

Abstract
WRKY transcription factors are plant-specific, zinc finger-type transcription factors. The WRKY superfamily is involved in abiotic stress responses in many crops including cotton, a major fiber crop that is widely cultivated and consumed throughout the world. Salinity is an important abiotic stress that results in considerable yield losses. In this study, we identified 109 WRKY genes (GarWRKYs) in a salt-tolerant wild cotton species Gossypium aridum from transcriptome sequencing data to elucidate the roles of these factors in cotton salt tolerance. According to their structural features, the predicted members were divided into three groups (Groups I–III), as previously described for Arabidopsis. Furthermore, 28 salt-responsive GarWRKY genes were identified from digital gene expression data and subjected to real-time quantitative RT-PCR analysis. The expression patterns of most GarWRKY genes revealed by this analysis are in good agreement with those revealed by RNA-Seq analysis. RT-PCR analysis revealed that 27 GarWRKY genes were expressed in roots and one was exclusively expressed in roots. Analysis of gene orthology and motif compositions indicated that WRKY members from Arabidopsis, rice and soybean generally shared the similar motifs within the same subgroup, suggesting they have the similar function. Overexpression-GarWRKY17 and –GarWRKY104 in Arabidopsis revealed that they could positively regulate salt tolerance of transgenic Arabidopsis during different development stages. The comprehensive data generated in this study provide a platform for elucidating the functions of WRKY transcription factors in salt tolerance of G. aridum. In addition, GarWRKYs related to salt tolerance identified in this study will be potential candidates for genetic improvement of cultivated cotton salt stress tolerance.
Introduction
Plants have developed a series of complex, effective systems to protect themselves from a variety of adverse environmental conditions during growth over the long course of evolution [1]. Studies have shown that these coping mechanisms mainly function via transcriptional activation or inhibition of transcription-related genes. Transcription factors play an important role in this process by activating or inhibiting the expression of target genes alone or via interactions with other proteins [2–4].
WRKY transcription factors are recently identified plant-specific transcription factors comprising one or two WRKY domains and a zinc-finger motif. The name WRKY is derived from the highly conserved WRKY domain, which consists of 60 amino acids. The WRKY domain comprises the highly conserved WRKYGQK sequence followed by a novel zinc-finger motif (C2H2orC2HC) located in the N-terminus [5]. Based on the number of WRKY domains and the pattern of zinc-finger motifs, WRKY proteins can be classified into three main groups and eight subgroups [5]. Group I WRKYs typically have two WRKY domains containing a C2H2 zinc-finger motif. The C-terminal WRKY domain has DNA binding activity. However, the N-terminal WRKY domain cannot bind to DNA alone, but it can assist the C-terminal WRKY domain in its binding to DNA, increasing its DNA binding affinity and specificity. In addition, the zinc-finger motif followed by the N-terminal WRKY domain may provide a protein–protein interaction interface; the zinc-finger motif’s structure is C2H2 (C-X4–5-C-X22–23-H-X-H, X is any amino acid). Group II WRKYs have a single WRKY domain with a C2H2 zinc-finger motif, which is the same as the C-terminal domain of Group I. Based on the phylogeny of the WRKY domains, group II WRKYs can be further divided into five subgroups: IIa (C-X5-CX23-HXH), IIb (C-X5-CX23-HXH), IIc (C-X4-CX23-HXH), IId (C-X5-CX23-HXH) and IIe (C-X5-CX23-HXH). Group III WRKYs also have a single WRKY domain, but their zinc-finger motif structure is C2HC (C-X7-C-X23-H-X-C). Based on the zinc finger structure, Group III can be further divided into two subgroups: IIIa and IIIb. The structure of subgroup IIIa zinc fingers is C-X7-C-X23-HXC, while that of subgroup IIIb is C-X7C-Xn-HXC (n ≥ 24) [6]. WRKYs specifically bind to the W-box, i.e., (T) (T) TGAC (C/T), in the promoter regions of target genes to regulate their transcription levels, enabling them to perform their biological functions [5].
Since the first report of SPF1 WRKY transcription factors from sweet potato in 1994 [7], numerous WRKY transcription factors have been experimentally identified from many other plant species including Arabidopsis, rice, tobacco, potato, cotton, barley, wheat, chamomile, soybean, cacao, grape, tomato, cucumber and so on [8–20]. Except for plant development, WRKY family genes also play important roles in plant biotic and abiotic stress responses including pathogen-induced defense programs and responses to drought, salt, stress et al.[20–22]. One of the most important functions of WRKYs appears to be regulation of the salt stress response. For example, AtWRKY25 and AtWRKY33 play a role in the salt stress response and improve salt tolerance in Arabidopsis [23]. OsWRKY11 and OsWRKY45 play a regulatory role in salt tolerance in transgenic rice [24,25]. GmWRKY54-overexpressing plants are more salt and drought tolerant than the wild type, and GmWRKY13 overexpression results in increased sensitivity to salt and mannitol stress [26]. Finally, overexpression of TaWRKY10 enhances drought and salt tolerance in transgenic tobacco plants [27].
In recent years, some WRKY transcription factors have also been reported in cotton. Guo et al. isolated GhWRKY3 from upland cotton (G. hirsutum), which is upregulated by the application of various phytohormones but not by cytokinin, auxin analog, drought, NaCl or cold [18]. Yu et al. identified GhWRKY15 from G. hirsutum, which is involved in disease resistance and plant development [28]. Shi et al. isolated a putative IId WRKY gene, GhWRKY39, from G. hirsutum and showed that it is activated by pathogens and salt stress. GhWRKY39-overexpressing Arabidopsis plants display enhanced tolerance to salt [29]. Zhou et al. identified 26 genes encoding putative WRKY proteins in G. hirsutum; five GhWRKY genes are upregulated in roots treated with NaCl [30]. With the release of the G. raimondii genome sequence, two comprehensive studies of G. raimondii WRKY family members were conducted, revealing 120 and 116 GrWRKY genes and 109 WRKY genes in G. arboreum [31–33], respectively; the expression of these genes under abiotic stress was analyzed in G. hirsutum [31,32]. All of these results suggest that WRKY genes are involved in the response to salinity stress in cotton.
High salinity is a major abiotic stress in cotton production worldwide. However, as described in detail previously [34], modern cotton cultivars, which are often grown under unstressed conditions, are the result of intensive selection to produce large amounts of specific types of fiber. Selection has unintentionally narrowed the genetic variability for salt tolerance. G. aridum is a D-genome diploid species from the Pacific coastal states of Mexico that shows remarkably tolerance to salt stress. Understanding how G. aridum responds to and develops tolerance to salt stress is the first step towards improving the adaptation of cotton to salt stress. In a previous study, we performed deep sequencing analysis of the G. aridum transcriptome in response to salt stress. A total of 281 unigenes were annotated as WRKY family members. Differentially expressed genes encoding transcription factors belonging to the WRKY family were predominant during different stages of salt stress [34]. These results suggest that WRKY family members are important regulators of the response to salt stress in G. aridum. In the current study, we focused on whole transcriptome-wide identification of salt-responsive members of the WRKY family in G. aridum. The results of this study increase our understanding of the GarWRKY gene family, which will facilitate the genetic improvement of salt stress tolerance in cotton.
