

Abstract
The mystacial vibrissae (whiskers) of nocturnal rodents play a major role in their sensorimotor behaviors. Relatively little information exists on the role of whiskers during early development. We characterized the contribution of whiskers to sensorimotor development in postnatal C57BL/6 mice. A comparison between intact and whisker-clipped mice in a battery of behavioral tests from postnatal day (P) 4 to 17 revealed that both male and female pups develop reflexive motor behavior even when the whiskers are clipped. Daily whisker trimming from P3 onwards results in diminished weight gain by P17, and impairment in whisker sensorimotor coordination behaviors, such as cliff avoidance and littermate huddling from P4 through P17, while facilitation of righting reflex at P4 and grasp response at P12. Since active whisker palpation does not start until 2 weeks of age, passive whisker touch during early neonatal stage must play a role in regulating these behaviors. Around the onset of exploratory behaviors (P12) neonatal whisker-clipped pups also display persistent searching movements when they encounter cage walls as a compensatory mechanism of sensorimotor development. Spontaneous whisker motion (whisking) is distinct from respiratory fluttering of whiskers. It is a symmetrical vibration of whiskers at a rate of approximately ∼8 Hz, and begins around P10. Oriented, bundled movements of whiskers at higher frequencies of ∼12 Hz during scanning object surfaces, i.e., palpation whisking, emerges at P14. The establishment of locomotive body coordination before eyes open accompanies palpation whisking, indicating an important role in the guidance of exploratory motor behaviors.
Keywords: somatosensory development, tactile sensation, whisking, sex differences, neonatal motor coordination
Introduction
Nocturnal rodents use their whiskers to scan the environment and navigate in the dark [1,2]. Tactile sense via specialized sensory organ, whiskers, is one of the primary sources for rodents to gather 3D shape and texture information in their world, while olfactory cues are another source to receive social and non-social information, both of which are integrated into nocturnal rodents' behavior [3,4]. The whiskers are arranged in five curvilinear rows bilaterally on the snout. They sweep back and forth during tactile exploration and object palpation [3-5]. Whisker tactile sensation allows the animal to distinguish subtle differences in surface texture and form [6,7] and location (distance) of objects [8-10], throughout their solitary or social life. In the brain, discrete and patterned neural modules represent the whiskers at each synaptic relay of the ascending trigeminal pathway leading to the neocortex [reviewed in 11]. This developmentally formed circuit controls and integrates sensorimotor processes from signals including somatic and tactile sensation to motor coordinative behavior. However, an understanding of the circuit mechanisms by which motor activity integrates somatosensory tactile information is currently limited [eg., 12,13].
“Whisking” is the patterned movement of whiskers by repetitive rapid protraction and retraction during scanning the environment [3,13]. Whiskers are present as small fine hairs on the snout of mice and rats when they are born and develop two distinct type of hairs: longer, prominent hairs formed two-dimensional grid of five rows on the snout (macrovibrissae), and shorter hairs covering the snout and lip surfaces (microvibrissae) [14,15]. Active (scanning) whisking does not emerge until the second postnatal (P) week; whisking gradually increases in frequency and amplitude and reaches adult form by the end of the third week [3,15]. Although whisker movements occur with a characteristic rhythm, movement patterns vary in relation to the texture of the surface the animal explores [6,16] and the way surfaces are contacted with bilateral whisker arrays [17-19]. Lateral asymmetry of whisking surfaces, in which each set of whiskers on the snout shows different patterns of movement depending on the surface contacted, develops in rats after P14 [20]. The stimulus dependent, rhythmic patterns of whisker movement allows them detailed shape/texture analysis [9,18,21]. A lack of function in whisker-derived somatosensory neural pathway induces several impairment in tactile and motor coordinative behaviors [22,23]. Thus, whisking leads to sensorimotor activity that eventually guides adaptive behaviors. The developmental process of whisking indicates that exploratory behavior and patterns would be coordinated with the emergence of whisking patterns. While whisking is a mechanism of active sensing of textures, several reports [24,25] have indicated that the sensory information through passive whisker touch may also play a role in functional adaptation in early neonatal stage when the whiskers are still immobile. Young rat pups display a twitch response to passive whisker contacts as early as P3 [26]. Whisker clipping in young pups (P4-6) causes pups to take more time to find a teat for suckling, [24,25,27]. Moreover, rat pups spend most of their time in huddles with littermates or dams, as a means of regulating their body temperature [24,28] and this behavior is disturbed when the whiskers are trimmed [26,27]. These findings suggest that the tactile sensory information via whiskers may regulate adaptive behaviors soon after the animal is born. Somatosensory pathway involves discrete chains of neuronal connections from discriminative touch and proprioceptive senses of whiskers, through thalamic neurons, and reaches the projections to somatosensory cortex [11,12]. A precise formation of the somatosensory pathway requires tactile stimuli via whiskers during neonatal stage [11.29], indicating an importance of whisker-tactile sensation in early neural integration process between somatosensory inputs and motor coordinative behaviors.
To date, most studies on whisking behavior have been done in rats. Few studies in adult mice used head-fixed preparations to observe whisking patterns to investigate a detailed whisker movement. Not much is known about the development of whisker sensation and related sensory-motor behaviors in mice, particularly in reference to how freely whisking behavior impacts sensory-motor development of the animal and its exploratory behaviors. The present study aims to fill this major gap. Our present study is undertaken to determine whether active whisking and passive sensation via whiskers take part in adaptive sensorimotor behaviors and their establishment during early stages of life. The battery of tests we chose is also important in assessing whisker sensation-related deficits in ever increasing numbers of genetically altered mouse lines, in which various aspects of whisker-related neural patterns and function in the brain are altered. For example, adenylate cyclase type 1 gene deficient mice, which are known as barreless mice, reveal significant impairment in tactile sensation as well as body coordination in various behavioral tests [63].
Materials and Methods
Animals
All experimental procedures were approved by the University of Maryland School of Medicine Institutional Animal Care and Use Committee. Mice were housed in an AAALAC accredited laboratory facility. C57BL/6 (B6) mice were originally obtained from Charles Liver (Frederick, MD) and maintained in our breeding colony. Mice used as the subjects were housed with their sire and dam in standard cages (28 × 17 × 12.5 cm) with filter-top lids. Subject pups were randomly chosen 3-6 pups from litters of B6 mice. To identify individual pups, each pup was tattooed with a small dot in the toe of the hindpaw (green paste, Ketchum Manufacturing Inc, Canada). All mice received water and standard rodent chow ad labium. The room in which the mice resided was environmentally controlled on a 12:12 h light:dark cycle (0600-1800 h lighting) at a temperature of 21 °C, relative humidity of 50-60%. To avoid the effect of repeated testing and habituation, each mouse pup was randomly assigned to experiments upto three times during developmental ages (e.g. mouse pup A was used on P-4, P9, and P14 for the battery of developmental experiments).
