

Abstract
Lack of vaccine and effective antiviral drugs against chikungunya virus (CHIKV) outbreaks have led to significant impact on health care in the developing world. Here, we evaluated the antiviral effects of tetracycline (TETRA) derivatives and other common antiviral agents against CHIKV. Our results showed that within the TETRA derivatives group, Doxycycline (DOXY) exhibited the highest inhibitory effect against CHIKV replication in Vero cells. On the other hand, in the antiviral group Ribavirin (RIBA) showed higher inhibitory effects against CHIKV replication compared to Aciclovir (ACIC). Interestingly, RIBA inhibitory effects were also higher than all but DOXY within the TETRA derivatives group. Docking studies of DOXY to viral cysteine protease and E2 envelope protein showed non-competitive interaction with docking energy of -6.6±0.1 and -6.4±0.1 kcal/mol respectively. The 50% effective concentration (EC50) of DOXY and RIBA was determined to be 10.95±2.12 μM and 15.51±1.62 μM respectively, while DOXY+RIBA (1:1 combination) showed an EC50 of 4.52±1.42 μM. When compared, DOXY showed higher inhibition of viral infectivity and entry than RIBA. In contrast however, RIBA showed higher inhibition against viral replication in target cells compared to DOXY. Assays using mice as animal models revealed that DOXY+RIBA effectively inhibited CHIKV replication and attenuated its infectivity in vivo. Further experimental and clinical studies are warranted to investigate their potential application for clinical intervention of CHIKV disease.
Introduction
Chikungunya virus (CHIKV) infection is a vector-borne disease transmitted to humans by the bite of female Aedes aegypti mosquitoes. The clinical significance of CHIKV has been emphasized by the epidemic outbreaks across the Indian sub-continent [1,2]. Since 2004, CHIKV-infected cases recorded across ~40 countries have been estimated to be >1.4–6.5 million [2]. Most of the CHIKV outbreaks were reported from tropical countries particularly across Africa, South-East Asia and the Indian Ocean [3]. Aedes albopictus has recently been shown to be a potential vector of CHIKV [4], which likely increases the possibilities of viral distribution across the non-epidemic regions of CHIKV infection such as Europe North America [5]. Very recently in 2014, in Central and South America, 174 cases of locally transmitted disease have been confirmed in El Salvador, Panama, Costa Rica, Venezuela and the Guianas. In addition, four cases of CHIKV infection have been confirmed in southern Florida of United States of America [6].
The lack of anti-viral drugs and vaccines remains a major challenge in the control of CHIKV outbreaks, as current treatment strategies rely on the alleviation of disease symptoms. Common symptoms of CHIKV infection includes high fever (~40°C), headache, back pain, myalgia, vomiting, nausea, conjunctivitis and arthralgia [1]. With the steady increase of CHIKV infection cases in recent years, there is indeed a dire need for newer and effective intervention strategies against this disease. The advent of broad-spectrum antimicrobial agents with minimal side-effects has immensely contributed to the advancement of modern therapeutics in medicine [7,8]. In particular, tetracycline derivatives (TETRA) are commonly used against bacterial infections, although antiviral functions of these antibiotics have not been completely investigated [9,10]. Doxycycline (DOXY), a semi-synthetic tetracycline antibiotic with significant effect against bacterial infections [11–13] works by preventing protein synthesis, and thus expands its usefulness in the clinical treatment of a broad array of bacterial infections [14]. DOXY have been shown to exert inhibitory effects against other microbial infections such as rickettsia, malaria [15] and chlamydia infections [16]. In addition, in vivo studies have indicated that treatment with DOXY attenuates acute lung injury in mice infected with virulent influenza H3N2 virus [17]. Interestingly, synergistic effects of DOXY administered together with oseltamivir provide an effective intervention strategy against swine flu infection [18]. Furthermore, in vitro studies showed that DOXY also exhibited significant inhibitory effects against antimurine retrovirus [19] and dengue virus replication in infected cell lines [20, 21]. Here, we established that a combination of DOXY and RIBA showed potent antiviral activities by inhibiting entry and replication of CHIKV in Vero cells, and reduced viral infectivity in vitro and in vivo.
Methods
Virus and cells
A clinical isolate of CHIKV (East/Central/South African genotype) was recovered from a patient’s serum and used for virus propagation by a single passage in C6/36 mosquito cells. The viral titre (107 p.f.u./ml) of CHIKV suspension was established by serial dilution on Vero cells using a plaque assay. Vero (African green monkey kidney cells, ATCC CCL-81) were obtained from the American Type Culture Collection (ATCC; Rockville, MD, USA) and cultured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum (FBS) as a growth medium or 2% FBS as a maintenance medium. All of the drugs used in this study were purchased from Sigma (Sigma Aldrich) and dissolved in DMSO to a final concentration of less than 1% of the total volume.
Cytotoxicity assay
Vero cells (1 × 104 cells per well of a 96-well plate) were treated with increasing concentrations (0, 6.25, 12.5, 25, 50, 100 μM) of the test compound for 72 h. The cytotoxic effect of the test compounds were determined based on an MTT (3-(4,5-dimethylthiazol 2-yl)-2,5-diphenyltetrazolium bromide) assay as previously reported [21]. The percentage of cell viability was calculated as follows: 100 − (Absorbance of treated cells / Absorbance of untreated cells) × 100.
Evaluation of antiviral activities of tetracycline derivatives
Vero cells were cultured in 24-well plates (1.5 × 105 cells per well) for 24 h at 37°C and 5% CO2. Virus culture supernatant was added to the cells at an MOI of 2, followed by incubation for 2 h with gentle shaking every 15 min to achieve optimal virus-to-cell contact. The cells were washed twice with fresh serum-free DMEM after removing the virus culture supernatant. Subsequently, new complete DMEM containing 15 μM of the test compounds was added to the cultures and incubated for 72 h. The viral titre was evaluated using a plaque formation assay.
Molecular modelling and docking of Doxycycline to CHIKV protease and E2 envelope glycoprotein
The crystal structures for the ligand DOXY (2XRL), the CHIKV protease receptor protein (3TRK) and the E2 envelope glycoprotein (3N41) were obtained from the RCSB Protein Data Bank (PDB) website (http://www.rcsb.org/pdb/home/home.do). The AutoDock Vina [22] molecular docking software was used to investigate the binding of DOXY to CHIKV protease and E2 envelope glycoprotein. The input files for AutoDock Vina were prepared using AutoDock Tools 1.5.6 [23] where a grid box of 30x30x30 Å3 was used by the default settings of 10 docked conformations for each run with the Iterated Local Search global optimizer algorithm. The docking simulation run was repeated three times and all data were presented as mean ± standard error. Lowest docked energy infers the best binding of ligand to receptor and the docked conformation images were produced by PyMOL 1.3 molecular modelling software, as well as for the hydrogen bond analysis.
