

Abstract
Primates, and particularly humans, are characterized by superior manual dexterity compared with other mammals. However, drawing the biomechanical link between hand morphology/behaviour and functional capabilities in non-human primates and fossil taxa has been challenging. We present a kinematic model of thumb–index precision grip and manipulative movement based on bony hand morphology in a broad sample of extant primates and fossil hominins. The model reveals that both joint mobility and digit proportions (scaled to hand size) are critical for determining precision grip and manipulation potential, but that having either a long thumb or great joint mobility alone does not necessarily yield high precision manipulation. The results suggest even the oldest available fossil hominins may have shared comparable precision grip manipulation with modern humans. In particular, the predicted human-like precision manipulation of Australopithecus afarensis, approximately one million years before the first stone tools, supports controversial archaeological evidence of tool-use in this taxon.
Keywords: grasping, Australopithecus, Neanderthal, manipulation, kinematic model, primates
1. Introduction
Enhanced manual grasping is considered to have been a key adaptation separating the earliest primates from other early mammals [1,2]. This ability continued to evolve among primates to become most refined in humans; coevolving with bipedalism, tool-use, brain enlargement and language [1–7]. Compared with other primates, the anatomy of the human hand helps to facilitate unique abilities, including forceful precision pinch grips between the pad of the thumb and the pads of the fingers and precision handling (manipulating objects within one hand) [3,5]. These dexterous abilities and associated anatomy are traditionally considered to have evolved in response to removing the hands from the constraints of locomotion as well as the mechanical demands of intensive tool-use and tool-production in our fossil hominin ancestors [3,5,8,9]. However, there is still much debate about the inferred manipulative capabilities of early fossil hominins, particularly with regard to tool-use [10–16] and potential subtle differences in precision grip movement or ability between Neanderthals and modern humans [17–19]. Furthermore, other primates, with markedly different hand morphology (e.g. hand proportions, thumb mobility) compared with that of humans, are also capable of using several different precision grips, including tip-to-tip or tip-to-side of the finger precision grips (rather than pad-to-pad as in humans), as well as in-hand movements [20], especially during feeding, tool-use [21–34] or experimental tool-making activities [35,36]. Unlike humans, the hands of other primates must balance the morphological and functional requirements of locomotion, feeding and manipulation, all of which vary tremendously across the order Primates. From an evolutionary standpoint, this prompts several questions about if and how primate grasping is derived from the requirements associated with feeding and/or locomotor behaviour, what grasping abilities are unique to humans and when these evolved, and what morphological constraints might limit precision grasping.
These questions are not fully understood in part because previous studies on primate grasping—and specifically precision grasping—have focused on digit posture (e.g. contact between the hand and the object; [21,22,30]) or morphology (e.g. length of the thumb relative to the fingers; [3–5,9]), both of which have been difficult to link directly to dynamic digit movement. Furthermore, investigation of the evolution of grasping is limited to inferences from bony morphology only, as this is all that is preserved in the fossil record [10,14,37]. Experimental studies, including the kinematics [38], electromyography of muscle use [8] or force experienced by the digits [39] during particular grasping tasks provide important information about the biomechanics of grasping, but may not be logistically or ethically feasible on extant primates and cannot be applied to extinct taxa. A lack of methods enabling the quantification of digit movement and dexterity directly from bony morphology has limited our understanding of evolution and variation of precision manipulation in primates.
Among the wide range of grip types within the primate grasping repertoire, the thumb–index finger pinch is particularly important for increased manual dexterity [3,40]. Nearly all forms of precision grip and many power grips [41] involve thumb–index opposition, making it the foundation of stable human grasping of small objects. Furthermore, thumb–index opposition was critical for tool-related behaviours in human evolutionary history [42] and remains crucial in modern humans for many precision manipulation tasks (e.g. handwriting, fine scraping) and nearly all types of human in-hand manipulation (where a grasped object is positioned by moving the fingers) [43]. It also provides the greatest range of in-hand manipulation workspace (WS) compared with grasps with more than two digits [44].
This study presents a novel method founded in mechanism kinematics that allows us to estimate the precision manipulation capabilities between the thumb (first ray) and index finger (second ray) across a broad sample of extant primates and fossil hominins. Here, we use ‘precision grip/grasp’ to refer to a specific static thumb–index finger posture in which an object is held between the fingertips, whereas ‘precision manipulation’ reflects the active movement of objects held between the fingertips and generally requires complex individual digit control. The kinematic hand model is based on the segment (i.e. metacarpal and phalanges) lengths of the thumb and index finger from associated hand skeletons and inferred or measured mobility (i.e. range of motion) of the first and second ray joints [45] (figure 1). In particular, the model incorporates inferred variation in trapeziometacarpal joint mobility across major primate clades (e.g. opposable versus pseudo-opposable thumbs). The manipulation WS of a given hand is calculated as the area over which the fingers can position a circular object of varying size while satisfying the mechanical constraints of the model (see Material and methods). Thus, WS is a measure of dynamic thumb–index manipulation, providing insights into both the range of locations over which an object can be grasped by the thumb and index finger, as well as the space over which the grasped object can be actively positioned by those digits after being grasped. As such, we consider a hand with a larger thumb–index finger manipulation WS to be one with a greater precision grip potential and overall increased dexterity than a hand with a smaller manipulation WS. Niewoehner et al. [19] used a similar method to investigate precision grip ability in the Neanderthal hand. However, our method differs from this previous study by employing a full kinematic analysis quantifying for the first time the manipulation WS. Furthermore, as our analysis is completely automated, we were able to apply the method to a large comparative sample, rather than qualitatively comparing a Neanderthal to a modern human.
Figure 1.
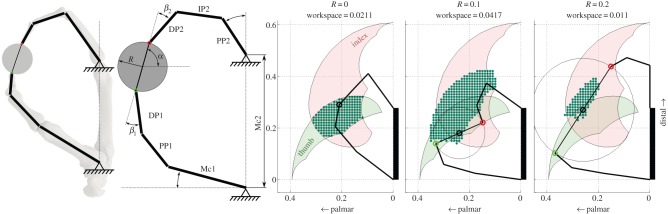
The thumb and index finger kinematic model. The model is based on three thumb segments (first metacarpal (Mc1), proximal phalanx (PP1) and distal phalanx (DP1)) and four index finger segments (Mc2, PP2, second intermediate phalanx (IP2) and DP2) of associated hand specimens. The digits touch a circular object of varying size (R, radius) and rotation (α). The relative orientation of the fingertips, touching two opposing points at the object, is β1 (thumb) and β2 (index finger). The area in which the object can be manipulated is then calculated. The three rightmost pictures show an example workspace (WS) of one representative human for three object sizes. In those pictures, one feasible configuration between the digits and object for each object size is shown. The dots represent the object positions for which a feasible configuration exists and therefore indicate the WS area. The shaded areas represent the positions the tip of thumb (green) and index (red) finger. (Online version in colour.)