Behavioral testing
We conducted a series of behavioral tests to investigate the development of motor and sensory performances in male and female B6 mice [30,31]. One group of mouse pups had their whiskers clipped bilaterally every other day starting at P3 to assess the significance of whiskers in behavioral development. Whiskers were visualized under an operating microscope and clipped with microscissors close to the skin surface (>0.5 mm in length). We chose P3 as the starting point based on the studied twitch response (initial reflective whisker movement observed on P3) in rat pups, although spontaneous twitching is observed before birth [26,32]. Mouse pups were assigned to one of the following four groups: male and female B6 pups with intact whiskers and male and female B6 pups with clipped whiskers (n=9, each group, each test day). We conducted tests on P4, 7, 9, 12, 14, and 17. The average of the litter size we used was 7.4 (range from 5 to 11). Three to five pups from a litter were chosen as the subjects. Our test battery consisted of reflexive sensorimotor ability task (grasp test, and surface righting test), vestibular and body coordination task (negative geotaxis test), whisker-sensorimotor task (cliff avoidance test), tactile sensation task (whisker stimulation test, body stimulation test), and social contact task (huddling test). Individual pups were tested for these behavioral tasks on each test day in the following order: surface righting test, cliff avoidance test, negative geotaxis test, grasp test, whisker stimulation test, tactile stimulation test, and huddling test. A set of test trials for each individual mouse was done within 30 min. In addition, pups were monitored every day, after a set of the trials were completed, for the appearance of developmental body landmarks and their weights were recorded before returned to their home cages.
Surface righting test
Surface righting is defined as the time (sec) a pup lying on its back takes to right itself on all 4 limbs. This is a reflex behavior and requires vestibular and spinal motor control [33,34]. Each mouse was placed on its back on a flat surface of a plastic Petri dish (20 cm diameter) and released. The pups were observed for up to 60 seconds. The latencies required to roll over and have all 4 paws in contact with the surface were recorded. The maximum time allowed per trial for righting was 60 sec, and if the mouse reached the cut-off time limit, the experimenter placed the pup in an upright position. Following the pup's upright positioning, exploratory behavior in the dish was observed for 60 sec. Exploratory behavior was calculated as the percent of time that pups showed head or body movement including rotating, locomotive shifting, and head moving. Each mouse was tested in 2 trials each test day.
Negative geotaxis test
Negative geotaxis is an innate response to gravitational cues; it requires vestibular and motor coordination to orient the body up slope on an angled plane [35-37]. Negative geotaxis is defined as the time (sec) it takes a pup placed head down on a 40-degree incline to turn 180-degree and begin crawling up the slope [38,39]. The pups were placed on an inclined plywood surface (20 × 20 cm) and observed for up to 30 seconds. Each pup was given 2 trials each test day.
Grasp test
This test is designed to determine the ability of the pup to grasp a thin rod stroking the palmar surface of each forepaw. The pups were placed in plastic Petri dishes (200 mm diameter, 20 mm height) and left unrestrained for 1 min. Then the experimenter touched and stroked against the palmar surface of a forepaw with a thin rod, and the responses were recorded. If pups showed signs of grasping the rod, then the rod was slowly drawn upward and it was checked if the pup keeps holding with its forepaw against the body weight of their upper body as the forelimb had sustained its grip. Each pup was examined in 2 trials of a rod contact on right and left forepaws. The palmar grasp reflex is characterized by closing of the digits in response to an object stroking the palmar surface [40,41]. The magnitude of this reflex was rated as absent (0 points), crooked digits but not grasped (0.5 point), or present (1 point) in each of the forelimbs separately. On the other hand, the grasp reaction was defined by clasping the rod with fingers and sustaining its forelimb for 1 sec [33,40] (the rod was slightly bent by resistance and the forelimb was stretched, while the pup had its hindlimbs on the ground). The score of the grasping reaction was made as absent (0 points), grasp but not sustained (0.5 point), or sustained (1 point) in each of the forelimbs separately. The mean of this score was calculated.
Cliff avoidance test
To test the ability to recognize the cliff edge [30,40], each mouse was placed on an edge of the platform elevated 5 cm from the table with the forepaws and nose over the edge. The latencies for backing and turning away more than 45° horizontal angle from the start point were counted. Each pup was tested in 2 trials each test day, with a cut-off time of 30 sec allowed for each trial. Behaviors of pups during the trial including the number of head dipping over the edge of the platform were also recorded.
Whisker stimulation test
In neonatal pups when whiskers are physically stimulated, behavioral reactions including twitching responses and head moving reactions are observed [25,42]. In the present study, one min after grasp test was completed, pups were kept in a Plexiglas Petri dish to avoid experimental handling. Unilateral whisker stimulation consisted of manual stimulation of the whiskers with a thin stainless rod (0.5 mm in diameter), for 20 sec (30 sweeps back and forth across the entire whisker field). Stimulation was repeated flexion of each whisker, without touching the intervibrissal fur or the snout skin. A similar procedure was applied to whisker-clipped pups, but a thin rod traced along the unilateral whisker pad without touching (spaced at 3-5 mm distance). Behaviors including twitch reflex and head moving reaction were observed during whisker stimulation on each side of the snout (each event was counted as 1 point). Whisker-clipped pups showed these reactions, to some degree, by sensing airflow from the rod tracing the snout skin or detecting rod movement by eyes after eyes opening.
Tactile stimulation
To evaluate the bodily tactile sensitivity irrespective of whisker sensation [30,40], pups were kept in a Plexiglas Petri dish for 1 min following whisker stimulation test. Then, several points along the body, including the top of the head, medial back, left and right forepaws, hindpaws, and the base of the tail (total 7 points) were manually poked 5 times with a thin stainless rod (0.5 mm diameter). The reaction of mice was analyzed in a scale in which 0 point indicates absence of response, and 1 point indicates presence of the response such as withdrawal or twitching. The scores were calculated as the percentage of the responses in the total stimulation (0-7 points).
Huddling test
The huddling test assessed the ability of pups to locate and maintain contact with another stationary pup referred to as the “target” pup (same age, non experimental littermate) [e.g., 24,43]. Before testing, each target pup was briefly (10 sec) exposed to an induction box in which a volume of isoflurane-saturated gas was filled [44]. The target pup was placed in a prone position in the center of a Plexiglas Petri dish, and the test pup was then placed perpendicular to the target pup with its head touching the target pup. During a 5 min observation, time spent in contact with the target pup was measured as partial huddling (<40% of body contacting with the target pup) and the full huddling (>40% of body contacting with the target pup), and all pup behaviors were monitored. Pups were returned to their home cage with the dam after completion of the session.
Whisking behavior
We assessed micro movement of whiskers in mouse pups when they contact (or not) a small object. In separate groups of pups (n=7 in each day), randomly chosen from 4 litters of B6, we assessed whisking behavior. These were freely moving pups throughout development from P7 to 19. To visualize the whiskers, a small drop of write-out ink was placed on the tip of the whiskers before testing. Pups were placed in a Plexiglas container (20 cm wide, 4 cm height) with a black floor. The whisking behavior was video recorded from above with a high-speed (120 fps) camera (EX-FH100, Casio), and later analyzed. The frequencies and the types of whisking toward the objects or in open air were quantified, by counting whisker movement in each frame of the video recordings for at least 3 sec length for each whisker motion. The total duration of each video recorded was minimum 4 minute [e.g., 22].
Statistical analysis
All data are presented as mean ± S.E.M. The data from developmental behavioral testing were analyzed by a three-way analysis of variance (ANOVA) with between-subject factors of whisker clipping (WT vs. WC) and sex (male vs. female) and a within-subject factor of age (P4, 7, 9, 12, 14, and 17) followed by the Bonferroni test, if differences were found. Whisking behaviors were evaluated by the appearance of whisking and the frequencies for each whisker movement type, by using a one-way ANOVA with age as a factor. Additionally, the frequency differences between the types of whisker movements were analyzed by a two-way ANOVA with subject factors of whisking type and age, followed by the Bonferroni test as a post hoc test. For all data, the accepted level of significance was p<0.05.