Materials and Methods
Plant materials and treatment conditions
Seeds from the wild Gossypium species G. aridum were kindly supplied by the National Wild Cotton Plantation in Hainan Island, China. The same treatment procedure was used as described in Xu et al. [34]. G. aridum plants were treated with 200 mM NaCl for 0, 1, 3, 6, 12, 24 and 72 h. Root and leaf tissues were collected at every stage of stress treatment. All tissues were immediately frozen in liquid nitrogen and stored at −70°C.
GarWRKY gene identification and chromosomal location
The annotated genome sequences of G. raimondii were downloaded from http://www.phytozome.net/cn/. The WRKY domain (PF03106) downloaded from PFAM protein family database was used as a query for identifying WRKY transcription factors by HMMER software version 3.0[35]. The annotated G. raimondii WRKY genes were used to establish a local nucleic acid database. Based on de novo transcriptome sequencing data with a pooled RNA sample from roots and leaves of 12 h salt-stressed and unstressed plants [34], more than 90,000 transcripts were identified and used to BLAST the G. raimondii WRKY gene database using a local BLAST program with E-value < 10−10. Redundant sequences were removed. The GarWRKY protein sequences were further analyzed to confirm the presence of WRKY domains using the InterPro program (protein sequence analysis & classification, http://www.ebi.ac.uk/interpro/) [36].
To determine the location of GarWRKY genes on chromosomes, the GarWRKY sequences were further used as query sequences for a BLASTN search against G. raimondii whole-genome scaffold data (http://www.phytozome.net). Finally, the locations of all 109 GarWRKYs in the G. raimondii genome were detected. Mapping of GarWRKY genes was performed using MapInspect (http://www.plantbreeding.wur.nl/UK/software_mapinspect.html). To better identify and classify the WRKY transcription factors (TFs) in Gossypium, a uniform nomenclature was assigned to the WRKY genes in G. aridum as in G. raimondii [31].
GarWRKY gene classification and phylogenetic analysis
To classify GarWRKY genes into different groups and subgroups, a total of 14 AtWRKY genes representing three main groups and seven subgroups were selected, including Group I (AtWRKY7 and AtWRKY28), Group IIa (AtWRKY25 and AtWRKY27), Group IIb (AtWRKY28 and AtWRKY36), Group IIc (AtWRKY40 and AtWRKY41), Group IId (AtWRKY42 and AtWRKY49), Group IIe (AtWRKY58 and AtWRKY60) and Group III (AtWRKY69 and AtWRKY70). Complete WRKY domains from all GarWRKY proteins and select AtWRKY proteins were used to align WRKY domains with ClustalW [37]. The consequential alignment was used to create a phylogenetic tree in MEGA v4.0 using the neighbor-joining (NJ) method with 1,000 bootstrap replications [38].
To assess structural divergence of WRKY genes in different species, the Multiple Expectation Maximization for Motif Elicitation program (MEME, ttp://meme.nbcr.net/meme/tools/meme) was used to identify conserved motifs in the encoded proteins. Parameters employed in the analysis were: minimum motif width, 6; maximum motif width, 50; and maximum number of motifs, 10.
Isolation of salt-induced WRKY family genes
In a previous study, digital gene expression (DGE) analysis of G. aridum under salt stress revealed that the expression levels of 28 of 109 WRKY genes were significantly altered under salt stress. Using gene-specific primers (S2 Table) designed based on corresponding homologous gene in the G. raimondii genome, WRKY genes were cloned by PCR and their transcripts were amplified from G. aridum roots treated with 200 mM NaCl for 12 hours. PCR was performed using EasyPfu DNA Polymerase (TransGen Biotech, China). The PCR products were cloned into the PTG19-T vector (TransGen Biotech, China) and sequencing was performed by Invitrogen (Shanghai, China). The subcellular localizations of proteins were predicted by CELLO v.2.5 (subcellular localization predictor, http://cello.life.nctu.edu.tw) [39].
Expression analysis of salt-induced GarWRKY members in G. aridum
RNA isolation
Total RNA from G. aridum leaves and roots was extracted using an improved CTAB method [40]. For RT-PCR, first-strand cDNA was synthesized with a BU-SuperScript RT Kit (Biouniquer, China) according to the manufacture’s protocol. For quantitative real-time PCR, first-strand cDNA was synthesized with PrimeScript RT Master Mix (Perfect Real Time) from TaKaRa (Dalian, China).
Semi-quantitative RT-PCR
Semi-quantitative RT-PCR was performed to detect the differential expression of GarWRKY genes in different tissues including root, stem, leaf, petal, stigma, stamen and calyx tissue under normal conditions. The RT-PCR condition were as follow: preheating at 94°C for 3 min, followed by 36 cycles of 95°C for 45 s, 60°C for 45 s and 72°C for 1 min, with a final extension at 72°C for 10 min to complete the reaction. The PCR products were separated on a 1.2% agarose gel and quantified using an Imaging System (Bioshine GelX 1520, China). The specific primers used for RT-PCR are listed in S3 Table. The cotton actin gene was used as an internal reference gene (S3 Table).
Quantitative real-time PCR
The qRT-PCR was performed on an ABI PRISM 7500 Real-Time PCR System. The qRT-PCR reaction system (20 μl) contained 10 μl 2×SYBR Premix Ex Taq, 1 μl cDNA temple, 0.4 μl each of forward and reverse primer (10 μΜ) and 0.4 μl ROX Reference Dye II (50×). The thermal cycling conditions were as follow: pre-denaturation at 95°C for 30 s, followed by 40 cycles at 95°C for 5 s and 60°C for 34 s. After the cycle was complete, melting curves analysis was performed at 60–95°C to verify the specificity of the amplicon for each primer pair. Quantitative real-time PCR was carried out with three biological replicates. The cotton actin gene was used as an internal reference gene. The specific primers used for qRT-PCR are listed in S3 Table. The relative gene expression values were analyzed by the 2-ΔΔt method.
Generation of GarWRKY17 and GarWRKY104 transgenic Arabidopsis plants
The cDNA sequence containing the full-length coding sequence of GarWRKY17 and GarWRKY104 were cloned into pCAMBIA2301 expression vector separately under the control of CaMV35S promoter between the Xba I and Kpn I sites. The recombinant plasmid pCAMBIA2301-GarWRKY17 and pCAMBIA2301-GarWRKY104 were transformed into Agrobacterium EH105 with electroporation method (V = 2.4kv). The Arabidopsis plants were transformed by the floral dip method [41]. Their T2 transgenic lines were obtained after screening of Kanamycin resistance, PCR and RT-PCR analysis.
Analysis of salt tolerance in transgenic Arabidopsis plants
To evaluate salt tolerance of GarWRKY17 and GarWRKY104 transgenic Arabidopsis plants during seed germination stage, 50 sterilized seeds of wide type (WT) and T2 generation transgenic lines (3 lines for GarWRKY17 and 2 lines for GarWRKY104) were sowed in MS medium with and without 150 mM NaCl. The experiment was conducted with three biological replicates. Germination rate was calculated after 10 days.
