

Abstract
Antibody and T-cell reactivities to Plasmodium vivax merozoite surface protein-9 (PvMSP9) were evaluated in a cross-sectional study of individuals naturally exposed to malaria infections living in Ribeirinha, a native riverine community and in Colina, a transmigrant community, Rondonia, Brazil. The antibody responses to PvMSP9-RIRII and PvMSP9-Nt domains in Ribeirinha were higher compared with Colina and correlated with age and time of malaria exposure. IgG2 was most prevalent for PvMSP9-RII in both communities, and IgG1 was the predominant isotype for PvMSP9-Nt and PvMSP9-RIRII in Ribeirinha. IFN-γ and IL-4 predominated in Ribeirinha, while IFN-γ predominated in Colina. Variation in exposure to P. vivax likely accounts for the differences observed in cytokine and antibody levels between the two populations studied.
Keywords: Plasmodium vivax, Malaria, Vaccine, Merozoite Surface Protein-9
1. Introduction
Malaria remains one of the most important parasitic diseases in the world, causing 300 to 500 million cases per annum. Although P. falciparum is responsible for the majority of these cases, P. vivax malaria has a wider geographic distribution and accounts for at least an estimated 80 million clinical cases annually throughout most of Asia, Oceania, southeastern Europe, Central and South America, and parts of Africa. Despite this widespread prevalence, P. vivax has long been overshadowed by the burden caused by P. falciparum malaria, which causes more than one million deaths in sub-Saharan Africa alone. P. vivax infections are chronic, can produce profound morbidity with anemia and can be incapacitating for days or weeks [1, 2]. P. vivax has the added complication of recurrent clinical episodes due to the reactivation of dormant liver parasite forms, the hypnozoites [3]. The disproportionate morbidity and mortality figures and an absence of an in vitro continuous culture are the primary reasons why studies on P. vivax have lagged far behind that of P. falciparum. However, the dramatic expansion of P. vivax and the emergence of strains resistant to chloroquine over the last decade [4, 5] and, more recently, to primaquine [6] have created the need for alternative prophylactic and therapeutic strategies including the search for novel targets for vaccine development.
Malaria asexual blood-stage vaccines aim to disrupt the interaction between Plasmodium merozoite proteins and erythrocyte surface by eliciting neutralizing antibodies [7, 8]. Among the P. vivax merozoite proteins being investigated as vaccine candidates are the reticulocyte binding proteins 1 and 2 (PvRBP1 and PvRBP2)[9], the Duffy binding protein (PvDBP)[10–12], the apical membrane antigen 1 (AMA-1)[13, 14], the merozoite surface protein 1 (PvMSP-1) [15], merozoite surface protein 3 (PvMSP3) [16] and, more recently, the merozoite surface protein 9 (PvMSP-9) [17].
There are abundant data generated through studies of human, simian and rodent malaria infections that both humoral and cellular immune responses are important in vaccine induced resistance for preventing or limiting the development and maintenance of erythrocytic stage parasites [18–21]. Several lines of evidence indicate that antibodies to merozoite surface proteins elicited by natural infection mediate the non-sterilizing immunity that characteristically develops in many residents of malaria endemic areas [21– 24]. Therefore proteins at the surface of Plasmodium merozoites are potential targets of malaria vaccines.
PvMSP-9, like all classical Plasmodium MSPs, is expressed during schizogony and becomes associated with the surface of merozoites in the course of schizont development and segmentation [17]. The deduced protein contains a hydrophobic signal sequence, a highly conserved N-terminal domain with a cluster of four cysteine residues, and a C-terminal region containing two species-specific blocks of repeated amino acids. Structurally related molecules have been identified in P. knowlesi (PkMSP-9), P. cynomolgi (PcyMSP-9), P. coatneyi (PcoMSP-9) and P. falciparum (PfMSP-9, also known as p101 or ABRA since its original designation) [17, 25, 26].
