immunol. 1989;47:187–376. doi: 10.1016/S0065-2776(08)60664-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ortaldo JR, Winkler-Pickett R, Kopp W, Kawasaki A, Nagashima K, Okumura K, et al. Relationship of large and small CD3- CD56+ lymphocytes mediating NK-associated activities. J Leukoc Biol. 1992;52:287–295. doi: 10.1002/jlb.52.3.287. [DOI] [PubMed] [Google Scholar]
- 7.Kärre K, Ljunggren HG, Piontek G, Kiessling R. selective rejection of H-2-deficient lymphoma variant suggests alternative immune defence strategy. Nature. 1986;319:675–678. doi: 10.1038/319675a0. [DOI] [PubMed] [Google Scholar]
- 8.Grimm EA, Mazumder A, Zhang HZ, Rosenberg SA. Lymphokine-activated killer cell phenomenon. Lysis of natural killer-resistant fresh solid tumor cells by interleukin 2-activated autologous human peripheral blood lymphocytes. J Exp Med. 1982;155:1823–1841. doi: 10.1084/jem.155.6.1823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Trinchieri G, Matsumoto-Kobayashi M, Clark SC, Seehra J, London L, Perussia B. Response of resting human peripheral blood natural killer cells to interleukin 2. J Exp Med. 1984;160:1147–1169. doi: 10.1084/jem.160.4.1147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Herberman RB, Hiserodt J, Vujanovic N, Balch C, Lotzova E, Bolhuis R, et al. Lymphokine-activated killer cell activity. Immunol Today. 1987;8:178–181. doi: 10.1016/0167-5699(87)90035-1. [DOI] [PubMed] [Google Scholar]
- 11.Talmadge JE, Meyers KM, Prieur DJ, Starkey JR. Role of NK cells in tumour growth and metastasis in beige mice. Nature. 1980;284:622–624. doi: 10.1038/284622a0. [DOI] [PubMed] [Google Scholar]
- 12.Gorelik E, Wiltrout RH, Okumura K, Habu S, Herberman RB. Role of NK cells in the control of metastatic spread and growth of tumor cells in mice. Int J Cancer. 1982;30:107–112. doi: 10.1002/ijc.2910300118. [DOI] [PubMed] [Google Scholar]
- 13.Okumura K, Habu S, Kasai M. The role of NK cells in resistance of in vivo tumors. Adv Exp Med Biol. 1982;155:773–784. doi: 10.1007/978-1-4684-4394-3_85. [DOI] [PubMed] [Google Scholar]
- 14.Hanna N. Inhibition of experimental tumor metastasis by selective activation of natural killer cells. Cancer Res. 1982;42:1337–1342. [PubMed] [Google Scholar]
- 15.Hanna N, Burton RC. Definitive evidence that natural killer (NK) cells inhibit experimental tumor metastases in vivo. J Immunol. 1981;127:1754–1758. [PubMed] [Google Scholar]
- 16.Barlozzari T, Leonhardt J, Wiltrout RH, Herberman RB, Reynolds CW. Direct evidence for the role of LGL in the inhibition of experimental tumor metastases. J Immunol. 1985;134:2783–2789. [PubMed] [Google Scholar]
- 17.Barlozzari T, Reynolds CW, Herberman RB. In vivo role of natural killer cells: involvement of large granular lymphocytes in the clearance of tumor cells in anti-asialo GM1-treated rats. J Immunol. 1983;131:1024–1027. [PubMed] [Google Scholar]
- 18.Roder JC, Haliotis T, Klein M, Korec S, Jett JR, Ortaldo J, et al. A new immunodeficiency disorder in humans involving NK cells. Nature. 1980;284:553–555. doi: 10.1038/284553a0. [DOI] [PubMed] [Google Scholar]
- 19.Sullivan JL, Byron KS, Brewster FE, Purtilo DT. Deficient natural killer cell activity in X-linked lymphoproliferative syndrome. Science. 1980;210:543–545. doi: 10.1126/science.6158759. [DOI] [PubMed] [Google Scholar]
