

Abstract
The purpose of this study was to clarify the differences between adolescent and collegiate baseball pitchers in the kinematic and kinetic profiles of the trunk and lower limbs during the pitching motion. The subjects were thirty-two adolescent baseball pitchers aged 12-15 years (APG) and thirty collegiate baseball pitchers aged 18-22 years (CPG). Three-dimensional motion analysis with a comprehensive lower-extremity model was used to evaluate kinematic and kinetic parameters during baseball pitching. The ground reaction forces (GRFs) of the pivot and stride legs during pitching were determined using two multicomponent force plates. The joint torques of hip, knee, and ankle were calculated by the inverse-dynamics computation of musculoskeletal human models using motion-capture data. To eliminate any effect of variation in body size, kinetic and GRFs data were normalized by dividing them by body mass. The velocity of a pitched ball was significantly higher (p < 0.01) in CPG (35.2 ± 1.9 m·s-1) than in the APG (30.7 ± 2.7 m·s-1). Most kinematic parameters for the lower limbs were similar between the CPG and the APG. Maximum Fy (toward the throwing direction) on the pivot leg and Fy and resultant forces on the stride leg at ball release were significantly greater in the CPG than in the APG (p < 0.05). Hip and knee joint torques on the lower limbs were significantly greater in the CPG than in the APG (p < 0.05). The present study indicates that the kinematics of lower limbs during baseball pitching are similar between adolescent and collegiate pitchers, but the momentum of the lower limbs during pitching is lower in adolescent pitchers than in collegiate ones, even when the difference in body mass is considered.
Key points.
Collegiate baseball pitchers can generate the hip and knee joint torques on the pivot leg for accelerating the body forward.
Collegiate baseball pitchers can generate the hip and knee joint torques to control/stabilize the stride leg in order to increase momentum on the stride leg during the arm acceleration phase.
The kinematics of the lower limbs during baseball pitching are similar between adolescent and collegiate pitchers, but the momentum of the lower limbs during pitching is lower in adolescent pitchers than in collegiate ones, even when the difference in body mass is considered.
Adolescent baseball pitchers cannot generate the hip and knee joint torques in the pivot and stride leg for transfer of the energy of trunk and the arm.
Key words: Pitching ball velocity, the open kinetic chain, joint moment, ground-reaction force, motion analysis
Introduction
In baseball, the velocity of the pitched ball is one of the most important factors for the outcomes of games. At present, many conditioning programs such as pitching, long-distance running, sprint running, and resistance training are adopted to improve the pitched ball velocity. These training programs are applied to not only collegiate players but also adolescent ones, in spite of large differences between these two populations in terms of body size, strength capability, and training experience. In addition, kinetic profiles in pitching motion differ considerably between adolescent and collegiate players. From the findings of Fleisig et al. (1999), while nearly all of the kinematic and temporal parameters during pitching are quite similar between young players aged 10 to 15 years and either high school or collegiate players, ball velocity and kinetic parameters differ among these age groups. As a reason for the observed differences, they suggested the age-related difference in muscle strength capability. However, it should be noted that the prior study focused on the motions of the upper limbs and trunk, although knee flexion angles in front foot contact and ball release were determined. To our knowledge, no studies have examined how kinematics and kinetics of the lower limbs during pitching motion differ between adolescent and collegiate pitchers.
Pitching motion is a high-demand athletic skill involving fine coordination of all body segments (Atwater, 1979), and the mechanics of the lower limbs are recognized as an integral part of the pitching motion (Elliott et al., 1988; Kageyama et al., 2014; Mac Williams et al., 1998; Matsuo et al., 2001; Milewski et al., 2012; Robb et al., 2010). The contributions of the lower extremities to baseball pitchers and their motions have been described as the open kinetic chain (Kreighbaum and Barthels, 1985), in which all body segments are required to move the upper-extremity joints into appropriate positions to minimize the loads on each segment and transmit the generated force from the legs to more distal segments (Kibler, 1995). The lower extremities and trunk provide the beginning of the open kinetic chain that ends with force transmission to the baseball at the time of its release (Elliott et al., 1988; Mac Williams et al., 1998; Matsuo et al., 2001). Thus, the lower limbs have been considered to be important for constructing a stable base in which arm motion can be more efficiently and safely generated along with providing rotational momentum (Burkhart et al., 2003; Kibler, 1991).
The contribution of lower limbs for producing high pitched ball velocity has been examined by measuring kinetic and kinematic parameters. Elliott et al. (1988) have suggested that the ability to drive the body over a stabilized stride leg is a feature of high-ball-velocity pitchers. Mac Williams et al. (1998) reported that the maximum ground-reaction forces (GRFs) values in the pitching direction were 0.35 and 0.72 per body weight for the pivot and stride legs, respectively, and wrist velocity at the time of ball release was related to both these variables. In addition, Kageyama et al. (2014) indicated that collegiate high-ball-velocity pitchers could generate greater momentum by hip and knee joints in pivot and stride leg. These findings indicate that greater momentum of lower limbs during pitching plays an important role to throw ball with high velocity. However, less information on how the kinematics and kinetic on lower limbs during pitching motion differs between adolescent collegiate baseball pitchers is available from previous studies. To clarify this may provide useful information on training and technical guidance for adolescent baseball pitchers.
The purpose of this study was to clarify the differences between adolescent and collegiate baseball pitchers in the kinematic and kinetic profiles of lower limbs as well as trunk during the pitching motion.
Methods
Subjects
Thirty-two adolescent baseball pitchers aged 12-15 years (APG; right-handed, n = 29; left-handed, n = 3) and thirty collegiate baseball pitchers aged 18-22 years (CPG; right-handed, n = 25; left-handed, n = 5) voluntarily participated in this study. Descriptive data on the physical characteristics of the subjects are shown in Table 1. This study was approved by the Ethics Committee of the National Institute of Fitness and Sports in Kanoya and was consistent with their requirements for human experimentation. Prior to the measurements, all subjects and the parents of the adolescents were fully informed of the purpose as well as the procedures of this study and possible risks of the measurements, and gave their written informed consent.
Table 1.
Physical characteristics and ball velocity difference between adolescent and collegiate pitchers. Values are expressed as mean (±SD).
| Adolescent pitcher (n = 32) |
Collegiate pitcher (n = 30) |
ES | |
|---|---|---|---|
| Age (yr) | 13.9 (.6) | 19.6 (.9)* | 7.44 |
| Height (m) | 1.64 (.08) | 1.77 (.05)* | 2.02 |
| Weight (kg) | 54.1 (10.5) | 72.7 (.9)* | 1.85 |
| Ball velocity (m·s-1) | 30.7 (2.7) | 35.2 (1.9)* | 1.96 |
ES; effect size value.