Results
Development of motor performance
Surface righting test
We evaluated the motor performance in mouse pups with or without whiskers. Although there was no sex difference in righting latencies, F(1,32)= 0.46, n.s., the latencies for body righting were longer in whisker-intact pups compared to whisker-clipped pups, F(1,32)= 14.86, p<.01, and this quickly decreased over age, F(5,160)= 88.27, p<.01. Subsequent analysis of clipping × age interaction, F(5,160)= 5.04, p<.01, revealed that mice with clipped whiskers show shorter latencies for righting than those with intact whiskers at only P4 (p<.01)(Fig. 1a).
Figure 1.

Motor performance in male and female B6 mouse pups with (WT) or without (WC) whiskers. Data are expressed as mean ± S.E.M. * indicates significant differences between WT and WC and # indicates age differences compared to P4. In the surface righting test, (a) the latencies for righting their body, and the percentage of observation for (b) locomotion and (c) nose rubbing at a wall during a 60-sec test were measured. In the negative geotaxis test, (d) the latencies for body rotation decrease over development. In the grasp test, the ratio of observation for (e) grasp reflex and (f) grasping response were measured.
Locomotor activity in a Petri dish following righting was calculated as the percentage that mice displayed locomotor movement during a 60 sec observation session. There was no significant difference in whisker clipped, F(1,32)= 0.04, n.s. or sex, F(1,32)= 0.26, but the locomotion increased with age, F(5,160)= 28.45, p<.01. Further analysis indicated that locomotion was greater at P12, 14, or 17 than at P4 (Fig. 1b).
Additionally, some pups repeatedly rubbed their snout horizontally at the surface of the wall. This was typically observed following P12 in whisker-clipped mice. An ANOVA for these scores revealed significantly higher percentage of snout rubbing in whisker-clipped pups than whisker-intact pups; Clipping: F(1,32)= 15.46, p<.01. There was no sex effect; Sex: F(1,32)= 1.06, n.s., while age effect was significant, F(5,160)= 14.12, p<.01. Subsequent analysis of whisker clipping × age interaction, F(5,160)= 6.08, p<.01, indicated that this stereotypic behavior is present at higher ratio at P12 (p<.01), P14 (p<.05), and P17 (p<.05) than P4, especially in whisker-clipped pups (Fig. 1c).
Negative geotaxis test
We measured the latencies for complete rotation from a prone position facing down a slope to upwards through development (Fig. 1d). There was no significant difference between whisker-clipped and whisker intact pups, F(1,32)= 3.08, p<.10, regardless of their sex, F(1,32)= 0.01, n.s. A small age-related difference was found; Age: F(5,160)= 13.03, p<.01, indicating that the latencies for rotation were shorter at P14 (p<.05)and P17 (p<.01) than P4.
Grasp test
The appearance of grasp reflex in both the right and left forepaws was evaluated throughout development (Fig. 1e). There was no significant difference between whisker-clipped and whisker-intact pups, F(1,32)= 0.01, n.s., or male and females, F(1,32)= 0.02, n.s. An age-related difference in the appearance of grasp reflex was found, Age: F(5,160)= 14.99, p<.01. The ratio of grasp reflex was greater at P7, 9, 12, and 14 than at P4.
Mice display active grasping to hold a small object. The grasp reaction was observed throughout development (Fig. 1f). No significant difference was found between male and female pups, F(1,32) =0.02, n.s. The main effects of clipping, F(1,32)= 6.98, p<.05, and age, F(5,160)= 184.84, p<.01, were significant. The interaction between clipping and age, F(5,160)= 8.96, p<.01, indicated that mice with clipped whiskers displayed a higher ratio of active grasping response than those with intact whiskers at P12, F(1,32)= 14.27, p<.01. Overall, pups showed significantly increased grasping response at P9, 12, 14, and 17 compared to P4, regardless of the whisker clipping.
Body weights
The appearance of developmental landmarks in body weights was not different between pups with intact and clipped whiskers; clipping: F(1,32)= 1.91, n.s., or sex: F(1,32)= 1.42, n.s., while there was significant age effect, F(5,160)= 395.72, p<.01. The whisker clipping × age interaction was significant, F(5,160)= 7.46, p<.01, but the other interactions were not significant. Overall, there was a significant difference between whisker-clipped and whisker-intact pups at P17 (p<.01) (Fig. 2a), but not at earlier ages [P17, intact males: 7.92 ± 0.42 g; whisker-clipped males: 6.90 ± 0.29 g; intact females: 8.16 ± 0.25 g; and whisker-clipped females: 6.97 ± 0.30 g].
Figure 2.

Whisker and tactile sensory performance in male and female B6 mouse pups with (WT) or without (WC) whiskers. Data are expressed as mean ± S.E.M. * indicates significant differences between WT and WC and # indicates age differences compared to P4. (a) The body weight gains in male and female B6 pups with or without whiskers. In the cliff avoidance test, (b) the latencies for avoidance from a cliff edge, and (c) the number of head dipping observed during a session are presented. In the whisker stimulation test, (d) the number of twitching response and (e) head moving response toward a 30 times whisker stimulation were measured. In the tactile stimulation test, (f) the percentage of twitching responses to total 7 stimulation on their body were calculated.
Whisker-related sensory motor performance
Cliff avoidance test
The latencies for avoiding a cliff edge were measured throughout development (Fig. 2b). On all days of testing, whisker-clipped mice showed longer latencies for cliff avoidance than those with intact whiskers, F(1,32)= 105.73, p<.01, while there was no significant difference between sexes, F(1,32)= 0.06, n.s. A significant difference in age was found, F(5,160)= 6.42, p<.01; the overall avoidance latency was shorter at P14 and 17 than at P4.
In both groups of mouse pups, head dipping at the edge of the platform during the test was observed beginning around P12 (Fig. 2c). Whisker intact pups revealed head dipping significantly more than whisker-clipped pups, F(1,32)= 80.89, p<.01, regardless of their sex, F(1,32)= 3.57, p<.10, especially at P12 (p<.01), P14 (p<.01), and P17 (p<.01). Overall, the number of head dipping increased at P12, P14, and P17 compared to P4, Age: F(5,160)= 66.69, p<.01.
Whisker stimulation test
When a physical stimulus is applied to their whiskers, mouse pups show a startle response such as twitching of the snout or the face, or head moving response such as orienting the head forward or backward in relation to the stimulus source. We investigated reactions to gentle stimulation of whiskers on either side of the snout in pups throughout development. Whisker-clipped pups showed spontaneous, coincident twitching or head moving behaviors during observation, but the scores for these indices were lower than whisker-intact pups. The scores for twitching reflex to whisker stimulation (Fig. 2d) indicate a clear difference between whisker-intact and whisker-clipped pups, F(1,32)= 475.41, p<.01, but no difference between the sexes, F(1,32)= 1.41, n.s. The scores of twitching reflex in whisker-clipped pups were a chance level since the rod stimulation was applied not to the whiskers, but nearby snout. The main effect of age was also significant, F(5,160)= 15.30, p<.01. The subsequent analysis for clipping and age interaction, F(5,160)= 12.47, p<.01, revealed that twitching response was greater at P12, 14, and 17 than P4 in whisker-intact pups.
We evaluated the scores of head moving response to whisker stimulation by a three-way ANOVA (Fig. 2e). We found a significant difference between whisker-intact and whisker-clipped pups, F(1,32)= 184.79, p<.01, while there was no difference between sex, F(1,32)= 0.06, n.s. The main effect of age was also significant, F(5,160)= 24.03, p<.01. Further analysis of whisker clipping and age interaction showed, F(5,160)= 20.54, p<.01, that significant differences exist between whisker-clipped and whisker-intact pups at P7, 9, 12, 14, and 17 (each p<.01). In the whisker-intact pups, the scores for head moving were greater at P9, 12, 14, and 17 than P4 (each p<.01).