- 20.Gui XE, Rinaldo CR, Jr, Ho M. Natural killer cell activity in renal transplant recipients receiving cyclosporine. Infect Immun. 1983;41:965–970. doi: 10.1128/iai.41.3.965-970.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hersey P, Edwards A, Honeyman M, McCarthy WH. Low natural-killer-cell activity in familial melanoma patients and their relatives. Br J Cancer. 1979;40:113–122. doi: 10.1038/bjc.1979.147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Sibbitt WL, Jr, Bankhurst AD, Jumonville AJ, Saiki J, Saiers JH, Doberneck RC. Defects in natural killer cell activity and interferon response in human lung carcinoma and malignant melanoma. Cancer Res. 1984;44:852–856. [PubMed] [Google Scholar]
- 23.Mickel RA, Kessler DJ, Taylor JM, Lichtenstein A. Natural killer cell cytotoxicity in the peripheral blood, cervical lymph nodes, and tumor of head and neck cancer patients. Cancer Res. 1988;48:5017–5022. [PubMed] [Google Scholar]
- 24.Pislarasu M, Oproiu A, Taranu D, Herberman RB, Sulica A. Modulation of natural killer cell activity by serum from cancer patients: preliminary results of a study of patients with colorectal adenocarcinoma or other types of cancer. Cancer Res. 1988;48:2596–2603. [PubMed] [Google Scholar]
- 25.Eremin O, Ashby J, Stephens JP. Human natural cytotoxicity in the blood and lymphoid organs of healthy donors and patients with malignant disease. Int J Cancer. 1978;21:35–41. doi: 10.1002/ijc.2910210108. [DOI] [PubMed] [Google Scholar]
- 26.Seltzer V, Doyle A, Kadish AS. Natural cytotoxicity in malignant and premalignant cervical neoplasia and enhancement of cytotoxicity with interferon. Gynecol Oncol. 1983;15:340–349. doi: 10.1016/0090-8258(83)90051-3. [DOI] [PubMed] [Google Scholar]
- 27.Strayer DR, Carter WA, Mayberry SD, Pequignot E, Brodsky I. Low natural cytotoxicity of peripheral blood mononuclear cells in individuals with high familial incidences of cancer. Cancer Res. 1984;44:370–374. [PubMed] [Google Scholar]
- 28.Nakajima T, Mizushima N, Kanai K. Relationship between natural killer activity and development of hepatocellular carcinoma in patients with cirrhosis of the liver. Jpn J Clin Oncol. 1987;17:327–332. [PubMed] [Google Scholar]
- 29.Schantz SP, Campbell BH, Guillamondegui OM. Pharyngeal carcinoma and natural killer cell activity. Am J Surg. 1986;152:467–474. doi: 10.1016/0002-9610(86)90326-0. [DOI] [PubMed] [Google Scholar]
- 30.Schantz SP, Savage HE, Racz T, Taylor DL, Sacks PG. Natural killer cells and metastases from pharyngeal carcinoma. Am J Surg. 1989;158:361–366. doi: 10.1016/0002-9610(89)90134-7. [DOI] [PubMed] [Google Scholar]
- 31.Schantz SP, Ordonez NG. Quantitation of natural killer cell function and risk of metastatic poorly differentiated head and neck cancer. Nat Immun Cell Growth Regul. 1991;10:278–288. [PubMed] [Google Scholar]
- 32.Pross HF, Lotzova E. Role of natural killer cells in cancer. Nat Immun. 1993;12:279–292. [PubMed] [Google Scholar]
- 33.Tartter PI, Steinberg B, Barron DM, Martinelli G. The prognostic significance of natural killer cytotoxicity in patients with colorectal cancer. Arch Surg. 1987;122:1264–1268. doi: 10.1001/archsurg.1987.01400230050009. [DOI] [PubMed] [Google Scholar]
- 34.Imai K, Matsuyama S, Miyake S, Suga K, Nakachi K. Natural cytotoxic activity of peripheral-blood lymphocytes and cancer incidence: an 11-year follow-up study of a general population. Lancet. 2000;356:1795–1799. doi: 10.1016/S0140-6736(00)03231-1. [DOI] [PubMed] [Google Scholar]