* p < 0.01, Significant difference between adolescent and collegiate pitchers
Experimental design
The participants threw a baseball from a portable pitching mound towards a strike zone marked on a home plate. The force plate was attached to the rigid steel frame of the portable pitching mound. The distance between the portable pitching mound and the home plate was the same as the official pitching distance (18.44 m). Ball velocity was measured using a radar gun (2ZM-1035, Mizuno Corporation, Osaka, Japan) positioned behind the strike zone and adjusted to the position of the ball release. Prior to the pitching trials, participants performed warm-up exercises including stretching. After the completion of the warm-up exercises, the subjects were asked to perform only fastball pitches 10 times at maximal effort with an interval of about 15 seconds between the trials. In the present study, the kinematic and kinetic data in the fastest pitch passing the strike zone were used for detailed analysis.
Data collection
The GRFs were collected with two multicomponent force plates (Z15907, 60 × 120 cm, Kistler Corporation, Winterthur, Switzerland) attached to the rigid steel frame of a custom-built pitching mound. To simulate the sloped geometry of a regulation pitching mound in official baseball rules, the inclination angle of the portable pitching mound was set at 4.8°. The GRFs of the pivot and stride legs during pitching was measured using two multicomponent force plates, each of which had a sampling rate of 2000 Hz. One force plate was set below the rubber to record push-off forces during the windup and initial portions of delivery, and a second force plate recorded the landing force.
Thirty-six reflective markers aligned to specific body landmarks (Figure 1) were attached directly onto the skin to minimize movement artifacts. Three-dimensional coordinates were measured using a motion analysis system (Eagle System, Motion Analysis Corporation, Santa Rosa, CA) with 12 Eagle cameras with a sampling rate of 500 Hz and a shutter speed of 2000 Hz. The root-mean-square error in the calculation of the three-dimensional marker location was found to be less than 1.0 mm. The three-dimensional coordinates and the GRFs were synchronized using software (Cortex 1.1.4.368, Motion Analysis Corporation, Santa Rosa, CA) and then calculated. Marker position data were filtered using a fourth-order Butterworth low-pass filter with a cut-off frequency of 13.4 Hz (Fleisig et al., 1999). The GRFs and three-dimensional coordinates were defined as follows: Y-axis, throwing direction; Z-axis, vertical axis; X-axis, third-base direction, perpendicular to the Y- and Z-axes. The X-axis was reversed between right- and left-handers; the first-base direction for the left hander was defined as “+”.
Figure 1.
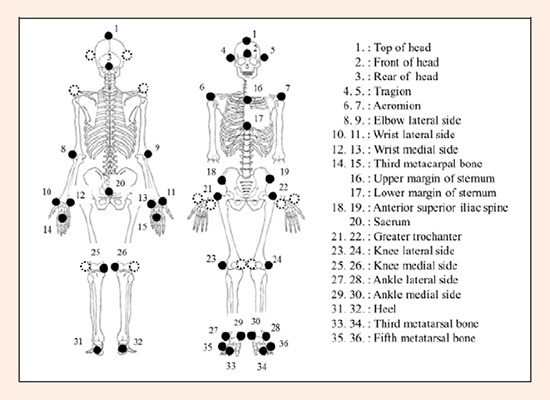
Placement of reflective markers on the body segment.
The ball velocity was measured using a radar gun. With a radar gun, the ball velocity may be lower than that gained with a marker on the ball. Therefore, we examined the difference in the velocity between the one got with a radar gun and the one gained with a marker using motion analysis system. As a result, the ball velocity gained with a radar gun had a significant correlation (r = 0.982, p < 0.01, n = 21) with that gained with a marker. In addition, there were no significant differences between the two methods (Radar gun: 32.6 ± 1.1 m·s-1 vs. Marker on ball: 32.8 ± 1.3 m·s-1). Thus, we adopted the measurement of the ball velocity by a radar gun in this study.
Data analysis
Kinematic and kinetic parameters were calculated with software (nMotion musculous 1.51, Motion Analysis Corporation, Santa Rosa, CA), utilizing the inverse-dynamics computation of musculoskeletal human models using motion-capture data (Nakamura et al., 2005). Kinematic parameters were calculated using the same methods as previously described elsewhere (Fleisig et al., 1996; Ishida and Hirano, 2004; Milewski et al., 2012; Stodden et al., 2001). The joint angles in the lower extremities were calculated using Euler equations of motion. Hip motion (coronal, sagittal, and transverse planes) and knee motion (sagittal plane) were calculated for both pivot and stride legs using standard angle definitions (Milewski et al., 2012; Figure 2 A-D, F). Stride length was measured and defined as the distance between the ankle joint centers at foot contact, expressed as a percentage of the subject’s height. Pelvis orientation was defined as the angle between a line connecting the two anterior superior iliac spine markers and the Y-axis in the XY plane (Stodden et al., 2001; Figure 2E). The upper torso orientation was defined as the angle between a line connecting the shoulder markers and the Y-axis in the XY plane (Stodden et al., 2001; Figure 2E). The pelvis and upper torso orientation angle was positive when they were “open” (i.e., their anterior aspect visible to the batter) and negative when they were “closed” (their posterior aspect visible to the batter) (Stodden et al., 2001; Ishida and Hirano, 2004; Figure 2E). Transverse plane rotation of the pelvis and upper torso orientation were measured with respect to the Y-axis (home plate). The pelvis and upper torso angle were at 90° of transverse rotation when they were square to the home plate. When the right and left anterior superior iliac spines were parallel to the home plate, the pelvic rotation equaled 90°. Trunk twist angle was defined as the difference between the pelvis and the upper torso angles (Ishida and Hirano, 2004; Figure 2E). Forward trunk tilt was the angle between the superior direction of the trunk and global Y (in the throwing direction) in the global YZ plane (Figure 2F). Forward trunk tilt was therefore 90° when the trunk was horizontal toward the target and 0° when the trunk was vertical (Fleisig et al., 1996; Matsuo et al., 2001; Stodden et al., 2001). For each displacement measurement, the corresponding velocity was calculated using the 5-point central difference method (Miller and Nelson, 1973). The joint torque was calculated at the hip, knee, and ankle using kinematic data, and inverse dynamics equations (Nakamura et al., 2005). To eliminate any effects of variation in body size, kinetic and GRFs data were normalized as divided by body weight. The pitching kinematics for left-handed subjects were calculated using the same conventions; however, it was necessary to mirror the world Z-axis so that all movements could be calculated, analyzed, and described from a right-hand point of view.
Figure 2.