We apply this kinematic model to an extant primate sample, including n = 360 associated hand specimens from 38 species of extant hominoids, Old World and New World monkeys and strepsirrhines, and a fossil hominin sample comprising specimens with associated and complete thumb and index fingers that span nearly four million years of hominin evolution. The fossil hominin sample includes the Pliocene fossil hominin Australopithecus afarensis composite hand [10,11], Pleistocene fossil hominins Australopithecus sediba MH2 [14], Homo neanderthalensis Kebara 2 [46], and early Homo sapiens Qafzeh 9 [47] and Ohalo II H2 [48]. Although this kinematic model simplifies the complexity of anatomy and movement in the primate hand, and particularly that of the thumb, this simplicity makes the model applicable to associated fossil specimens for which knowledge about joint movements, soft-tissue anatomy or possibly other bones (e.g. carpals) is unknown. This model offers for the first time a method of assessing manipulative movement within the hand based on bony morphology alone. It can help to establish new links between behavioural studies and morphology, and may reveal potential manipulative capabilities in taxa for which behavioural or biomechanical studies have not yet been done (e.g. most strepsirrhines) or are not possible (i.e. fossil taxa and kinematic studies of most extant non-human primates). Here, we focus on the thumb–index finger movement of several extant primate taxa known for their dexterity (precision grip ability, manipulative skills or tool use) and fossil hominins. The model can be applied to extant and fossil primate and non-primate taxa alike, and can be modified to include different grips or ranges and planes of motion in future biomechanical studies.
Using this kinematic model, we predict that humans will have the largest thumb–index finger WS across all object sizes and that non-human primate taxa considered to be most dexterous in the wild and/or captivity—specifically African apes, baboons, macaques and capuchin monkeys—will have higher WS values than other primates [22,23,25–31]. We expect Pongo to have a different WS from other dexterous species because, although they are also adept tool-users, they have short thumbs and often use a variety of finger-only and non-hand (e.g. mouth or foot) strategies to grasp and use tools [24,30,49]. Among fossil taxa, we hypothesize that Au. afarensis will have a smaller thumb–index finger WS than Au. sediba and later Homo, and that Au. sediba will group with later Homo because of its long thumb [14]. We further predict that H. neanderthalensis will have distinct WS pattern, though not necessarily smaller, compared with early and recent H. sapiens owing to differences in relative segment lengths of the thumb [17].
2. Material and methods
2.1. Extant sample
Our dataset consists of 360 associated hand specimens from 38 primate species (figure 4 and electronic supplementary material, table S2), including hominoids, Old World monkeys, New World monkeys and several species of strepsirrhines (sexes pooled). The human sample includes African, European and small-bodied Khoisan individuals, all of which have very similar relative segment length proportions. Interarticular (IA) length of the first and second metacarpals and maximum length of the proximal, intermediate and distal phalanges of the first and second rays were measured on associated osteological specimens (i.e. instead of using species means for each segment). For part (n = 230) of the sample that did not have IA length for the metacarpals, total length (TL) was used to estimate IA length using regression based on n = 130 specimens (electronic supplementary material, figure S4)
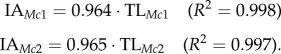 |
Figure 4.

Thumb–index finger proportion and workspace (WS) values for three objects sizes across the comparative extant and fossil primate sample. Left plot shows a ratio of thumb length to index finger length and the right plot shows the WS values for object sizes R = 0 (in which thumb and index finger are touching), R = 0.1 and R = 0.2. The vertical line represents the mean, and the symbols represent the results for each individual for the respective object sizes. The ‘mob’ column indicates mobility of the trapeziometacarpal joint according to Rose [45], where a = 37.6, b = 32.8, c = 21.4, d = 12.5 and e = 10.6°.
The high correlation allowed transformation of TL measurements to IA length without the introduction of significant error. Associated specimens with at least one segment outside a ±2.5σ range were removed from further analyses. This included the removal of five H. sapiens, two P. pygmaeus, two Macaca fascicularis, two Loris tardigradus, two Tarsius syrichta, one Microcebus murinus and one Nycticebus coucang individuals.
2.2. Fossil sample
Thumb and index finger segment lengths were measured directly from the fossils (electronic supplementary material, table S2). The Au. afarensis (3.9–2.9 Ma) composite hand is comprised fossils that are not associated with the same individual [10,11], and thus, there is debate regarding the hand proportions in this taxon and the potential for human-like precision grip [10,11,15,16,50]. However, we include Au. afarensis as an estimate of the potential primitive condition in hominins. Specimens included in the model are: Mc1 AL333w-39, PP1 AL333–69 and DP1 AL33–159 for the thumb, and Mc2 AL333–48, PP2 AL333–93, IP2 AL333–32 and DP2 is a mean length of the only two non-pollical distal phalanges, AL333w-11 and -50.
Australopithecus sediba (1.98 Ma) MH2 is the only (published) almost complete hand of early fossil hominin individual, and its morphology demonstrates a mosaic of primitive, African ape-like features and derived, human-like features [14]. MH2 is missing the distal phalanx of the index finger (DP2). Therefore, two models of Au. sediba were constructed estimating the length of the DP2 based on both a ratio of IP2/DP2 length in Pan troglodytes and in H. sapiens.
2.3. The kinematic model
The thumb and index finger are each modelled as a three link model, such that the basal joint is the trapeziometacarpal joint in the thumb and the metacarpophalangeal (McP) joint in the index finger (figure 1). The model and calculations are done in Matlab (MathWorks Inc., Natick, MA). The segment lengths are derived from the associated osteological hand specimens, but act as kinematic link lengths in the model. Thus, the joint centres are located at the end of the bone in the model, whereas in reality, the joints centres are located in the centre of the epiphyses.
It is assumed that the carpometacarpal joints of the thumb and index finger derive from a single point in morphospace, even though morphologically they are separated (Mc1 articulates with the trapezium, whereas the Mc2 articulates with the trapezoid and capitate in a more distal and medial anatomical position). Thus, the assumed distance between the trapeziometacarpal joint and the McP joint of the index finger is the IA length of the Mc2.
The thumb and index finger are restricted to movement in a single plane, even though the primary plane of movement for both digits is not coplanar. All joints are decoupled (i.e. can function independently), and each joint has one degree of freedom, acting as a hinge joint in flexion and extension only. Thus, abduction and adduction of the thumb and other out-of-the-plane movements are not modelled. Primates vary strongly in the morphology of the trapeziometacarpal joint and range of mobility. Therefore, the limit of movement for this joint follows the group mean estimations of flexion by Rose [45] for Homo (37.6°), great apes (32.8°), cercopithecoids (21.4°), colobines (12.5°), New World monkeys (10.6°) and strepsirrhines (10.6°). The same value was used for all species within the respective groups. Although these values are estimated from the bony morphology [45], they are measured in a variety of different primate species and incorporate important differences in opposable (hominoids and Old World monkeys) and pseudo-opposable (New World monkeys and strepsirrhines) thumb mobility across the breadth of our sample [4]. The limit of movement for all other joints is based by necessity on the maximum range of motion in human joints, because this information is not known for all non-human primates. Maximum flexion values for the index finger are 85.5°, 102° and 72° for the McP, first interphalangeal joint (IPJ), and second IPJ, respectively [51]. In the thumb, maximum flexion of the McP is 59° and the IPJ is 67° [52].
In the kinematic model, the digits move to grip a circular object (figure 2) that varies in size, with a radius (scaled to hand size) of R = 0 (i.e. where the thumb and index finger touch) to R = 0.3. The object sizes are scaled to the size of the hand, such that a radius of 1 is equal to the TL of rays 1 and 2. Only a single point on the fingertip can contact to the object, the digits must oppose each other perfectly on the object (i.e. 180° apart on the object), and the digit and object must be non-intersecting (i.e. collisions limit the range of motion). When both digits are fully extended (i.e. straight), the angle between them is 90°. The relative angle β between the diameter connecting the grasp points and the distal finger segments (DP1, DP2) is restricted to [−90 … 90] degrees.