- 35.van den Broek ME, Kägi D, Ossendorp F, Toes R, Vamvakas S, Lutz WK, et al. Decreased tumor surveillance in perforin-deficient mice. J Exp Med. 1996;184:1781–1790. doi: 10.1084/jem.184.5.1781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Smyth MJ, Thia KY, Street SE, MacGregor D, Godfrey DI, Trapani JA. Perforin-mediated cytotoxicity is critical for surveillance of spontaneous lymphoma. J Exp Med. 2000;192:755–760. doi: 10.1084/jem.192.5.755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kägi D, Ledermann B, Bürki K, Seiler P, Odermatt B, Olsen KJ, et al. Cytotoxicity mediated by T cells and natural killer cells is greatly impaired in perforin-deficient mice. Nature. 1994;369:31–37. doi: 10.1038/369031a0. [DOI] [PubMed] [Google Scholar]
- 38.Takeda K, Smyth MJ, Cretney E, Hayakaw Y, Kayagaki N, Yagita H, et al. Critical role for tumor necrosis factor-related apoptosis-inducing ligand in immune surveillance against tumor development. J Exp Med. 2002;195:161–169. doi: 10.1084/jem.20011171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Cretney E, Takeda K, Yagita H, Glaccum M, Peschon JJ, Smyth MJ. Increased susceptibility to tumor initiation and metastasis in TNF-related apoptosis-inducing ligand-deficient mice. J Immunol. 2002;168:1356–1361. doi: 10.4049/jimmunol.168.3.1356. [DOI] [PubMed] [Google Scholar]
- 40.Shankaran V, Ikeda H, Bruce AT, White JM, Swanson PE, Old LJ, et al. IFNγ and lymphocytes prevent primary tumour development and shape tumour immunogenicity. Nature. 2001;410:1107–1111. doi: 10.1038/35074122. [DOI] [PubMed] [Google Scholar]
- 41.Kaplan DH, Shankaran V, Dighe AS, Stockert E, Aguet M, Old LJ, et al. Demonstration of an interferon γ-dependent tumor surveillance system in immunocompetent mice. Proc Natl Acad Sci U S A. 1998;95:7556–7561. doi: 10.1073/pnas.95.13.7556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Street SE, Cretney E, Smyth MJ. Perforin and interferon-γ activities independently control tumor initiation, growth, and metastasis. Blood. 2001;97:192–197. doi: 10.1182/blood.v97.1.192. [DOI] [PubMed] [Google Scholar]
- 43.Dighe AS, Richards E, Old LJ, Schreiber RD. Enhanced in vivo growth and resistance to rejection of tumor cells expressing dominant negative IFN γ receptors. Immunity. 1994;1:447–456. doi: 10.1016/1074-7613(94)90087-6. [DOI] [PubMed] [Google Scholar]
- 44.Lanier LL. NK cell receptors. Annu Rev Immunol. 1998;16:359–393. doi: 10.1146/annurev.immunol.16.1.359. [DOI] [PubMed] [Google Scholar]
- 45.Raulet DH. Roles of the NKG2D immunoreceptor and its ligands. Nat Rev Immunol. 2003;3:781–790. doi: 10.1038/nri1199. [DOI] [PubMed] [Google Scholar]
- 46.Lanier LL. A renaissance for the tumor immunosurveillance hypothesis. Nat Med. 2001;7:1178–1180. doi: 10.1038/nm1101-1178. [DOI] [PubMed] [Google Scholar]
- 47.Diefenbach A, Raulet DH. The innate immune response to tumors and its role in the induction of T-cell immunity. Immunol Rev. 2002;188:9–21. doi: 10.1034/j.1600-065x.2002.18802.x. [DOI] [PubMed] [Google Scholar]
- 48.Long EO. Regulation of immune responses through inhibitory receptors. Annu Rev Immunol. 1999;17:875–904. doi: 10.1146/annurev.immunol.17.1.875. [DOI] [PubMed] [Google Scholar]
- 49.Raulet DH, Vance RE, McMahon CW. Regulation of the natural killer cell receptor repertoire. Annu Rev Immunol. 2001;19:291–330. doi: 10.1146/annurev.immunol.19.1.291. [DOI] [PubMed] [Google Scholar]
- 50.Michaelsson J, Teixeira de Matos C, Achour A, Lanier LL, Kärre K, Soderstrom K. A signal peptide derived from hsp60 binds HLA-E and interferes with CD94/NKG2A recognition. J Exp Med. 2002;196:1403–1414. doi: 10.1084/jem.20020797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Long EO, Rajagopalan S. Stress signals activate natural killer cells. J Exp Med. 2002;196:1399–1402. doi: 10.1084/jem.20021747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Klein C, Sato T, Meguid MM, Miyata G. From food to nutritional support to specific nutraceuticals: a journey across time in the treatment of disease. J Gastroenterol. 2000;35(Suppl 12):1–6. [PubMed] [Google Scholar]
