

Abstract
AIM: To investigate the roles and interactions of rho-associated protein kinase (ROCK)1 and miR-124 in human colorectal cancer (CRC).
METHODS: Expression of ROCK1 protein was examined by Western blotting, and quantitative reverse transcriptase PCR was performed to measure expression of ROCK1 mRNA and miR-124. Two cancer cell lines were transfected with pre-miR-124 (mimic) and anti-miR-124 (inhibitor) and the effects on ROCK1 protein and mRNA expression were observed. In addition, cell proliferation was assessed via a 5-ethynyl-2′ deoxyuridine assay. Soft agar formation assay, and cell migration and invasion assays were used to determine the effect of survivin on the transformation and invasion activity of CRC cells.
RESULTS: miR-124 was significantly downregulated in CRC compared to normal specimens (0.603 ± 0.092 vs 1.147 ± 0.286, P = 0.016) and in metastatic compared to nonmetastatic CRC specimens (0.416 ± 0.047 vs 0.696 ± 0.089, P = 0.020). Expression of miR-124 was significantly associated with CRC metastasis, tumor T and N stages, and tumor grade (all P < 0.05). ROCK1 protein was significantly increased in CRC compared to normal tissues (1.896 ± 0.258 vs 0.866 ± 0.136, P = 0.026), whereas ROCK1 mRNA expression was unaltered (2.613 ± 0.251 vs 2.325 ± 0.246). miR-124 and ROCK1 were inversely expressed in CRC tissues and cell lines. ROCK1 mRNA was unaltered in cells transfected with miR-124 mimic and miR-124 inhibitor, compared to normal controls. There was a significant reduction in ROCK1 protein in cells transfected with miR-124 mimic and a significant increase in cells transfected with miR-124 inhibitor (Ps < 0.05). Transformation and invasion of cells transfected with miR-124 inhibitor were significantly increased compared to those in normal controls (P < 0.05). Cells transfected with miR-124 inhibitor showed increased cell proliferation.
CONCLUSION: miR-124 promotes hyperplasia and contributes to invasion of CRC cells, but downregulates ROCK1. ROCK1 and miR-124 may play important roles in CRC.
Keywords: Cell invasion, Colorectal cancer, miR-124, Rho-associated protein kinase
Core tip: miR-124 inhibits neoplastic transformation, cell proliferation, and metastasis, and downregulates rho-associated protein kinase (ROCK)1 in some cancers. In this study, we investigated the roles and interactions of ROCK1 and miR-124 in human colorectal cancer (CRC). miR-124 promoted cell hyperplasia and contributed to invasion, but downregulated ROCK1 in CRC. ROCK1 and miR-124 may play important roles in CRC.
INTRODUCTION
Colorectal cancer (CRC) is one of the most common malignances and the third leading cause of cancer-related death worldwide, with an estimated incidence of 1 million new cases and mortality of > 600000 deaths annually[1,2]. Recent progress in diagnosis and therapy has helped to save the lives of many patients at early stages of this malignancy, but the prognosis for patients with advanced disease or metastasis is still poor[1,2]. Therefore, further investigation into the molecular pathogenesis of CRC and the consequential development of novel targeted therapeutics are needed.
miRNAs are non-protein-coding small RNAs of 19-25 nucleotides that are cleaved from 70-100-nucleotide hairpin pre-miRNA precursors by the enzyme Drosha[3,4]. miRNAs bind to complementary sequences in the 3’-untranslated regions of their target mRNAs and induce mRNA degradation or translational repression[5]. Recent evidence shows that abnormal expression levels of miRNAs are associated with several human cancers, and that they play crucial roles in cell proliferation, differentiation, and apoptosis[6]. Increasing evidence reveals that miRNA dysfunction is associated with several human cancers[7,8]. Volinia et al[9] studied the miRNA expression pattern in solid cancers (2532 samples, 31 cancer types, 120 miRNAs), and found that miR-124 was expressed at a low level in many solid cancers[9-11], and acted as a tumor suppressor[12,13]. However, its role in CRC remains elusive.
Rho-associated protein kinase (ROCK)1 is a member of the rho-associated serine/threonine kinase family, which facilitates reorganization of the actin cytoskeleton during motion[14]. ROCK1 functions as an oncogene, and possesses a wide range of functions, including cellular migration, invasion, and metastasis[15]. ROCK1 is increased in many cancers, including glioma, osteosarcoma, prostate cancer, and gastric cancer[13,16]. ROCK1 is positively correlated with tumor-node-metastasis (TNM) stage and lymph node metastasis in gastric cancer[17]. ROCK1 is targeted by several miRNAs, including miR-135a, miR-145, and miR-148a[12,15,17]. Hu et al[18] found that ROCK1 was a direct target of miR-124 in gastric cancer. However, relatively little is known regarding the underlying mechanisms through which miR-124 regulates ROCK1 in CRC. In this study, we aimed to elucidate the roles and interactions between ROCK1 and miR-124 in human CRC.
MATERIALS AND METHODS
Ethics statement
Tissue specimens (68 tissue pairs) from CRC patients were obtained and histologically confirmed by a pathologist at the Second Affiliated Hospital of Henan University of Traditional Chinese Medicine (Zhengzhou, China). Written informed consent was obtained from all patients and the study was approved by the Ethics Committee of Henan University of Traditional Chinese Medicine on Human Research (No. H20130816).
Cell culture
Three colon cancer cell lines (HCT116, HT29, and SW620) and one human colonic mucosa epithelial cell line (NCM460) were obtained from the Xie He Cell Bank of the Chinese Academy of Medical Sciences (Beijing, China). HCT116 and HT29 cells were cultured in McCoy’s 5A medium (Invitrogen of Thermo Fisher Scientific, Waltham, MA, United States), and SW620 cells were cultured in RPMI 1640 containing 10% fetal bovine serum (FBS; Sigma-Aldrich, St. Louis, MO, United States). All the cells were cultured in a humidified 37 °C incubator supplemented with 5% CO2.