To obtain further evidence that whether the overexpression of GarWRKY17 and GarWRKY104 could confer resistance to salt stress during vegetative growth, sterilized seeds of WT and T2 transgenic Arabidopsis were sowed in soil and grew for 20 days. Plants were growing in a pot supplemented with 150ml water or 150mM NaCl. Phenotype symptoms were observed after 4 weeks.
To determine the activities of antioxidant enzymes, three-week-old WT seedlings and T2 seedlings of 2 GarWRKY17–overexpressing (GarWRKY17-10 and GarWRKY17-25) were exposed to 150 mM NaCl for 24h. Leaves were collected from 10 plants of wild type and two transgenic lines. The activities of superoxide dismutase (SOD) and peroxidase (POD) was assayed using the procedure described by Liu et al. [42]. The tests were performed in triplicate.
Results
Identification of WRKY genes in G. aridum and their chromosomal locations
In a previous study, we identified more than 90,000 transcripts from de novo transcriptome sequencing of G. aridum under salt stress [34]. To perform genome-wide analysis of WRKY genes in G. aridum, all of these transcripts were used to BLAST the G. raimondii WRKY gene database using a local BLAST program. A total of 109 genes in G. aridum were ultimately identified as possible members of the WRKY superfamily and named according to their position from the top to the bottom of cotton chromosomes 1–13 (Fig 1). To facilitate communication, a uniform nomenclature for the 109 GarWRKY genes identified in this work was adopted based on that used in G. raimondii by Cai et al. [31]. The authors identified 120 candidate WRKY genes in the G. raimondii genome. By comparison, 11 WRKY genes (WRKY7, 16, 33, 42, 44, 49, 89, 96, 102, 109 and 112) were not identified in the current study.
Fig 1. Chromosomal locations of GarWRKY genes in the G. raimondii genome.

The candidate WRKY genes in G. aridum were designated GarWRKY1 to GarWRKY120 based on their orders on chromosomes. Eleven WRKY genes in G. aridum (GarWRKY7, 16, 33, 42, 44, 49, 89, 96, 102, 109 and 112) not identified in the current study were replaced by corresponding WRKY genes in the G. raimondii genome based on a report by Cai et al. (2014). Salt-responsive GarWRKYs are indicated in red. The scale is in megabases (Mb).
GarWRKY genes are distributed on all 13 G. aridum chromosomes. Of these, chromosome 7 has the highest number of GarWRKY genes (13) and chromosome 5 has the lowest (3). The distribution patterns of these genes on individual chromosomes reveal some regions with numerous GarWRKY gene clusters. For example, GarWRKY genes located on chromosomes 4, 8, 9 and 11 appear to be concentrated on the lower or upper ends of the arms of chromosomes, respectively (Fig 1).
Classification and phylogenetic analysis of GarWRKY genes
The WRKY domain and the zinc finger motif are the most prominent structural features of WRKYs. The WRKY domain contains the highly conserved heptapeptide stretch WRKYGQK at its N-terminus followed by a zinc-finger motif. A total of 126 WRKY domains were found in the 109 GarWRKY protein sequences.
The amino acid sequences of 126 WRKY domains were aligned using ClustalW with default settings. Based on AtWRKY classification and the WRKY domain features of GarWRKYs, the GarWRKYs were mainly classified into three groups, designated Group I to Group III (Fig 2). Seventeen GarWRKYs were assigned to Group I; these proteins contain two WRKY domains, an N-terminal WD (NTWD) and a C-terminal WD (CTWD). The NTWD zinc-finger type is C-X4-C-X22-HXH and the CTWD zinc-finger type is C-X4-CX23-HXH. Eighty GarWRKYs were assigned to Group II, which contain a single WRKY domain. The zinc-finger type is also C2H2, but its structure is C-X4–5-C-X23-HXH, which is structurally different from the Group I zinc finger motif. These 80 Group II GarWRKYs were further divided into five subgroups: subgroup IIa (7), IIb (15) IIc (30), IId (15) and IIe (13). Twelve GarWRKYs were assigned to Group III; these proteins contain a single WRKY domain with a C2HC zinc-finger motif (C-X7-C-X23-HXC). Detailed information about the GarWRKYs can be found in S1 Table.
Fig 2. Phylogenetic analysis of WRKY domains in G. aridum.

WRKY protein name with the suffix ‘N’ or ‘C’ indicates the N-terminal WRKY domains or the C-terminal WRKY domains. The black arcs indicate different groups of WRKY domains. ● represent AtWRKY proteins, ► represent salt-responsive GarWRKY proteins.
Cloning and classification of salt-induced WRKY family genes
In a previous study, we performed digital gene expression profile analysis of G. aridum during different stages of salt stress and identified 28 WRKY ESTs that were differentially expressed in roots or leaves during at least one stage [34]. In the current study, using gene-specific primers designed based on the corresponding homologous genes in the G. raimondii genome, we performed PCR cloning of WRKY genes and amplified the transcripts from roots of G. aridum treated with 200 mM NaCl for 12 h. We obtained 28 cDNA sequences of WRKY genes with complete open reading frames (ORFs; GenBank accession numbers KM438453–KM438480; Table 1). The ORF length ranged from 474 bp to 1,848 bp, with an average length of 1,048 bp. The identified GarWRKY genes encode proteins ranging from 157 to 615 amino acids (aa) in length with an average of 348 aa. The similarities between GarWRKY and GrWRKY based on their full-length nucleotide sequences ranged from 93.5% to 99.88%. All 28 salt-responsive GarWRKY proteins were predicted to be located in the nucleus.
Table 1. Characterization of 28 salt-responsive GarWRKY members.