- 53.Nagao F, Nakayama M, Muto T, Okumura K. Effects of a fermented milk drink containing Lactobacillus casei strain Shirota on the immune system in healthy human subjects. Biosci Biotechnol Biochem. 2000;64:2706–2708. doi: 10.1271/bbb.64.2706. [DOI] [PubMed] [Google Scholar]
- 54.Ohashi Y, Nakai S, Tsukamoto T, Masumori N, Akaza H, Miyanag N, et al. Habitual intake of lactic acid bacteria and risk reduction of bladder cancer. Urol Int. 2002;68:273–280. doi: 10.1159/000058450. [DOI] [PubMed] [Google Scholar]
- 55.Kyo E, Uda N, Ushijima M, Kasuga S, Itakura Y. Prevention of psychological stress-induced immune suppression by aged garlic extract. Phytomedicine. 1999;6:325–330. doi: 10.1016/S0944-7113(99)80053-1. [DOI] [PubMed] [Google Scholar]
- 56.Schink M. Mistletoe therapy for human cancer: the role of the natural killer cells. Anticancer Drugs. 1997;8(Suppl 1):S47–S51. doi: 10.1097/00001813-199704001-00011. [DOI] [PubMed] [Google Scholar]
- 57.Yoon TJ, Yoo YC, Kang TB, Baek YJ, Huh CS, Song SK, et al. Prophylactic effect of Korean mistletoe (Viscum album coloratum) extract on tumor metastasis is mediated by enhancement of NK cell activity. Int J Immunopharmacol. 1998;20:163–172. doi: 10.1016/s0192-0561(98)00024-1. [DOI] [PubMed] [Google Scholar]
- 58.Kim JH, Mun YJ, Woo WH, Jeon KS, An NH, Park JS. Effects of the ethanol extract of Cichorium intybus on the immunotoxicity by ethanol in mice. Int J Immunopharmacol. 2002;2:733–744. doi: 10.1016/s1567-5769(02)00008-5. [DOI] [PubMed] [Google Scholar]
- 59.Currier NL, Miller SC. The effect of immunization with killed tumor cells, with/without feeding of Echinacea purpurea in an erythroleukemic mouse model. J Altern Complement Med. 2002;8:49–58. doi: 10.1089/107555302753507177. [DOI] [PubMed] [Google Scholar]
- 60.Sriwanthana B, Chavalittumrong P. In vitro effect of Derris scandens on normal lymphocyte proliferation and its activities on natural killer cells in normal and HIV-1 infected patients. J Ethnopharmacol. 2001;76:125–129. doi: 10.1016/s0378-8741(01)00223-9. [DOI] [PubMed] [Google Scholar]
- 61.Abuharfeil NM, Salim M, Von Kleist S. Augmentation of natural killer cell activity in vivo against tumour cells by some wild plants from Jordan. Phytother Res. 2001;15:109–113. doi: 10.1002/ptr.692. [DOI] [PubMed] [Google Scholar]
- 62.Kurashige S, Jin R, Akuzawa Y, Endo F. Anticarcinogenic effects of shikaron, a preparation of eight Chinese herbs in mice treated with a carcinogen, N-butyl-N′-butanolnitrosoamine. Cancer Invest. 1998;16:166–169. doi: 10.3109/07357909809050031. [DOI] [PubMed] [Google Scholar]
- 63.Fujimiya Y, Suzuki Y, Oshiman K, Kobori H, Moriguchi K, Nakashima H, et al. Selective tumoricidal effect of soluble proteoglucan extracted from the basidiomycete, Agaricus blazei Murill, mediated via natural killer cell activation and apoptosis. Cancer Immunol Immunother. 1998;46:147–159. doi: 10.1007/s002620050473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Suresh K, Vasudevan DM. Augmentation of murine natural killer cell and antibody dependent cellular cytotoxicity activities by Phyllanthus emblica, a new immunomodulator. J Ethnopharmacol. 1994;44:55–60. doi: 10.1016/0378-8741(94)90099-x. [DOI] [PubMed] [Google Scholar]