Tactile response
Mice show twitching responses when gently poked along the body with a thin rod. We measured this response throughout development. There were no significant differences between whisker-intact and whisker-clipped pups, F(1,32)= 0.41, n.s., or sexes, F(1,32)= 0.56, n.s. However, there was an age-related difference, F(5,160)= 12.48, p<.01, the tactile response ratio was higher at P12, 14, and 17 than at P4 (Fig. 2f).
Huddling
Mouse pups typically huddle with their littermates or caregivers in their nest and even outside of the nest [43]. They adjust their huddling posture and position in the group to manage their body temperature [24,43]. We assessed huddling tendencies and postures in B6 mice with or without whiskers through development. The percentage of huddling observed during the test was separately calculated by the huddling postures; partial (<40% body contacts) and full contact huddle (>40% body contacts) (Fig. 3a). For data of partial huddling, there were no significant differences in either whisker-clipped, F(1,32)= 0.03, n.s., sex, F(1,32)= 0.05, n.s., or age, F(5,160)= 0.98, n.s. On the other hand, the percentage of full huddling during the session showed a significant decrease in whisker-clipped pups compared to whisker-intact pups, F(1,32)= 135.43, p<.01, throughout tested ages (Fig. 3b). There was no significant differences between sexes, F(1,32)= 0.18, n.s. There was an age difference, Age: F(5,160)= 3.49, p<.01, P14 pups displayed more huddling compared to P4 pups, regardless of having whiskers or not.
Figure 3.

Huddling with a littermate was observed in male and female B6 mouse pups with (WT) or without (WC) whiskers. Data are expressed as mean ± S.E.M. * indicates significant differences between WT and WC and # indicates age differences compared to P4. (a) partial huddling, defined as <40% of pup's body contact with a littermate, and (b) full huddling, defined as >40% of pup's body contact with a littermate that was observed from above camera recording are presented.
Whisking behavior
Mice are capable of whisking across objects with high frequency of whisker vibration [e.g., 15,19]. We utilized a high-speed camera and analyzed the frequencies and types of whisker vibration during development from P7 to 19. We found three different types of whisker movements: a) The whiskers slightly swayed horizontally with the snout movement in respiration (respiratory fluctuation). b) Symmetrical movement of whiskers back and forth that are spread out (symmetry whisking). c) Oriented bundled movement of whiskers especially protracted toward an object (active whisking) (Fig. 4a). A slight fluctuation of whiskers with respiration was observed at all ages from P7 to 19. The ratio of whisking in the total number of events that a snout of the pups approached an object (more than 5 events for each mouse) was calculated. This revealed that active movement of whiskers (symmetry whisking) emerges around P10. A one-way ANOVA on these ratios revealed a significant age difference, F(12,72)= 68.40, p<.01, and indicated that the ratio of whisking during object contact was significantly higher after P10 compared to P7.
Figure 4.
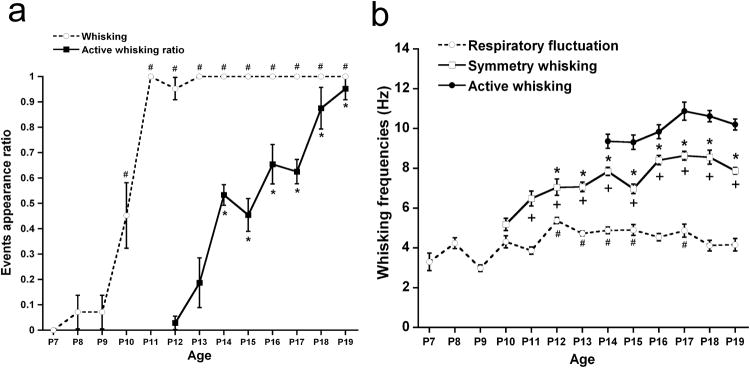
Development of whisking behavior observed in B6 mouse pups from P7 through P19. Data are expressed as mean ± S.E.M. (a) The ratio of appearance of whisking behavior (dashed line) and the ratio of active whisking in total observed whisking during an object contact (solid line) were calculated. # indicates age differences compared to P7; * indicates significant age differences compared to P12. Whisking behavior in B6 pups emerges around P10 and is consolidated by P11. When exploring an object, pups show active whisking after P14 and the appearance ratio increases over ages. (b) The frequencies of whisker movement including respiratory fluctuation (dashed line), symmetry whisking (open square, solid line), and active whisking (closed circle, solid line) were measured. # indicates age differences in respiratory fraction compared to P7. * indicates significant age difmferences in symmetry whisking compared to P10; and + indicates frequencies differences between respiratory fraction and symmetry whisking.
Following the emergence of symmetry whisking, the active whisking appeared around P14. We calculated the ratio of active whisking in total whisking during contacting an object (more than 5 events for each mouse)(Fig. 4a). The ratio of active whisking was about 0.5 on P14 and increased thereafter to 0.9 on P19. A one-way ANOVA conducted on the scores of active whisking ratio from P12 to 19 showed a significant age effect, F(7,42)= 27.93, p<.01. Subsequent analysis indicated that the ratio was higher after P14 compared to P12.
The frequencies of whisker movement for each type of whisking were measured as time of fluctuation per sec (Hz)(Fig. 4b). The frequencies for respiratory movement (∼5 Hz) changed with age, F(12,72)= 6.38, p<.01; the frequencies for respiratory fluctuation were greater at P12, 13, 14, 15, and 17 compared to P7. Symmetry whisking started in all pups at P10 (∼ 8Hz). The frequencies for whisker vibration were compared from P10 through 19 and we found that the frequencies for symmetry whisking were faster from P12 compared to P10 and from P17 compared to P11, F(9,54)= 12.36, p<.01. Finally, active whisking emerged in all pups observed from P14 onwards (∼11 Hz). Age differences in frequencies for active whisking were evaluated by a one-way ANOVA and no significant difference was found, F(5,30)= 2.08, n.s.
To assess developmental diversity of whisking frequencies, we compared the frequencies of whisker vibration between respiratory fluctuation and symmetry whisking from P10 to 19. The frequencies were significantly different between the two types of whisker vibration, F(1,6)= 1337.2, p<.01, and age, F(9,54)= 10.05, p<.01. The interaction between type of vibration and age was also significant, F(9,54)= 8.11, p<.01. Subsequent analysis indicated that the vibration frequencies were faster in symmetry whisking than respiratory fluctuation after P11 (each p<.01), but not at P10.
The frequency differences were also evaluated between the types of whisking; symmetry vs. active whisking from P14 to 19. The frequencies for active whisking were significantly faster than those for symmetry whisking, F(1,6)= 85.28, p<.01, while the main effect of age was also significant, F(5,30)= 5.90, p<.01. The interaction between type of whisking and age was not significant, F(5,30)= 1.05, n.s. Thus, the whisking frequency is faster in active whisking than symmetry whisking from P14 to 19.
Discussion
Development of motor performance
We assessed the sensorimotor development of B6 pups from P4 to 17 and compared to that of pups deprived of the sensory apparatus following bilateral whisker trimming. Overall results of the present test battery indicated no sex differences in motor performance throughout development. During early neonatal stage, motor performance is gradually developed from partial reflex to full body control and the motor neural system is integrated with somatosensory nerve circuit. Previous studies documented the maturation of locomotion and posture in the neonatal rat and mouse [45,46]. By P2, rat pups can move by dragging and then they can pivot their head. At P5, the head can be raised but angled downwards. Around P11, a stable quadruped posture is gained and the posterior body can be supported off the ground [46]. Subsequently, standing on four limbs sustaining the body emerges at around P12 [15,41]. Our present results support and extend these observations in B6 mice and reveal that the sensory motor system develops through neonatal stage from a micro reflex movement at around P7 to integrated, coordinated behavior at around P12.