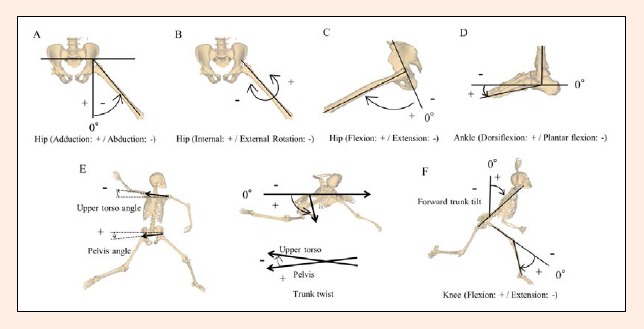
Definitions of kinematic variables. (A) Hip adduction/abduction, (B) hip (internal/external rotation), (C) hip (flexion/extension), (D) ankle (dorsiflexion/plantar flexion), (E) upper torso, pelvis angles and trunk twist, (F) forward trunk tilt and knee (flexion/extension).
To simplify interpretation of the results, throwing motion was divided it into six phases as previously defined for baseball pitching: windup, stride, arm cocking, arm acceleration, arm deceleration, and follow-through (Fleisig et al., 1996; 1999; Stodden et al., 2001). However, to simplify the phase of pitching motion, this study divided it into two phases (Figure 3) as previously described (Kageyama et al., 2014). The position during pitching define the points in time when the knee of the stride leg reached maximal height (MKH), the anterior (Y: toward the throwing direction) push-off force on the pivot leg reached a maximal value (MAP), the stride foot made contact with the ground (SFC), the shoulder joint reached maximal external rotation (MER), and the ball was released (REL). Ground contact was defined by the resultant force of the stride leg that was greater than 50 N. Data was analyzed from two phases in the present study. These two phases were defined as from MKH to SFC (phase 1), and from SFC to REL (phase 2). The GRFs on the pivot leg was mainly measured in the phase 1, whereas that on the stride leg was measured in the phase 2. The GRFs on the pivot leg was measured after SFC but its magnitude was small. Therefore, the GRFs on the pivot leg in the phase 2 was not analyzed. The GRFs on the stride leg was not measured because the stride foot was in the air until SFC. Temporal data were calculated, with the time of MKH defined as 0%, the time of SFC defined as 100%, and the time of REL defined as 200%. The angles of the trunk and lower legs were measured at five instances: MKH, MAP, SFC, MER, and REL.
Figure 3.
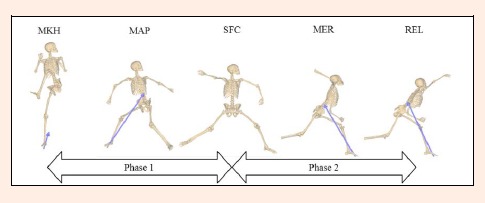
Phases of pitching motion. MKH; Maximal stride knee height. MAP; Maximal anterior push-off force. SFC; Stride foot contacts ground. MER; Maximum shoulder external rotation. REL; Ball release. Values measured from MHL until a particular event, expressed in time (s) or percentage of phase 1 (where 0% corresponds to the instant of maximal height of the knee of the stride leg and 100% corresponds to the instant of stride foot contact) and phase 2 (where 100% corresponds to the instant of stride foot contact and 200% corresponds to the instant of ball release).
Statistical analysis
Descriptive data are presented as means ± SD. A two-way repeated measures analysis of variance (ANOVA) (group ×time) was used to test the effects of group and time and their interaction on the kinematics and kinetic parameters. When a significant interaction was found, an unpaired Student’s t-test with a Bonferroni correction was used to test the difference in the measured variables between the APG and the CPG. In addition, the effect size (Cohen’s d) was calculated to express the magnitude of the difference between the two means. The threshold level values were < 0.20 (trivial), 0.20 – 0.49 (small), 0.50 – 0.79 (medium), and ≧ 0.80 (large) (Faul et al., 2007). The signifycance level was set at p < 0.05. All data were analyzed using SPSS Statistics 19 software (IBM Corporation, Chicago, IL).
Results
Table 1 shows the physical characteristics and ball velocity between the APG and the CPG. The physical characteristics and the pitched ball velocity were significantly greater in the CPG than in the APG (p < 0.01).
Kinematic parameters
Table 2 shows descriptive data on lower-limb kinematic parameters between the APG and the CPG. Pivot and stride hip abduction angles at MAP and maximum hip abduction and extension angular velocities were significantly lower in the CPG than in the APG (p < 0.05). Stride length (absolute value) and pivot and stride hip external rotation angles at MKH, pivot external rotation angle at MAP, pivot flexion angle at MAP, stride hip flexion angle at MKH and REL, stride knee flexion angle at SFC, pivot ankle dorsiflexion angle at SFC, stride ankle dorsiflexion angle at MKH, maximum hip flexion angle in pivot and stride legs and maximum hip internal rotation angular velocity were significantly greater in the CPG than in the APG (p < 0.05).
Table 2.
Lower-limb kinematic parameter difference between adolescent and collegiate pitchers. Values are expressed as mean (±SD).
| Variable | Adolescent pitcher (n = 32) |
Collegiate pitcher (n = 30) |
ES | Adolescent pitcher (n = 32) |
Collegiate pitcher (n = 30) |
ES |
|---|---|---|---|---|---|---|
| Phase time / Strength length | ||||||