- 65.Yu Y, Kasahara T, Sato T, Asano K, Yu G, Fang J, et al. Role of endogenous interferon-g on the enhancement of splenic NK cell activity by electroacupuncture stimulation in mice. J Neuroimmunol. 1998;90:176–186. doi: 10.1016/s0165-5728(98)00143-x. [DOI] [PubMed] [Google Scholar]
- 66.Choi GS, Oha SD, Han JB, Bae HS, Cho YW, Yun YS, et al. Modulation of natural killer cell activity affected by electroacupuncture through lateral hypothalamic area in rats. Neurosci Lett. 2002;329:1–4. doi: 10.1016/s0304-3940(02)00551-7. [DOI] [PubMed] [Google Scholar]
- 67.Kiecolt-Glaser JK, Glaser R, Williger D, Stout J, Messick G, Sheppard S, et al. Psychosocial enhancement of immunocompetence in a geriatric population. Health Psychol. 1985;4:25–41. doi: 10.1037//0278-6133.4.1.25. [DOI] [PubMed] [Google Scholar]
- 68.Gruber BL, Hersh SP, Hall NR, Waletzky LR, Kunz JF, Carpenter JK, et al. Immunological responses of breast cancer patients to behavioral interventions. Biofeedback Self Regul. 1993;18:1–22. doi: 10.1007/BF00999510. [DOI] [PubMed] [Google Scholar]
- 69.Diego MA, Field T, Hernandez-Reif M, Shaw K, Friedman L, Ironson G, et al. HIV adolescents show improved immune function following massage therapy. Int J Neurosci. 2001;106:35–45. doi: 10.3109/00207450109149736. [DOI] [PubMed] [Google Scholar]
- 70.Bittman BB, Berk LS, Felten DL, Westengard J, Simonton OC, Pappas J, et al. Composite effects of group drumming music therapy on modulation of neuroendocrine-immune parameters in normal subjects. Altern Ther Health Med. 2001;7:38–47. [PubMed] [Google Scholar]
- 71.Berk LS, Felten DL, Tan SA, Bittman BB, Westengard J. Modulation of neuroimmune parameters during the eustress of humor-associated mirthful laughter. Altern Ther Health Med. 2001;7:62–72. 74–166. [PubMed] [Google Scholar]
- 72.Bennett MP, Zeller JM, Rosenberg L, McCann J. The effect of mirthful laughter on stress and natural killer cell activity. Altern Ther Health Med. 2003;9:38–45. [PubMed] [Google Scholar]
- 73.Iwama H, Akama Y. Skin rubdown with a dry towel activates natural killer cells in bedridden old patients. Med Sci Monit. 2002;8:CR611–CR615. [PubMed] [Google Scholar]
- 74.Fox PA, Henderson DC, Barton SE, Champion AJ, Rollin MS, Catalan J, et al. Immunological markers of frequently recurrent genital herpes simplex virus and their response to hypnotherapy: a pilot study. Int J STD AIDS. 1999;10:730–734. doi: 10.1258/0956462991913420. [DOI] [PubMed] [Google Scholar]
- 75.Nagatomi R, Kaifu T, Okutsu M, Zhang X, Kanemi O, Ohmori H. Modulation of the immune system by the autonomic nervous system and its implication in immunological changes after training. Exerc Immunol Rev. 2000;6:54–74. [PubMed] [Google Scholar]
- 76.Nieman DC. Exercise immunology: practical applications. Int J Sports Med. 1997;18(Suppl 1):S91–S100. doi: 10.1055/s-2007-972705. [DOI] [PubMed] [Google Scholar]
- 77.Pedersen BK, Bruunsgaard H, Klokker M, Kappel M, MacLean DA, Nielsen HB, et al. Exercise-induced immunomodulation–possible roles of neuroendocrine and metabolic factors. Int J Sports Med. 1997;18(Suppl 1):S2–S7. doi: 10.1055/s-2007-972695. [DOI] [PubMed] [Google Scholar]
- 78.Shinkai S, Watanabe S, Asai H, Shek PN. Cortisol response to exercise and post-exercise suppression of blood lymphocyte subset counts. Int J Sports Med. 1996;17:597–603. doi: 10.1055/s-2007-972901. [DOI] [PubMed] [Google Scholar]
- 79.Pedersen BK, Nieman DC. Exercise immunology: integration and regulation. Immunol Today. 1998;19:204–206. doi: 10.1016/s0167-5699(98)01255-9. [DOI] [PubMed] [Google Scholar]