Viral inactivation assay
We employed a virus inactivation assay to assess the ability of the test compounds to inactivate CHIKV and prevent subsequent infection,. Vero cells were seeded in 24-well microplates (1.5 x 105 cells/well) for 24 hrs. The test compounds (15 μM of DOXY or RIBA and a 15 μM mixture (1:1) of DOXY+RIBA) were mixed with the virus culture supernatant (MOI of 2) and incubated at 37°C for 1 h and then inoculated onto the Vero cells with gentle shaking every 15 min for 2 h. The virus and test compound mix was then removed, and the cells were washed twice with fresh serum-free DMEM to remove any residual virus. New complete DMEM was later added, and the cultures were incubated for 72 h at 37°C supplemented with 5% CO2. The viral titre of the CHIKV suspension was determined through plaque formation assays.
Viral attachment assay
This assay was performed to determine the ability of the test compounds to inhibit virus entry into host cells. Vero cells were grown in 24-well microplates (1.5 x 105 cells/well) for 24 hrs. Cell culture media were removed and the cells were then washed three times with PBS. New media containing virus and test compound mix (concentrations as previously described) were separately added and the cells were incubated for 1 hr at 4°C. The media were later removed and the cells were washed extensively with cold PBS to remove unadsorbed virus. Cells were incubated for 72 h and the viral titre was quantified by plaque formation assays.
Post-infection assay
To investigate the inhibitory effect of the test compounds against CHIKV post-infection, Vero cells were seeded in a 24-well tissue culture plate (1.5 × 105 cells/well), incubated for 24 h under optimal conditions and infected with CHIKV (MOI of 2). Post-infection treatment was performed after inoculating Vero cells with CHIKV for 2 h at 37°C, and subsequently complete DMEM media containing the test compounds (15 μM of DOXY or RIBA and a 15 μM mixture (1:1) of DOXY+RIBA) were added. The cultures were then incubated for 72 h at 37°C and 5% CO2; three wells of infected cells for each experiment were maintained without test compounds as controls and the viral titre was quantified by plaque formation assays. Cell viability was measured after infection and treatment by MTT assay.
Virus quantification by plaque formation assay
A 10-fold serial dilution of culture supernatant of CHIKV- infected cells was added to fresh Vero cells grown in 24-well plates (1.5 × 105 cells) and incubated for 1 h at 37°C. The cells were overlaid with DMEM containing 1.1% methylcellulose. Viral plaques were stained with crystal violet dye after 5-day incubation. Virus titres were calculated according to the following formula: Titre (p.f.u./ml) = number of plaques × volume of the diluted virus added to the well × dilution factor of the virus used to infect the well in which the plaques were enumerated.
ELISA-like cell-based assay
A dose response assay was performed to determine the effect of DOXY, RIBA and DOXY+RIBA on CHIKV replication in Vero cells. The concentrations of test compounds DOXY and RIBA were 0, 5, 10, 15 and 20 μM while the final concentrations of DOXY+RIBA were 0, 5, 10, 25, 50 and 100 μM. The assay was initiated by seeding Vero cells at 1 ×104 cells/well (carried out in triplicate) under optimal conditions (37°C, 5% CO2 in a humidified incubator) in 96-well plates. Virus was mixed with test compounds (MOI of 2) and inoculated to the cells followed by incubation at 37°C for 2 h with gentle shaking every 15 min. At the end of incubation, cells were washed three times with PBS and incubated for 72 h. Culture media was then removed and cells were washed again with PBS before fixing with ice-cold methanol for 15 min at -20°C. After these washing steps, cells were then incubated with blocking buffer containing bovine serum albumin for 1 h at room temperature. CHIKV antibody (Abcam, UK, Cat. no. ab126796) was later added, and cells were left to incubate overnight at 4°C. Following overnight incubation, cells were washed three times with PBS and incubated for 30 min with anti-mouse IgG conjugated with alkaline phosphatase. The cells were then washed again with PBS and the absorbance between 490–650 nm was measured using an ELISA reader. EC50 was calculated using non-linear regression fitting (GraphPad Prism version 5.01).
A similar experimental procedure was carried out for different time points (4, 8, 24, 48 and 72 h) following treatment of infected cells with 15 μM of DOXY or RIBA, or 15 μM of DOXY+RIBA (1:1). The results of ELISA experiments were recorded as mean of the 6-plicate experiments.
Antiviral and toxicity testing in vivo
Animal experiments were carried out in accordance with the University of Malaya guidelines on the Care and Use of Laboratory Animals, and research protocols applied in the investigation had been approved by the Animal Ethics Committee of the University of Malaya. Adult ICR mice were used in acute toxicity and viral infection experiments. Acute toxicity assay was performed by administering the test compounds individually or in combination intraperitoneally to three groups of animals (n = 6 each group). The first two groups of animals were given low dose (5 mg/kg) and high dose (50 mg/kg) respectively, while the third group was kept without treatment The range of the doses was selected based on previous in vivo studies on RIBA [24, 25] and DOXY [26]. The animals were observed for 24 h for signs of toxicity, and after 14 days post-treatment the animals were sacrificed for histological examinations. Viral infection experiments was performed using four groups of animals (n = 8 each). The four groups included positive control (animals were intraperitoneally inoculated with 1 × 106 plaque-forming units (PFU) of the purified CHIKV as described previously [27], RIBA-treated group (infected-animals treated with 15 mg/kg of RIBA), DOXY+RIBA-treated group (infected-animals treated with 15 mg/kg of RIBA and 10 mg/kg of DOXY) and the fourth group was administrated with PBS as a mock-administrated group. Single dose of the test compounds were administrated intraperitonally after 6 h post-infection. Mice were observed for 7 days post-infection and sacrificed for viral quantification and histopathology analysis. Spleen and liver were collected, weighed and fixed in 4% formaldehyde in PBS, and embedded in paraffin using a Leica ASP300 S tissue processor and a Leica EG1160 embedding station (Leica). Sections (4 μm) were stained with hematoxylin–eosin (H&E).
Statistical analysis
All of the assays were performed in triplicate, and the statistical analyses were performed using GraphPad Prism version 5.01 (GraphPad Software, San Diego, CA). P values of <0.05 were considered significant.