PvMSP-9 is considered as a potentially important vaccine candidate based on its surface association and the demonstration that a P. vivax monoclonal antibody and polyclonal antiserum against the native PcyMSP-9 can interfere with the entry of P. vivax merozoites into red blood cells in short-term in vitro cultures [25]. Moreover, PvMSP-9 appears to be highly conserved across Plasmodium species. Rabbit antibodies raised against the native PcyMSP-9 cross-react with MSP-9 of P. vivax, P. knowlesi and P. cynomolgi in immunofluorescence and immunoblot assays [17]. Similarly, a rabbit antiserum raised against a recombinant N-terminal fragment, PvMSP-9-Nt, cross-reacts with MSP-9 from each of these species and weakly with P. falciparum by immunoblot [17]. We recently demonstrated that immunization of BALB/c mice with recombinant PvMSP9-Nt and the second block of repeats (PvMSP9-RII) generates a cellular response with secretion of IFN-γ and IL-4 in addition to a strong humoral response with high titers of specific antibodies that recognize the parasite’s native protein [27].
Given the cumulative data supporting the vaccine potential of PvMSP-9, the present study which for the first time examines the acquired cellular and humoral immune responses against PvMSP-9 in humans living in malaria-endemic areas of Brazil and naturally exposed to P. vivax infections will provide important information on whether immune responses specific to this antigen are generated in natural infections and their immunogenic potential as vaccine candidates.
2. Material and Methods
2.1. Study area and volunteers
A cross-sectional cohort study was conducted in populations living in two distinct communities in the malaria endemic region of Rondonia state, Brazil, where P. vivax malaria accounts for more than 75% of all malaria cases in the last five years (Brazilian Ministry of Health, 2007, unpublished data). Samples and survey data were collected during the dry months of June–August of 2004, coinciding with the period of increased malaria transmission in Rondonia State. The two communities of Colina and Ribeirinha have been described previously [28, 29]. Briefly, Colina consists primarily of transmigrants from non-endemic areas of Brazil (63.8%) who have lived in the region for 10 years or more. They reside in rural settlements alongside unpaved roads that traverse the transnational highway BR-364 approximately 50–100 km southeast of the capital, Porto Velho. Ribeirinha encompasses riverine communities along the banks of the Madeira River and its tributaries approximately 30–60 km northeast of the capital. Families in Ribeirinha have lived in the malaria-endemic region for over 25 years, and the majority (91.9%) is natives of the rain forest region. In both communities, more than 80% of the individuals live close to the forest and mosquito breeding places and have outdoor activities early in the morning and in the evening. In these areas year-round transmission of P. vivax and P. falciparum occur, increasing during the dry months (June and July). The study population consisted of 121 donors from Colina and 185 donors from Ribeirinha. In addition, 24 naive control subjects were included from non endemic regions of Rio de Janeiro, Brazil and Atlanta, Georgia, USA.
2.2. Epidemiological survey
In order to evaluate epidemiological factors that may influence the immune response against PvMSP-9, all donors were interviewed after informed consent was obtained. Questions on the survey related to demographics, time of residence in the endemic area, personal and family histories of malaria, use of malaria prophylaxis, presence of malaria symptoms, and personal knowledge of malaria. Survey data was entered into a database created with Epi Info 2002 (Centers for Disease Control and Prevention, Atlanta, GA). Written informed consent was obtained from all adult donors or from parents of donors in the case of minors. The study was reviewed and approved by the Fundação Oswaldo Cruz Ethical Committee and National Ethical Committee of Brazil.
2.3. Collection of human blood samples and malaria diagnosis
Venous peripheral blood was collected into heparinized tubes, and peripheral blood mononuclear cells (PBMC) were isolated by Ficoll/Hypaque (Pharmacia, Piscataway, NJ) density gradient centrifugation and used in the ELISPOT assays. Plasma was stored at −20°C and thin and thick blood smears of all donors were examined for malaria parasites. The numbers of donors positive for P. vivax and P. falciparum at the time of blood collection are listed in Table 1. All smear-positive donors were subsequently treated for P. vivax or P. falciparum per the regimen recommended by the Brazilian Ministry of Health.
Table 1.