- 80.Okamoto S, Ibaraki K, Hayashi S, Saito M. Ventromedial hypothalamus suppresses splenic lymphocyte activity through sympathetic innervation. Brain Res. 1996;739:308–313. doi: 10.1016/s0006-8993(96)00840-2. [DOI] [PubMed] [Google Scholar]
- 81.Carr DJ, Gebhardt BM, Paul D. α adrenergic and mu-2 opioid receptors are involved in morphine-induced suppression of splenocyte natural killer activity. J Pharmacol Exp Ther. 1993;264:1179–1186. [PubMed] [Google Scholar]
- 82.Nielsen HB, Secher NH, Kristensen JH, Christensen NJ, Espersen K, Pedersen BK. Splenectomy impairs lymphocytosis during maximal exercise. Am J Physiol. 1997;272:R1847–R1852. doi: 10.1152/ajpregu.1997.272.6.R1847. [DOI] [PubMed] [Google Scholar]
- 83.Stein PK, Ehsani AA, Domitrovich PP, Kleiger RE, Rottman JN. Effect of exercise training on heart rate variability in healthy older adults. Am Heart J. 1999;138:567–576. doi: 10.1016/s0002-8703(99)70162-6. [DOI] [PubMed] [Google Scholar]
- 84.Rogausch H, del Rey A, Kabiersch A, Reschke W, Ortel J, Besedovsky H, et al. Endotoxin impedes vasoconstriction in the spleen: role of endogenous interleukin-1 and sympathetic innervation. Am J Physiol. 1997;272:R2048–R2054. doi: 10.1152/ajpregu.1997.272.6.R2048. [DOI] [PubMed] [Google Scholar]
- 85.Lysle DT, How T. Endogenous opioids regulate the expression of inducible nitric oxide synthase by splenocytes. J Pharmacol Exp Ther. 1999;288:502–508. [PubMed] [Google Scholar]
- 86.Stevens-Felten SY, Bellinger DL. Noradrenergic and peptidergic innervation of lymphoid organs. Chem Immunol. 1997;69:99–131. doi: 10.1159/000058655. [DOI] [PubMed] [Google Scholar]
- 87.Friedman EM, Irwin MR. Modulation of immune cell function by the autonomic nervous system. Pharmacol Ther. 1997;74:27–38. doi: 10.1016/s0163-7258(96)00200-8. [DOI] [PubMed] [Google Scholar]
- 88.Hasko G, Shanley TP, Egnaczyk G, Nemeth ZH, Salzman AL, Vizi ES, et al. Exogenous and endogenous catecholamines inhibit the production of macrophage inflammatory protein (MIP) 1 α via a β adrenoceptor mediated mechanism. Br J Pharmacol. 1998;125:1297–1303. doi: 10.1038/sj.bjp.0702179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Niijima A, Meguid MM. Influence of systemic arginine-lysine on immune organ function: an electrophysiological study. Brain Res Bull. 1998;45:437–441. doi: 10.1016/s0361-9230(97)00349-3. [DOI] [PubMed] [Google Scholar]
- 90.Kagitani F, Kimura A, Sato A, Suzuki A. The role of the spinal cord as a reflex center for the somatically induced reflex responses of splenic sympathetic and natural killer cell activity in anesthetized rats. Neurosci Lett. 1996;217:109–112. [PubMed] [Google Scholar]
- 91.Madden KS, Thyagarajan S, Felten DL. Alterations in sympathetic noradrenergic innervation in lymphoid organs with age. Ann NY Acad Sci. 1998;840:262–268. doi: 10.1111/j.1749-6632.1998.tb09566.x. [DOI] [PubMed] [Google Scholar]
- 92.ThyagaRajan S, Madden KS, Kalvass JC, Dimitrova SS, Felten SY, Felten DL. L-deprenyl-induced increase in IL-2 and NK cell activity accompanies restoration of noradrenergic nerve fibers in the spleens of old F344 rats. J Neuroimmunol. 1998;92:9–21. doi: 10.1016/s0165-5728(98)00039-3. [DOI] [PubMed] [Google Scholar]
- 93.Schedlowski M, Hosch W, Oberbeck R, Benschop RJ, Jacobs R, Raab HR, et al. Catecholamines modulate human NK cell circulation and function via spleen-independent β 2-adrenergic mechanisms. J Immunol. 1996;156:93–99. [PubMed] [Google Scholar]