Human samples
Paired resected surgical specimens from primary tumor and adjacent non-tumor sites were obtained from CRC patients [male (n = 47), female (n = 21); median age: 57 years; colon cancer (n = 37) and rectal cancer (n = 31)] who underwent surgery at the Affiliated Hospital of Henan University of Traditional Chinese Medicine, according to a standard protocol, before any therapeutic intervention. Adjacent non-tumor mucosa, ≥ 6 cm from the tumor, was removed. The specimens were snap-frozen in liquid nitrogen and stored at -80 °C for molecular analyses. The remaining tissue specimens were fixed in 10% formalin and embedded in paraffin for routine histologic examination.
Western blotting
Total proteins were extracted from tissues using a total protein extraction kit (Keygen, Nanjing, China). The concentrations of total proteins were measured using a BCA Protein Assay Kit (Keygen). A total of 80 μg protein was separated using SDS-PAGE and transferred onto polyvinylidene difluoride membranes; the membranes were then blocked in 5% fat-free milk at room temperature for 2 h. After incubation with rabbit or goat primary antibodies against ROCK1 (ab80590; Abcam, Cambridge, United Kingdom) at a dilution of 1:10000 or GAPDH (Santa Cruz Biotechnology, Dallas, TX, United States) at a dilution of 1:200 at 4 °C overnight, the membranes were probed with goat anti-rabbit or mouse anti-goat secondary antibodies at a dilution of 1:5000 at room temperature for 2 h. The signals were detected using a Super ECL Plus Kit (Keygen) and determined by quantitative analysis using UVP software (UVP, LLC, Upland, CA, United States). The integral optical density ratio of ROCK1/GAPDH indicated the relative expression of ROCK1 protein.
Total RNA isolation and cDNA synthesis
TRIzol reagent (CWbio, Beijing, China) was used to isolate total RNA from the snap-frozen tissues. The isolated RNA was treated with DNase I (Invitrogen). The RNA concentration and purity were determined using a NanoDrop ND-1000 (NanoDrop Products, Wilmington, DE, United States). The ratio of 28S/18S was analyzed by Glyko Bandscan 5.0. RNA quality and quantity were determined spectrophotometrically at 260 and 280 nm, respectively. Reverse transcription of RNA was performed using the NCode miRNA First-Strand cDNA Synthesis Kit (Invitrogen).
Quantitative reverse transcriptase-PCR
Quantitative reverse transcriptase (qRT)-PCR was performed using the Light Cycler 2.0 Real-Time PCR System (Roche, Penzberg, Germany) in a total volume of 20 μL in glass capillaries containing 2 μL cDNA, 0.8 μL each primer, and 10 μL Light Cycler TaqMan Master Mix (Invitrogen). PCR for miR-124 was initiated using a 10-min denaturation step at 95 °C, followed by termination with a 30-s cooling step at 40 °C. The cycling protocol consisted of denaturation at 95 °C for 15 s and annealing at 60 °C for 1 min for 40 cycles. Fluorescence detection was performed at the end of each step. PCR for ROCK1 was initiated with a 10-min denaturation at 95 °C. Amplification was carried out for 40 cycles of 15 s at 95 °C and 1 min at 60 °C, followed by an extension step of 5 min at 72 °C. All reactions were performed in duplicate. The PCR products were confirmed by melting curve analysis. We used the mathematical delta-delta method (ratio = 2-ΔΔCT) developed by PE Applied Biosystems (Foster City, CA, United States) to compare relative expression between treatments.
RNAi assay
HCT-116 and HT-29 cells were incubated in a six-well tissue culture dish without antibiotics for 24 h prior to transfection, when they had reached 60%-80% confluence. Negative control siRNA, specific miR-124 inhibitor and mimic siRNA (Invitrogen) transfection reagent complexes were mixed with Lipofectamine 2000 (Invitrogen) and then added to the cells. After 6 h at 37 °C, the medium was changed and the cells were cultured in RPMI 1640 supplemented with 10% heat-inactivated FBS. Silencing of miRNA-124 and ROCK1 was determined by qRT-PCR and Western blotting.
5-ethynyl-2′ deoxyuridine (EdU) proliferation assay
Transfected HCT-116 and HT-29 cells were plated in 24-well plates at 4 × 104 cells/well, allowed to adhere, washed with PBS, and incubated for 2 h in serum-free RPMI containing 10 μmol/L EdU (Guangzhou RiboBio Co. Ltd., Guangzhou, China). The cells were washed with PBS, fixed, and permeabilized in PBS containing 2% formaldehyde, 0.5% Triton ×100, and 300 mmol/L sucrose for 15 min. After washing with PBS, cells were blocked using 10% FBS in PBS for 30 min, and incorporated EdU was detected by incubation with a fluorescent azide coupling solution (Apollo; Guangzhou RiboBio Co. Ltd.) for 30 min. The cells were washed three times with PBS containing 0.05% Tween 20, incubated with the DNA staining dye Hoechst 33342 for 30 min, and washed in PBS. Images were captured using a fluorescent microscope, and the average nuclear fluorescent intensity was calculated from at least 50 non-S phase cells randomly selected in five different fields of view.
Soft agar colony-formation assay
A bottom layer (0.6% low-melt agarose) was prepared with RPMI 1640 medium containing 10% FBS, 100 U/mL penicillin, and 100 mg/mL streptomycin. A top layer (0.3% low-melt agarose) was prepared with the same RPMI 1640 medium as described above plus 5000 cells. Plates were incubated at 37.8 °C in 5% CO2 in a humidified incubator for approximately 2 wk. The plates were scanned and photographed, and the number of colonies was quantified using Quantity one version 4.0.3 software (Bio-Rad Laboratories, Hercules, CA, United States).