Figure 2.

Relative segment lengths of the thumb and index finger in extant primates. The sum of all segments within each specimen is scaled to 1. The Pan sample includes both chimpanzees and bonobos combined, as digit proportions were very similar between the two species. The H. sapiens sample includes African, European and small-bodied Khoisan individuals. (Online version in colour.)
Soft tissues and muscle forces are not considered nor does the model take into account muscular and neural couplings between finger joints (which are not known or available for the majority of the specimens considered); this model is purely kinematic. Therefore, whether it is biologically possible for a given hand to apply forces in a particular direction, whereas grasping the object in a given configuration is not tested. However, the simplicity of the model makes it applicable to fossil specimens for which only information about bony morphology is preserved. Human joint ranges of motion were used for all joints of the fossil H. sapiens and H. neanderthalensis specimens, because overall hand morphology is generally very similar [18]. Given the mosaic human-like and Pan-like morphology of Au. afarensis and Au. sediba hands, two models were created for each; one using a Pan joint limit for the trapeziometacarpal joint and human joint limits for the remaining joints, and the other using a human limits for all joints.
2.4. Workspace calculation
To determine the area over which an object can be manipulated, a circular object is placed into the model. The object has two grasping points: index finger (red dot) and thumb (green dot; figure 1). The parameters of the object are the centre location p ∈R2 and orientation (α). Furthermore, for each object position the relative orientation of fingertip and object (β1 and β2) can be varied. For a given object location p, there are infinite object rotations α and hand configurations defined by β1 and β2. In order to count an object position p as valid, only one set of α, β1 and β2 that results in a feasible configuration (where both fingertips are able to touch their respective grasping points and each joint is within its defined limits) is necessary. Multiple solutions for one position will not give an additional benefit in the WS calculation.
For the actual calculation, the plane is discretized into equally spaced points, where the interpoint distance in x- and y-directions is dx. A smaller distance results in a finer grid, which leads to a higher precision of the WS calculation. For the actual WS calculations, dx is set to 0.01. For each location in the WS, the object is rotated between α = 0–360°. In our case, one revolution of the object is discretized into 120 steps. Furthermore, the relative angle between object and finger is varied, βi = −90° to 90°. Those angles are discretized into the same number of bins as α. For each of those configurations (p, α, β1, β2), it is checked whether a feasible finger configuration exists. Overall, the sum of all object positions p, where at least one valid grasp configuration of α and β was found will determine the hand WS.
We look at a large diversity of primates that vary greatly in hand morphology and body mass. To facilitate comparison of hands of widely different sizes, all of the thumb and index finger segment lengths for each individual are normalized to a common length of 1
This is the most straightforward normalization, as it relies only on parameters that are used for modelling the hand, as body mass is not known for most osteological specimens.
3. Results
3.1. Relative segment lengths in extant primates
A key morphology that has been linked to precision grip ability is the length of the thumb relative to the fingers [3,5]. The broad comparative primate sample we have collected here demonstrates substantial variation in thumb-to-index finger length ratio and the relative segment lengths within each ray (figure 2). Within the index finger, most of the variation across primates is in the length of the metacarpal and proximal phalanx, whereas all segments within the thumb vary strongly. Humans are distinct among primates in having the longest thumb relative to their index finger, which facilitates the ability to oppose the pad of the thumb to the distal digits and provides greater control of an object during precision handling [3–5]. However, other primates show a variety of different thumb–index finger proportions to accommodate requirements of locomotion or feeding, including reduced thumb length (e.g. Presbytis), reduced index finger length (e.g. Loris), elongated finger length (e.g. Pan, Hylobates) or reduced metacarpals, but elongated phalanges (e.g. Daubentonia; figure 2 and electronic supplementary material, figure S1) [4,53,54]. The primate taxa documented as being particularly dexterous during feeding or tool-use also vary greatly in their thumb–index finger proportions (figure 2). At one extreme, Pongo and less so Pan have a relatively long index finger and short thumb, whereas at the other extreme, Papio and less so Sapajus, show the opposite proportions with a relatively long thumb and are more similar to H. sapiens in this way. This variation in relative segment lengths has biomechanical consequences for thumb–index finger opposition.
3.2. Thumb–index finger workspace in extant primates
The precision grip WS for each individual and each taxon was calculated for grasping and manipulating a circular object varying in size (scaled to hand size) from R = 0 to 0.3, increasing incrementally by 0.01 (figure 1). The WS for R = 0 corresponds to the area over which the tips of the thumb, and the index finger can touch. As the size of the object increases up to R = 0.3, the WS increases until a point when the object size becomes a constraint on digit movement. The WS for all primates was zero for objects with an R > 0.3 (about the size of a ripe cantaloupe for the average human hand). Figure 1 shows an example WS for a representative human. The general behaviour is similar for all primate taxa, but the y-intersect and the maximum height and position vary for each individual and species.
Figure 3 and table 1 show the average and peak precision manipulation WS relative to object size in a sample of primate taxa that vary substantially in their locomotor and feeding behaviours, relative digit/segment lengths or inferred trapeziometacarpal joint mobility (figures 2 and 4 and electronic supplementary material, figure S2; also see Material and methods). Across all samples, humans have the largest WS for all objects with an R < 0.15 (slightly larger than a tennis ball for the average human hand; figure 3). The peak human WS is substantially higher and at a smaller object size (R = 0.08) than those of all other primates (table 1 and electronic supplementary material, table S1). Within non-human primates, Gorilla have the closest WS to humans, followed by Pan (including both chimpanzees and bonobos), but the peak WS for all African apes is at a much larger object size than that of humans. Papio and Macaca mean and peak WS is broadly similar to that of Pan, both being slightly better at manipulating mid-sized objects but slightly less adept with very small or very large objects, despite a substantially different thumb–index finger ratio (figures 2 and 3) and lower trapeziometacarpal joint mobility (see Material and methods). Sapajus shows a large range of intraspecific variation (as reflected in the segment lengths in figure 2), but its peak WS is for relatively small objects and its overall WS is generally similar to Gorilla for manipulating smaller objects even though Sapajus has more limited thumb mobility. However, Sapajus falls below African apes, Papio and Macaca in manipulating larger objects (R > 0.07). Pongo, with a relatively short thumb, has a much smaller WS than all other taxa, apart from manipulating relatively large objects (R > 0.18; electronic supplementary material, figure S2).
Figure 3.

Workspace (WS) values relative to object size in a sample of extant primates. The bold lines or symbols represent the mean value for each taxon and the lightly shaded areas represent the standard deviation. The maximal WS line represents the highest achievable WS for each object size and was determined by finding the combination of segment lengths that resulted in the highest possible WS. H. sapiens have a much larger WS for smaller objects while Pongo has the smallest WS for most of the object sizes. All taxa generally have a similar WS for objects greater than R = 0.2. (Online version in colour.)
Table 1.