In the tactile response tasks (whisker and skin stimulation), mouse pups were required to sense a contact of the rod and display a body reaction to the tactile stimulus. Although the partial reaction to touch sense such as startling, twitching, or grasp reflex was observed from P4, the body responsivity to touch sense of whisker and fur/skin was markedly improved around P12. This development of sensorimotor process indicates that tactile/somatic sensory system would be integrated with motor performance during the neonatal stage, after P12. Such integration process has been reported in the primary somatosensory cortex, in which the discrete neural module, so-called “barrels” is formed during the early stage of life by P15 via tactile integral stimulation from whiskers [10,11]. Each barrel is in anatomic correspondence with one specific mystacial vibrissa on the snout of the animal [47], and thus a deprivation of whisker stimulus during the early stage of life results in a malformation of barrel cortex structure [e.g., 23,48] and thus motor coordinative behavior.
Righting reflex involves vestibular sensation including signals about head and body orientation, proprioceptive touch sensation; and coordinative body movement to express reflexive response [e.g. 33,34]. When the subject is placed supine on the ground (contact righting), tactile stimuli on the body and face would also guide righting reflex [34]. It is noteworthy that contact righting as performed in the present experiment appears normal, while air-righting which represents more vestibular sense appears impaired following gestational space (no gravity) exposure in rat pups [49]. Similarly, negative geotaxis response also requires vestibular sensation (head angle and orientation) and body balance to sustain and rotate their body on the declined surface [35,37]. The righting reflex quickly developed after P7 in B6 pups regardless of gender or the presence of whiskers. Rotating response in the negative geotaxis test also showed a quick development and reached a plateau on P12, regardless of sex or the presence of whiskers. Differential development observed in two vestibular/sensorimotor tests may reflect a difference in required motor patterns/performance. Righting reflex requires reflexive muscle response driven by vestibular or proprioceptive touch sense on the ground. Rotating response in the negative geotaxis test pre-requires consistent sustaining their body by limbs and coordinative body balance to rotate their entire body on the declined surface. Additionally, air righting that represents pure vestibular sense begins to appear about P6-7 in rat pups [34,50]. Taken together, it is suggested that simple sensation including somatic and vestibular sense or tactile sense would develop after P7, while motor performance requires entire body coordination and complex motor control such as locomotor activity, geotaxis, and rotating response all of which improved after P12. This is consistently shown in grasp response, in which grasp reflex is induced by a brief palm skin touch, was developed after P7, and decreased after P17, regardless of sex or whisker function. On the other hand, the forelimb grasp reaction required increased muscle strength as well as coordinative muscle control including toe-palm and forelimb muscles to keep sustaining its upper body by grasping was improved after P12. Grasp reflex showed a quick decline after P17 when pups tended to loss interested in the presented rod or would push away instead of displaying gentle grasping.
To our surprise, pups with clipped whiskers performed better in (contact) righting reflex at P4 and grasping (handling) reaction at P12 compared to those having intact whiskers. Both tasks require development of neural integration from “non-whisker” somatic sensations to body muscle control. Particularly, the righting reflex on the ground requires signal integration from supine touch sense to muscle reflective control, was expressed on P4, while pure vestibular sensation measured by air righting may appear at about P7 and is completed by P14-16 [34,51]. These findings indicate that present righting reflex on the ground at P4 represents not vestibular function, but more distinctly represents proprioceptive tactile cues and coordinative movement regulating reflex. No impairments in tactile touch sensation were detected in whisker-clipped pups at P4. Therefore, better performance in righting reflex at P4 would indicate fine signal integration from touch sense to expressing reflexive reaction. Negative geotaxis in the present experiment utilized a severe decline (>40 degree) would not considerably reflect vestibular sensation [37], but requires coordinative movement, did not show a difference in performance by whisker clipping. Thus, it is still unclear how the development of the vestibular sensation or its integration is influenced by whiskers (tactile) deprivation. The signals in the vestibular system are subtended by pathways running from the vestibular nuclei to the thalamus, cerebellum, and the somatosensory cortex [52]. Vestibular system projections are shown to overlap with the somatosensory system [52,53]. For example, neurons in the vestibular nuclei project to thalamic nuclei where they converge with visual and somatosensory tracts [52,53]. This thalamocortical somatosensory system is formed through stimulus-dependent cortical circuits during the early stage of life [10,11]. Thus, it is possible that lack of whisker sensation (which is a primary source of tactile sense) from early neonatal stage facilitates the development of stimulus-elicited motor coordination excluding whisker touch sensation as a compensatory mechanism. As such, grasping reaction requires sensorimotor coordination including paw-tactile sensation to keep holding the rod as well as coordinative muscle control to sustain its upper body weight by forelimb muscle. Perhaps, whisker sensation and control would form coordinative networks with body somatic sensation and vestibular sensation during early developmental stage. Neural networks in whisker-clipped mouse pups may abbreviate the coordinating process of tactile sensation with whisker sensation.
Development of whisker sensory behavior
Several studies have shown that deafferentation of the whiskers or whisker clipping disrupts tactile discrimination and control of the discriminative whisking pattern [e.g. 16,22,23]. In the present study, whisker trimming in mouse pups from P3 onwards led to reduce body weight gain by P17. This observation per se indicates the importance of whiskers in normal functional development. Deprivation of whisker sensation by whisker trimming may induce a reduction of nutritional energy supplies by disrupting nipple searching [25] and/or aggregation with the dam and littermates for regulating homeostasis of body temperature [24,25]. Sullivan et al [25] demonstrated an importance of whiskers in nipple searching in rats but significant effect was observed only during P3-5. It is likely that olfactory cues play more significant role in searching nipple/milk and mother's cues during the first week of life [43,54]. In the present study, a reduction of body weights in whisker clipped pups was found during P14-17 period, suggesting that a transition from mother's milk to solid food occurs during P15-17 [55] may be affected by whisker presence. Galef et al. [56,57] illustrated that pups do not imitate the mother in her feeding behavior but are simply attracted by her to the feeding area during P15-17. Pups were housed with their dams but they needed to search and find solid food by themselves as the onset of self-feeding. Whisker clipped mice clearly showed a decreased investigation in the open arena and toward placed objects and significant impairment in texture/shape recognition [22,63]. It is highly possible that the deafferentation of whiskers induces delayed transition to solid food or impairment in solid food feeding.
In addition to delayed body weight gain, mice with clipped whiskers displayed a marked reduction of cliff avoidance compared to those with intact whiskers throughout the ages tested, suggesting that the sensory information from whiskers plays a role in functional adaptation even in early neonatal stages (∼P4) while the whiskers are still immobile. Starting at P12, normal pups began to show head dipping at the cliff edge to “evaluate” safety in the environment, which is considerable as a typical exploratory behavior with whiskers in pups, while whisker-clipped pups displayed few head dipping at the edge. This observation indicates an appearance of active, locomotive searching behavior that emerges around P12 and a role for whiskers associated with the emergence of active searching. As described below, this age is the emergence of whisking, but not yet active palpation whisking. When the whiskers are gently stimulated in the whisker sensation test, normal pups are able to partially show a twitching response in the snout or face at P4 and this response increases with age. Head-flipping response appears at P12 in response to whisker stimulation. On the other hand, tactile sensitivity in other parts of the body develops to adultlike (weaned age) levels by P12, regardless of whisker sensory deprivation. This again indicates that although whiskers are able to sense tactile stimuli before P4, coordination of tactile response via whiskers or bodily skin is established around P12. Moreover, Whisker-clipped mouse pups showed persistent snout rubbing at the surface of the wall after P12, which is a stereotypic lateral back and forth movement of the snout, most likely involved in shape recognition in the environment. This movement is considered to be an active exploration by pivoting the head with whiskers touching and it emerges around P12. Whisker-clipped pups may use microvibrissae, instead of whiskers (macrovibrissae) to search the surface of the wall, as a compensatory mechanism.