Migration and invasion assays
Cell migration and invasion assays were performed using a Transwell chamber. For migration, 2 × 104 transfected cells in serum-free medium were plated into the top chamber (BD Biosciences, Franklin Lakes, NJ, United States). For invasion, the same density of cells was seeded into the top chamber, which was precoated with Matrigel (BD Biosciences). After incubation for 24 h, the membranes were fixed and stained with 0.1% crystal violet. Cells passing through the membranes were counted under a microscope (Olympus Corp., Shinjuku, Tokyo, Japan).
Statistical analysis
SPSS version 13.0 software (SPSS Inc., Chicago, IL, United States) was used for the data analysis. Each assay was performed a minimum of three times. The data are expressed as the mean ± SD and Student’s t test and one-way analysis of variance were used to determine the significance of the difference in multiple comparisons. The Mann-Whitney U test was used to determine the associations of miR-124 expression and CRC clinicopathologic features. P < 0.05 indicated statistical significance.
RESULTS
Expression of miR-124 and ROCK1 in tissue specimens and relationship to tumor features
We investigated miR-124 and ROCK1 expression in 68 paired tissue specimens, including 38 cases of nonmetastatic CRC and 30 cases of metastatic CRC. Expression of ROCK1 was significantly increased in CRC compared with noncancerous tissues (1.896 ± 0.258 vs 0.866 ± 0.136, P = 0.026) (Figure 1A and B) and between metastatic tissues and non-metastatic CRC tissues (2.576 ± 0.269 vs 1.566 ± 0.126, P = 0.017) (Figure 1C and D). In contrast, higher expression of miR-124 was found in normal compared with CRC tissues (1.147 ± 0.286 vs 0.603 ± 0.092, P = 0.016), and in nonmetastatic compared to metastatic CRC tissues (0.696 ± 0.089 vs 0.416 ± 0.047, P = 0.020) (Figure 2A and B). For expression of ROCK1 mRNA, there was no difference between the CRC and the normal tissues (2.325 ± 0.246 vs 2.613 ± 0.251) or between metastatic and nonmetastatic CRC tissues (2.723 ± 0.283 vs 2.577 ± 0.276) (Figure 2C and D).
Figure 1.
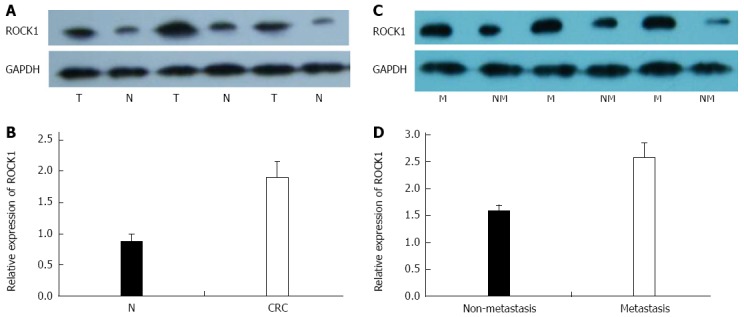
Expression of rho-associated protein kinase 1 protein in 68 colorectal cancer and noncancerous tissue specimens. A: Representative Western blot of rho-associated protein kinase (ROCK)1 expression in colorectal cancer (CRC) (T) and normal (N) tissues; B: Densitometry analysis of ROCK1 expression in N and CRC tissues relative to GAPDH; C: Representative Western blot ROCK1 expression in metastatic (M) and non-metastatic (NM) CRC specimens; D: Densitometry analysis of ROCK1 expression in metastatic and non-metastatic CRC tissues relative to GAPDH. All experiments were repeated three times.
Figure 2.
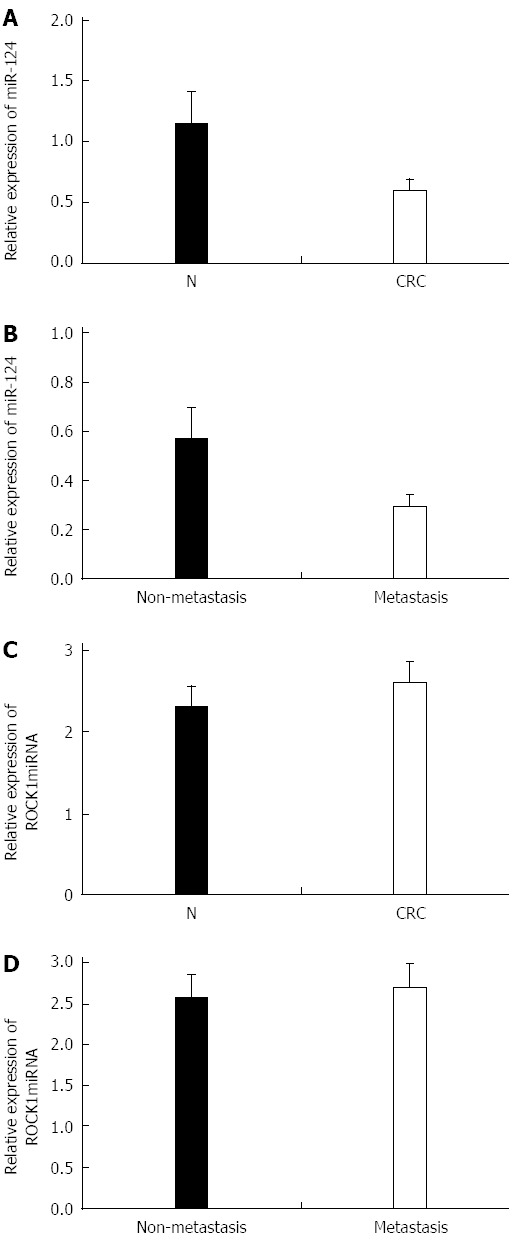
Expression of miR-124 and rho-associated protein kinase 1 mRNA in 68 colorectal cancer and noncancerous tissue specimens. Quantitative real time-PCR data for miR-124 in A: Colorectal cancer (CRC) tissues relative to normal (N) tissues; and B: 30 metastatic and 38 non-metastatic CRC tissues; Quantitative real time-PCR data for rho-associated protein kinase 1 (ROCK1) in C: CRC and N tissues; and D: Metastatic and non-metastatic CRC tissues. All experiments were repeated three times.