| Phase 1 time (s) | .9 (.2) | .9 (.3) | .03 | |||
| Phase 2 time (s) | .2 (.0) | .2 (.0) | .27 | |||
| Total Pitch Time (s) | 1.1 (.2) | 1.1 (.3) | .01 | |||
| Stride length (m) | 1.4 (.1) | 1.5 (.1)** | 1.33 | |||
| Stride length (%height) | 82.9 (6.1) | 84.6 (4.2) | .33 | |||
| Angles | Pivot leg | Stride leg | ||||
| Hip Coronal Plane (Adduction:+; Abduction:- ) | ||||||
| Angle at MKH (°) | -23.1 (5.8) | -24.6 (5.9) | .26 | 20.9 (14.9) | 24.1 (12.8) | .23 |
| Angle at MAP (°) | -25.9 (9.8) | -19.7 (9.8)* | .62 | -31.7 (13.5) | -21.0 (13.2)** | .78 |
| Angle at SFC (°) | -42.5 (4.5) | -42.1 (5.4) | .08 | -38.5 (6.8) | -38.1 (6.0) | .06 |
| Angle at MER (°) | 27.7 (18.8) | 26.2 (21.7) | .08 | |||
| Angle at REL (°) | 33.9 (16.7) | 37.1 (14.0) | .20 | |||
| Hip Transverse Plane (Internal Rotation:+; External Rotation:- ) | ||||||
| Angle at MKH (°) | -18.3 (7.1) | -23.8 (6.0)** | .83 | -32.7 (10.3) | -39.9 (11.6) * | .64 |
| Angle at MAP (°) | -25.1 (6.5) | -31.0 (8.2)** | .79 | -39.3 (9.5) | -38.3 (8.2) | .11 |
| Angle at SFC (°) | -21.9 (8.8) | -25.5 (8.2) | .41 | -45.4 (8.3) | -48.5 (8.6) | .37 |
| Angle at MER (°) | -22.0 (12.2) | -23.7 (9.3) | .15 | |||
| Angle at REL (°) | -18.3 (10.4) | -16.1 (8.8) | .22 | |||
| Hip Sagittal Plane (Flexion:+; Extension:- ) | ||||||
| Angle at MKH (°) | 15.4 (7.7) | 16.3 (6.2) | .13 | 100.3 (10.3) | 108.9 (6.8)** | .96 |
| Angle at MAP (°) | 51.0 (10.0) | 59.2 (8.7) ** | .86 | 45.0 (13.1) | 46.0 (14.1) | .07 |
| Angle at SFC (°) | 20.5 (14.2) | 23.5 (13.7) | .21 | 58.1 (11.1) | 61.9 (12.5) | .31 |
| Angle at MER (°) | 105.4 (14.2) | 109.4 (13.8) | .28 | |||
| Angle at REL (°) | 97.9 (15.5) | 106.2 (10.0)* | .62 | |||
| Knee Sagittal Plane (Flexion:+; Extension:- ) | ||||||
| Angle at MKH (°) | 16.0 (7.0) | 17.0 (6.4) | .14 | 111.1 (18.5) | 112.0 (13.5) | .05 |
| Angle at MAP (°) | 50.3 (9.2) | 49.5 (9.4) | .09 | 35.9 (17.9) | 33.4 (15.0) | .15 |
| Angle at SFC (°) | 30.8 (13.1) | 28.1 (10.2) | .22 | 40.0 (8.2) | 46.8 (8.5)** | .80 |
| Angle at MER (°) | 39.0 (12.1) | 44.2 (10.9) | .44 | |||
| Angle at REL (°) | 31.4 (15.6) | 38.0 (13.7) | .44 | |||
| Ankle Sagittal Plane (Dorsiflexion:+; Plantar flexion:- ) | ||||||
| Angle at MKH (°) | 2.3 (5.3) | 2.1 (6.9) | .03 | .6 (18.4) | 11.6 (14.2)* | .66 |
| Angle at MAP (°) | -5.9 (9.6) | -1.4 (9.5) | .46 | 6.8 (11.5) | 10.8 (11.4) | .35 |
| Angle at SFC (°) | 21.8 (17.8) | 31.6 (10.3) * | .66 | 13.3 (12.8) | 16.6 (14.4) | .24 |
| Angle at MER (°) | 19.2 (5.5) | 19.7 (5.8) | .08 | |||
| Angle at REL (°) | 20.9 (6.5) | 21.4 (6.1) | .08 | |||
| Joint Angular Velocities | Pivot leg | Stride leg | ||||
| Max Hip Flex Angle (°) | 57.9 (9.3) | 62.6 (8.1) * | .52 | 106.4 (13.8) | 112.5 (8.8)* | .51 |
| Max Knee Flex Angle (°) | 58.6 (9.9) | 56.7 (10.1) | .19 | 47.8 (7.1) | 51.7 (8.3) | .49 |
| Max Hip Add AV (°/s ) | 117.9 (66.0) | 140.7 (47.3) | .39 | 852.0 (213.9) | 830.6 (160.9) | .11 |
| Max Hip Abd AV (°/s ) | 248.0 (73.7) | 279.8 (78.4) | .41 | 123.3 (137.9) | 38.8 (116.4)* | .65 |
| Max Hip IntRot AV (°/s ) | 110.8 (77.6) | 171.0 (122.7) * | .58 | 459.7 (13.8) | 499.7 (157.3) | .27 |
| Max Hip ExtRot AV(°/s ) | 72.0 (26.5) | 69.1 (25.0) | .11 | 82.4 (89.0) | 59.0 (63.1) | .30 |
| Max Hip Flex AV (°/s ) | 133.5 (41.5) | 140.1 (40.1) | .16 | 609.5 (98.8) | 632.9 (107.0) | .22 |
| Max Hip Ext AV (°/s ) | 515.6 (111.8) | 549.1 (100.8) | .30 | 262.7 (102.2) | 204.2 (98.5)* | .57 |
| Max Knee Ext AV (°/s ) | 239.0 (85.4) | 230.0 (76.6) | .11 | 245.7 (95.1) | 220.9 (117.8) | .23 |
| Knee Ext AV at MER (°/s ) | 167.9 (126.0) | 132.1 (122.8) | .28 | |||
| Knee Ext AV at REL (°/s ) | 199.4 (109.0) | 161.1 (117.3) | .33 | |||
ES; Effect Size value; MKH; Maximal stride knee height. MAP; Maximal anterior push-off force. SFC; Stride foot contacts ground. MER; Maximum shoulder external rotation. REL; Ball release. AV: Angular Velocity; Max: Maximum; Flex: Flexion; Ext: Extension; Add: Adduction; Abd: Abduction; IntRot:Internal Rotation; ExtRot: External Rotation.
* p < 0.05, Significant difference between adolescent and collegiate pitchers.
** p < 0.01, Significant difference between adolescent and collegiate pitchers.
Table 3 shows a comparison between the APG and the CPG in terms of trunk kinematic parameters. Pelvis, negative trunk twist and trunk tilt angles at MAP and maximum trunk tilt angular velocity were significantly lower in the CPG than in the APG (p < 0.05). Maximum upper torso angular velocity and upper torso and pelvis angular velocities at MER were significantly greater in the CPG than in the APG (p < 0.05).
Table 3.
Trunk kinematic parameter difference between adolescent and collegiate pitchers. Values are expressed as mean (±SD).