| GarWRKY | Group | ORF (bp) | Polypeptide (aa) | GrWRKY | Similarity (%) | Subcellular localization | GenBank accession number |
|---|---|---|---|---|---|---|---|
| GarWRKY5 | III | 921 | 306 | WRKY5 | 98.05 | Nuclear | KM438453 |
| GarWRKY6 | IIc | 1077 | 358 | WRKY6 | 99.07 | Nuclear | KM438454 |
| GarWRKY9 | IId | 951 | 316 | WRKY9 | 98.74 | Nuclear | KM438455 |
| GarWRKY17 | IIc | 957 | 318 | WRKY17 | 98.03 | Nuclear | KM438456 |
| GarWRKY22 | I | 1530 | 509 | WRKY22 | 99.22 | Nuclear | KM438457 |
| GarWRKY27 | IIe | 1386 | 461 | WRKY27 | 99.49 | Nuclear | KM438458 |
| GarWRKY28 | IIa | 933 | 310 | WRKY28 | 99.41 | Nuclear | KM438459 |
| GarWRKY29 | IIc | 960 | 319 | WRKY29 | 99.06 | Nuclear | KM438460 |
| GarWRKY31 | IIb | 1698 | 565 | WRKY31 | 99.04 | Nuclear | KM438461 |
| GarWRKY38 | IIc | 474 | 157 | WRKY38 | 99.37 | Nuclear | KM438462 |
| GarWRKY43 | IIe | 828 | 275 | WRKY43 | 98.56 | Nuclear | KM438463 |
| GarWRKY51 | IIa | 876 | 291 | WRKY51 | 93.5 | Nuclear | KM438464 |
| GarWRKY52 | IIa | 786 | 261 | WRKY52 | 98.85 | Nuclear | KM438465 |
| GarWRKY54 | IIb | 978 | 325 | WRKY54 | 98.56 | Nuclear | KM438466 |
| GarWRKY56 | III | 1008 | 335 | WRKY56 | 99.01 | Nuclear | KM438467 |
| GarWRKY65 | IIc | 867 | 288 | WRKY65 | 99.88 | Nuclear | KM438468 |
| GarWRKY67 | IIe | 978 | 325 | WRKY67 | 99.39 | Nuclear | KM438469 |
| GarWRKY72 | IId | 1020 | 339 | WRKY72 | 97.65 | Nuclear | KM438470 |
| GarWRKY75 | IIb | 1443 | 480 | WRKY75 | 98.83 | Nuclear | KM438471 |
| GarWRKY78 | IIc | 885 | 294 | WRKY78 | 98.8 | Nuclear | KM438472 |
| GarWRKY90 | III | 999 | 332 | WRKY90 | 98.8 | Nuclear | KM438473 |
| GarWRKY95 | I | 1215 | 404 | WRKY95 | 99.42 | Nuclear | KM438474 |
| GarWRKY104 | IIc | 966 | 321 | WRKY104 | 98.65 | Nuclear | KM438475 |
| GarWRKY105 | IIe | 717 | 238 | WRKY105 | 99.02 | Nuclear | KM438476 |
| GarWRKY107 | IIb | 1848 | 615 | WRKY107 | 98.11 | Nuclear | KM438477 |
| GarWRKY113 | IIe | 1194 | 397 | WRKY113 | 98.49 | Nuclear | KM438478 |
| GarWRKY114 | III | 876 | 291 | WRKY114 | 95.97 | Nuclear | KM438479 |
| GarWRKY117 | IIc | 786 | 261 | WRKY117 | 99.46 | Nuclear | KM438480 |
Expression patterns of GarWRKY genes in various tissues revealed by semi-quantitative RT-PCR analysis
Using RT-PCR, we examined the expression patterns of GarWRKY genes in plants grown under normal growth conditions in seven different tissues including root, stem, leaf, petal, stigma, stamen and calyx tissue. As shown in Fig 3, 27 GarWRKY genes were expressed in roots, while 6 genes (GarWRKY5, GarWRKY17, GarWRKY27, GarWRKY75, GarWRKY90 and GarWRKY104) were expressed in all tissues (with varying expression levels). Moreover, 21 genes had different tissue-specific expression profiles and GarWRKY51 had very low transcript abundance in all tissues. Notably, GarWRKY52 was exclusively expressed in roots.
Fig 3. Expression patterns of GarWRKY genes in various tissues.

Semi-quantitative RT-PCR was conducted under normal growth condition. The cotton actin gene was used as an internal reference.
Validation of significant changes in GarWRKY expression in response to salt stress using real-time PCR
Gene expression patterns can provide important clues for determining gene function. To validate the results obtained from RNA-Seq, we performed quantitative RT-PCR analysis (Q-PCR) of the differentially expressed GarWRKY genes to confirm their levels of expression in roots and leaves after salt stress treatment. Quantitative expression analysis of these genes was performed using samples obtained from independent experiments carried out under the same conditions used to obtain RNA samples for RNA-Seq analysis. RNA was extracted from roots and leaves at 0, 1, 3, 6, 12, 24 and 72 h post salt stress, while RNA-Seq analysis was performed with fewer time points (0, 3, 12 and 72 h). As shown in Fig 4, the GarWRKY genes displayed distinct expression patterns in response to salt stress. Real-time PCR generally produced similar results to those obtained by differential expression profiling by RNA-Seq. However, in contrast to the RNA-Seq results, qRT-PCR revealed reduced expression of GarWRKY95 and GarWRKY114 in leaves.
Fig 4. Expression profiles of GarWRKY genes under NaCl stress.
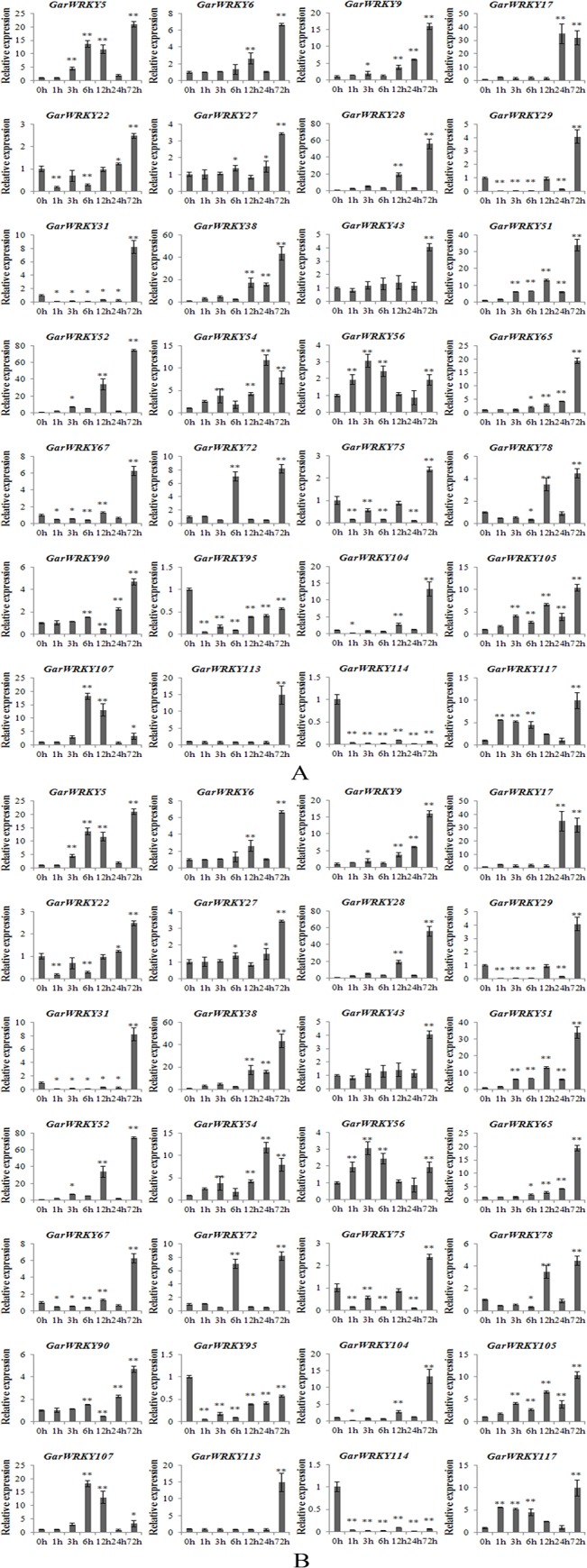
A: roots; B: leaves. The cotton actin gene was used as an internal reference. The data are mean ± SE of three biological replicates. * and ** indicate statistical significance at the 0.05 and 0.01 probability level, respectively.