The relationships between miR-124 expression and clinicopathologic variables of CRC are summarized in Table 1. There was no significant relationship between miR-124 expression and patient age or sex. However, miR-124 was significantly associated with CRC metastasis, tumor T and N stages, and tumor grade (all P < 0.05).
Table 1.
Associations between clinicopathologic factors and miR-124 expression
| Parameters | n |
miR-124 expression1 |
P value | ||
| Low | Middle | High | |||
| Age | 0.823 | ||||
| ≤ mean | 23 | 11 | 8 | 4 | |
| > mean | 45 | 24 | 16 | 5 | |
| Sex | 0.921 | ||||
| Male | 47 | 24 | 19 | 4 | |
| Female | 21 | 11 | 5 | 5 | |
| T stage2 | 0.022 | ||||
| T1 | 19 | 4 | 12 | 3 | |
| T2 | 17 | 9 | 6 | 2 | |
| T3 | 15 | 10 | 3 | 2 | |
| T4 | 15 | 12 | 2 | 1 | |
| N stage2 | 0.019 | ||||
| N0 | 23 | 6 | 10 | 7 | |
| N1 | 19 | 8 | 7 | 4 | |
| N2 | 17 | 8 | 6 | 3 | |
| N3 | 9 | 6 | 3 | 0 | |
| Metastasis | 0.006 | ||||
| Metastasis | 30 | 22 | 6 | 2 | |
| Non-metastasis | 38 | 14 | 17 | 7 | |
| Grade3 | 0.015 | ||||
| Well differentiated | 26 | 10 | 13 | 3 | |
| Moderately differentiated | 12 | 10 | 9 | 3 | |
| Poorly differentiated | 18 | 15 | 1 | 2 | |
Low, T/N < 1.2; middle, T/N = 1.2-10; high, T/N > 10;
2009 Tumor-Node-Metastasis staging classification system;
Fuhrman classification system: well differentiated = grade 1 and 2; moderately differentiated = grade 3; and poorly differentiated = grade 4.
Expression of miR-124 and ROCK1 in CRC cell lines
We determined the expression of miR-124 and ROCK1 (mRNA and protein) in three CRC cell lines and one normal colorectal cell line (NCM460) (Figure 3). Compared to NCM460 (1.306 ± 0.116), miR-124 expression was significantly reduced in HCT116, HT29, and SW620 cell lines (0.566 ± 0.036, 0.608 ± 0.032, and 0.526 ± 0.028, respectively; all P < 0.01) (Figure 3C). Western blotting showed a higher expression of ROCK1 protein in the three CRC cell lines compared to the NCM460 (HCT116: 1.983 ± 0.0768, HT29: 2.086 ± 0.078, and SW620: 2.226 ± 0.086 vs 0.936 ± 0.066; all P < 0.01) (Figure 3A and B). For ROCK1 mRNA, however, there was no significant difference between NCM460 and cancer cell lines (HCT116: 2.679 ± 0.076, HT29: 2.814 ± 0.086, SW620: 2.703 ± 0.046, NCM460: 2.426 ± 0.056) (Figure 3D).
Figure 3.
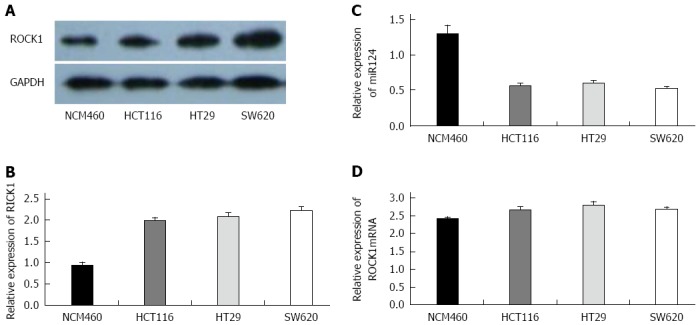
Expression of miR-124 and rho-associated protein kinase 1 in colorectal cancer cell lines. A: Representative Western blot of rho-associated protein kinase 1 (ROCK1) expression in the human colonic mucosa epithelial cell line (NCM460) and colorectal cancer (CRC) cell lines (HCT116, HT29 and SW620); B: Densitometry analysis of ROCK1 expression relative to GAPDH; Quantitative real time-PCR data for C: miR-124; and D: ROCK1 mRNA. All experiments were repeated three times.
miR-124 downregulates ROCK1 protein in cultured HCT116 and HT29cells
We determined whether transfection of HCT116 and HT29 cells with miR-124 mimic or miR-124 inhibitor affected ROCK1 expression. In HCT116 and HT29 cells characterized by low miR-124 expression, downregulation of endogenous miR-124 with miR-124 inhibitor led to a significant increase in ROCK1 protein levels compared to the negative controls (HCT116: 3.356 ± 0.0916 vs 1.956 ± 0.516, HT29: 3.996 ± 0.0981 vs 2.456 ± 0.0706; both P < 0.01) (Figure 4A and B). In contrast, there was a significant decrease in ROCK1 protein levels in cells transfected with miR-124 mimic (HCT116: 0.986 ± 0.0506 vs 1.956 ± 0.516, HT29: 0.916 ± 0.0483 vs 2.456 ± 0.0706; both P < 0.01). For ROCK1 mRNA, there was no difference in cells transfected with miR-124 inhibitor or mimic compare to the negative controls (HCT116: 2.686 ± 0.0515 vs 2.567 ± 0.062, HT29: 3.016 ± 0.0538 vs 3.012 ± 0.072) (Figure 4D).