Position of the peak workspace and object size for a sample of extant and fossil primate taxa. The real object size corresponds to the object size that is achieved by scaling the relative object size back to real world units. Australopith ‘human’ and ‘Pan’ refer to use of human versus Pan trapeziometacarpal joint range of motion in two separate kinematic models. In Au. sediba MH2, the length of the DP2 was also inferred using both human and Pan DP2/IP2 proportions (see Material and methods).
| Taxon | peak workspace | relative object size (R) | real object size R (mm) |
|---|---|---|---|
| haplorhines | |||
| Homo sapiens | 0.043 | 0.08 | 19.00 |
| Pan paniscus | 0.029 | 0.11 | 28.70 |
| P. troglodytes | 0.028 | 0.10 | 26.2 |
| Gorilla gorilla | 0.031 | 0.10 | 28.70 |
| Pongo pygmaeus | 0.022 | 0.14 | 37.50 |
| Hylobates lar | 0.027 | 0.11 | 2.20 |
| Papio hamadryas | 0.030 | 0.10 | 15.60 |
| Macaca fascicularis | 0.028 | 0.10 | 10.00 |
| Macaca mulatta | 0.028 | 0.10 | 11.40 |
| Presbytis cristata | 0.016 | 0.16 | 18.00 |
| Sapajus apella | 0.027 | 0.08 | 7.90 |
| Cebus albifrons | 0.029 | 0.08 | 8.70 |
| Alouatta semiculus | 0.029 | 0.08 | 11.50 |
| Tarsius bancanus | 0.023 | 0.11 | 5.80 |
| strepsirrhines | |||
| Lepilemur leucopus | 0.029 | 0.08 | 4.00 |
| Propithecus verreauxi | 0.029 | 0.09 | 9.60 |
| Avahi laniger | 0.024 | 0.11 | 7.40 |
| Euoticus elegantulus | 0.031 | 0.09 | 4.20 |
| Loris tardigradus | 0.029 | 0.09 | 2.90 |
| Nycticebus coucang | 0.030 | 0.09 | 4.00 |
| Daubentonia madagascariensis | 0.029 | 0.08 | 10.00 |
| fossils | |||
| early H. sapiens Qafzeh 9 | 0.044 | 0.08 | 2.90 |
| early H. sapiens Ohalo II H2 | 0.042 | 0.07 | 2.50 |
| H. neanderthalensis Kebara 2 | 0.043 | 0.08 | 3.00 |
| Au. sediba MH2 (human) | 0.044 | 0.08 | 2.10 |
| Au. sediba MH2 (Pan) | 0.040 | 0.09 | 2.40 |
| Au. afarensis comp (human) | 0.042 | 0.08 | 2.40 |
| Au. afarensis comp (Pan) | 0.040 | 0.08 | 2.40 |
Across our primate sample, there are some general patterns in precision manipulation WS despite differences in thumb–index finger proportions and joint mobility (figure 4). Although there is large variation in WS for small object sizes up to about R = 0.1, manipulation WS generally converges across all primates, including humans, for the larger R > 0.15 objects (figure 4). In general, all taxa show their greatest WS with a relative object size of around R = 0.1, whereas manipulation of objects with an R = 0 is similar or smaller to the WS of R = 0.2 objects. The only exceptions to this general primate pattern are humans (and fossil hominins, see below), New World monkeys (Cebus, Alouatta and with some overlap, Sapajus) and aye-ayes (Daubentonia), in which the WS for R = 0 is substantially greater than that of R = 0.2.
These results highlight the enhanced human ability to precision grasp small objects, but also reveal that different combinations of relative thumb–index finger proportions and joint mobility can produce similar manipulation capabilities (figure 4 and electronic supplementary material, figure S2).
3.3. Thumb–index finger proportions and manipulation workspace in fossil hominins
While there are some subtle differences, fossil hominin finger proportions, manipulation WS values all fall within the range of recent H. sapiens variation (figure 4). Australopith hands demonstrate a combination of Pan-like and human-like morphology and thus inferring mobility of the thumb is challenging. Therefore, we created two models each for Au. afarensis and Au. sediba using either a Pan-like or human-like trapeziometacarpal range of motion (see Material and methods). Furthermore, the Au. afarensis composite hand is based on unassociated fossils from multiple individuals, and thus this study presents only one possible estimate of precision manipulation in this taxon. The Au. afarensis thumb–index proportions here are generally similar to those of H. sapiens but with a slightly longer index finger (figure 4; electronic supplementary material, figure S3). In contrast, the associated MH2 hand of Au. sediba demonstrates a relatively long thumb, particularly the first metacarpal, compared with other fossil hominins and recent H. sapiens (figure 4 and electronic supplementary material, figure S3) [14]. Despite this variation in morphology and mobility, the mean manipulation WS for all Au. afarensis and Au. sediba thumb and index finger kinematic models fall within the recent H. sapiens range of variation. Using a Pan-like range of trapeziometacarpal mobility, both australopiths fall slightly below the mean and peak WS of H. sapiens and H. neanderthalensis, especially for small objects (R < 0.1; figure 5 and table 1). When using a human-like range of trapeziometacarpal mobility, Au. afarensis has the same mean and peak manipulation WS as recent H. sapiens, whereas Au. sediba falls out slightly higher than recent H. sapiens and fossil Homo, in particular for object sizes R > 0.08.
Figure 5.

Workspace (WS) values relative to object size in fossil hominins compared with extant Pan and H. sapiens. The bold lines and symbols represent the mean value for each taxon and the lightly shaded areas for Pan and H. sapiens represent the standard deviation. The maximal WS line represents the highest achievable WS for each object size and was determined by finding the combination of segment lengths that resulted in the highest possible WS. All fossil hominins have a larger WS for smaller objects (R < 0.15) compared with Pan. Using Pan-like trapeziometacarpal mobility, Au. afarensis composite hand and Au. sediba have a lower WS than fossil and extant Homo, but with human-like mobility are similar to these taxa. All taxa generally have a similar WS for objects greater than R = 0.15. (Online version in colour.)
Given their generally recent human-like morphology, all H. neanderthalensis and early H. sapiens kinematic models used a recent human range of motion at the trapeziometacarpal joint. However, compared with recent H. sapiens, some fossil Homo specimens demonstrate subtle differences in thumb–index finger proportions (figure 4 and electronic supplementary material, figure S3). H. neanderthalensis demonstrate distinct pollical proportions with a relatively longer pollical distal phalanx but shorter proximal phalanx [17]. Early H. sapiens Qafzeh 9 has a relatively longer thumb than Ohalo II H2, although both fall within the range of variation of recent H. sapiens (figure 4). Despite these differences, all fossil Homo generally have a similar mean and peak manipulation WS for a relatively small object size, although H. neanderthalensis and early H. sapiens Qafzeh 9 have slightly higher WSs for very small objects (R > 0.05; figure 5).
Overall, there is little variation in the potential thumb–index finger WS across australopiths (especially using human range of mobility), fossil Homo, and recent H. sapiens despite subtle differences in thumb–index finger proportions and potential thumb mobility.