Most GarWRKY genes in roots were activated within 1 h following salt treatment, with peak expression during the middle period and a return to basal levels during the latest period (72 h; Fig 4). However, in leaves, the majority of the analyzed WRKY genes were activated later than 6 h after salt treatment, with peak expression during the latest period (72 h). Expression analysis of WRKY genes during salt stress revealed delayed induction kinetics for most of the analyzed genes in leaves compared with roots, indicating that plants perceive salt stress and activate their defense machinery rapidly in roots.
Based on hierarchical clustering, the expression patterns of the GarWRKY genes were divided into five groups, i.e., Cluster A, Cluster B, Cluster C, Cluster D and Cluster E (Fig 5). Cluster A includes three genes (GarWRKY9, GarWRKY27 and GarWRKY107) that were slightly downregulated in roots but upregulated in leaves. Cluster B includes one gene (GarWRKY22) that was downregulated in roots and leaves. Cluster C contains nine genes (GarWRKY90, GarWRKY72, GarWRKY113, GarWRKY75, GarWRKY31, GarWRKY78, GarWRKY104, GarWRKY67 and GarWRKY29) that were upregulated in roots and downregulated in leaves at early stages (before 24 h) but upregulated at later stage (72 h). Cluster D contains two genes (GarWRKY95 and GarWRKY114) that were upregulated in roots but downregulated in leaves. Finally, Cluster E contains 13 genes (GarWRKY56, GarWRKY5, GarWRKY6, GarWRKY105, GarWRKY17, GarWRKY38, GarWRKY51, GarWRKY52, GarWRKY54, GarWRKY65, GarWRKY117, GarWRKY28 and GarWRKY43) that were upregulated in roots and leaves.
Fig 5. Heat map of the expression patterns of 28 salt-responsive GarWRKYs from RNA-Seq and qRT-PCR data.

Changes in expression levels are displayed from green (downregulated) to red (upregulated), as shown in the color gradient at the bottom right corner (color figure online). Heat maps were generated and hierarchical clustering was performed using Cluster 3.0 based on log2 fold-change data in response to NaCl stress. The normalized expression values from RNA-Seq data were provided in S4 Table.
Comparison of orthologous GarWRKY genes from Arabidopsis, rice and soybean
To further implicate the functions of GarWRKYs in plant defenses to stresses, the orthologs in Arabidopsis, rice and soybean of 28 salt-responsive WRKY members were identified using BLASTP (E < 1e−20), and the 58 top hit (17 in Arabidopsis, 19 in rice and 22 in soybean, respectively) were collected. A phylogenetic tree was constructed with the MEGA 4.0 software and conserved motif compositions were analyzed by MEME program.
Our results showed that all 86 WRKY members from different species could be unambiguously classified as Group I, GroupIIa, IIb, IIc, IId, IIe and Group III (Fig 6). In total, 10 motifs, named, motif 1–10, were identified. Motifs 1, 2, 3, 4, were located in the WRKY domains, while the other motifs were dispersed around the WRKY domains. Motifs 1 and 2 were conserved in all WRKY family members; motif 5 was conserved in GroupIIa and IIb except for GarWRKY54; motif 4 was specific to Group I WRKY members. In Group I, only GarWRKY22 and OsWRKY78 contained motif 10. In Group IId and IIe, motif 10 was found only in GarWRKY9, GarWRKY27, GarWRKY113, OsWRKY14 and OsWRKY31. Overall, similar motif compositions were found in WRKY members of Arabidopsis, rice and soybean within the same subgroup, indicating the WRKY members have the similar function in these plants. It is worthwhile to note that 20 orthologous WRKY members in Arabidopsis, rice and/or soybean were also induced under salt stress according to previous reports [20–26, 42–44] (Fig 6), further implying these 28 GarWRKY members may play an important role in underlying mechanism of gene regulation under salt stress.
Fig 6. Phylogenetic relationships and motif compositions of 28 salt-responsive GarWRKY members and their orthologs of Arabidopsis, rice and soybean.

Unrooted phylogenetic tree was constructed by using MEGA4.0. The motifs identified by MEME software are represented by colored boxes. * indicated the genes induced under salt tress.
Salt tolerance validation of GarWRKY17 and GarWRKY104 in transgenic Arabidopsis
GarWRKY17 and GarWRKY104 showed relatively higher differentially expression level in qRT-PCR analysis (Fig 4). Therefore, they were selected to generate overexpression transgenic Arabidopsis plants to further evaluate their function in response to salt stress,. When these lines germinated on the medium adding 150 mM NaCl, the germination rates of two GarWRKY104-overexpressing lines (GarWRKY104-1 and GarWRKY104-2) were significantly higher than that of WT (80.0% and 81.3% vs 22.0%), However, no significant difference was observed between WT and three GarWRKY17-overexpressing lines (GarWRKY17-4, -10, -25) (Fig 7).
Fig 7. Overexpressing GarWRKYs Arabidopsis plants at germination stage under saltstress.
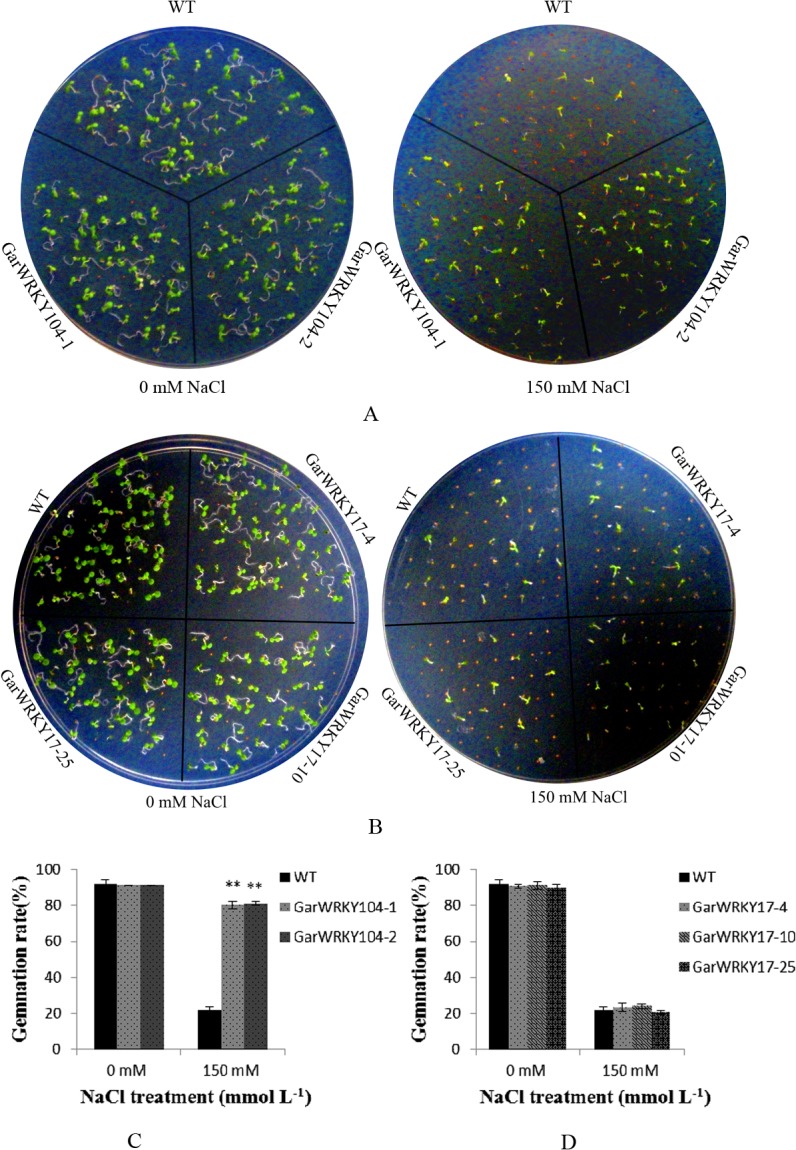
(A) Seedlings of the WT and 2 GarWRKY104-overexpressing lines germinated on MS medium adding 150 mM NaCl; (B) Seeds from the WT and 3 GarWRKY17-overexpressing lines were germinated on MS medium; (C) Germination rate of WT and 2 GarWRKY104-overexpressing lines; (D) Germination rate of WT and 3 GarWRKY17-overexpressing lines. The data are mean ± SE of three biological replicates. * and ** indicate statistical significance at the 0.05 and 0.01 probability level, respectively.