Figure 4.

In vitro effects of miR-124 regulation. A: Representative Western blot of rho-associated protein kinase 1 (ROCK1) expression in HCT116 and HT29 cells transfected with negative control (NC), miR-124 mimic and miR-124 inhibitor; B: Relative densitometry analysis of ROCK1 expression relative to GAPDH; Quantitative real time-PCR data for C: miR-124; and D: ROCK1 mRNA.
Knockdown of miR-124 induces cell proliferation
We determined the effect of knockdown of miR-124 on cell viability and proliferation using an EdU assay (Figure 4). To confirm the increased number of HCT116 and HT29 cells following treatment with miR-124 inhibitor, cells were labeled with EdU to measure active DNA synthesis and Hoechst 33342 to show all cell nuclei (Figure 4A). According to fluorescent microscopy, the mean percentage of new cells that incorporated EdU was 26.6% and 21.6% in the negative control siRNA group, 41.6% and 40.8% in cells transfected with miR-124 inhibitor (HCT116: 0.416 ± 0.0105 vs 0.266 ± 0.0121, HT29: 0.408 ± 0.0131 vs 0.216 ± 0.0147; both P < 0.01) (Figure 5A and B). We observed that the proliferative ability of HCT116 cells transfected with miR-124 mimic decreased with increasing concentrations of miR-124 mimic from 80 nmol/μL to 120 nmol/μL, with a time of transfection from 1 d to 2 d (P < 0.05) (Figure 5C).
Figure 5.
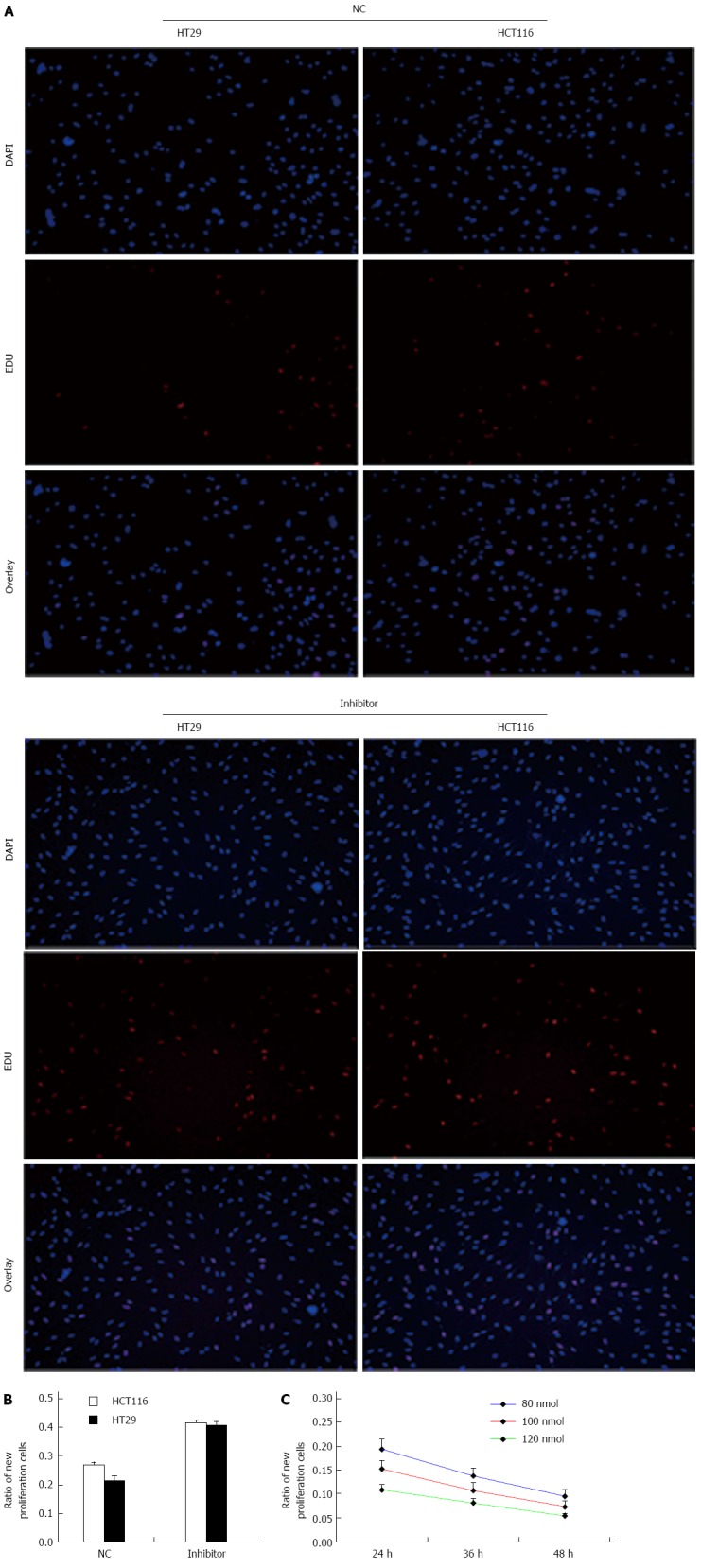
miR-124 knockdown promotes cell proliferation. A: New generation of cells were detected by 5-ethynyl-2’ deoxyuridine (EDU; red), nuclei are stained blue (DAPI); B: Proliferative activity of HCT116 and HT29 cells transfected with negative control (NC) and the miR-124 inhibitor; C: Proliferative ability of HCT16 cells over time when transfected with various doses of miR-124 mimic. All experiments were repeated three times.