4. Discussion
This study models, for the first time, digit movement during precision grasping and manipulation in a broad sample of humans, non-human primates and fossil hominins. Our kinematic thumb–index finger model allows one to assess tip-to-tip precision manipulation potential from the bony morphology of associated hand skeletons. This model is largely based (by necessity of available data) on human kinematics, and thus likely provides the most realistic estimate of digit movement in humans and fossil hominins, while perhaps overestimating or underestimating precision grip potential in non-human primates (see Material and methods). Although the model incorporates variation in thumb mobility (e.g. opposable versus pseudo-opposable thumbs), it does not incorporate the complexity of additional planes of motion (e.g. abduction–adduction), muscle and tendon function, or neurological and myological couplings across segments or digits as these data are unknown for the vast majority of extant primates and are unavailable for fossil specimens. This model also does not take into consideration the associated cognitive or ecological factors that may be required for or promote particular grasping behaviours. The simplicity of the model, however, makes it ideal for comparative analyses across extant and fossil primate and/or non-primate taxa alike, and offers a novel method of assessing how variation in bony morphology can affect digit movement. With further research, the model can be adapted to include different ranges of motion in multiple planes and potentially different grasping behaviours (e.g. pad-to-pad or power grip) between the thumb, index finger or other fingers. Nevertheless, results from the current model provide support for our hypotheses and also reveal several general patterns in primate thumb–index finger precision manipulation potential, despite large variation in hand morphology.
Across our broad sample of primates, the highest manipulation WS for all taxa is for non-zero object sizes when the tip of the thumb and index finger are not touching. The majority of primates demonstrate a similar WS pattern, in which maximal WS is generally between a relative object size of R = 0.05 and 0.15 (figure 4 and electronic supplementary material, figure S2). This similar pattern is also a consequence of the similarity of segment length proportions between primates, where many species have similar ratios between two adjacent segments. This ‘sweet spot’ around the maximal WS is a balance of trade-offs: grasping an object can be thought of as extending the length of the digit by the object radius to increase the WS size, but as the object gets larger relative to the size of the hand, it begins to collide and interfere with digit movement. Across primates, different combinations of thumb–index finger proportions and joint mobility can provide a similar manipulation WS. For example, a relatively short thumb but high trapeziometacarpal joint mobility, as in African apes and Hylobates, yields similar WS values as a relatively long thumb and limited thumb mobility of many Lemuriformes and Lorisiformes [55]. Although the dexterity of great apes has been well documented [20–24,27,32], recent experimental studies suggest that the manipulative abilities in some strepsirrhines may be underestimated [56,57], and our results offer support for future research in these taxa. At the same time, these results also demonstrate that a relatively long thumb—a key morphological feature often linked to precision grip ability in extant and fossil apes [4,5,10,14,58]—does not necessarily translate into greater thumb–index finger manipulation. Although humans and fossil hominins have a higher manipulation WS, especially for smaller objects, several strepsirrhines, including Propithecus and most Lorisiformes, have a similar or higher thumb–index finger ratio compared with humans (owing to a markedly reduced length of the index finger), but substantially lower mean manipulation WS. However, the segment proportions of most non-human primates are at a biomechanical disadvantage relative to the human proportions, such that even with higher human-like trapeziometacarpal joint mobility, their WSs are substantially lower than that of humans. These results highlight the functional importance of joint mobility and a kinematic ‘balance’ in hand proportions.
Results from this kinematic model offer support for most of our hypotheses. First, we predicted that extant humans would have the largest thumb–index finger WS for all object sizes compared with other primates. Humans have a much higher mean and peak WS for small-to-medium objects (i.e. R = 0.0–0.15; R = 0–35 mm for an average human hand size) than all other extant primates. This is consistent with comparative biomechanical and behavioural studies highlighting the enhanced human ability to securely hold and manipulate small objects between the thumb and index finger [3,5,23,30]. However, humans have a slightly lower WS than other dexterous primates for larger objects (i.e. R > 0.15), suggesting that the human hand is particularly well suited to manipulate smaller objects (figure 3 and electronic supplementary material, figure S2). These results thus provide further empirical support for Napier's [3, p. 652] original observation that thumb–index finger precision grips are key to human manipulative abilities. Although our current model does not investigate the manipulative WS of pad-to-pad precision grips that are distinct to humans, it is interesting that even a tip-to-tip precision grip model—a grip that many other primates are capable of using—shows a potential optimization for smaller objects in humans relative to other taxa. As to why the human hand may have evolved to be particularly adept at manipulating small- and medium-sized objects may be related to a particular focus in our early evolutionary history on flake tool-use [12], food foraging or processing of small items similar to that of extant baboons [59], or any number of other manipulative behaviours [11,16]. We predict that modelling the distinctive pad-to-pad precision grip, in which the distal IPJ is extended rather than flexed, will further highlight the enhanced human ability to manipulate small objects in particular compared with other primates.
Second, we predicted that non-human primates traditionally considered more dexterous in the wild and/or captivity would have higher WS values than other non-human primates. This hypothesis is partially supported by the high mean WS in Gorilla and the high WS specifically for small objects in Sapajus (as well as Cebus). Although Gorilla do not regularly use tools in the wild like their chimpanzee cousins, they often use the thumb and index finger to grasp small objects [30,60] and exhibit a larger variety of precision grips during food preparation [27,60,61]. Similarly, Sapajus has evolved myological and neurological features that facilitate digit individualization [62] and a ‘functionally’ opposable thumb [26]. Sapajus exhibits considerable manipulative skills with small food items between the thumb and index finger, compared with the whole-hand grip strategies of other New World monkeys (i.e. Saimiri and Saguinus; [26,63,64]). Although Sapajus is more well known for its nut-cracking activities, these are done with large (i.e. 25% of their body mass) stone anvils that require bimanual power grips rather than precision grasping [65]. Cebus has a similarly high WS for small objects and is also known to manipulate leaves as tools in the wild [66]. However, Alouatta demonstrate similar WS values to Sapajus and Cebus and are not known to have enhanced manipulative skills in the wild or captivity (though this has not yet been studied). Furthermore, the other primates known to be particularly dexterous—Pan, Papio and Macaca—have the same mean and peak manipulation WS as the general ‘primate’ pattern (despite large variation in thumb–index proportions and joint mobility), which does not support our hypothesis that these taxa would have higher thumb–index finger manipulation than other primates. Pongo, as predicted, demonstrates a much lower mean and peak thumb–index finger WS compared with all other primates (except Presbytis) for small- and medium-sized objects (R < 0.18), which may help to explain why they often use within-finger grips or the mouth to manipulate objects [24,49] (electronic supplementary material, figure S2).
Third, we found general support for our predictions among the fossil taxa. Overall, all fossil australopiths and Homo fall within the range of variation in recent H. sapiens manipulation WS. Using either Pan-like or human-like trapeziometacarpal joint mobility, Au. afarensis mean thumb–index finger WS falls at or just slightly below the recent H. sapiens and fossil Homo means (figure 4). This study uses just one possible hand configuration for Au. afarensis with thumb–index finger proportions that are similar to recent H. sapiens. Other research has demonstrated that hand proportions in Au. afarensis may be more similar to Gorilla [15]. Furthermore, Au. afarensis Mc1 and trapezium articular morphology suggests that range of motion at this joint may have been more similar to Pan [50,67] and thus it is possible that Au. afarensis may have had a smaller thumb–index manipulation WS than later hominins and was, in particular, less adept at manipulating small objects.