To further validate the salt tolerance of GarWRKY17 and GarWRKY104, 20 day-old overexpressing transgenic plants were treated with150 mM NaCl solution using water as the control. Four weeks later, growth of the WT seedlings was completely inhibited while the GarWRKY17–overexpressing seedlings remained green and continued to grow, especially for GarWRKY17-10 and GarWRKY17-25 lines (Fig 8). Similar to WT, the GarWRKY104-overexpressing lines displayed the inhibited growth (data not shown). These data indicate that GarWRKY17 positively regulate salt tolerance in a wide range of vegetative growth, while GarWRKY104 exhibited the salt terlance only during seed germination.
Fig 8. Effect of salt treatment on GarWRKY17-overexpressing Arabidopsis plants at vegetative growth stage.

(A) Plants of WT and 3 GarWRKY17-overexpressing Arabidopsis lines were watered with 150 mM NaCl solution; (B) The expression level of GarWRKY17 in WT and transgenic T2 lines; (C) SOD activity; (D) POD activity. The data are mean ± SE of three biological replicates. * and ** indicate statistical significance at the 0.05 and 0.01 probability level, respectively.
To study the physiological response of overexpressing-GarWRKY17 Arabidopsis plants to salt stress, two transgenic lines (GarWRKY17-10 and GarWRKY17-25) which showed better growth than WT plants during vegetative stage under salt stress were selected for further enzyme activities analysis. The activities of SOD and POD were determined in leaves of the control and the salt-treated Arabidopsis plants (Fig 8). The activity of SOD was much higher in the transgenic plants as compared to the WT plants with or without salt stress. In particular, the SOD activity in the two transgenic lines increased 2.6 times (GarWRKY17-10) and 2.8 times (GarWRKY17-25) compared with that in the wild type plants under salt stress. The activity of POD was significant higher in the transgenic plants as compared to the WT plants with or without salt stress. The activity of POD in the two transgenic lines increased 41% (GarWRKY17-10) and 36% (GarWRKY17-25) compared with that in WT plants under salt stress. These data indicate that overexpression of GarWRKY17 confer a more efficient antioxidant system to counteract the oxidative stress caused by salt stress.
Discussion
A few studies have revealed WRKY family members in G. hirsutum (cultivated species) and G. raimondii (for which complete genome sequence information is available), and the expression patterns of various WRKY TFs in response to abiotic stress (including salt stress) have been analyzed in upland cotton. As described in detail previously [34], modern cotton cultivars are the result of intensive selection to produce large amounts of specific types of fiber under unstressed conditions. Selection has unintentionally narrowed the genetic variability for salt tolerance. Therefore, improving salt tolerance in upland cotton requires the use of valuable alleles from other Gossypium species. In this study, we selected G. aridum, a D-genome diploid species that grows in the Pacific coastal states of Mexico, to conduct transcriptome-wide identification of WRKY family members. We identified 28 salt-responsive GarWRKY genes and validated their expression patterns in G. aridum using RNA-Seq and qRT-PCR technology. In a recent study, Cai et al. (2014) identified 120 candidate WRKY genes from G. raimondii and revealed that 17 WRKY genes are significantly induced by salt treatment in G. hirsutum [31]. Using uniform nomenclature, we found that WRKY22 in G. hirsutum and GarWRKY22 in G. aridum had similar expression patterns in G. hirsutum and G. aridum (significantly downregulated after salt treatment). However, WRKY6 in G. hirsutum and GarWRKY6 in G. aridum exhibited opposite expression patterns (downregulated in G. hirsutum and upregulated in G. aridum), indicating that some orthologous WRKY genes between G. hirsutum and G. aridum may have similar functions in regulating the stress response, while some may not.
In this study, we identified a total of 109 WRKY genes in G. aridum via de novo transcriptome sequencing of G. aridum under salt stress. In a recent report, Cai et al. identified 120 candidate WRKY genes from the G. raimondii genome, while the 11 remaining WRKY genes (WRKY7, 16, 33, 42, 44, 49, 89, 96, 102, 109 and 112) were not identified in the current study. Since G. aridum and G. raimondii belong to the same subgenome (D genome), they are expected to have the same number of WRKY genes in their genomes. The transcript abundance of these 11 WRKY genes was too low for them to be detected in the RNA-Seq libraries. These genes may be pseudogenes, or they may be expressed at specific developmental stages, under specific conditions or in specific tissues. In Cai’s study, the expression patterns of WRKY16 and WRKY44 were examined in different tissues. WRKY16 was not expressed in roots and was only slightly expressed in leaves, but it was preferentially expressed in fibers. WRKY44 exhibited very low transcript abundance in roots and leaves but preferential expression in anthers. These WRKY genes, which are expressed under specific conditions or in specific plant tissues, may be involved in the growth and development of different organs.
A number of studies have shown that WRKY genes are induced by salt stress and that overexpression of some WRKY genes alters stress tolerance in plants. In Arabidopsis, 18 out of 35 WRKY genes from roots are induced by salt stress, such as AtWRKY8, 25, 33, 46, 57 and so on [43]. In rice, 27 WRKY transcription factor genes are induced in response to salt stress, 26 of which are upregulated [44]. In soybean, 22 of 64 WRKY genes are differentially expressed under salt stress [26]. In transgenic rice, overexpression of OsWRKY11/01g43650 and OsWRKY45/05g2577 results in enhanced salt, heat and drought tolerance [25]. Overexpression of OsWRKY45 results in enhanced salt and drought tolerance, in addition to increased disease resistance [24]. In Arabidopsis, overexpression of either AtWRKY25 or AtWRKY33 increases salt tolerance [23], while overexpression of AtWRKY18 or AtWRKY60 increases plant sensitivity to salt [45]. GmWRKY54-overexpressing plants are more salt and drought tolerant than the control, and GmWRKY13 overexpression results in increased sensitivity to salt and mannitol stress [26]. In the current study, integrated transcriptome analysis indicated that 28 WRKY genes in G. aridum were differentially regulated in response to salt stress conditions. To further explore the functions of GarWRKYs that may be involved in plant defense responses to salt stress, we identified orthologous pairs among GarWRKYs, AtWRKYs, OsWRKYs and GmWRKYs based on full-length protein sequence similarities. Of the 28 salt-induced GarWRKY genes, 20 orthologs in Arabidopsis, rice and soybean also exhibit significant induction under salt stress. Among these, GarWRKY22, GarWRKY65 and GarWRKY5 are orthologous to AtWRKY33, GmWRKY54 and OsWRKY45, respectively. Motif compositions analysis showed that similar motif compositions were generally shared by WRKY members from Arabidopsis, rice and soybean within the same subgroup. These findings suggest that these members are important regulators of the response to salt stress. We also observed there was a possible relation between motif composition and gene function. GarWRKY22 contained motif 10, which was downregulated under salt stress. However, its orthologous gene in Arabidopsis (AtWRKY33) didn’t possessed motif 10, which was upregulated under salt stress [23]. Further study is required to clarify whether motif 10 is a key composition in regulating alteration of gene expression and functional divergence in different species.