| Variable | Adolescent pitcher (n = 32) |
Collegiate pitcher (n = 30) |
ES |
|---|---|---|---|
| Angles | |||
| Upper Torso | |||
| Angle at MKH (°) | -17.5 (51.6) | -24.3 (16.9) | .17 |
| Angle at MAP (°) | -35.3 (13.7) | -32.9 (11.3) | .18 |
| Angle at SFC (°) | -32.1 (13.8) | -33.4 (9.5) | .11 |
| Angle at MER (°) | 82.7(12) | 79.8 (23.7) | .15 |
| Angle at REL (°) | 118.0 (8.2) | 122.5 (9.9) | .49 |
| Pelvis | |||
| Angle at MKH (°) | -36.4 (19.2) | -34.4 (16.7) | .11 |
| Angle at MAP (°) | -11.1 (12.7) | -17.6 (11.5) * | .53 |
| Angle at SFC (°) | 16.7 (10.4) | 14.8 (9.7) | .19 |
| Angle at MER (°) | 93.8 (8.8) | 90.2 (14.7) | .30 |
| Angle at REL (°) | 100.9 (8.0) | 101.9 (7.3) | .12 |
| Trunk twist | |||
| Angle at MKH (°) | 18.9 (58.8) | 10.1 (11.5) | .20 |
| Angle at MAP (°) | -24.2 (16.3) | -15.3 (14.6) * | .56 |
| Angle at SFC (°) | -48.8 (14.5) | -48.2 (11.6) | .04 |
| Angle at MER (°) | -11.1 (12.2) | -10.4 (12.7) | .06 |
| Angle at REL (°) | 17.1 (9.3) | 20.6 (8.3) | .39 |
| Trunk tilt | |||
| Angle at MKH (°) | -3.5 (6.2) | -3.6 (4.7) | .02 |
| Angle at MAP (°) | -9.3 (8.5) | -15.7 (6.2) ** | .83 |
| Angle at SFC (°) | -2.1 (6.6) | -4.0 (4.8) | .33 |
| Angle at MER (°) | 15.5 (6.1) | 12.9 (7.8) | .37 |
| Angle at REL (°) | 29.3 (7.6) | 25.4 (7.6) | .51 |
| Angular Velocities | |||
| Maximum Upper Torso Angular Velocity (°/s ) | 1170.2 (161.8) | 1273.0 (141.9) ** | .66 |
| Maximum Pelvis Angular Velocity (°/s ) | 727.4 (118.9) | 714.0 (71.2) | .13 |
| Maximum Trunk Positive Twist Angular Velocity (°/s ) | 821.7 (175.5) | 871.2 (149.9) | .30 |
| Maximum Trunk Negative Twist Angular Velocity (°/s ) | 359.0 (145.9) | 397.2 (118.2) | .28 |
| Maximum Trunk Tilt Angular Velocity (°/s ) | 381.3 (82.9) | 339.6 (59.3) * | .57 |
| Upper Torso Angular velocity at MER (°/s) | 1052.1 (188.1) | 1172.5 (235.8) * | .56 |
| Pelvis Angular velocity at MER (°/s) | 306.4 (129.6) | 376.7 (119.2) * | .55 |
ES; Effect Size value. MKH; Maximal stride knee height. MAP; Maximal anterior push-off force. SFC; Stride foot contacts ground. MER; Maximum shoulder external rotation. REL; Ball release.
* p < 0.05, Significant difference between adolescent and collegiate pitchers.
** p < 0.01, Significant difference between adolescent and collegiate pitchers
Kinetic parameters
Table 4 shows a comparison between the APG and the CPG in terms of GRFs. Fy on the pivot leg at MAP and Fy and resultant forces on the stride leg at REL were significantly greater in the CPG than in the APG (p < 0.05). Fz and resultant forces on the pivot leg at MKH were significantly lower in CPG than in APG (p < 0.05).
Table 4.
GRFs in pivot and stride leg difference between adolescent and collegiate pitchers. Values are expressed as mean (±SD).
| Variable | Adolescent pitcher (n = 32) |
Collegiate pitcher (n = 30) |
ES | Adolescent pitcher (n = 32) |
Collegiate pitcher (n = 30) |
ES |
|---|---|---|---|---|---|---|
| GRFs | Force Fx | Force Fz | ||||
| Force on pivot leg at MKH (N/kg) | -.2 (.3) | -.3 (.3) | .14 | 8.0 (1.5) | 6.8 (1.8)** | .69 |
| Force on pivot leg at MAP (N/kg) | -.3 (.7) | -.3 (.7) | .08 | 10.5 (1.9) | 10.8 (2.4) | .16 |
| Force on stride leg at MER (N/kg) | .6 (.9) | .4 (1.0) | .28 | 18.4 (1.8) | 18.4 (1.7) | .00 |
| Force on stride leg at REL (N/kg) | .9 (.7) | 1.2 (.9) | .39 | 16.6 (2.5) | 17.7 (2.2) | .47 |
| Force Fy | Resultant forces | |||||
| Force on pivot leg at MKH (N/kg) | .7 (.4) | .8 (.5) | .25 | 8.1 (1.5) | 7.0 (1.7)** | .69 |
| Force on pivot leg at MAP (N/kg) | 7.2 (1.3) | 8.4 (1.7)** | .74 | 12.8 (2.2) | 13.8 (2.7) | .40 |
| Force on stride leg at MER (N/kg) | -10.2 (1.6) | -10.9 (1.7) | .43 | 21.1 (2.1) | 21.5 (2.1) | .16 |
| Force on stride leg at REL (N/kg) | -7.9 (2.0) | -9.3 (1.8)** | .70 | 18.4 (3.0) | 20.1 (2.6)* | .57 |
| Maxima and minima of GRFs | Pivot leg | Stride leg | ||||
| Maximum Fx (N/kg) | 1.4 (.6) | 1.3 (.6) | .18 | 1.2 | 1.4 (.7) | .23 |
| Maximum Fy (N/kg) | 7.2 (1.3) | 8.4 (1.7) ** | .74 | |||
| Maximum Fz (N/kg) | 12.5 (1.7) | 12.9 (2.1) | .23 | 19.5 (2.0) | 19.2 (1.7) | .16 |
| Maximum Resultant forces (N/kg) | 14.0 (2.2) | 14.9 (2.6) | .37 | 22.2 (2.5) | 22.2 (2.0) | .03 |
| Minimum Fx (N/kg) | -.7 (.5) | -.7 (.5) | .06 | -1.3 (.8) | -1.1 (.7) | .22 |
| Minimum Fy (N/kg) | -10.9 (1.8) | -11.1 (1.7) | .11 | |||
ES; Effect Size value. MKH; Maximal stride knee height. MAP; Maximal anterior push-off force. SFC; Stride foot contacts ground. MER; Maximum shoulder external rotation. REL; Ball release.
* p < 0.05, Significant difference between adolescent and collegiate pitchers.
** p < 0.01, Significant difference between adolescent and collegiate pitchers
Table 5 shows a comparison between the APG and the CPG in terms of the joint torques of the lower limbs. The joint torques of pivot hip abduction at MAP, stride hip adduction at SFC, stride hip abduction at REL, pivot hip internal rotation at MAP, stride hip external rotation at SFC, pivot knee extension at MAP, and stride knee extension at MER and REL were significantly greater in the CPG than in the APG (p < 0.05). Pivot hip extension torque at MAP and SFC, pivot ankle dorsiflexion torque at SFC and stride ankle plantar flexion at MAP were significantly lower in the CPG than in the APG.
Table 5.