miR-124 inhibits metastasis, invasion, and clonogenic survival of CRC cells
To study the role of miR-124 in the regulation of growth and invasion of CRC cells, a colony-formation assay was performed to evaluate whether miR-124 knockdown synergistically promoted HCT116 and HT29 cell transformation (Figure 6A). The assay demonstrated that the number of colonies generated by cells transfected with the miR-124 inhibitor was further increased compared to that generated by the controls (HCT116: 42 ± 3.2 vs 30 ± 2.2, HT29: 37 ± 2.6 vs 24 ± 2, both P < 0.01) (Figure 6A and B).
Figure 6.
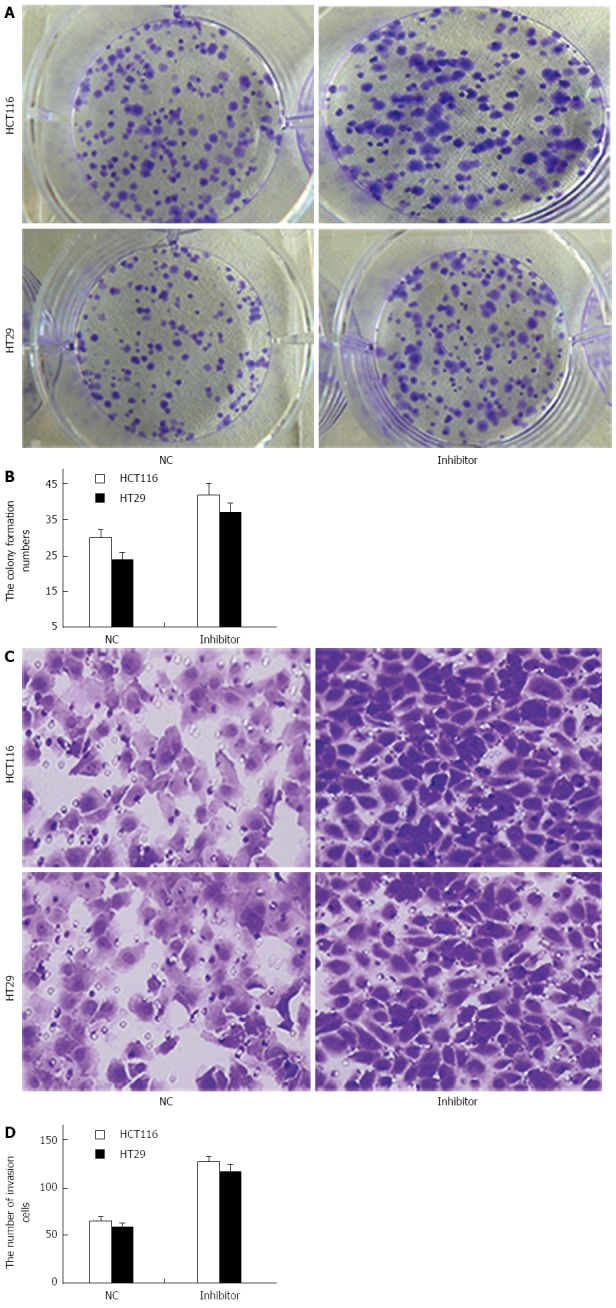
miR-124 inhibits growth and invasion of HCT116 and HT29 cells. A: Colony-formation assay for HCT116 and HT29 cells transfected with negative control (NC) and miR-124 inhibitor was performed on day 14; B: Quantification of colony formation with miR-124 inhibitor; C: Cell migration and invasion assays were performed for HCT116 and HT29 cells transfected with NC and miR-124 inhibitor; D: Quantification of the number of invasive cells with NC and miR-124 inhibitor. All experiments were repeated three times.
Migration and invasion assays showed that downregulation of miR-124 markedly increased proliferation of HCT116 and HT29 cells compare to negative controls (HCT116: 126 ± 7 vs 65 ± 4, HT29: 115 ± 8 vs 58 ± 5, both P < 0.01) (Figure 6C and D).
DISCUSSION
There are few CRC-specific biomarkers that have been used clinically for diagnosis and prediction of prognosis. Although many miRNAs are aberrantly altered in CRC, their underlying molecular mechanisms in the development and progression of CRC remain poorly understood[19]. Lewis et al[20] identified the targets of vertebrate miRNAs using an algorithm called TargetScan. They found that the conserved 5′ region of mammalian miRNAs is most important for target identification. Karginov et al[21] devised a direct biochemical method for miRNA target discovery that combined RNA-induced silencing complex purification with microarray analysis of bound mRNAs. They found that examining the complete spectrum of miR-124 targets in 293 cells yielded a set that were downregulated at the mRNA level, as previously observed, and a set whose mRNA levels were unaffected by miR-124a. Therefore, to explore the function of miRNAs specifically involved in CRC development and progression would greatly help expand our knowledge of CRC, and may provide new targets for its diagnosis and therapy. miR-124 and ROCK1 may not be specific markers for CRC, but they may be used to benefit patients due to their significance in prediction of tumor prognosis. An inverse association between miR-124 and ROCK1 protein was observed in CRC. miR-124 expression was significantly downregulated in CRC tissues and cell lines. In contrast, ROCK1 protein expression was significantly increased, whereas ROCK1 mRNA expression showed consistent results. We observed a significant reduction in ROCK1 protein and mRNA levels in cells transfected with miR-124 mimic, but a significant increase in cells transfected with miR-124 inhibitor. The results suggest that miR-124 post-transcriptionally and negatively regulates ROCK1 by repressing translation in CRC.