Using a Pan-like trapeziometacarpal joint mobility, Au. sediba falls below the recent and fossil Homo mean WS despite having a relatively longer thumb (figures 4 and 5; electronic supplementary material, figure S3). Like Au. afarensis, the Pan-like model for Au. sediba has a lower WS for manipulating small objects (R < 0.1), highlighting the importance of thumb mobility (rather than thumb length in this case) for fine-tuned precision manipulation (figure 5). However, the trapeziometacarpal morphology of the Au. sediba Mc1 indicates a larger range of mobility than Au. afarensis [14], suggesting that human-like Au. sediba model may be a better estimate of thumb–index manipulation in this early fossil hominin. Using human-like trapeziometacarpal mobility, Au. sediba has a slightly higher mean WS than recent H. sapiens, but generally has the same manipulation WS as recent and fossil Homo, as predicted. Nevertheless, the fact that the Au. afarensis and Au. sediba thumb–index manipulation potential falls generally within the Homo range of variation is consistent with previous suggestions that both australopiths may have been capable of at least some human-like precision grips [5,14,16,67]. This functional assessment is further supported by archaeological evidence of tool-use in Au. afarensis-bearing deposits at 3.4 Ma [12] and the first recognizable stone tools at approximately 2.6 Ma [68]. These results also suggest that Au. afarensis may have had greater dexterity than is required for cutting with a stone [32], including manipulative and tool-related behaviours that may not preserve in the archaeological record.
Finally, the results do not support for our prediction that H. neanderthalensis (represented here only by Kebara 2) will have distinct, though not necessarily smaller, manipulation WS pattern compared with early and recent H. sapiens owing to differences in relative segment lengths of the thumb [17]. H. neanderthalensis has a similar mean WS and identical peak WS for the same relatively small object size as that of recent H. sapiens (figure 4 and table 1). When WS is viewed across different object sizes, H. neanderthalensis falls above the recent H. sapiens WS range of variation for very small objects (R < 0.05; figure 5). However, early H. sapiens Qafzeh 9 displays a similar WS pattern suggesting that potential increase in thumb–index precision grip of small objects is not related to the distinct H. neanderthalensis thumb proportions. The inclusion of additional associated H. neanderthalensis hand skeletons is needed to test whether there is subtle, yet distinct, pattern in the manipulation WS compared with H. sapiens. For example, Niewoehner et al. [19] found that the thumb–index tip contact ability of Neandertal La Ferrassie I was similar to that of modern humans, but this similarity was not quantified.
These results derived from a broad sample of extant and fossil primates demonstrate that this kinematic model of thumb–index finger precision manipulation based on digit segment lengths provides a useful tool for assessing digit movement. This model provides the first opportunity to assess how variation in bony morphology and joint mobility can affect precision grip and precision manipulation. As such, it is applicable and adaptable to variety of extant and fossil tetrapod taxa for which manual (or pedal) grasping ability is of interest.
Supplementary Material
Supplementary Material
Acknowledgements
We thank Dr Pierre Lemelin for generously sharing his data on extant strepsirrhines. We thank L. Berger, J. Kibii and B. Zipfel and the University of the Witwatersrand, W. Kimbel, G. Semishaw, G. Shimelies and the National Museum of Ethiopia, V. Volpato and Senckenberg Museum Frankfurt, and Y. Rak, I. Hershkovitz, D. Nadel and Tel Aviv University for access to fossil specimens, and F. Mayer, S. Jancke and the Museum für Naturkunde Berlin, C. Boesch, J.-J. Hublin and the Max- Planck-Institut für evolutionäre Anthropologie, S. Kirchengast and the Universität Wien, E. Gilissen, W. Wendelen and Musée royal de L'Afrique central, J. Moggi Cecchi, S. Bortoluzzi and the University of Florence, K. Zyskowski, L. Jones and the Yale Peabody Museum, S. King, S. Matarazzo and the University of Massachussetts Amherst, J. Chupasko and the Harvard Museum of Comparative Zoology, E. Westwig and the American Museum of Natural History, F. Zachos, A. Bibl and the Naturhistorisches Museum Wien for access to comparative extant material.
Data accessibility
The segment measurements that were collected by the authors have been uploaded as part of the electronic supplementary material.
Funding statement
We are grateful for funding from the Gustavus and Louise Pfeiffer Research Foundation and the US National Science Foundation CAREER Award grant IIS-0953856 (AMD), and European Research Council Starting grant no. 336301 and Natural Sciences and Engineering Research Council of Canada (T.L.K.). We also thank the Action Transversale du Muséum National d'Histoire Naturelle: Formes possibles, Formes réalisées and Collections vivantes (Paris, France; E.P.).
Authors' contributions
E.P. and A.M.D. devised the analysis. T.F. and T.L.K. collected segment length data. T.F. created the model and figures. T.F. and T.L.K wrote the manuscript. All authors interpreted the results and edited the manuscript.
Competing interests
The authors declare no competing interests.
References
- 1.Wood Jones F. 1916. Arboreal man. London, UK: Edward Arnold. [Google Scholar]
- 2.Cartmill M. 1974. Rethinking primate origins. Science 184, 436–443. ( 10.1126/science.184.4135.436) [DOI] [PubMed] [Google Scholar]
- 3.Napier JR. 1960. Studies of the hands of living primates. Proc. Zool. Soc. London 134, 647–657. ( 10.1111/j.1469-7998.1960.tb05606.x) [DOI] [Google Scholar]
- 4.Napier JR. 1993. Hands. Princeton, NJ: Princeton University Press. [Google Scholar]
- 5.Marzke MW. 1997. Precision grips, hand morphology, and tools. Am. J. Phys. Anthropol. 102, 91–110. () [DOI] [PubMed] [Google Scholar]
- 6.Ehrsson HH, Fagergren A, Forssberg H. 2001. differential fronto-parietal activation depending on force used in a precision grip task: an fMRI study. J. Neurophysiol. 85, 2613–2623. [DOI] [PubMed] [Google Scholar]
- 7.Christiansen MH, Kirby S. 2003. Language evolution: consensus and controversies. Trends Cogn. Sci. 7, 300–307. ( 10.1016/S1364-6613(03)00136-0) [DOI] [PubMed] [Google Scholar]