Recent studies have identified and revealed the expression patterns of various salt-responsive WRKY TFs in cotton [18, 28–34,]. However, functional analysis of these TFs in an important crop such as cotton has rarely been performed. To date, only one WRKY gene in cotton has been functionally characterized [29], while the biological functions of most cotton WRKY genes remain largely unknown. In this study, we performed the first transcriptome-wide analysis of salt-responsive members of the WRKY family in G. aridum, a highly salt-tolerant wild species. Functional analysis by overexpression of GarWRKY17 and GarWRKY104 in Arabidopsis indicated they could positively regulate salt tolerance in Arabidopsis different development stages.This study demonstrates that a number of WRKY genes might be involved in the response to salt stress, and it provides clues for the selection of candidate genes for use in improving the stress tolerance of upland cotton.
Supporting Information
(DOC)
(DOC)
(DOC)
(DOC)
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by grants from the Major Project of the National Transgene (2014ZX08005-004), the National Natural Science Foundation of China (NSFC) (31171595) and the Jiangsu Independent Innovation Funds of Agricultural Technology (CX (13)3059).
References
- 1. Glazebrook J (2001) Genes controlling expression of defense responses in Arabidopsis–2001 status. Current Opinion in Plant Biology 4: 301–308. [DOI] [PubMed] [Google Scholar]
- 2. Chen W, Provart NJ, Glazebrook J, Katagiri F, Chang H, Eulgem T, et al. (2002) Expression profile matrix of Arabidopsis transcription factor genes suggests their putative functions in response to environmental stresses. The Plant Cell 14: 559–574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Kalde M, Barth M, Somssich IE, Lippok B (2003) Members of the Arabidopsis WRKY group III transcription factors are part of different plant defense signaling pathways. Molecular Plant-Microbe Interactions 16: 295–305. [DOI] [PubMed] [Google Scholar]
- 4. Pabo CO, Sauer RT (1992) Transcription factors: structural families and principles of DNA recognition. Annual Review of Biochemistry 61: 1053–1095. [DOI] [PubMed] [Google Scholar]
- 5. Rushton PJ, Somssich IE, Ringler P, Shen QJ (2010) WRKY transcription factors. Trends in Plant Science 15: 247–258. 10.1016/j.tplants.2010.02.006 [DOI] [PubMed] [Google Scholar]
- 6. Zhang Y, Wang L (2005) The WRKY transcription factor superfamily: its origin in eukaryotes and expansion in plants. BMC Evolutionary Biology 5: 1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Ishiguro S, Nakamura K (1994) Characterization of a cDNA encoding a novel DNA-binding protein, SPF1, that recognizes SP8 sequences in the 5′ upstream regions of genes coding for sporamin and β-amylase from sweet potato. Molecular and General Genetics 244: 563–571. [DOI] [PubMed] [Google Scholar]
- 8. Sun C, Palmqvist S, Olsson H, Borén M, Ahlandsberg S, Jansson C (2003) A novel WRKY transcription factor, SUSIBA2, participates in sugar signaling in barley by binding to the sugar-responsive elements of the iso1 promoter. The Plant Cell 15: 2076–2092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Chen C, Chen Z (2000) Isolation and characterization of two pathogen-and salicylic acid-induced genes encoding WRKY DNA-binding proteins from tobacco. Plant Molecular Biology 42: 387–396. [DOI] [PubMed] [Google Scholar]
- 10. Zhang Z, Xie Z, Zou X, Casaretto J, Ho TD, Shen Q (2004) A rice WRKY gene encodes a transcriptional repressor of the gibberellin signaling pathway in aleurone cells. Plant Physiology 134: 1500–1513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Lambais MR (2001) In silico differential display of defense-related expressed sequence tags from sugarcane tissues infected with diazotrophic endophytes. Genetics and Molecular Biology 24: 103–111. [Google Scholar]
- 12. Borrone JW, Kuhn DN, Schnell RJ (2004) Isolation, characterization, and development of WRKY genes as useful genetic markers in Theobroma cacao . Theoretical and Applied Genetics 109: 495–507. [DOI] [PubMed] [Google Scholar]
- 13. Pnueli L, Hallak-Herr E, Rozenberg M, Cohen M, Goloubinoff P, Kaplan A, et al. (2002) Molecular and biochemical mechanisms associated with dormancy and drought tolerance in the desert legume Retama raetam . The Plant Journal 31: 319–330. [DOI] [PubMed] [Google Scholar]
- 14. Ashida Y, Nishimoto M, Matsushima A, Watanabe J, Hirata T (2002) Molecular cloning and mRNA expression of geraniol-inducible genes in cultured shoot primordia of Matricaria chamomilla . Bioscience, Biotechnology, and Biochemistry 66: 2511–2514. [DOI] [PubMed] [Google Scholar]
- 15. Li H, Xu Y, Xiao Y, Zhu Z, Xie X, Zhao H, et al. (2010) Expression and functional analysis of two genes encoding transcription factors, VpWRKY1 and VpWRKY2, isolated from Chinese wild Vitis pseudoreticulata . Planta 232: 1325–1337. 10.1007/s00425-010-1258-y [DOI] [PubMed] [Google Scholar]
- 16. Xu Y, Wang J, Wang S, Wang J, Chen X (2004) Characterization of GaWRKY1, a cotton transcription factor that regulates the sesquiterpene synthase gene (+)-δ-cadinene synthase-A. Plant Physiology 135: 507–515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Ling J, Jiang W, Zhang Y, Yu H, Mao Z, Gu X, et al. (2011) Genome-wide analysis of WRKY gene family in Cucumis sativus . BMC Genomics 12: 471 10.1186/1471-2164-12-471 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Guo R, Yu F, Gao Z, An H, Cao X, Guo X (2011) GhWRKY3, a novel cotton (Gossypium hirsutum L.) WRKY gene, is involved in diverse stress responses. Molecular Biology Reports 38: 49–58. 10.1007/s11033-010-0076-4 [DOI] [PubMed] [Google Scholar]
- 19. Huang S, Gao Y, Liu J, Peng X, Niu X, Fei Z, et al. (2012) Genome-wide analysis of WRKY transcription factors in Solanum lycopersicum . Molecular Genetics and Genomics 287: 495–513. 10.1007/s00438-012-0696-6 [DOI] [PubMed] [Google Scholar]