Lower-limb joint torque difference between adolescent and collegiate pitchers. Values are expressed as mean (±SD).
| Variable | Adolescent pitcher (n = 32) |
Collegiate pitcher (n = 30) |
ES | Adolescent pitcher (n = 32) |
Collegiate pitcher (n = 30) |
ES |
|---|---|---|---|---|---|---|
| Joint torques | Pivot leg | Stride leg | ||||
| Hip Coronal Plane (Adduction:+; Abduction:- ) | ||||||
| Joint torque at MKH (Nm/kg) | -.5 (.3) | -.4 (.3) | .17 | -.1 (.1) | -.1 (.1) | .02 |
| Joint torque at MAP (Nm/kg) | -1.1 (.6) | -1.8 (.9)** | .86 | .2 (.3) | .1 (.3) | .27 |
| Joint torque at SFC (Nm/kg) | .7 (.8) | -.4 (1.1) | .35 | .7 (.3) | 1.0 (.4) ** | 1.02 |
| Joint torque at MER (Nm/kg) | -1.7 (.5) | -1.5 (.8) | .22 | |||
| Joint torque at REL (Nm/kg) | -1.6 (.6) | -1.9 (.4) | .62 | |||
| Hip Transverse Plane (Internal Rotation:+; External Rotation:- ) | ||||||
| Joint torque at MKH (Nm/kg) | -.1 (.1) | -.0 (.1) | .30 | .0 (.1) | .1 (.1) | .21 |
| Joint torque at MAP (Nm/kg) | .3 (.3) | .7 (.6)** | .80 | -.1 (.1) | -.1 (.1) | .07 |
| Joint torque at SFC (Nm/kg) | -.3 (.3) | -.2 (.3) | .31 | -.2 (.1) | -.4 (.2) ** | 1.06 |
| Joint torque at MER (Nm/kg) | .5 (.3) | .4 (.4) | .27 | |||
| Joint torque at REL (Nm/kg) | .4 (.3) | .5 (.3) | .24 | |||
| Hip Sagittal Plane (Flexion:+; Extension:- ) | ||||||
| Joint torque at MKH (Nm/kg) | .0 (.3) | .1 (.7) | .36 | .1 (.2) | .1 (.3) | .22 |
| Joint torque at MAP (Nm/kg) | 51.0 (10.0) | -.5 (1.0) * | .58 | .1 (.3) | .2 (.2) | .22 |
| Joint torque at SFC (Nm/kg) | 20.5 (14.2) | -1.0 (.7) * | .54 | .0 (.2) | -.1 (.4) | .33 |
| Joint torque at MER (Nm/kg) | -2.0 (.7) | -2.2 (.7) | .22 | |||
| Joint torque at REL (Nm/kg) | -1.9 (.7) | -2.2 (.8) | .47 | |||
| Knee Sagittal Plane (Flexion:+; Extension:- ) | ||||||
| Joint torque at MKH (Nm/kg) | -.1 (.3) | -.3 (.4) | .39 | .0 (.1) | .0 (.1) | .22 |
| Joint torque at MAP (Nm/kg) | -1.0 (.6) | -1.7 (.7) | .91 | .1 (.1) | .1 (.1) | .29 |
| Joint torque at SFC (Nm/kg) | .3 (.4) | .4 (.4) | .11 | -.1 (.2) | -.1 (.2) | .21 |
| Joint torque at MER (Nm/kg) | -1.1 (.8) | -1.5 (.6) * | .58 | |||
| Joint torque at REL (Nm/kg) | -.5 (1.0) | -1.0 (.9) * | .53 | |||
| Ankle Sagittal Plane (Dorsiflexion:+; Plantar flexion:- ) | ||||||
| Joint torque at MKH (Nm/kg) | .3 (.2) | .3 (.2) | .29 | .0 (.0) | .0 (.0) | .26 |
| Joint torque at MAP (Nm/kg) | 1.1 (.5) | 1.0 (.5) | .16 | .0 (.0) | .0 (.0) * | .62 |
| Joint torque at SFC (Nm/kg) | .7 (.5) | .4 (.4) * | .59 | .0 (.1) | -.1 (.0) | .38 |
| Joint torque at MER (Nm/kg) | .9 (.5) | .7 (.5) | .25 | |||
| Joint torque at REL (Nm/kg) | .9 (.5) | .8 (.5) | .13 | |||
| Maximum joint torques | Pivot leg | Stride leg | ||||
| Max Hip Adduction Torque (Nm/kg) | 1.0 (.9) | .8 (.8) | .17 | .8 (.3) | 1.0 (.4) ** | .83 |
| Max Hip Abduction Torque (Nm/kg) | 2.0 (.5) | 2.4 (.8) * | .66 | 1.9 (.5) | 2.0 (.5) | .27 |
| Max Hip Internal Rot Torque (Nm/kg) | .6 (.3) | .9 (.5) * | .64 | .6 (.3) | .6 (.3) | .15 |
| Max Hip External Rot Torque (Nm/kg) | .4 (.3) | .4 (.3) | .26 | .3 (.1) | .4 (.2) ** | .87 |
| Max Hip Flexion Torque (Nm/kg) | .2 (.4) | .6 (.9) * | .59 | .0 (.2) | -.1 (.4) | .12 |
| Max Hip Extension Torque (Nm/kg) | 1.8 (.6) | 1.5 (.7) | .45 | 2.4 (.6) | 2.5 (.7) | .18 |
| Max Knee Flexion Torque (Nm/kg) | .5 (.4) | .5 (.4) | .13 | .2 (.4) | .0 (.4) | .49 |
| Max Knee Extension Torque (Nm/kg) | 1.6 (.5) | 2.2 (.6) ** | .93 | 1.7 (.4) | 1.9 (.4) * | .55 |
| Max Ankle Dorsiflexion Torque (Nm/kg) | 1.2 (.5) | 1.2 (.4) | .04 | 1.0 (.5) | .9 (.5) | .14 |
ES; Effect Size value. MKH; Maximal stride knee height. MAP; Maximal anterior push-off force. SFC; Stride foot contacts ground. MER; Maximum shoulder external rotation. REL; Ball release. Max; Maximum. Rot; Rotation.
* p < 0.05, Significant difference between adolescent and collegiate pitchers.
** p < 0.01, Significant difference between adolescent and collegiate pitchers
The maximum joint torques of hip abduction, hip internal rotation, hip flexion, and knee extension in pivot leg and hip adduction, hip external rotation, and knee extension in the stride leg were significantly greater in the CPG than in the APG (p < 0.05). Appearance of the maximum stride hip abduction torque (APG: 185.4 ± 8.2% time vs. CPG: 194.4 ± 6.2% time) and stride hip flexion torque (APG: 104.6 ± 7.9% time vs. CPG: 101.0 ± 3.0% time) were significantly later in the CPG than in the APG (p < 0.05).
Discussion
The pitched ball velocities for CPG (36.0 ± 1.6 m·s-1) and APG (31.0 ± 2.9 m·s-1) are higher than those reported previously for university baseball pitchers (33-35 m/s, Felter and Dapena, 1986; Fleisig et al., 1999; Sakurai et al., 1993; Stodden et al., 2001) and adolescent baseball pitchers aged 10 to 15 years (26.3-28.0 m·s-1; Dun et al., 2008; Fleisig et al., 1999), respectively. Thus, in the comparison within similar age group, the CPG and APG had greater pitching ability than those examined in previous studies.