- 8.Marzke MW, Toth N, Schick K, Reece S, Steinberg B, Hunt K, Linscheid RL, An KN. 1998. EMG study of hand muscle recruitment during hard hammer percussion manufacture of Oldowan tools. Am. J. Phys. Anthropol. 105, 315–332. () [DOI] [PubMed] [Google Scholar]
- 9.Tocheri MW, Orr CM, Jacofsky MC, Marzke MW. 2008. The evolutionary history of the hominin hand since the last common ancestor of Pan and Homo. J. Anat. 212, 544–562. ( 10.1111/j.1469-7580.2008.00865.x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Marzke MW. 1983. Joint functions and grips of the Australopithecus afarensis hand, with special reference to the region of the capitate. J. Hum. Evol. 12, 197–211. ( 10.1016/S0047-2484(83)80025-6) [DOI] [Google Scholar]
- 11.Alba DM, Moyà-Solà S, Köhler M. 2003. Morphological affinities of the Australopithecus afarensis hand on the basis of manual proportions and relative thumb length. J. Hum. Evol. 44, 225–254. ( 10.1016/S0047-2484(02)00207-5) [DOI] [PubMed] [Google Scholar]
- 12.McPherron SP, Alemseged Z, Marean CW, Wynn JG, Reed D, Geraads D, Bobe R, Béarat HA. 2010. Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ago at Dikika, Ethiopia. Nature 466, 857–860. ( 10.1038/nature09248) [DOI] [PubMed] [Google Scholar]
- 13.Domínguez-Rodrigo M, Pickering TR, Bunn HT. 2011. Reply to McPherron et al.: Doubting Dikika is about data, not paradigms. Proc. Natl Acad. Sci. USA 108, E117 ( 10.1073/pnas.1104647108) [DOI] [Google Scholar]
- 14.Kivell TL, Kibii JM, Churchill SE, Schmid P, Berger LR. 2011. Australopithecus sediba hand demonstrates mosaic evolution of locomotor and manipulative abilities. Science 333, 1411–1417. ( 10.1126/science.1202625) [DOI] [PubMed] [Google Scholar]
- 15.Rolian C, Gordon AD. 2013. Reassessing manual proportions in Australopithecus afarensis. Am. J. Phys. Anthropol. 152, 393–406. ( 10.1002/ajpa.22365) [DOI] [PubMed] [Google Scholar]
- 16.Almécija S, Alba DM. 2014. On manual proportions and pad-to-pad precision grasping in Australopithecus afarensis. J. Hum. Evol. 73, 88–92. ( 10.1016/j.jhevol.2014.02.006) [DOI] [PubMed] [Google Scholar]
- 17.Trinkaus E, Villemeur I. 1991. Mechanical advantages of the Neanderthal thumb in flexion: a test of an hypothesis. Am. J. Phys. Anthropol. 84, 249–260. ( 10.1002/ajpa.1330840303) [DOI] [PubMed] [Google Scholar]
- 18.Niewoehner WA. 2001. Behavioral inferences from the Skhul/Qafzeh early modern human hand remains. Proc. Natl Acad. Sci. USA 98, 2979–2984. ( 10.1073/pnas.041588898) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Niewoehner WA, Bergstrom A, Eichele D, Zuroff M, Clark JT. 2003. Digital analysis: manual dexterity in Neanderthals. Nature 422, 395 ( 10.1038/422395a) [DOI] [PubMed] [Google Scholar]
- 20.Crast J, Fragaszy D, Hayashi M, Matsuzawa T. 2009. Dynamic in-hand movements in adult and young juvenile chimpanzees (Pan troglodytes). Am. J. Phys. Anthropol. 138, 274–285. ( 10.1002/ajpa.20925) [DOI] [PubMed] [Google Scholar]
- 21.Boesch C, Boesch H. 1993. Different hand postures for pounding nuts with natural hammers by wild chimpanzees. In Hands of primates (eds Preuschoft H, Chivers D.), pp. 31–43. Vienna, Australia: Springer. [Google Scholar]
- 22.Christel M. 1993. Grasping techniques and hand preferences in Hominoidea. In Hands of primates (eds Preuschoft H, Chivers D.), pp. 91–108. Vienna, Australia: Springer. [Google Scholar]
- 23.Marzke MW, Wullstein KL. 1996. Chimpanzee and human grips: a new classification with a focus on evolutionary morphology. Int. J. Primatol. 17, 117–139. ( 10.1007/BF02696162) [DOI] [Google Scholar]
- 24.Van Schaik CP, Fox EA, Sitompul AF. 1996. Manufacture and use of tools in wild Sumatran orangutans. Naturwissenschaften 83, 186–188. ( 10.1007/BF01143062) [DOI] [PubMed] [Google Scholar]
- 25.Westergaard GC, Suomi SJ. 1997. Capuchin monkey (Cebus apella) grips for the use of stone tools. Am. J. Phys. Anthropol. 103, 131–135. () [DOI] [PubMed] [Google Scholar]
- 26.Christel MI, Fragaszy D. 2000. Manual function in Cebus apella. Digital mobility, preshaping, and endurance in repetitive grasping. Int. J. Primatol. 21, 697–719. ( 10.1023/A:1005521522418) [DOI] [Google Scholar]
- 27.Byrne RW, Corp N, Byrne JM. 2001. Manual dexterity in the gorilla: bimanual and digit role differentiation in a natural task. Anim. Cogn. 4, 347–361. ( 10.1007/s100710100083) [DOI] [PubMed] [Google Scholar]
- 28.Gumert MD, Kluck M, Malaivijitnond S. 2009. The physical characteristics and usage patterns of stone axe and pounding hammers used by long-tailed macaques in the Andaman Sea region of Thailand. Am. J. Primatol. 71, 594–608. ( 10.1002/ajp.20694) [DOI] [PubMed] [Google Scholar]
- 29.Macfarlane NBW, Graziano MSA. 2009. Diversity of grip in Macaca mulatta. Exp. Brain Res. 197, 255–268. ( 10.1007/s00221-009-1909-z) [DOI] [PubMed] [Google Scholar]
- 30.Pouydebat E, Gorce P, Coppens Y, Bels V. 2009. Biomechanical study of grasping according to the volume of the object: human versus non-human primates. J. Biomech. 42, 266–272. ( 10.1016/j.jbiomech.2008.10.026) [DOI] [PubMed] [Google Scholar]
- 31.Pouydebat E, Reghem E, Borel A, Gorce P. 2011. Diversity of grip in adults and young humans and chimpanzees (Pan troglodytes). Behav. Brain Res. 218, 21–28. ( 10.1016/j.bbr.2010.11.021) [DOI] [PubMed] [Google Scholar]
- 32.Tonooka R, Matsuzawa T. 1995. Hand preferences of captive chimpanzees (Pan troglodytes) in simple reaching for food. Int. J. Primatol. 16, 17–35. ( 10.1007/BF02700151) [DOI] [Google Scholar]
- 33.Haslam M, et al. 2009. Primate archaeology. Nature 460, 339–344. ( 10.1038/nature08188) [DOI] [PubMed] [Google Scholar]
- 34.McGrew WC. 2013. Is primate tool use special? Chimpanzee and New Caledonian crow compared. Phil. Trans. R. Soc. Lond. B 368, 20120422 ( 10.1098/rstb.2012.0422) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Toth N, Schick KD, Savage-Rumbaugh ES, Sevcik RA, Rumbaugh DM. 1993. Pan the tool-maker: investigations into the stone tool-making and tool-using capabilities of a bonobo (Pan paniscus). J. Archaeol. Sci. 20, 81–91. ( 10.1006/jasc.1993.1006) [DOI] [Google Scholar]
- 36.Schick KD, Toth N, Garufi G, Savage-Rumbaugh ES, Rumbaugh D, Sevcik R. 1999. Continuing investigations into the stone tool-making and tool-using capabilities of a bonobo (Pan paniscus). J. Archaeol. Sci. 26, 821–832. ( 10.1006/jasc.1998.0350) [DOI] [Google Scholar]
- 37.Napier J. 1962. The evolution of the hand. Sci. Am. 207, 56–62. ( 10.1038/scientificamerican1262-56) [DOI] [PubMed] [Google Scholar]
- 38.Reghem E, Chèze L, Coppens Y, Pouydebat E. 2013. Unconstrained 3D-kinematics of prehension in five primates: lemur, capuchin, gorilla, chimpanzee, human. J. Hum. Evol. 65, 303–312. ( 10.1016/j.jhevol.2013.06.011) [DOI] [PubMed] [Google Scholar]