- 20. Seki M, Narusaka M, Ishida J, Nanjo T, Fujita M, Oono Y, et al. (2002) Monitoring the expression profiles of 7000 Arabidopsis genes under drought, cold and high-salinity stresses using a full-length cDNA microarray. The Plant Journal 31: 279–292. [DOI] [PubMed] [Google Scholar]
- 21. Chen L, Song Y, Li S, Zhang L, Zou C, Yu D (2012) The role of WRKY transcription factors in plant abiotic stresses. Biochimica et Biophysica Acta-Gene Regulatory Mechanisms 1819: 120–128. [DOI] [PubMed] [Google Scholar]
- 22. Cheong Y, Chang H, Gupta R, Wang X, Zhu T, Luan S (2002) Transcriptional profiling reveals novel interactions between wounding, pathogen, abiotic stress, and hormonal responses in Arabidopsis . Plant Physiology 129: 661–677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Jiang Y, Deyholos MK (2009) Functional characterization of Arabidopsis NaCl-inducible WRKY25 and WRKY33 transcription factors in abiotic stresses. Plant MolecularBiology 69: 91–105. 10.1007/s11103-008-9408-3 [DOI] [PubMed] [Google Scholar]
- 24. Qiu Y, Yu D (2009) Over-expression of the stress-induced OsWRKY45 enhances disease resistance and drought tolerance in Arabidopsis . Environmental and Experimental Botany 65: 35–47. [Google Scholar]
- 25. Wu X, Kishitani S, Ito Y, Toriyama K (2009) Accumulation of raffinose in rice seedlings overexpressing OsWRKY11 in relation to desiccation tolerance. Plant Biotechnology 26: 431–434. [Google Scholar]
- 26. Zhou Q, Tian A, Zou H, Xie Z, Lei G, Huang J, et al. (2008) Soybean WRKY-type transcription factor genes, GmWRKY13, GmWRKY21, and GmWRKY54, confer differential tolerance to abiotic stresses in transgenic Arabidopsis plants. Plant Biotechnology Journal 6: 486–503. 10.1111/j.1467-7652.2008.00336.x [DOI] [PubMed] [Google Scholar]
- 27. Wang C, Deng P, Chen L, Wang X, Ma H, Hu W, et al. (2013) A wheat WRKY transcription factor TaWRKY10 confers tolerance to multiple abiotic stresses in transgenic tobacco. PloS one 8: e65120 10.1371/journal.pone.0065120 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Yu F, Hua X, Lu W, Wu C, Cao X, Guo X (2012) GhWRKY15, a member of the WRKY transcription factor family identified from cotton (Gossypium hirsutum L.), is involved in disease resistance and plant development. BMC Plant Biology 12: 144 10.1186/1471-2229-12-144 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Shi W, Liu D, Hao L, Wu C, Guo X, Li H (2014) GhWRKY39, a member of the WRKY transcription factor family in cotton, has a positive role in disease resistance and salt stress tolerance. Plant Cell, Tissue and Organ Culture 18: 1–16. [Google Scholar]
- 30.Zhou L, Wang N, Kong L, Gong S, Li Y, Li X (2014) Molecular characterization of 26 cotton WRKY genes that are expressed differentially in tissues and are induced in seedlings under high salinity and osmotic stress. Plant Cell, Tissue and Organ Culture 1–16.
- 31.Cai C, Niu E, Du H, Zhao L, Feng Y, Guo W (2014) Genome-wide analysis of the WRKY transcription factor gene family in Gossypium raimondii and the expression of orthologs in cultivated tetraploid cotton. The Crop Journal.
- 32.Dou L, Zhang X, Pang C, Song M, Wei H, Fan S, et al. (2014) Genome-wide analysis of the WRKY gene family in cotton. Molecular Genetics and Genomics: 1–19. [DOI] [PubMed]
- 33. Ding M, Chen J, Jiang Y, Lin L, Cao Y, Wang M, et al. (2015) Genome‑wide investigation and transcriptome analysis of the WRKY gene family in Gossypium , Mol Genet Genomics, 290:151–171. 10.1007/s00438-014-0904-7 [DOI] [PubMed] [Google Scholar]
- 34. Xu P, Liu Z, Fan X, Gao J, Zhang X, Zhang X, et al. (2013) De novo transcriptome sequencing and comparative analysis of differentially expressed genes in Gossypium aridum under salt stress. Gene 525: 26–34. 10.1016/j.gene.2013.04.066 [DOI] [PubMed] [Google Scholar]
- 35. Punta M, Coggill PC, Eberhardt RY, Mistry J, Tate J, Boursnell C, et al. (2012) The Pfam protein families database. Nucleic Acids Research 40: D290–D301. 10.1093/nar/gkr1065 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Jones P, Binns D, Chang H, Fraser M, Li W, McAnulla C, et al. (2014) InterProScan 5: genome-scale protein function classification. Bioinformatics: btu031. [DOI] [PMC free article] [PubMed]
- 37. Li KB (2003) ClustalW-MPI: ClustalW analysis using distributed and parallel computing. Bioinformatics 19: 1585–1586. [DOI] [PubMed] [Google Scholar]
- 38. Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Molecular Biology and Evolution 24: 1596–1599. [DOI] [PubMed] [Google Scholar]
- 39. Yu C, Chen Y, Lu C, Hwang J (2006) Prediction of protein subcellular localization. Proteins: Structure, Function, and Bioinformatics 64: 643–651. [DOI] [PubMed] [Google Scholar]
- 40. Hu G, Yu S (2007) Extraction of high-quality total RNA in cotton leaf with improved CTAB Method. Cotton science 19: 69–70. [Google Scholar]
- 41. Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agrobacterium mediated transformation of Arabidopsis thaliana. Plant J. 16, 735–743. [DOI] [PubMed] [Google Scholar]
- 42. Liu RX, Guo WQ, Chen BL, Zhou ZG (2008) Effects of N fertilization on root development and activity of water-stressed cotton plants, Agricultural Water Management, 95:1261–1270. [Google Scholar]
- 43. Jiang Y, Deyholos MK (2006) Comprehensive transcriptional profiling of NaCl-stressed Arabidopsis roots reveals novel classes of responsive genes. BMC Plant Biology 6: 25 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Ramamoorthy R, Jiang S, Kumar N, Venkatesh PN, Ramachandran S (2008) A comprehensive transcriptional profiling of the WRKY gene family in rice under various abiotic and phytohormone treatments. Plant and Cell Physiology 49: 865–879. 10.1093/pcp/pcn061 [DOI] [PubMed] [Google Scholar]
- 45. Chen H, Lai Z, Shi J, Xiao Y, Chen Z, Xu X (2010) Roles of Arabidopsis WRKY18, WRKY40 and WRKY60 transcription factors in plant responses to abscisic acid and abiotic stress. BMC Plant Biology 10: 281 10.1186/1471-2229-10-281 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOC)
(DOC)
(DOC)
(DOC)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.