Results
Determination of potential cytotoxic effects of antimicrobial agents on Vero cells
We determined the potential cytotoxic effects of antimicrobial agents tested in the current investigation on Vero cells. TETRA derivatives were evaluated for their antiviral properties against CHIKV, and therefore the cytotoxic effects of these compounds were first evaluated. The maximal non-toxic dose (MNTD value) was determined in this study to be approximately 25 μM that showed 80% cell viability. Therefore, all subsequent in vitro cell culture experiments were carried out using doses less than 25 μM for all compounds under study (S1 Fig).
DOXY and RIBA showed increased anti-CHIKV activity by plaque formation assay
The antiviral activity of TETRA derivatives (TETRA, DOXY and ROLI) and the antiviral drugs (ACIC and RIBA) against CHIKV was investigated by plaque formation assay (S2 Fig). Our results showed that the CHIKV load expressed as plaque forming units per ml was significantly reduced following treatment with TETRA derivatives in addition to the antiviral drugs (One Way ANOVA, P<0.01) compared with untreated cells (Fig 1A and 1B). Interestingly, DOXY showed highest inhibitory effect against CHIKV replication in Vero cells (1.50 × 107 p.f.u./ml) compared to the control (6.43 × 107 p.f.u./ml), ROLI (3.76 × 107 p.f.u./ml) and TETRA (2.76 × 107 p.f.u./ml). In addition, RIBA showed higher inhibitory effect (P<0.01) against CHIKV (0.96 × 107 p.f.u./ml) compared to ACIC (2.30 × 107 p.f.u./ml) and other TETRA derivatives except DOXY. Based on these results, we concluded that two antimicrobial agents i.e DOXY and RIBA showed high virus inhibition potential against CHIKV and were therefore selected for further anti-CHIKV investigations.
Fig 1. The inhibitory effect of antimicrobial agents against CHIKV replication in Vero cells.

(A) Viral load was significantly reduced after the treatment with TETRA derivatives and the antiviral drugs compared with the untreated CHIKV-infected cells. DOXY showed highest inhibitory effect against CHIKV replication in Vero cells compared to the control, ROLI and TETRA. RIBA showed higher inhibitory effect (p<0.01) against CHIKV compared to ACIC and other TETRA derivatives except DOXY. (B) The chemical structure of the medical compounds (One way ANOVA, P<0.01).
Docking studies of DOXY to CHIKV cysteine protease and E2 glycoprotein
DOXY showed best binding with the CHIKV cysteine protease (docking energy, -6.6±0.1 kcal/mol), and similar binding was observed with CHIKV E2 glycoprotein (docking energy,-6.4±0.1 kcal/mol) for 10 docked conformations. Since there is no hydrogen bond between DOXY and the viral protease, the interaction is theoretically due to non-polar and van der Waal interactions (Fig 2A). In addition, the predicted binding of DOXY to E2 glycoprotein did not occur at the predicted binding site that involves the residues N207 to N218. DOXY binding with E2 glycoprotein was through the hydrogen bonds that were initiated with L16, P240 and L241 residues of E2 glycoprotein on the basis of most negative docked energy by AutoDock Vina (Fig 2B).
Fig 2. Docking studies of DOXY to CHIKV cysteine protease and E2 glycoprotein.

The input files for AutoDock Vina were prepared using AutoDock Tools 1.5.6 where a grid box of 30x30x30 Å3 was used by the default settings of 10 docked conformations for each run with the Iterated Local Search global optimizer algorithm. (A) DOXY and viral protease binding is due to non-polar and van der Waal interactions. The predicted binding of DOXY to protease was not at the binding site indicating to possible non-competitive inhibition of DOXY against CHIKV protease. (B) Binding of DOXY to viral E2 glycoprotein involved the amino acids L16, P240 and L241 in addition to known E2 binding site to cell receptors (circled).
DOXY+RIBA led to considerable reduction of CHIKV particles in Vero cells
Given that DOXY and RIBA showed increased inhibition potentials against CHIKV in our investigations, we next characterised the antiviral effectiveness of DOXY, RIBA and DOXY+RIBA, against CHIKV using a ELISA-like cell-based assay and fluorescence microscopy following immunostaining. Our results showed that the test compounds inhibited CHIKV replication in Vero cells in a dose-dependent manner (Fig 3A). The 50% effective concentration (EC50) for DOXY was estimated to be 15.51±1.62 μM and for RIBA it was 10.95±2.12 μM, while DOXY+RIBA (1:1) showed an EC50 of 4.52±1.42 μM. Interestingly, DOXY+RIBA showed 95% viral inhibition at 15 μM, even though at same concentration DOXY and RIBA showed only 51% and 68% of viral inhibition respectively (Fig 3A). Infectious particles of CHIKV were reduced after the treatment with DOXY, RIBA and DOXY+RIBA in a time dependent manner with considerable improvement on performance when the test compound were administered together (Fig 3B). Together, we concluded that DOXY+RIBA led to considerable improvement in the reduction of CHIKV particles in Vero cells.
Fig 3. Inhibition of CHIKV after the treatment with DOXY, RIBA and DOXY+RIBA.

(A) ELISA-like cell-based assay was used to evaluate the antiviral activity of the drugs. Viral antigen was detected by CHIKV antibody and anti-mouse IgG conjugated with alkaline phosphatase. The 50% effective concentration (EC50) of the DOXY was calculated to be 10.95±2.12 μM and the RIBA drug was 15.51±1.62 μM, while DOXY+RIBA (1:1) showed an EC50 of 4.52±1.42 μM. (B) Levels of viral proteins after the treatment with drugs were measured by ELISA-like cell-based assay of triplicates at 4, 8, 24, 48 and 72 h post infection. The OD values were measured by an ELISA reader at 490–650 nm. The result showed considerable improvement on performance when the test compound were administered together compared to individual treatment (Two way ANOVA, *P<0.05).
DOXY+RIBA inhibited viral entry of Vero cells
We investigated whether DOXY+RIBA act by inhibiting viral entry of Vero cells. First, we incubated CHIKV with the test compound and then inoculated the cells to examine whether the compounds can inactivate CHIKV and inhibit subsequent events of viral entry (Fig 4A). Treatment of the infected cells with DOXY+RIBA showed the highest inhibitory effect (91%) compared with individual compounds. The difference between DOXY+RIBA and DOXY alone was statistically insignificant (One Way ANOVA, P>0.05); while it was highly significant (One Way ANOVA, P< 0.01) compared to RIBA. In addition, DOXY showed approximately 68% of viral inhibition compared to 45% of RIBA (P<0.05). Similarly, the results of viral attachment assay showed that DOXY+RIBA exhibited the highest inhibition effects against viral infectivity (92%) as represented in Fig 4B. The results showed potential inhibition of DOXY (71%) compared with RIBA (44%) against CHIKV (One Way ANOVA, P<0.05).