- 39.Williams EM, Gordon AD, Richmond BG. 2012. Hand pressure distribution during Oldowan stone tool production. J. Hum. Evol. 62, 520–532. ( 10.1016/j.jhevol.2012.02.005) [DOI] [PubMed] [Google Scholar]
- 40.Landsmeer JM. 1962. Power grip and precision handling. Ann. Rheum. Dis. 21, 164–170. ( 10.1136/ard.21.2.164) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Cutkosky MR. 1989. On grasp choice, grasp models, and the design of hands for manufacturing tasks. Robot. Autom. IEEE Trans. 5, 269–279. ( 10.1109/70.34763) [DOI] [Google Scholar]
- 42.Marzke MW, Shackley MS. 1986. Hominid hand use in the pliocene and pleistocene: evidence from experimental archaeology and comparative morphology. J. Hum. Evol. 15, 439–460. ( 10.1016/S0047-2484(86)80027-6) [DOI] [Google Scholar]
- 43.Bullock IM, Ma RR, Dollar AM. 2013. A hand-centric classification of human and robot dexterous manipulation. Haptics, IEEE Trans. 6, 129–144. ( 10.1109/toh.2012.53) [DOI] [PubMed] [Google Scholar]
- 44.Bullock IM, Feix T, Dollar AM. 2014. Dexterous workspace of human two- and three-fingered precision manipulation. IEEE Haptics Symp., 41–47. ( 10.1109/HAPTICS.2014.6775431) [DOI] [PubMed] [Google Scholar]
- 45.Rose MD. 1992. Kinematics of the trapezium-1st metacarpal joint in extant anthropoids and Miocene hominoids. J. Hum. Evol. 22, 255–266. ( 10.1016/0047-2484(92)90058-h) [DOI] [Google Scholar]
- 46.Arensburg B, et al. 1985. Une sépulture néandertalienne dans la grotte de Kébara (Israël). Comptes-rendus des séances l'Académie des Sci. Série 2, Mécanique-physique, Chim. Sci. l'univers, Sci. la terre 300, 227–230. [Google Scholar]
- 47.Van der Meersch B. 1981. Les hommes fossiles de Gafzeh. Israël: Les hommes fossiles de Gafzeh (Israël). Paris, France: Centre National de la Recherche Scientifique. [Google Scholar]
- 48.Hershkovitz I, Speirs MS, Frayer D, Nadel D, Wish-Baratz S, Arensburg B. 1995. Ohalo II H2: a 19,000-year-old skeleton from a water-logged site at the Sea of Galilee, Israel. Am. J. Phys. Anthropol. 96, 215–234. ( 10.1002/ajpa.1330960302) [DOI] [PubMed] [Google Scholar]
- 49.O'Malley RC, McGrew WC. 2000. Oral tool use by captive orangutans (Pongo pygmaeus). Folia Primatol. (Basel). 71, 334–341. ( 10.1159/000021756) [DOI] [PubMed] [Google Scholar]
- 50.Bush ME, Lovejoy CO, Johanson DC, Coppens Y. 1982. Hominid carpal, metacarpal, and phalangeal bones recovered from the Hadar formation: 1974–1977 collections. Am. J. Phys. Anthropol. 57, 651–677. ( 10.1002/ajpa.1330570410) [DOI] [Google Scholar]
- 51.Mallon WJ, Brown HR, Nunley JA. 1991. Digital ranges of motion: normal values in young adults. J. Hand Surg. Am. 16, 882–887. ( 10.1016/S0363-5023(10)80155-8) [DOI] [PubMed] [Google Scholar]
- 52.Jenkins M, Bamberger H, Black L, Nowinski R. 1998. Thumb joint flexion. What is normal? J. Hand Surg. J. Br. Soc. Surg. Hand 23, 796–797. ( 10.1016/s0266-7681(98)80100-9) [DOI] [PubMed] [Google Scholar]
- 53.Schultz AH. 1936. Characters common to higher primates and characters specific for man. Q. Rev. Biol. 11, 259–283. ( 10.1086/394508) [DOI] [Google Scholar]
- 54.Jouffroy FK, Godinot M, Nakano Y. 1993. Biometrical characteristics of primate hands. In Hands of primates (eds Preuschoft H, Chivers D.), pp. 133–171. Vienna, Austria: Springer. [Google Scholar]
- 55.Bishop A. 1964. Use of the hand in lower primates. In Evolutionary and genetic biology of primates, vol. 2 (ed. Buettner-Janusch J.), pp. 133–225. New York, NY: Academic Press. [Google Scholar]
- 56.Schöneich S. 1993. Hand usage in the ring-tailed lemur (Lemur catta Linnaeus 1758) when solving manipulative tasks. In Hands of primates (eds Preuschoft H, Chivers D.), pp. 7–20. Vienna, Austria: Springer. [Google Scholar]
- 57.Santos LR, Mahajan N, Barnes JL. 2005. How prosimian primates represent tools: experiments with two lemur species (Eulemur fulvus and Lemur catta). J. Comp. Psychol. 119, 394–403. ( 10.1037/0735-7036.119.4.394) [DOI] [PubMed] [Google Scholar]
- 58.Moyà-Solà S, Köhler M, Rook L. 1999. Evidence of hominid-like precision grip capability in the hand of the Miocene ape Oreopithecus. Proc. Natl Acad. Sci. USA 96, 313–317. ( 10.1073/pnas.96.1.313) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Torigoe T. 1985. Comparison of object manipulation among 74 species of non-human primates. Primates 26, 182–194. ( 10.1007/bf02382017) [DOI] [Google Scholar]
- 60.Pouydebat E, Berge C, Gorce P, Coppens Y. 2006. La préhension chez les Primates: précision, outils et perspectives évolutives. C. R. Palevol. 5, 597–602. ( 10.1016/j.crpv.2005.10.011) [DOI] [Google Scholar]
- 61.Neufuss J, Deschner T, Robbins M, Pouydebat E, Kivell T. 2013. Hand use during non-locomotor behaviours in the wild African apes. FOLIA Primatol. 84, 307. [Google Scholar]
- 62.Bortoff G, Strick P. 1993. Corticospinal terminations in two new-world primates: further evidence that corticomotoneuronal connections provide part of the neural substrate for manual dexterity. J. Neurosci. 13, 5105–5118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Costello MB, Fragaszy DM. 1988. Prehension in Cebus and Saimiri: I. Grip type and hand preference. Am. J. Primatol. 15, 235–245. ( 10.1002/ajp.1350150306) [DOI] [PubMed] [Google Scholar]
- 64.Lemelin P, Grafton B. 1998. Grasping performance in Saguinus midas and the evolution of hand prehensility in primates. In Primate locomotion (eds Strasser E, Fleagle J, Rosenberger A, McHenry H.), pp. 131–144. New York, NY: Springer. [Google Scholar]
- 65.Fragaszy DM, Visalberghi E, Fedigan LM. 2004. The complete capuchin: the biology of the genus Cebus. Cambridge, UK: Cambridge University Press. [Google Scholar]
- 66.Phillips KA. 1998. Tool use in wild capuchin monkeys (Cebus albifrons trinitatis). Am. J. Primatol. 46, 259–261. () [DOI] [PubMed] [Google Scholar]
- 67.Tocheri MW, Marzke MW, Liu D, Bae M, Jones GP, Williams RC, Razdan A. 2003. Functional capabilities of modern and fossil hominid hands: three-dimensional analysis of trapezia. Am. J. Phys. Anthropol. 122, 101–112. ( 10.1002/ajpa.10235) [DOI] [PubMed] [Google Scholar]
- 68.Semaw S, Renne P, Harris JWK, Feibel CS, Bernor RL, Fesseha N, Mowbray K. 1997. 2.5-million-year-old stone tools from Gona, Ethiopia. Nature 385, 333–336. ( 10.1038/385333a0) [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The segment measurements that were collected by the authors have been uploaded as part of the electronic supplementary material.