As a tumor suppressor, miR-124 is significantly decreased in a variety of human malignancies, including CRC, glioblastoma, hepatocellular carcinoma, medulloblastoma, and gastric cancer[22,23]. Zhao et al[24] found that downregulation of miR-124 promoted malignant progression of glioblastoma cells via increasing expression of a novel protein similar to human protein phosphatase 1, regulatory (inhibitor) subunit 13 like. Xie et al[23] reported that miR-124 inhibited proliferation and induced apoptosis by targeting enhancer of zeste homolog 2 in gastric cancer. In our study, expression of miR-124 was associated with CRC metastasis, supporting our observation that miR-124 inhibits invasion-related processes. Cells transfected with miR-124 inhibitor showed increased cell proliferation and transformation capacity, according to EdU and soft agar formation assays. Therefore, we assume that downregulation of ROCK1 by miR-124 may result in proliferation or metastasis in CRC. Our biologic evidence suggests that targeting miR-124 provides a good strategy to block tumor proliferation and metastasis. From the clinical viewpoint, low expression of miR-124 or high expression of ROCK1 might be considered as a risk factor for tumor progression. Thus, strict systemic therapy after surgery, such as immunotherapy, angiogenesis inhibitors, chemotherapy, and radiotherapy with regular investigation might improve prognosis.
In conclusion, our present study indicates that ROCK1 is negatively regulated at the post-transcriptional level by miR-124, and that miR-124 reduces proliferation and invasion/metastasis in CRC. The results suggest that, as a tumor suppressor, miR-124 may play a role in tumor growth and metastasis. Our findings further suggest that rescue strategies against miR-124, or strategies interfering with the miR-124/ROCK1 interaction, or that inhibit ROCK1 expression, may have potential therapeutic application in CRC in the future.
COMMENTS
Background
miR-124 is decreased in many cancers and acts as a tumor suppressor. Rho-associated protein kinase (ROCK)1 functions as an oncogene and possesses a wide range of functions, including cellular migration, invasion, and metastasis. miR-124 inhibits neoplastic transformation, cell proliferation, and metastasis, and downregulates ROCK1 in gastric cancer.
Research frontiers
There are few colorectal cancer (CRC)-specific biomarkers that have been used clinically for diagnosis and prediction of prognosis. Although many miRNAs are aberrantly altered in CRC, their underlying molecular mechanisms in CRC development and progression remain poorly understood.
Innovations and breakthroughs
In the present study, ROCK1 was a direct target of miR-124 in gastric cancer. However, relatively little is known regarding the underlying mechanisms through which miR-124 regulates ROCK1 in CRC. In this study, the authors elucidated the roles and interactions between ROCK1 and miR-124 in human CRC.
Applications
As a tumor suppressor, miR-124 may play a role in tumor growth and metastasis. The results for miR-124 and ROCK1 suggest that rescue strategies against miR-124, or strategies interfering with the miR-124/ROCK1 interaction, or inhibition of ROCK1 expression, may provide therapeutic potential in CRC in the future.
Terminology
miRNAs are non-protein-coding small RNAs of 19-25 nucleotides that are cleaved from 70-100-nucleotide hairpin pre-miRNA precursors by the enzyme Drosha. ROCK1 is a member of the Rho-associated serine/threonine kinase family, which facilitates reorganization of the actin cytoskeleton during motion.
Peer-review
This is an interesting work about a potential linkage between mir-124 and ROCK1. The authors studied expression of and relationship between miRNA-124 and ROCK1 in colorectal cancer specimens and in normal colorectal cells in vitro.
Footnotes
Ethics approval: The study was approved by the Ethics Committee of Henan University of Traditional Chinese Medicine on Human Research (No. H20130816).
Conflict-of-interest: All authors have no conflict of interest related to the manuscript.
Data sharing: No additional data are available.