Fig 4. Mode of inhibition of DOXY and its combination with RIBA against CHIKV.
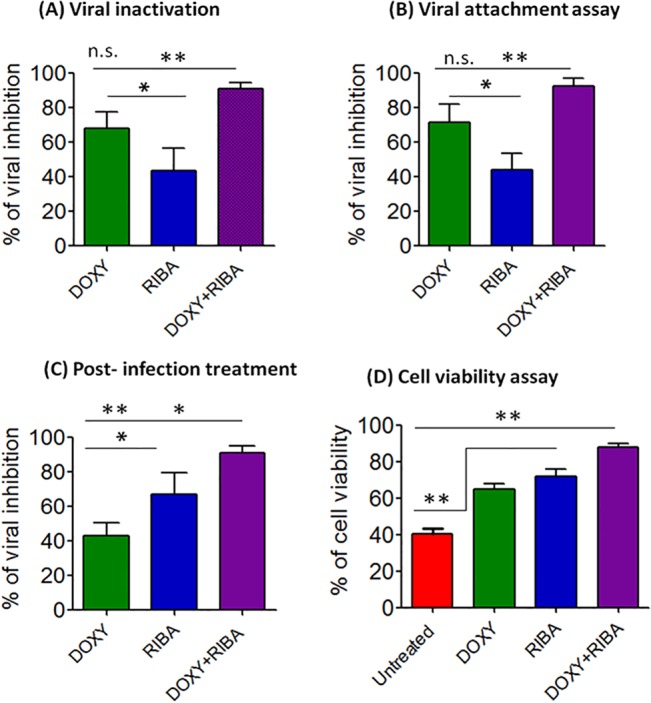
(A) Viral inactivation assay showed significant viral inhibition after the treatment with DOXY compared with RIBA; while DOXY+RIBA showed the highest inhibition percentage. (B) Viral attachment assay showed that DOXY significantly inhibited virus attachment to target cells compared to RIBA and the combination of DOXY+RIBA showed the highest inhibitory effect. (C) Post-infection treatment showed that DOXY inhibitory effect against CHIKV was lower than the inhibitory effect of RIBA. The combination treatment significantly inhibited virus replication compared with the individual treatments. (D) Cell viability of CHIKV-infected cells after treatment with DOXY, RIBA and a DOXY+RIBA compared with untreated cells (One Way ANOVA, *P<0.05, **P<0.01).
Presence of DOXY+RIBA in the culture media significantly reduced CHIKV replication post-infection
We then incubated Vero cells with CHIKV at 37°C for 2 h and subsequently treated the cells with the test compounds to determine their inhibitory effects against viral replication post-infection. The inhibitory effect of DOXY against CHIKV post-infection was lower (43%) than the inhibitory effect showed by RIBA (67%). The presence of the two drugs in the culture media of the infected cells was significantly more effective (One Way ANOVA, P<0.01) in reducing viral replication (91%) compared to individual treatments with DOXY or RIBA (Fig 4C). The MTT assay showed that the viability of CHIKV-infected cells was significantly (One Way ANOVA, P<0.01) improved after treatment with DOXY (65%), RIBA (71.6%) and DOXY+RIBA (88.2%) compared with untreated cells (40.3%) as presented in Fig 4D.
Morphological and histological changes in the liver and spleen of CHIKV-infected mice after the treatment with DOXY+RIBA
Since the in vitro results showed the DOXY+RIBA treatment led to improved inhibition in CHIKV replication, we next set out to examine if DOXY+RIBA treatment have an effect on reducing CHIKV viral load and infectivity in vivo compared to the general antiviral drug RIBA. The experiment of acute toxicity was performed by administering the animals with low (5 mg/kg) and high dose (50 mg/kg) of the drugs, individually or in combination, intraperitoneally. The test compounds at these doses showed no signs of toxicity throughout the 24 h of observation. Histological examinations of liver, spleen and kidney were similar with untreated control group after up to 14 days post-treatment (data not shown).
To observe the effects of drug treatment on cell morphology and histology, CHIKV-infected mice were first grouped into DOXY-, RIBA- and DOXY+RIBA- treated animals besides infected animals without treatment and normal animals as controls. After 7 days post-infection, CHIKV-infected mice usually show liver and spleen hypertrophy (Image A in S3 Fig). The liver of non-treated CHIKV-infected mice were also larger and irregular in shape compared to normal controls. We noticed that treatment with DOXY+RIBA resulted in considerable enhancement in liver size and shape, although these effects were not observed when mice were treated with DOXY or RIBA alone (Image B in S3 Fig). The spleen of [non-treated] CHIKV-infected mice also increased in size compared to normal mice. However, it was noted that DOXY+RIBA treatment markedly reduced the size of the spleen. Again, treatment with either DOXY or RIBA alone showed insignificant effect on spleen size (Image C in S3 Fig). To support these observation, liver and spleen weight was also measured as viral infection is expected to cause considerable weight increase. A similar trend of results were obtained, where treatment with DOXY+RIBA led to a significant reduction in liver (p<0.05) and spleen (p<0.01) weight compared to individual treatment with DOXY or RIBA (Fig 5A and 5B). Quantification of viral titer in the blood of infected mice by plaque formation assay also showed that treatment with DOXY+RIBA exhibited a significantly higher reduction compared to control (ONE WAY ANOVA, P<0.01). We also noted that although treatment with RIBA significantly (ONE WAY ANOVA, P<0.05) reduced viral titer, treatment with DOXY alone showed insignificant reduction in viral load (Fig 5C).
Fig 5. CHIKV-infected mice showed liver and spleen hypertrophy, but reduction in size was evident after treatment.

(A) Treatment with DOXY+RIBA led to significant reduction in liver weight (p<0.05) compared with infected group. (B) Treatment with DOXY+RIBA showed considerable reduction in spleen weight (p<0.01), as evidenced by Kruskal-Wallis test followed by Dunn’s post-test. (C) Viral titer in blood of infected mice was quantified by plaque formation assay (limit of detection was 10 pfu) after 7 days post infection. Although RIBA significantly (One Way ANOVA, P<0.05) reduced viral titer, DOXY+RIBA showed a more prominent reduction compared to control (One Way ANOVA, P<0.01).