Sensory deprivation during early stage of life affects whisker-related exploratory behavior later in life [23,48]. Significance of passive whisker sensation can be demonstrated in huddling, which is a typical social behavior during neonatal stage. Mouse pups strongly prefer to huddle with littermates to regulate body temperature and energy consumption [24,43]. When a pup detects littermates via tactile sense, it moves and adjusts body position to make a tight contact with littermates and spend a substantial amount of time in huddles. Mouse pups with clipped whiskers showed a similar tendency for partial huddle, but a reduction of full huddle, compared to those with intact whiskers throughout the ages tested. This indicates that lack of whiskers during contacting with a littermate causes impairment in adjusting movement of body position. Even after eye opening at P14-15 [33], whisker-clipped pups still show decreased full huddling, underscoring the significance of whisker tactile sensation in maintaining behavioral adaptation throughout neonatal period.
Development of whisking behavior
Rat and mouse pups are born with a set of fine, immobile whisker hairs on the snout [3]. A high frequency of whisker movement, so-called “whisking” begins around P11 to 13 in rat pups [20,33]. Adult whisking shows asymmetry of whisker movement during contacting an object [13,17]. This contact-related modulation of whisking behavior in rats is shown to emerge gradually from P11 and it is established by P17 [15,20]. We found that B6 mouse pups began to show whisking around P10 during exploring the environment (see Fig. 5). At P10, whisking frequencies are transient and slow as those in respiratory fluctuation. In freely-moving mouse pups, a frequency of whisking as symmetry movements, around 6-8 Hz, could be observed in pups from P11 to 19, consistent with the findings in rat pups [15,20,58]. We observed another, distinct movement of whiskers in the form of protracted, bundled movement of whiskers toward an object that pups palpate with their whiskers. In particular, it is observed when whiskers make contact with an object. Then, the mouse controls the subsequent whisk to ensure that whiskers contacting an object make the maximum protraction [1,59], which would be the same category of whisking in previous findings in rats, which is called active whisking [16,59] or contact-induced whisking [2]. This active palpation whisking began around P13 when pups were contacting and scanning an object presented to them. The ratio that pups display active palpation whisking when contacting an object increased from 0.5 at P14 to over 0.9 at P19, and the frequencies of whisker movement as active palpation whisking were around 10-11 Hz, which is quite higher than symmetry (free-air) whisking. Gene-manipulated adult mice showing a lack of functional neural connection in whisker-barrel system demonstrated a remarked impairment in whisker related behavior as well as inaccuracy of active palpation whisking and reduced frequencies of whisking [22]. These findings as well as the previous finding in neonatally whisker trimmed rats [33,48] indicate an importance of whisking frequencies and patterns changed depending on situations in normal adaptive behavior. During active palpation whisking, whisking frequency may result in enhanced number of contacts made with a surface of the object of interest. Asymmetrical changes in whisking amplitude reported in rats [16,17,60,61] may serve a similar functional end: increased exploration of the surface and space in the direction in which the animal is moving, and gathering information [2,21].
Figure 5.
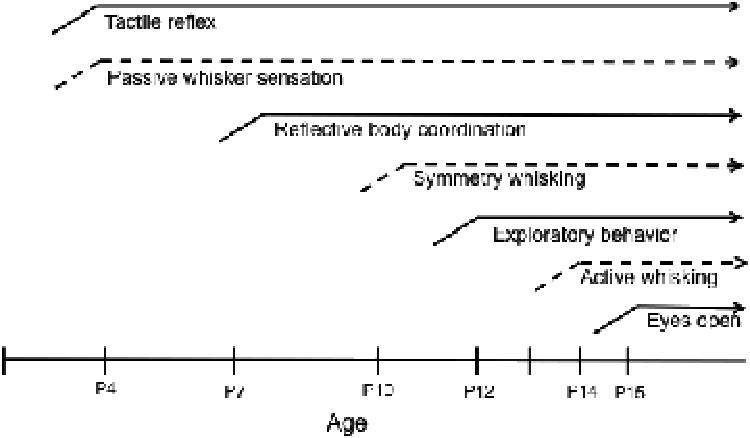
A schematic diagram for development of whisker sensation and exploratory motor coordination in C57BL/6 mouse. Tactile reflex is represented by measuring whisker and skin twitch response and grasp reflection. Reflexive body coordination is shown in measuring righting reflex and negative geotaxis. Exploratory behavior is represented by measuring locomotor activity and cliff avoidance.
Prior to the onset of whisking, neonatal mice (∼P4) are capable of sensing and responding to whisker stimulation, which is also shown in tactile learning paradigms in rats [25,29]. However, at early ages, the pups are not yet able to orient their head or whiskers to the stimulus source. At the onset of whisking around P11 several sensory motor reflexes develop, but integrative motor performance including body coordination and locomotion is not yet established (summarized in Fig. 5). At P13 to 14, a context-dependent (palpation) whisking which is pre-requisite for active scanning of the environment and distinguishing the surface of interest emerges. At this stage of development, integrative motor performance is established (P12), accompanied by eyes open (P15) [e.g. 62]. Sensory information gained through whiskers is critical for passive touch and guidance of body position particularly during early neonatal stage; active whisking develops in parallel with the increasing motor capacities that allow the animal to orient and navigate in the environment. The coordinative development of whisker performance with exploratory motor behavior suggests an importance of tactile sensation in expression of adaptive behavior in postnatal mouse pups.
Highlights.
Whisker function in early development was evaluated in C56BL6 mice
Whisker-clipped pups showed accelerated body coordination development
Passive sensation by whiskers impacts on coordination of body position
Active whisking emerges at P-14 helps searching the surface
Acknowledgments
Supported by NIH/NINDS NS039050 (RSE) Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the above stated funding agencies.