Open-Access: This article is an open-access article which was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Peer-review started: November 22, 2014
First decision: December 26, 2014
Article in press: February 11, 2015
P- Reviewer: De Nardi P, Haier J, Linnebacher M S- Editor: Ma YJ L- Editor: AmEditor E- Editor: Ma S
References
- 1.Parkin DM, Bray F, Ferlay J, Pisani P. Global cancer statistics, 2002. CA Cancer J Clin. 2005;55:74–108. doi: 10.3322/canjclin.55.2.74. [DOI] [PubMed] [Google Scholar]
- 2.Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011;61:69–90. doi: 10.3322/caac.20107. [DOI] [PubMed] [Google Scholar]
- 3.Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. doi: 10.1016/s0092-8674(04)00045-5. [DOI] [PubMed] [Google Scholar]
- 4.Zeng Y. Principles of micro-RNA production and maturation. Oncogene. 2006;25:6156–6162. doi: 10.1038/sj.onc.1209908. [DOI] [PubMed] [Google Scholar]
- 5.Engels BM, Hutvagner G. Principles and effects of microRNA-mediated post-transcriptional gene regulation. Oncogene. 2006;25:6163–6169. doi: 10.1038/sj.onc.1209909. [DOI] [PubMed] [Google Scholar]
- 6.Esquela-Kerscher A, Slack FJ. Oncomirs - microRNAs with a role in cancer. Nat Rev Cancer. 2006;6:259–269. doi: 10.1038/nrc1840. [DOI] [PubMed] [Google Scholar]
- 7.Chu D, Zhao Z, Li Y, Li J, Zheng J, Wang W, Zhao Q, Ji G. Increased microRNA-630 expression in gastric cancer is associated with poor overall survival. PLoS One. 2014;9:e90526. doi: 10.1371/journal.pone.0090526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ding H, Wu YL, Wang YX, Zhu FF. Characterization of the microRNA expression profile of cervical squamous cell carcinoma metastases. Asian Pac J Cancer Prev. 2014;15:1675–1679. doi: 10.7314/apjcp.2014.15.4.1675. [DOI] [PubMed] [Google Scholar]
- 9.Volinia S, Galasso M, Costinean S, Tagliavini L, Gamberoni G, Drusco A, Marchesini J, Mascellani N, Sana ME, Abu Jarour R, et al. Reprogramming of miRNA networks in cancer and leukemia. Genome Res. 2010;20:589–599. doi: 10.1101/gr.098046.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lu Z, Liu M, Stribinskis V, Klinge CM, Ramos KS, Colburn NH, Li Y. MicroRNA-21 promotes cell transformation by targeting the programmed cell death 4 gene. Oncogene. 2008;27:4373–4379. doi: 10.1038/onc.2008.72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Medina PP, Slack FJ. microRNAs and cancer: an overview. Cell Cycle. 2008;7:2485–2492. doi: 10.4161/cc.7.16.6453. [DOI] [PubMed] [Google Scholar]
- 12.Li J, Song Y, Wang Y, Luo J, Yu W. MicroRNA-148a suppresses epithelial-to-mesenchymal transition by targeting ROCK1 in non-small cell lung cancer cells. Mol Cell Biochem. 2013;380:277–282. doi: 10.1007/s11010-013-1682-y. [DOI] [PubMed] [Google Scholar]
- 13.Zhang H, Wang Q, Zhao Q, Di W. MiR-124 inhibits the migration and invasion of ovarian cancer cells by targeting SphK1. J Ovarian Res. 2013;6:84. doi: 10.1186/1757-2215-6-84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wen X, Ding L, Wang JJ, Qi M, Hammonds J, Chu H, Chen X, Hunter E, Spearman P. ROCK1 and LIM kinase modulate retrovirus particle release and cell-cell transmission events. J Virol. 2014;88:6906–6921. doi: 10.1128/JVI.00023-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Shin JY, Kim YI, Cho SJ, Lee MK, Kook MC, Lee JH, Lee SS, Ashktorab H, Smoot DT, Ryu KW, et al. MicroRNA 135a suppresses lymph node metastasis through down-regulation of ROCK1 in early gastric cancer. PLoS One. 2014;9:e85205. doi: 10.1371/journal.pone.0085205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Zhou P, Huang G, Zhao Y, Zhong D, Xu Z, Zeng Y, Zhang Y, Li S, He F. MicroRNA-363-mediated downregulation of S1PR1 suppresses the proliferation of hepatocellular carcinoma cells. Cell Signal. 2014;26:1347–1354. doi: 10.1016/j.cellsig.2014.02.020. [DOI] [PubMed] [Google Scholar]
- 17.Wan X, Cheng Q, Peng R, Ma Z, Chen Z, Cao Y, Jiang B. ROCK1, a novel target of miR-145, promotes glioma cell invasion. Mol Med Rep. 2014;9:1877–1882. doi: 10.3892/mmr.2014.1982. [DOI] [PubMed] [Google Scholar]
- 18.Hu CB, Li QL, Hu JF, Zhang Q, Xie JP, Deng L. miR-124 inhibits growth and invasion of gastric cancer by targeting ROCK1. Asian Pac J Cancer Prev. 2014;15:6543–6546. doi: 10.7314/apjcp.2014.15.16.6543. [DOI] [PubMed] [Google Scholar]
- 19.Hong L, Han Y, Yang J, Zhang H, Zhao Q, Wu K, Fan D. MicroRNAs in gastrointestinal cancer: prognostic significance and potential role in chemoresistance. Expert Opin Biol Ther. 2014;14:1103–1111. doi: 10.1517/14712598.2014.907787. [DOI] [PubMed] [Google Scholar]
- 20.Lewis BP, Shih IH, Jones-Rhoades MW, Bartel DP, Burge CB. Prediction of mammalian microRNA targets. Cell. 2003;115:787–798. doi: 10.1016/s0092-8674(03)01018-3. [DOI] [PubMed] [Google Scholar]
- 21.Karginov FV, Conaco C, Xuan Z, Schmidt BH, Parker JS, Mandel G, Hannon GJ. A biochemical approach to identifying microRNA targets. Proc Natl Acad Sci USA. 2007;104:19291–19296. doi: 10.1073/pnas.0709971104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Li KK, Pang JC, Ching AK, Wong CK, Kong X, Wang Y, Zhou L, Chen Z, Ng HK. miR-124 is frequently down-regulated in medulloblastoma and is a negative regulator of SLC16A1. Hum Pathol. 2009;40:1234–1243. doi: 10.1016/j.humpath.2009.02.003. [DOI] [PubMed] [Google Scholar]
- 23.Xie L, Zhang Z, Tan Z, He R, Zeng X, Xie Y, Li S, Tang G, Tang H, He X. MicroRNA-124 inhibits proliferation and induces apoptosis by directly repressing EZH2 in gastric cancer. Mol Cell Biochem. 2014;392:153–159. doi: 10.1007/s11010-014-2028-0. [DOI] [PubMed] [Google Scholar]
- 24.Zhao WH, Wu SQ, Zhang YD. Downregulation of miR-124 promotes the growth and invasiveness of glioblastoma cells involving upregulation of PPP1R13L. Int J Mol Med. 2013;32:101–107. doi: 10.3892/ijmm.2013.1365. [DOI] [PubMed] [Google Scholar]