The liver of infected mice showed accumulated infiltrated fluid due to inflammation, with high presence of swollen hepatocyte and few shrinking apoptotic cells (Fig 6). DOXY or RIBA-treated mice showed reduction in abnormal cells and inflammation. Treatment with DOXY+RIBA showed lesser amounts of shrinking apoptotic cells while other infection features disappeared. In addition, we note the existence of binucleate hepatocytes suggesting accelerated liver cell renewal. Similarly, the spleen of infected mice showed shrinking apoptotic cells and accumulated infiltrated fluid due to inflammation, and this symptoms were reduced when treated with DOXY+RIBA (Fig 6).
Fig 6. Histopathology of the liver and spleen of CHIKV-infected mice after the treatment with DOXY-RIBA.
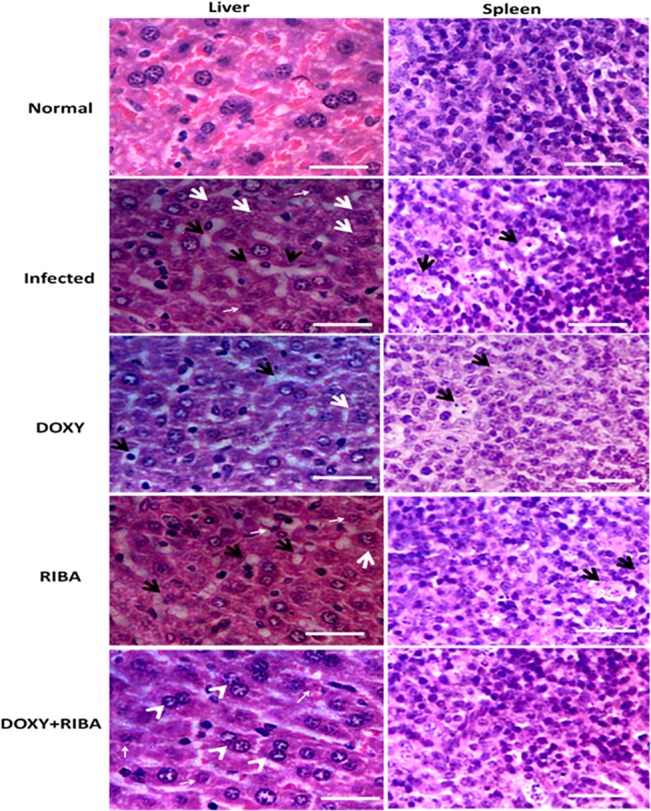
The liver of infected mice showed accumulated infiltrated fluid due to inflammation (black arrows), many swollen hepatocyte (white arrows) and few shrinking apoptotic cells (small arrows). DOXY- or RIBA- treated mice showed reduction in abnormal cells and inflammation. DOXY+RIBA showed lesser shrinking apoptotic cells (small arrows) while other infection features disappeared. The existence of binucleate hepatocytes indicated accelerated liver cell renewal (arrowheads). The spleen of infected mice showed shrinking apoptotic cells and accumulated infiltrated fluid due to inflammation (black arrows), and this symptoms were reduced due to treatment with DOXY+RIBA (scale bars 50 μM).
Discussion
Current treatment strategies against CHIKV infection are aimed at alleviating the symptoms associated with CHIKV disease, and this is largely due to unavailability of effective anti-CHIKV drugs or vaccines for human use. Furthermore, treatment with common antiviral drugs like RIBA or ACIC is almost always ineffective due to unsuitable mechanism of action possessed by these drugs. For instance, it has been recently reported that Hepatitis C virus (HCV) replication in persistently infected cultures induces an autophagy response that impairs RIBA uptake by preventing the expression of equilibrative nucleoside transporter 1 [28]. To increase the effectiveness of this drug, RIBA was administered in combination with other antiviral agents, and results have suggested significant improvement in their antiviral activities against HCV infection, such as pegylated interferon-alpha2a [29, 30], peginterferon alfa-2a and telaprevir [31] and alpha-cyclodextrin [32]. Furthermore, the combination of RIBA with thiosemicarbazone derivative have resulted in considerable success against Japanese Encephalitis Virus [33], whereas a combination of RIBA with cidofovir showed high activity against human adenovirus [34]. Because most of these compounds are relatively expensive, an alternative combination of RIBA with other common antibiotics that showed significant antiviral activities such as DOXY is an attractive alternative. Previous studies have indeed shown that DOXY possessed significant antiviral activity against influenza virus [18], retrovirus [19] dengue virus [20, 21]. To that end, the current study was designed to evaluate the inhibitory effects of TETRA derivatives and certain common antiviral drugs (RIBA and ACIC) against CHIKV infection in vitro and in vivo.
Our observations showed that viral yield was significantly reduced in CHIKV-infected cells after treatment with TETRA derivatives and other antiviral drugs. Of particular note, outstanding inhibitory effects against CHIKV were observed following treatment with DOXY individually or in combination with RIBA. In the present investigation, we determined the inhibitory effects of DOXY against CHIKV as well as its possible effects with the virus life cycle in Vero cells. In parallel with our previous reports [21, 35] the current study showed that TETRA derivatives exhibited similar cytotoxicity effects where the CC50 values were more than 100 μM. On the other hand, both RIBA and ACIC showed lower cytotoxic effects against Vero cells. TETRA derivatives at the doses with more than 80% cell viability showed various levels of the inhibitory effects against CHIKV. Of these, DOXY inhibited CHIKV at the level similar to RIBA. Previous studies have also reported similar antiviral functions of DOXY against various viruses [19–21]. For instance, the antiviral activity of DOXY has been reported against retroviruses where a significant reduction in retrovirus titre was observed after incubation of infected cells with DOXY [19]. Our previous work showed that DOXY inhibits dengue serine protease (NS2B-NS3 protease) and virus entry into the target cells [21]. In this study, when the virus and drug were exposed concurrently on Vero cells, we found that DOXY inhibited CHIKV at EC50 of 10.9 μM, which is lower than RIBA (15.5 μM). RIBA has been shown to possess broad spectrum antiviral effects that act by inhibiting viral replication [36]. Based on this fact we postulated that the inhibition effect of DOXY is directed more towards viral entry rather than viral replication. The combination of DOXY and RIBA showed the highest inhibitory effect (EC50, 4.5 μM). This could be due to the fact that the combination of these two drugs led to the targeting of virus life cycle at both the entry stage (by DOXY) and at the RNA replication stage (by RIBA). Therefore, we performed additional experiments and assays to delineate the antiviral activity of DOXY alone or in combination with RIBA in the aspects of viral infectivity, entry and replication.