- 94.Benschop RJ, Nijkamp FP, Ballieux RE, Heijnen CJ. The effects of b-adrenoceptor stimulation on adhesion of human natural killer cells to cultured endothelium. Br J Pharmacol. 1994;113:1311–1316. doi: 10.1111/j.1476-5381.1994.tb17141.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Carlson SL, Beiting DJ, Kiani CA, Abell KM, McGillis JP. Catecholamines decrease lymphocyte adhesion to cytokine-activated endothelial cells. Brain Behav Immun. 1996;10:55–67. doi: 10.1006/brbi.1996.0005. [DOI] [PubMed] [Google Scholar]
- 96.Suzuki S, Toyabe S, Moroda T, Tada T, Tsukahara A, Iiai T, et al. Circadian rhythm of leucocytes and lymphocytes subsets and its possible correlation with the function of the autonomic nervous system. Clin Exp Immunol. 1997;110:500–508. doi: 10.1046/j.1365-2249.1997.4411460.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Benschop RJ, Nieuwenhuis EE, Tromp EA, Godaert GL, Ballieux RE, van Doornen LJ. Effects of β-adrenergic blockade on immunologic and cardiovascular changes induced by mental stress. Circulation. 1994;89:762–769. doi: 10.1161/01.cir.89.2.762. [DOI] [PubMed] [Google Scholar]
- 98.Klokker M, Secher NH, Madsen P, Pedersen M, Pedersen BK. Adrenergic β1− and β1+ 2-receptor blockade suppress the natural killer cell response to head-up tilt in humans. J Appl Physiol. 1997;83:1492–1498. doi: 10.1152/jappl.1997.83.5.1492. [DOI] [PubMed] [Google Scholar]
- 99.Carlson SL, Fox S, Abell KM. Catecholamine modulation of lymphocyte homing to lymphoid tissues. Brain Behav Immun. 1997;11:307–320. doi: 10.1006/brbi.1997.0501. [DOI] [PubMed] [Google Scholar]
- 100.Pelley RP, Strickland FM. Plants, polysaccharides, and the treatment and prevention of neoplasia. Crit Rev Oncog. 2000;11:189–225. [PubMed] [Google Scholar]
- 101.Janeway CA., Jr The immune system evolved to discriminate infectious nonself from noninfectious self. Immunol Today. 1992;13:11–16. doi: 10.1016/0167-5699(92)90198-G. [DOI] [PubMed] [Google Scholar]
- 102.Takeda K, Kaisho T, Akira S. Toll-like receptors. Annu Rev Immunol. 2003;21:335–376. doi: 10.1146/annurev.immunol.21.120601.141126. [DOI] [PubMed] [Google Scholar]
- 103.Akira S, Takeda K, Kaisho T. Toll-like receptors: critical proteins linking innate and acquired immunity. Nat Immunol. 2001;2:675–680. doi: 10.1038/90609. [DOI] [PubMed] [Google Scholar]
- 104.Yeh W-C, Chen N-J. Another toll road. Nature. 2003;424:736–737. doi: 10.1038/424736a. [DOI] [PubMed] [Google Scholar]
- 105.Oshiumi H, Matsumoto M, Funami K, Akazawa T, Seya T. TICAM-1, an adopter molecules that participates in Toll-like receptor 3-mediated interferin-b induction. Nat Immunol. 2003;4:161–167. doi: 10.1038/ni886. [DOI] [PubMed] [Google Scholar]
- 106.Underhill DM, Ozinsky A, Hajjar AM, Stevens A, Wilson CB, Bassetti M, et al. The Toll-like receptor 2 is recruited to macrophage phagosomes and discriminates between pathogens. Nature. 1999;401:811–815. doi: 10.1038/44605. [DOI] [PubMed] [Google Scholar]
- 107.Gantner BN, Simmons RM, Canavera SJ, Akira S, Underhill DM. Collaborative induction of inflammatory responses by dectin-1 and Toll-like receptor 2. J Exp Med. 2003;197:1107–1117. doi: 10.1084/jem.20021787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Weis WI, Taylor ME, Drickamer K. The C-type lectin superfamily in the immune system. Immunol Rev. 1998;163:19–34. doi: 10.1111/j.1600-065x.1998.tb01185.x. [DOI] [PubMed] [Google Scholar]
- 109.Hofer E, Sobanov Y, Brostjan C, Lehrach H, Duchler M. The centromeric part of the human natural killer (NK) receptor complex: lectin-like receptor genes expressed in NK, dendritic and endothelial cells. Immunol Rev. 2001;181:5–19. doi: 10.1034/j.1600-065x.2001.1810101.x. [DOI] [PubMed] [Google Scholar]