Footnotes
The authors have no conflicts of interest to declare.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Hartmann MJ. A night in the life of a rat: vibrissal mechanics and tactile exploration. Ann New York Acad Sci. 2011;1225:110–8. doi: 10.1111/j.1749-6632.2011.06007.x. [DOI] [PubMed] [Google Scholar]
- 2.Mitchinson B, Martin CJ, Grant RA, Prescott TJ. Feedback control in active sensing: rat exploratory whisking is modulated by environmental contact. Proc Royal Soci B: Biol Sci. 2007;274(1613):1035–41. doi: 10.1098/rspb.2006.0347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Welker WI. Analysis of sniffing of the albino rat. Behaviour. 1964;22:223–44. [Google Scholar]
- 4.Wineski LE. Movements of the cranial vibrissae in the golden hamster (Mesocricetus auratus) J Zool. 1983;200:261–80. [Google Scholar]
- 5.Carvell GE, Simons DJ. Biometric analyses of vibrissal tactile discrimination in the rat. J Neurosci. 1990;10:2638–48. doi: 10.1523/JNEUROSCI.10-08-02638.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Carvell GE, Simons DJ. Task- and subject-related differences in sensorimotor behavior during active touch. Somatosen Motor Res. 1995;12:1–9. doi: 10.3109/08990229509063138. [DOI] [PubMed] [Google Scholar]
- 7.Prigg T, Goldreich D, Carvell GE, Simons DJ. Texture discrimination and unit recordings in the rat whisker/barrel system. Physiol Behav. 2002;77(4-5):671–5. doi: 10.1016/s0031-9384(02)00917-4. [DOI] [PubMed] [Google Scholar]
- 8.Knutsen PM, Pietr M, Ahissar E. Haptic object localization in the vibrissal system: behavior and performance. J Neurosci. 2006;26:8451–64. doi: 10.1523/JNEUROSCI.1516-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.O'Connor DH, Clack NG, Huber D, Komiyama T, Myers EW, Svoboda K. Vibrissa-based object localization in head-fixed mice. J Neurosci. 2010;30:1947–67. doi: 10.1523/JNEUROSCI.3762-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.O'Connor DH, Peron SP, Huber D, Svoboda K. Neural activity in barrel cortex underlying vibrissa-based object localization in mice. Neuron. 2010;67:1048–61. doi: 10.1016/j.neuron.2010.08.026. [DOI] [PubMed] [Google Scholar]
- 11.Erzurumlu RS, Gaspar P. Development and critical period plasticity of the barrel cortex. Eur J Neurosci. 2012;35(10):1540–53. doi: 10.1111/j.1460-9568.2012.08075.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lee S, Kruglikov I, Huang ZJ, Fishell G, Rudy B. A disinhibitory circuit mediates motor integration in the somatosensory cortex. Nature Neurosci. 2013;16(11):1662–70. doi: 10.1038/nn.3544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zucker E, Welker WI. Coding of somatic sensory input by vibrissae neurons in the rat's trigeminal ganglion. Brain Res. 1969;12(1):138–56. doi: 10.1016/0006-8993(69)90061-4. [DOI] [PubMed] [Google Scholar]
- 14.Hartmann MJ. Active sensing capabilities of the rat whisker system. Autonomous Robots. 2001;11:249–254. [Google Scholar]
- 15.Grant RA, Mitchinson B, Prescott TJ. The development of whisker control in rats in relation to locomotion. Dev Psychobiol. 2011;54:151–68. doi: 10.1002/dev.20591. [DOI] [PubMed] [Google Scholar]
- 16.Carvell GE, Simons DJ. Abnormal tactile experience early in life disrupts active touch. J Neurosci. 1996;16:2750–7. doi: 10.1523/JNEUROSCI.16-08-02750.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Towal RB, Hartmann MJ. Right-left asymmetries in the whisking behavior of rats anticipate head movements. J Neurosci. 2006;26(34):8838–46. doi: 10.1523/JNEUROSCI.0581-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Towal RB, Hartmann MJ. Variability in velocity profiles during free-air whisking behavior of unrestrained rats. J Neurophysiol. 2008;100(2):740–52. doi: 10.1152/jn.01295.2007. [DOI] [PubMed] [Google Scholar]
- 19.Grant RA, Mitchinson B, Fox CW, Prescott TJ. Active touch sensing in the rat: anticipatory and regulatory control of whisker movements during surface exploration. J Neurophysiol. 2009;101:862–72. doi: 10.1152/jn.90783.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Grant RA, Sperber AL, Prescott TJ. The role of orienting in vibrissal touch sensing. Front Behav Neurosci. 2012;6:39. doi: 10.3389/fnbeh.2012.00039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Mitchinson B, Grant RA, Arkley K, Rankov V, Perkon I, Prescott TJ. Active vibrissal sensing in rodents and marsupials. Phil Trans Royal Soci B: Biol Sci. 2013;366(1581):3037–48. doi: 10.1098/rstb.2011.0156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Arakawa H, Suzuki A, Zhao S, Tsytsarev V, Lo FS, Hayashi Y, Itohara S, Iwasato T, Erzurumlu RS. Thalamic NMDA receptor function is necessary for patterning of the thalamocortical somatosensory map and for sensorimotor behaviors. J Neurosci. 2014;34(36):12001–14. doi: 10.1523/JNEUROSCI.1663-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Papaioannou S, Brigham L, Krieger P. Sensory deprivation during early development causes an increased exploratory behavior in a whisker-dependent decision task. Brain Behav. 2013;3:24–34. doi: 10.1002/brb3.102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Alberts JR. Huddling by rat pups: Group behavioral mechanisms of temperature regulation and energy conservation. J Comp Physiol Psychol. 1978;92:231–45. doi: 10.1037/h0077459. [DOI] [PubMed] [Google Scholar]
- 25.Sullivan RM, Landers MS, Flemming J, Vaught C, Young TA, Polan JH. Characterizing the functional significance of the neonatal rat vibrissae prior to the onset of whisking. Somatosen Motor Res. 2003;20(2):157–62. doi: 10.1080/0899022031000105190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Blumberg MS. Beyond dreams: Do sleep-related movements contribute to brain development? Front Neurol. 2010;1:140. doi: 10.3389/fneur.2010.00140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kenyon CAP, Keeble S, Cronin P. The role of perioral sensation in nipple attachment by weanling rat pups. Dev Psychobiol. 1982;15:409–21. doi: 10.1002/dev.420150503. [DOI] [PubMed] [Google Scholar]
- 28.Schank JC, Alberts JR. Self-organized huddles of rat pups modeled b simple rules of individual behavior. J Theoret Biol. 1997;189:11–25. doi: 10.1006/jtbi.1997.0488. [DOI] [PubMed] [Google Scholar]
- 29.Landers MS, Sullivan RM. Vibrissae-evoked behavior and conditioning before functional ontogeny of the somatosensory vibrissae cortex. J Neurosci. 1999;19(12):5131–7. doi: 10.1523/JNEUROSCI.19-12-05131.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Crawley JN. Behavioral phenotyping of transgenic and knockout mice: experimental design and evaluation of general health, sensory functions, motor abilities, and specific behavioral tests. Brain Res. 1999;835:18–26. doi: 10.1016/s0006-8993(98)01258-x. [DOI] [PubMed] [Google Scholar]
- 31.Metz GA, Schwab ME. Behavioral characterization in a comprehensive mouse test battery reveals motor and sensory impairments in growth-associated protein-43 null mutant mice. Neurosci. 2004;129:563–74. doi: 10.1016/j.neuroscience.2004.07.053. [DOI] [PubMed] [Google Scholar]