Our computational studies showed the predicted binding of DOXY to CHIKV cysteine protease occur at the active site that is important to catalyse viral polyprotein and then viral replication. The active site of CHIKV cysteine protease was reported to involve W1084, C1013 and H1083 residues [37]. Therefore, DOXY may be considered as a non-competitive inhibitor against CHIKV protease. Furthermore, the computationally predicted binding site of CHIKV E2 glycoprotein involves residues N218 to N207 [38]. Although the binding of DOXY with E2 glycoprotein was not at the expected binding site, DOXY showed binding affinity with L16, L241 and P240 through three hydrogen bonds. This binding may impair the important conformational changes of E2 binding site to bind with cell membrane receptors that eventually facilitate virus entry. Previous computational docking studies with dengue virus have indeed shown that DOXY inhibits virus plaque formation by disrupting the conformational changes in the viral envelope that are necessary for virus entry [20]. To verify these findings, further analysis was carried out in vitro to prove that DOXY can inactivate CHIKV and prevent virus entry.
DOXY showed higher potential in reducing viral entry into target cells compared with RIBA. CHIKV was inactivated after 1 h incubation with DOXY, indicating that the drug may have an ability to bind with virus envelop. Similar potential was observed when the virus was incubated with DOXY and cells at 4°C showing that the drug inhibited viral entry into the target cells. Taken together, these results are consistent with our computational data that showed binding affinity between DOXY and CHIKV envelop that may impair important conformational changes of E2 protein during virus entry. Such activity demonstrates the ability of DOXY in inhibiting virus propagation at the cell entry stage. The literature showed broad-spectrum antiviral activity of DOXY [17–20]. Thus, the other mechanism of inhibition that compromise the possible binding of DOXY with specific cell membrane components needs to be considered. Interestingly, post-infection treatment showed low potential effects of DOXY against viral replication in the target cells compared to RIBA, confirming that the main inhibitory effect of DOXY at the entry stage of virus infection.
Our in vivo study showed that treatment with DOXY+RIBA led to significant reduction in virus titre in the blood of ICR mice at 7 days post-infection. In this study we used high dose (106 PFU/mice) of CHIKV to examine the ability of the drugs to combat viral infection. This might result in longer persistence of the virus in the serum of ICR mice. It has been illustrated that mice age, strain and dose of inoculation have an effect on virus persistence post-infection. For example, CHIKV genomic RNA persisted in muscle tissues at least 3 weeks post- infection in 14-day-old C57BL/6J mice [39]; while viral RNA was eliminated after 5–6 days in 6- to 10- weeks mice of the same strain [40]. In addition, C57BL/6 mice showed early clearance of HSV-1 in the brain stem when compared with ICR mice [41]. It has been reported that high inoculation dose has a significant correlation the period and level of the viremia in monkey model [42] and in C57BL/6J mice (3- to 5- weeks old up to 7- to 10- days post- infection) [27]. The results of this study are consistent with previous studies that showed CHIKV persistence in the serum for 7- to 10- days post-infection. [27, 39].
The results of histopathology study indicated that viral infection causes the accumulation of infiltrated fluid due to inflammation, with high presence of swollen hepatocyte and few shrinking apoptotic cells in the liver and spleen. Treatment with DOXY+RIBA led to considerable reduction in pathological signs compared with individual treatment with DOXY or RIBA. Similar studies had shown that combining DOXY with oseltamivir, an antiviral drug, provides an effective treatment for swine origin influenza infected patient [18]. In addition, combining DOXY with monocaprin (1-monoglyceride of capric acid, an anti-herpes- virus drug) provides an effective treatment for herpes labialis, significantly reducing time to healing and pain compared with monocaprin alone [43]. This observation could be attributed to the antiviral activity of DOXY in addition to its feature as an anti-inflammatory drug. It has been reported that DOXY is an anti-inflammatory agent based on its potential inhibition of matrix metalloproteinases such as collagenase, gelatinase and stromelysin, which are part of the inflammatory response and contribute to the breakdown of body tissues [43, 44]. Therefore, in this study, the in vivo investigation showed a convincing outcome of the combined treatment (DOXY+RIBA) compared with RIBA alone, to mice infected with CHIKV. We postulate the anti-CHIKV activity of DOXY+RIBA would potentially inhibit CHIKV and assist in attenuating viral clinical symptoms due to the anti-inflammatory effects of DOXY.
In conclusion, we convincingly showed that DOXY exhibited significant anti-CHIKV activity that acts by impairing viral entry of Vero cells. DOXY+RIBA reduced viral infectivity and replication in infected cells. Further experimental and clinical studies should be carried out to investigate their potential utilization for the attenuation of the clinical symptoms pathognomonic to CHIKV disease.
Supporting Information
Vero cells were treated with increased concentrations (0, 6.25, 12.5, 25, 50, 100 μM) of the drugs for 72 h in 6-plicates for each drug. The maximal non-toxic dose was determined to be approximately 25 μM, that showed 80% cell viability and above. Therefore, all subsequent in vitro cell culture experiments were carried out using doses less than 25 μM for all tested compounds.
(TIF)
Viral load expressed as plaque forming units per ml (p.f.u./ml) was significantly reduced after the treatment with TETRA derivatives and the antiviral drugs compared with the untreated CHIKV-infected cells.
(TIF)
(A) Mice were infected with CHIKV and treated with RIBA alone or in combination with DOXY. After 7 days post-infection, CHIKV-infected mice showed liver and spleen hypertrophy (B) Liver of CHIKV-infected mice was larger and irregular compared to normal. DOXY+RIBA showed considerable enhancement in liver size and shape but not RIBA or DOXY alone. (C) Spleen of CHIKV-infected mice increased in size compared to normal. DOXY+RIBA treatment successfully reduced the size of spleen while treatment with either RIBA or DOXY individually showed insignificant effect on spleen size.
(TIF)
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
Support was provided by the University of Malaya, UMRG Grant RG309-14AFR AND BK-043-2014.