Maximum Fy on the pivot leg was significantly greater in the CPG than in the APG (Table 4). Elliott et al. (1988) reported that the ability to drive the body over a stabilized stride leg is a feature of high-ball-velocity pitchers. In addition, Mac Williams et al. (1998) indicated that the maxima of GRFs (Fy, Fz, and resultant forces) on the pivot leg and Fz and Fy at MAP were highly correlated with wrist velocity at the time of ball release. On the basis of these results, Mac Williams et al. (1998) suggested that the landing leg serves as an anchor in transforming the forward and vertical momentum into rotational components; posteriorly directed forces at the landing foot reflect an overall balance of the inertial forces of the body moving forward to create ball velocity. Taking this into account, the greater maximum Fy on the pivot leg in collegiate baseball pitchers may be interpreted as that, compared with adolescent pitchers, they can generate the inertial forces for moving the body forward before stride foot contact.
In the pivot leg, joint torques during hip abduction, hip internal rotation, hip flexion, and knee extension were significantly greater in the CPG than in the APG (Table 5). The study that focused on the joint torques of the lower limbs during pitching motion is only a report of collegiate baseball pitchers by Kageyama et al. (2014). The current result indicates that the joint torques of the pivot leg during pitching motion in collegiate baseball pitchers were similar to those reported in Kageyama et al. (2014). Kageyama et al. (2014) found that collegiate high-ball-velocity pitchers could generate greater momentum by hip extension/abduction and knee extension in the pivot leg for accelerating the body forward. Campbell et al. (2010) observed that the activities of the gastrocnemius, vastus medialis, gluteus maximus, and biceps femoris of the pivot leg from stride knee peak flexion to stride foot contact, expressed as the values relative to their respective maximal voluntary isometric contractions, were 75, 68, 73, and 48%, respectively, which promoted concentric plantar flexion, knee extension, and hip extension. Considering these findings, it may be assumed that the observed differences between the adolescent and collegiate pitchers in the hip and knee joint torques during pitching motion could to be attributed to those in the muscular activities around the hip and knee and in the ability for accelerating the body forward.
Fy and resultant forces on the stride leg at REL were significantly greater in the CPG than in the APG (Table 4). The energy of the lower limbs during pitching is transferred to the trunk and arms (Elliott et al., 1988; Kageyama et al., 2014; Mac Williams et al., 1998; Matsuo et al., 2001; Milewski et al., 2012; Robb et al., 2010). Elliott et al. (1988) suggested that the ability to drive the body over a stabilized stride leg was a characteristic of high-ball-velocity pitchers. Mac Williams et al. (1998) reported that the maxima of GRFs (Fy, Fz, and resultant forces) on the stride leg and Fy, Fz, and resultant forces at REL correlated highly with wrist velocity at the time of ball release. Taking these findings into account together with the current results, it is likely that collegiate baseball pitchers can generate the inertial forces from MER to REL, which cause the upper body to move forward.
In the stride leg, joint torques during hip adduction, hip external rotation, and knee extension were significantly greater in the CPG than in the APG (Table 5). Knee extension torques on the stride leg at MER and REL were significantly greater in the CPG than in the APG (Table 5). The current result indicates that as compared to Kageyama et al. (2014), the joint torques of the stride leg during pitching motion in collegiate baseball pitchers were similar. Campbell et al. (2010) reported that the high activation of the vastus medialis in the stride leg during the phase 3 (from SFC to REL) indicates their important roles in controlling/stabilizing knee joint positions, whereas the upper extremities and torso forcefully rotate about the stride hip. Considering these finding, the current results support the findings of Kageyama et al. (2014) and the greater joint torque of the hip and knee for collegiate baseball pitchers may be assumed to contribute for controlling and/or stabilizing their stride legs in the phase from MER to REL.
Upper-torso and pelvis angular velocities were significantly greater in the CPG than in the APG (Table 3). Trunk rotation during pitching was shown to be an important factor for pitchers throwing at high velocity (Fleisig et al., 1999, Matsuo et al., 2001, Stodden et al., 2001). According to a report by Fleisig et al. (1999), college and professional baseball pitchers generally achieved higher upper-torso velocities than adolescent pitchers. In addition, Stodden et al. (2001) suggested that an increase in momentum transfer caused by increased pelvis and upper-torso velocities would increase the force at the shoulder and elbow, which is needed to accelerate the throwing arm. The current results support the findings of Fleisig et al. (1999) and suggest that as compared to adolescent baseball pitchers, collegiate baseball pitchers can generate the momentum of the lower limbs for increasing the energy of the trunk rotation and the arm.
The present study provides evidence that the difference in the pitched ball velocity between adolescent and collegiate baseball pitchers can be attributed to that in the momentum of the lower limbs, rather than the kinematics. Although high levels of lower-limb strength are necessary in pitching, the fact that the pitchers throwing at high velocity generated greater momentum of the lower limbs during pitching motion indicates improvements in dynamic muscular strength/power (Campbell et al., 2010; Elliott et al., 1988; Kageyama et al., 2014; Mac Williams et al., 1998; Matsuo et al., 2001). Fleisig et al. (1999) suggested that the increases in kinetic and velocity variables were due to increased strength and muscle mass in the higher-level pitchers. Notably, adolescent baseball pitchers cannot develop hip and knee joint torques corresponding to their body size compared with collegiate baseball pitchers. In addition, weakness in the knee and hip has been implicated as a potential area for a break in the open kinetic chain in the pitching cycle (Burkhart et al., 2003). In other words, it seems that in addition to a small momentum of the lower limbs, adolescent pitchers cannot perform properly the open kinetic chain which transfers the energy of the lower limbs during pitching to the trunk and arms.
A limitation of the current study was that subjects were throwing only fastballs. In baseball game, subjects would be pitching not only the fastball but also the breaking ball (e.g., curveball, change-up, slider, etc.). Fleisig et al. (2006) reported that collegiate baseball pitchers were significant differences in kinematic between the fastball and curveball. According to a report by Dun et al. (2008), youth baseball pitchers were different in kinematic and temporal among the 3 pitch types (fastball, curveball, change-up). Therefore, the current results may reflect only fastball. In the future, it will be necessary to examine differences in the kinematic and kinetic profiles of the trunk and lower limbs during baseball pitching between adolescent and collegiate baseball pitchers, in relation between fastball and breaking ball.
Conclusion
The present study indicates that the kinematics of the lower limbs during baseball pitching are similar between adolescent and collegiate pitchers, but the momentum of the lower limbs during pitching is lower in adolescent pitchers than in collegiate ones, even when the difference in body mass is considered. Thus, the current results indicates that as compared to collegiate baseball pitchers, adolescent baseball pitchers cannot generate the hip and knee joint torques in the pivot and stride leg, which contribute to transfer the energy of trunk and the arm.