- 32.Tiriac A, Uitermarkt BD, Fanning AS, Sokoloff G, Blumberg MS. Rapid whisker movements in sleeping newborn rats. Curr Biol. 2012;22(21):2075–2080. doi: 10.1016/j.cub.2012.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Roubertoux P, Semal C, Ragueneau S. Early development in mice: II. Sensory motor behavior and genetic analysis. Physiol Behav. 1985;35:659–66. doi: 10.1016/0031-9384(85)90393-2. [DOI] [PubMed] [Google Scholar]
- 34.Pellis SM, Pellis VC. Development of righting when falling from a bipedal standing posture: evidence for the dissociation of dynamic and static righting reflexes in rats. Physiol Behav. 1994;56(4):659–63. doi: 10.1016/0031-9384(94)90223-2. [DOI] [PubMed] [Google Scholar]
- 35.Thiessen DD, Lindzey G. Negative geotaxis in mice: effect of balancing practice on incline behavior in C57BL/6J male mice. Anim Behav. 1967;15:113–6. doi: 10.1016/s0003-3472(67)80020-4. [DOI] [PubMed] [Google Scholar]
- 36.Moser VC. Neurobehavioral screening in rodents. Cur Prot Toxicol. 1999;11:2.1–2.16. doi: 10.1002/0471140856.tx1102s06. [DOI] [PubMed] [Google Scholar]
- 37.Motz BA, Alberts JR. The validity and utility of geotaxis in young rodents. Neurotoxicol Teratol. 2005;27:529–33. doi: 10.1016/j.ntt.2005.06.005. [DOI] [PubMed] [Google Scholar]
- 38.Vorhees CV, Brunner RL, Butcher RE. Psychotropic drugs as behavioral teratogens. Science. 1979;205:1220–5. doi: 10.1126/science.472738. [DOI] [PubMed] [Google Scholar]
- 39.Vorhees CV, Butcher RE, Brunner RL, Sobotka TJ. A developmental test battery for neurobehavioral toxicity in the rats: a preliminary analysis using monosodium glutamate, calcium carrageenan, and hydroxyurea. Toxicol Applied Pharmacol. 1979;50(2):267–82. doi: 10.1016/0041-008x(79)90152-2. [DOI] [PubMed] [Google Scholar]
- 40.Fox WM. Reflex-ontogeny and behavioral development of the mouse. Anim Behav. 1965;13:234–41. doi: 10.1016/0003-3472(65)90041-2. [DOI] [PubMed] [Google Scholar]
- 41.Palanza P, Parmigiani S, vom Saal FS. Effects of prenatal exposure to low doses of diethylstilbestrol, o,p'DDT, and methoxychlor on postnatal growth and neurobehavioral development in male and female mice. Horm Behav. 2001;40:252–65. doi: 10.1006/hbeh.2001.1697. [DOI] [PubMed] [Google Scholar]
- 42.McNamara KCS, Lisembee AM, Lifshitz J. The whisker nuisance task identifies a late-onset, persistent sensory sensitivity in diffuse brain-injured rats. J Neurotrauma. 2010;27:695–706. doi: 10.1089/neu.2009.1237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Alberts JR. Huddling by rat pups: ontogeny of individual and group behavior. Dev Psychobiol. 2007;49(1):22–32. doi: 10.1002/dev.20190. [DOI] [PubMed] [Google Scholar]
- 44.Drobac E, Durand E, Laudenbach V, Mantz J, Gallego J. A simple method for short-term controlled anesthesia in newborn mice. Physiol Behav. 2004;82(2-3):279–83. doi: 10.1016/j.physbeh.2004.03.013. [DOI] [PubMed] [Google Scholar]
- 45.Clarac F, Brocard F, Binay L. The maturation of locomotor networks. Prog Brain Res. 2004;143:57–66. doi: 10.1016/S0079-6123(03)43006-9. [DOI] [PubMed] [Google Scholar]
- 46.Lelard T, Jamon M, Gasc JP, Vidal PP. Postural development in rats. Exp Neurol. 2006;202:112–24. doi: 10.1016/j.expneurol.2006.05.018. [DOI] [PubMed] [Google Scholar]
- 47.Erzurumlu RS, Kind PC. Neural activity: sculptor of ‘barrels’ in the neocortex. Trends Neurosci. 2001;24(10):589–95. doi: 10.1016/s0166-2236(00)01958-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Shoykhet M, Land PW, Simons D. Whisker trimming begun at birth or on postnatal day 12 affects excitatory and inhibitory receptive fields of layer IV barrel neurons. J Neurophysiol. 2005;94:3987–95. doi: 10.1152/jn.00569.2005. [DOI] [PubMed] [Google Scholar]
- 49.Ronca AE, Alberts JR. Effects of prenatal spaceflight on vestibular responses in neonatal rats. J Appl Physiol. 2000;89:2318–24. doi: 10.1152/jappl.2000.89.6.2318. [DOI] [PubMed] [Google Scholar]
- 50.Laouris Y, Kalli-Laouri J, Schwartze P. The postnatal development of the air-righting reaction in albino rats. Quantitative analysis of normal development and the effect of preventing neck-torso and torso-pelvis rotations. Behav Brain Res. 1990;37:37–44. doi: 10.1016/0166-4328(90)90070-u. [DOI] [PubMed] [Google Scholar]
- 51.Hard E, Larsson K. Development of air righting in rats. Brain Behav Evol. 1975;11:53–9. doi: 10.1159/000123624. [DOI] [PubMed] [Google Scholar]
- 52.Jamon M. The development of vestibular system and related functions in mammals: impact of gravity. Front Int Neurosci. 2014;8(11):1–13. doi: 10.3389/fnint.2014.00011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Cullen KE. The vestibular system: multimodal integration and encoding of self-motion for motor control. Trends Neurosci. 2012;35(3):185–96. doi: 10.1016/j.tins.2011.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.McLean JH, Harley CW. Olfactory learning in the rat pup: a model that may permit visualization of a mammalian memory trace. Neuroreport. 2004;15(11):1691–7. doi: 10.1097/01.wnr.0000134988.51310.c3. [DOI] [PubMed] [Google Scholar]
- 55.Rosenblatt JS. The development of maternal responsiveness in the rat. Am J Orthopsychiatry. 1969;39(1):36–56. doi: 10.1111/j.1939-0025.1969.tb00619.x. [DOI] [PubMed] [Google Scholar]
- 56.Galef BG, Jr, Clark MM. Social factors in the position avoidance and feeding behavior of wild and domesticated rat pups. J Comp Physiol Psychol. 1971;75(3):341–57. doi: 10.1037/h0030937. [DOI] [PubMed] [Google Scholar]
- 57.Galef BG, Jr, Clark MM. Mother's milk and adult presence: two factors determining initial dietary selection by weaning rats. J Comp Physiol Psychol. 1972;78:220–5. doi: 10.1037/h0032293. [DOI] [PubMed] [Google Scholar]
- 58.Hill DN, Bermejo R, Zeigler HP, Kleinfeld D. Biomechanics of the vibrissa motor plant in rat: rhythmic whisking consists of triphasic neuromuscular activity. J Neurosci. 2008;28(13):3438–55. doi: 10.1523/JNEUROSCI.5008-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Sachdev RN, Sato T, Ebner FF. Divergent movement of adjacent whiskers. J Neurophysiol. 2002;87:1440–8. doi: 10.1152/jn.00539.2001. [DOI] [PubMed] [Google Scholar]
- 60.Landers M, Zeigler HP. Development of rodent whisking: trigeminal input and central pattern generation. Somatosen Motor Res. 2006;23:1–10. doi: 10.1080/08990220600700768. [DOI] [PubMed] [Google Scholar]
- 61.Berg RW, Kleinfeld D. Rhythmic whisking by rat: retraction as well as protraction of the vibrissae is under active muscular control. J Neurophysiol. 2003;89(1):104–17. doi: 10.1152/jn.00600.2002. [DOI] [PubMed] [Google Scholar]
- 62.Sachdev RN, Berg RW, Champney G, Kleinfeld D, Ebner FF. Unilateral vibrissa contact: changes in amplitude but not timing of rhythmic whisking. Somatosen Motor Res. 2003;20(2):163–9. doi: 10.1080/08990220311000405208. [DOI] [PubMed] [Google Scholar]
- 63.Arakawa H, Akkentli F, Erzurumlu RS. Region-specific disruption of Adenylate cyclase type 1 gene differentially affects somatosensorimotor behaviors in mice. eNeuro. 2014:0007–14. [PMC free article] [PubMed] [Google Scholar]