References
- 1. Pialoux G, Gauzere BA, Jaureguiberry S, Strobel M. Chikungunya, an epidemic arbovirosis. Lancet Infect Dis 2007: 7: 319–327. [DOI] [PubMed] [Google Scholar]
- 2. Powers AM, Logue CH. Changing patterns of Chikungunya virus: re-emergence of a zoonotic arbovirus. J Gen Virol 2007: 88: 2363–2377. [DOI] [PubMed] [Google Scholar]
- 3. Staples JE, Breiman RF, Powers AM. Chikungunya fever: an epidemiological review of a re-emerging infectious disease. Clin Infect Dis 2009; 15 (49): 942e8 10.1086/605496 [DOI] [PubMed] [Google Scholar]
- 4. Tsetsarkin KA, Vanlandingham DL, McGee CE, Higgs S. A single mutationin chikungunya virus affects vector specificity and epidemic potential. PLoS Pathog 2007; 3(12): 1895–1906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Caminade C, Medlock JM, Ducheyne E, McIntyre KM, Leach S, Baylis M, et al. Suitability of European climate for the Asian tiger mosquito Aedes albopictus: recent trends and future scenarios. J R Soc 2012; 9(75): 2708–2717. 10.1098/rsif.2012.0138 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Bajak A. US assesses virus of the Caribbean Nat. 2014; 512: 124–125. 10.1038/512124a [DOI] [PubMed] [Google Scholar]
- 7. Braschi I, Blasioli S, Gigli L, Gessa CE, Alberti A, Martucci A. Removal of sulfonamide antibiotics from water: evidence of adsorption into an organophilic zeolite Y by its structural modifications. J. Hazard. Mater 2010; 178: 218–225. 10.1016/j.jhazmat.2010.01.066 [DOI] [PubMed] [Google Scholar]
- 8. Ji LL, Liu FL, Xu ZY, Zheng SR, Zhu DQ. Adsorption of pharmaceutical antibiotics on template-synthesized ordered micro- and mesoporous carbons, Environ. Sci. Technol 2010; 44: 3116–3122. 10.1021/es903716s [DOI] [PubMed] [Google Scholar]
- 9. Homem V, Santos L. Degradation and removal methods of antibiotics from aqueous matrices. J. Environ. Manage 2011; 92: 2304–2347. 10.1016/j.jenvman.2011.05.023 [DOI] [PubMed] [Google Scholar]
- 10. Chao Y, Zhu W, Wu X, Hou F, Xun S, Wu P, et al. Application of graphene-like layered molybdenum disulfide and its excellent adsorption behaviour for Doxycycline antibiotic. Chem Eng J 2011; 243: 60–67. [Google Scholar]
- 11. Zhu J, Liu S, Liu Z, Li Y, Tian J, Hu X. A highly sensitive and selective assay of Doxycycline by dual wavelength overlapping resonance Rayleigh scattering. SPECTROCHIM ACTA A 2014; 124: 237–242. 10.1016/j.saa.2013.12.114 [DOI] [PubMed] [Google Scholar]
- 12. Polat KY, Tosun MS, Ertekin V, Aydinli B, Emre S. Brucella infection with pancytopenia after pediatric liver transplantation. Transpl Infect Dis 2012; 14: 326–329. 10.1111/j.1399-3062.2011.00709.x [DOI] [PubMed] [Google Scholar]
- 13. Waltera MS, Franka MJ, Satué M, Monjo M, Ronold HJ, Lyngstadaas SP, et al. Bioactive implant surface with electrochemically bound Doxycycline promotes bone formation markers in vitro and in vivo. Dent mater 2014; 30: 200–214. 10.1016/j.dental.2013.11.006 [DOI] [PubMed] [Google Scholar]
- 14.Drugs.com, Doxycycline, http://www.drugs.com/pro/doxycycline.html
- 15. Macdougall C, Chambers HF. Protein synthesis inhibitors and miscellaneous antibacterial agents In: Brunton LL, Chabner AB, Knollmann CB, eds. Goodman & Gilman’s The Pharmacological Basis of Therapeutics. 12th ed. New York, NY: McGraw-Hill; 2011; 1537–1538. [Google Scholar]
- 16. Kirchner JT, Hemmert D. Sexually transmitted diseases in women. Chlamydia trachomatis and herpes simplex infections. Postgrad Med 2000; 107(55–58): 55–61. [DOI] [PubMed] [Google Scholar]
- 17. Ng HH, Narasaraju T, Phoon MC, Sim MK, Seet JE, Chow VT. Doxycycline treatment attenuates acute lung injury in mice infected with virulent influenza H3N2 virus: Involvement of matrix metalloproteinases. Exp Mol Pathol 2012; 92: 287–295. 10.1016/j.yexmp.2012.03.003 [DOI] [PubMed] [Google Scholar]
- 18. Quispe-Laime AM, Bracco JD, Barberio PA, Campagne CG, Rolfo VE, Umberger R, et al. H1N1 influenza A virus-associated acute lung injury: response to combination oseltamivir and prolonged corticosteroid treatment. J Intensive Care Med 2010; 36: 33–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Sturtz FG. Antimurine retroviral effect of Doxycycline. Methods Find Exp Clin Pharmacol 1998; 20: 643–647. [DOI] [PubMed] [Google Scholar]
- 20. Yang JM, Chen YF, Tu YY, Yen KR, Yang YL. Combinatorial computational approaches to identify tetracycline derivatives as flavivirus inhibitors. PLoS One 2007; 2: e428 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Rothan HA, Mohamed Z, Paydar M, Rahman NA, Yusof R. Inhibitory effect of Doxycycline against dengue virus replication in vitro. Arch Virol 2013; 10.1007/s00705-013-1880-7 [DOI] [PubMed]
- 22. Trott AO, Olson J. AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading, J Comput Chem 2010; 31: 455–461. 10.1002/jcc.21334 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Michel FS. Python A programming language for software integration and development. J Mol Graphics Mod 1999; 17: 57–61. [PubMed] [Google Scholar]
- 24. Wyde PR, Wilson SZ, Gilbert BE, Smith RH. Protection of mice from lethal influenza virus infection with high dose-short duration RIBA aerosol. Antimicrob Agents Chemother 1986; 30: 942–944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Rowe T, Banner D, Farooqui A, Ng DC, Kelvin AA, Rubino S , et al. In vivo RIBA activity against severe pandemic H1N1 influenza A/Mexico/4108/2009. J Gen Virol 2010; 91: 2898–2906. 10.1099/vir.0.024323-0 [DOI] [PubMed] [Google Scholar]