- 110.Ross GD, Vetvicka V, Yan J, Xia Y, Vetvickova J. Therapeutic intervention with complement and β-glucan in cancer. Immunopharmacology. 1999;42:61–74. doi: 10.1016/s0162-3109(99)00013-2. [DOI] [PubMed] [Google Scholar]
- 111.Tzianabos AO. Polysaccharide immunomodulators as therapeutic agents: structural aspects and biologic function. Clin Microbiol Rev. 2000;13:523–533. doi: 10.1128/cmr.13.4.523-533.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Muta T, Iwanaga S. The role of hemolymph coagulation in innate immunity. Curr Opin Immunol. 1996;8:41–47. doi: 10.1016/s0952-7915(96)80103-8. [DOI] [PubMed] [Google Scholar]
- 113.Battle J, Ha T, Li C, Della Beffa V, Rice P, Kalbfleisch J, et al. Ligand binding to the (1→3)-β-D-glucan receptor stimulates NFκB activation, but not apoptosis in U937 cells. Biochem Biophys Res Commun. 1998;249:499–504. doi: 10.1006/bbrc.1998.9175. [DOI] [PubMed] [Google Scholar]
- 114.Zimmerman JW, Lindermuth J, Fish PA, Palace GP, Stevenson TT, DeMong DE. A novel carbohydrate–glycosphingolipid interaction between a β-(1–3)-glucan immunomodulator, PGG-glucan, and lactosylceramide of human leukocytes. J Biol Chem. 1998;273:22014–22020. doi: 10.1074/jbc.273.34.22014. [DOI] [PubMed] [Google Scholar]
- 115.Rice PJ, Kelley JL, Kogan G, Ensley HE, Kalbfleisch JH, Browder IW, et al. Human monocyte scavenger receptors are pattern recognition receptors for (1→3)-β-D-glucans. J Leukoc Biol. 2002;72:140–146. [PubMed] [Google Scholar]
- 116.Brown GD, Gordon S Immune recognition. A new receptor for b-glucans. Nature. 2001;413:36–37. doi: 10.1038/35092620. [DOI] [PubMed] [Google Scholar]
- 117.Willment JA, Gordon S, Brown GD. Characterization of the human β-glucan receptor and its alternatively spliced isoforms. J Biol Chem. 2001;276:43818–43823. doi: 10.1074/jbc.M107715200. [DOI] [PubMed] [Google Scholar]
- 118.Taylor PR, Brown GD, Reid DM, Willment JA, Martinez-Pomares L, Gordon S, et al. The β-glucan receptor, dectin-1, is predominantly expressed on the surface of cells of the monocyte/macrophage and neutrophil lineages. J Immunol. 2002;169:3876–3882. doi: 10.4049/jimmunol.169.7.3876. [DOI] [PubMed] [Google Scholar]
- 119.Brown GD, Gordon S. Fungal β-glucans and mammalian immunity. Immunity. 2003;19:311–315. doi: 10.1016/s1074-7613(03)00233-4. [DOI] [PubMed] [Google Scholar]
- 120.Lise LD, Audibert F. Immunoadjuvants and analogs of immunomodulatory bacterial structures. Curr Opin Immunol. 1989;2:269–274. doi: 10.1016/0952-7915(89)90199-4. [DOI] [PubMed] [Google Scholar]
- 121.Parham P. NK cell receptors: of missing sugar and missing self. Curr Biol. 2000;10:R195–R197. doi: 10.1016/s0960-9822(00)00350-x. [DOI] [PubMed] [Google Scholar]
- 122.Rudd PM, Wormald MR, Stanfield RL, Huang M, Mattsson N, Speir JA, et al. Roles for glycosylation of cell surface receptors involved in cellular immune recognition. J Mol Biol. 1999;293:351–366. doi: 10.1006/jmbi.1999.3104. [DOI] [PubMed] [Google Scholar]
- 123.Portoukalian J. Immunogenicity of glycolipids. Clin Rev Allergy Immunol. 2000;19:73–78. doi: 10.1385/criai:19:1:73. [DOI] [PubMed] [Google Scholar]
- 124.Rabinovich GA, Rubinstein N, Fainboim L. Unlocking the secrets of galectins: a challenge at the frontier of glyco-immunology. J Leukoc Biol. 2002;71:741–752. [PubMed] [Google Scholar]
- 125.Wasser SP. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl Microbiol Biotechnol. 2002;60:258–274. doi: 10.1007/s00253-002-1076-7. [DOI] [PubMed] [Google Scholar]