Acknowledgments
This study was not funded. The authors would like to thank all individuals who participated in this study.
Biographies

Masahiro KAGEYAMA
Employment
Graduate School of Physical Education, National Institute of Fitness and Sports in Kanoya
Degree
MSc
Research interests
Biomechanics
E-mail: m127003@sky.nifs-k.ac.jp

Takashi SUGIYAMA
Employment
Graduate School of Physical Education, National Institute of Fitness and Sports in Kanoya
Degree
MSc
Research interests
Biomechanics
E-mail: m127004@sky.nifs-k.ac.jp

Hiroaki KANEHISA
Employment
National Institute of Fitness and Sports in Kanoya
Degree
PhD
Research interests
Exercise physiology
E-mail: hkane@nifs-k.ac.jp

Akira MAEDA
Employment
National Institute of Fitness and Sports in Kanoya
Degree
PhD
Research interests
Biomechanics
E-mail: amaeda@nifs-k.ac.jp
References
- Atwater A.E. (1979) Biomechanics of overarm throwing movements and of throwing injuries. Exercise and Sport Sciences Reviews 7, 43-85. [PubMed] [Google Scholar]
- Burkhart S.S., Morgan C.D., Kibler W.B. (2003) The disabled throwing shoulder: Spectrum of pathology part I: Patho anatomy and biomechanics. Arthroscopy: The Journal of Arthroscopic & Related Surgery 19(4), 404-420. [DOI] [PubMed] [Google Scholar]
- Campbell B.M., Stodden D.F., Nixon M.K. (2010) Lower extremity muscle activation during baseball pitching. Journal of Strength and Conditioning Research 24(4), 964-971. [DOI] [PubMed] [Google Scholar]
- Dun S., Loftice J., Fleisig G.S., Kingsley D., Andrews J.C. (2008) A biomechanical comparison of youth baseball pitches: Is the curveball potentially harmful? The American Journal of Sports Medicine 36(4), 686-692. [DOI] [PubMed] [Google Scholar]
- Elliott B., Grove J.R., Gibson B. (1988) Timing of the lower limb drive and throwing limb movement in baseball pitching. International Journal of Sport Biomechanics 4, 59-67. [Google Scholar]
- Escamilla R.F., Fleisig G.S., Barrentine S.W., Zheng N., Andrews J.R. (1998) Kinematic comparisons of throwing different types of baseball pitches. Journal of Applied Biomechanics 14, 1-23. [Google Scholar]
- Faul F., Erdfelder E., Lang A.G., Buchner A. (2007) G*Power 3: a flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods 39(2), 175-191. [DOI] [PubMed] [Google Scholar]
- Felter M., Dapena J. (1986) Dynamics of the shoulder and elbow joints of the throwing arm during a baseball pitch. International of Sport Biomechanics 2(4), 235-259. [Google Scholar]
- Fleisig G.S., Barrantine S., Zheng N., Escamilla R., Andrews J. (1999) Kinematic and kinetic comparison of baseball pitching among various levels of development. Journal of Biomechanics 32(12), 1371-1375. [DOI] [PubMed] [Google Scholar]
- Fleisig G.S., Escamilla R.F., Andrews J.R., Matsuo T. (1996) Kinematic and Kinetic Comparison Between Baseball Pitching and Football Passing. Journal of Applied Biomechanics 12, 207-224. [Google Scholar]
- Fleisig G.S., Kingsley D.S., Loftice J.W., Dinnen K.P., Ranganathan R., Dun S., Escamilla R.F., Andrews J.R. (2006) Kinetic comparison among the fastball, curveball, change-up, and slider in collegiate baseball pitchers. The American Journal of Sports Medicine 34(3), 423-430 [DOI] [PubMed] [Google Scholar]
- Ishida K., Hirano Y. (2004) Effects of Non-throwing Arm on Trunk and Throwing Arm Movements in Baseball Pitching. International Journal of Sport and Health Science 2, 119-128. [Google Scholar]
- Kageyama M., Sugiyama T., Takai Y., Kanehisa H., Maeda A. (2014) Kinematic and kinetic profiles of the lower limbs during baseball pitching in collegiate baseball pitchers. Journal of Sports Science and Medicine 13(4), 742-750. [PMC free article] [PubMed] [Google Scholar]
- Kibler W.B. (1991) Role of the scapula in the overhead throwing motion. Contemporary Orthopaedics 22, 525-532. [Google Scholar]
- Kibler W.B. (1995) Specificity and sensitivity of the anterior slide test in throwing athletes with superior glenoid labral tears. Arthroscopy: The Journal of Arthroscopic and Related Surgery 11, 296-300. [DOI] [PubMed] [Google Scholar]
- Kreighbaum E., Barthels K.M. (1985) Biomechanics - A qualitative approach for studying human movement. 2nd edition Burgess, Minneapolis: 585-616. [Google Scholar]
- Mac Williams B.A., Choi T., Perezous M.K., Chao E.Y., McFarland E.G. (1998) Characteristic ground-reaction forces in baseball pitching. The American Journal of Sports Medicine 26(1), 66-71. [DOI] [PubMed] [Google Scholar]
- Matsuo T., Escamilla R.F., Fleisig G.S., Barrentine S.W., Andrews J.R. (2001) Comparison of kinematic and temporal parameters between different pitch velocity groups. Journal of Applied Biomechanics 17, 1-13. [Google Scholar]
- Milewski M.D., Ounpuu S., Solomito M., Westwell M., Nissen C.W. (2012) Adolescent baseball pitching technique: lower extremity biomechanical analysis. Journal of Applied Biomechanics 28(5), 491-501. [DOI] [PubMed] [Google Scholar]
- Nakamura Y., Yamane K., Fujita Y., Suzuki I. (2005) Somatosensory Computation for man–machine interface from motion-capture data and musculoskeletal human model. IEEE Transactions on Robotics 21(1), 58-66. [Google Scholar]
- Robb A.J., Fleisig G., Wilk K., Macrina L., Bolt B., Pajaczkowski J. (2010) Passive ranges of motion of the hips and their relationship with pitching biomechanics and ball velocity in professional baseball pitchers. American Journal of Sports Medicine 38, 2487-2493. [DOI] [PubMed] [Google Scholar]
- Sakurai S., Ikegami Y., Okamoto A., Yabe K., Toyoshima S. (1993) A three-dimensional cinematographic analysis of upper limb movement during fastball and curveball baseball pitches. Journal of Applied Biomechanics 9, 47-65. [Google Scholar]
- Stodden D.F., Fleisig G.S., McLean S.P., Lyman S.L., Andrews J.R. (2001) Relationship of pelvis and upper torso kinematics to pitched baseball velocity. Journal of Applied Biomechanics 17, 164-172. [DOI] [PubMed] [Google Scholar]