- 26. Tremblay P, Meiner Z, Galou M, Heinrich C, Petromilli C, Lisse T , et al. Doxycycline control of prion protein transgene expression modulates prion disease in mice. Proc Natl Acad Sci U S A 1998; 13: 12580–12585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Parashar D, Paingankar MS, Kumar S, Gokhale MD, Sudeep AB, Shinde SB , et al. Administration of E2 and NS1 siRNAs Inhibit Chikungunya Virus Replication In Vitro and Protects Mice Infected with the Virus. PLoS Negl Trop Dis 2013; 7: e2405 10.1371/journal.pntd.0002405 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Panigrahi R, Chandra PK, Ferraris P, Kurt R, Song K, Garry RF , et al. Persistent HCV infection impairs ribavirin antiviral activity through clathrin-mediated trafficking of equilibrative nucleoside transporter 1. J Virol 2014; 22: VI.02492-14. 10.1186/1687-9856-2014-22 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Fuster D, Huertas JA, Gomez G, Sola R, Gonzalez Garcia J, Vilaró J, et al. Baseline factors associated with haematological toxicity that leads to a dosage reduction of pegylated interferon-alpha2a and ribavirin in HIV- and HCV-coinfected patients on HCV antiviral therapy. Antivir Ther 2005; 10: 841–847. [PubMed] [Google Scholar]
- 30. Bizollon T, Adham M, Pradat P, Chevallier M, Ducerf C, Baulieux J , et al. Triple antiviral therapy with amantadine for IFN-ribavirin nonresponders with recurrent posttransplantation hepatitis C. Transplantation 2005; 79: 325–329. [DOI] [PubMed] [Google Scholar]
- 31. Lawitz E, Rodriguez-Torres M, Muir AJ, Kieffer TL, McNair L, Khunvichai A, et al. Antiviral effects and safety of telaprevir, peginterferon alfa-2a, and ribavirin for 28 days in hepatitis C patients. J Hepatol 2008; 49: 163–169. 10.1016/j.jhep.2008.03.027 [DOI] [PubMed] [Google Scholar]
- 32. Jeulin H, Grancher N, Kedzierewicz F, Finance C, Le Faou AE, Venard V, et al. In vivo antiviral activity of ribavirin/alpha-cyclodextrin complex: evaluation on experimental measles virus encephalitis in mice. Int J Pharm 2008; 357: 148–153. 10.1016/j.ijpharm.2008.01.043 [DOI] [PubMed] [Google Scholar]
- 33. Sebastian L, Desai A, Yogeeswari P, Sriram D, Madhusudana SN, Ravi V, et al. Combination of N-methylisatin-beta-thiosemicarbazone derivative (SCH16) with ribavirin and mycophenolic acid potentiates the antiviral activity of SCH16 against Japanese encephalitis virus in vitro. Lett Appl Microbiol 2012; 55: 234–239. 10.1111/j.1472-765X.2012.03282.x [DOI] [PubMed] [Google Scholar]
- 34. Darr S, Madisch I, Heim A. Antiviral activity of cidofovir and ribavirin against the new human adenovirus subtype 14a that is associated with severe pneumonia. Clin Infect Dis 2008; 47: 731–732. 10.1086/590970 [DOI] [PubMed] [Google Scholar]
- 35. Rothan HA, Buckle MJ, Ammar YA, Paydar M, Shatrah O, Noorsaadah AR, et al. Study the antiviral activity of some derivatives of tetracycline and non-steroid anti-inflammatory drugs towards dengue virus. Trop Biomed 2013; 30(4): 681–690. [PubMed] [Google Scholar]
- 36. Crotty S, Maag D, Jamie JA, Zhong W, Johnson YN, Hong Z, et al. The broad-spectrum antiviral ribonucleoside RIBA is an RNA virus mutagen. Nat Med 2000; 6: 1375–1379. [DOI] [PubMed] [Google Scholar]
- 37. Pastorino BAM, Peyrefitte CN, Almeras L, Grandadam M, Rolland D, Tolou HJ, et al. Expression and biochemical characterization of nsP2 cysteine protease of Chikungunya virus. Virus Res 2008;13: 293–298 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Asnet MJ, Paramasivan R, Tyagi BK, Surender M, Shenbagarathai R. Identification of structural motifs in the E2 glycoprotein of Chikungunya involved in virus–host interaction. Journal of Biomol Struct Dyn 2013; 31: 1077–1085. 10.1080/07391102.2012.721496 [DOI] [PubMed] [Google Scholar]
- 39. Morrison TE, Oko L, Montgomery SA, Whitmore AC, Lotstein AR, Gunn BM, et al. A mouse model of chikungunya virus-induced musculoskeletal inflammatory disease: evidence of arthritis, tenosynovitis, myositis, and persistence. Am J Pathol 2011; 178(1):32–40. 10.1016/j.ajpath.2010.11.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Poo YS, Nakaya H, Gardner J, Larcher T, Schroder WA, Le TT, et al. CCR2 deficiency promotes exacerbated chronic erosive neutrophil-dominated chikungunya virus arthritis. J Virol. 2014; 88(12):6862–72. 10.1128/JVI.03364-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Yao HW, Ling P, Chen SH, Tung YY, Chen SH. Factors affecting herpes simplex virus reactivation from the explanted mouse brain. Virology. 2012; 10;433(1):116–23. 10.1016/j.virol.2012.07.018 [DOI] [PubMed] [Google Scholar]
- 42. Labadie K, Larcher T, Joubert C, Mannioui A, Delache B, Brochard P, et al. Chikungunya disease in nonhuman primates involves long-term viral persistence in macrophages. J Clin Invest 2010; 120: 894–906 10.1172/JCI40104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Skulason S, Holbrook WP, Thormar H, Gunnarsson GB, Kristmundsdottir T. A study of the clinical activity of a gel combining monocaprin and Doxycycline: a novel treatment for herpes labialis. J Oral Pathol Med 2012; 41:61–67. 10.1111/j.1600-0714.2011.01037.x [DOI] [PubMed] [Google Scholar]
- 44. Manning MW, Cassis LA, Daugherty A. Differential effects of Doxycycline, a broad-spectrum matrix metalloproteinase inhibitor, on angiotensin II-induced atherosclerosis and abdominal aortic aneurysms. Arterioscler Thromb Vasc Biol 2003; 23: 483–488. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Vero cells were treated with increased concentrations (0, 6.25, 12.5, 25, 50, 100 μM) of the drugs for 72 h in 6-plicates for each drug. The maximal non-toxic dose was determined to be approximately 25 μM, that showed 80% cell viability and above. Therefore, all subsequent in vitro cell culture experiments were carried out using doses less than 25 μM for all tested compounds.
(TIF)
Viral load expressed as plaque forming units per ml (p.f.u./ml) was significantly reduced after the treatment with TETRA derivatives and the antiviral drugs compared with the untreated CHIKV-infected cells.
(TIF)
(A) Mice were infected with CHIKV and treated with RIBA alone or in combination with DOXY. After 7 days post-infection, CHIKV-infected mice showed liver and spleen hypertrophy (B) Liver of CHIKV-infected mice was larger and irregular compared to normal. DOXY+RIBA showed considerable enhancement in liver size and shape but not RIBA or DOXY alone. (C) Spleen of CHIKV-infected mice increased in size compared to normal. DOXY+RIBA treatment successfully reduced the size of spleen while treatment with either RIBA or DOXY individually showed insignificant effect on spleen size.
(TIF)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.