Summary of epidemiological and malaria history characteristics in Colina (CO) and Ribeirinha (RI) population, Rondonia State, Brazil.
| CO | RI | P valuesa | ||
|---|---|---|---|---|
| Gender | ||||
| Male – n (%) | 61 (50,4%) | 86 (46,5%) | ||
| Female – n (%) | 60 (49,6%) | 99 (53,5%) | χ2= 0.171 | p=0.2998 |
| TOTAL (n) | 121 | 185 | ||
| Age (M ± SD) | 37.5±16.3 | 35.5±16.6 | t=1.021 | p=0.5960 |
| Years of residence in malaria endemic area (Median) | 19 | 27 | MW=21.906 | p<0.0001 |
| Years of residence in Rondonia State (Median) | 18 | 27 | MW=31.084 | p<0.0001 |
| Years of residence in present address (Median) | 6 | 13 | MW=22.313 | p<0.0001 |
| Total number of past malaria episodes (Median) | 8 | 5 | NSb | NSb |
| Infections in the last 6 months (Median) | 1 | 0 | MW=20.789 | p<0.0001 |
| Months since the last infection (Median) | 12 | 24 | MW=20.055 | p<0.0001 |
| Diagnosis | ||||
| Negative (n) | 106 | 164 | ||
| P. vivax (n) | 6 | 16 | ||
| P. falciparum (n) | 10 | 8 | ||
We consider statistically significant p values lower than 0.05, the differences in medians for the study population data were tested by non-parametric Mann-Whitney, normalizing transformations were performed on raw data before testing by one-way analysis of variance where appropriate and differences in proportions were evaluated by chi-square (χ2) test.
Not statistically significant.
2.4. PvMSP-9 recombinant proteins
The N-terminal domain (aa 34–367) and the second block of tandem repeats (aa 874–972) of PvMSP-9 (Belem strain) were expressed in pGEX (Amersham-Pharmacia) as GST fusion proteins, PvMSP9-Nt and PvMSP9RII, as described (29). The C-terminal domain was similarly expressed as a GST fusion protein. Briefly, the C-terminal region of the msp9 gene containing the repeated blocks I and II (aa 729–972) was amplified by PCR using P. vivax Belem strain genomic DNA as a template and forward and reverse primers containing the ATTB1 and ATTB2 recombination sites, respectively. The Gateway System (Invitrogen, Carlsbad, CA) was used to prepare the expression clone following the manufacturer’s instructions. The PCR product was then subcloned into the shuttle vector pDONR 201 by incubating overnight at 25°C in the presence of the BP clonase enzyme, creating the BRI-RII/pDONR entry clone. The one-tube reaction was followed by adding the destination vector pDEST 15 (N-terminal Glutathione-S Tranferase- tag) and the LR clonase to the entry clone. Library efficiency DH5 competent cells were transformed with the whole mixture and transformant clones were selected by plating on ampicillin (100 μg ml-1)/LB plates. Positive clone were confirmed by big dye terminator v3.1 sequencing technology and used for protein expression. rPvMSP9-BRI-RII was expressed in the E. coli strain BL21 salt-inducible (SI). The expression clone was inoculated into 40 ml of 2× TY liquid medium containing 100 μg ml-1 of ampicillin. Cells were cultured at 280 rpm overnight, then diluted 1:10 and grown to OD600=0.8 at 37°C. Induction was started by addition of NaCl to a final concentration of 0.3M and shaking at room temperature for 16 hours. The cells were then centrifuged and the pellet was suspended in phosphate buffered saline (PBS) containing 0.25% Triton X100 and protease inhibitors, sonicated and centrifuged. The soluble fraction was incubated with pre-washed Glutathione Sepharose 4B (Amersham, Piscataway NJ) beads in batch purification. Prior to elution, the beads were washed three times with PBS and fractions were eluted at 4°C using 10mM reduced glutathione in 50mM Tris pH 8.0 buffer. Before quantization of the total protein, fractions were pooled and dialyzed against three changes of PBS. A schematic representation of the localization of the PvMSP9 recombinant constructs is shown in Figure 1.
Figure 1.

Schematic representation of PvMSP9, the constructs of recombinant proteins used for serological analysis (continuous lines) and the synthetic peptides used for cellular analysis (discontinuous lines).
2.5. Prediction of PvMSP9 promiscuous T cell epitopes
In silico analysis was used to predict putative class II restricted T cell epitopes. The amino acid sequence of PvMSP9 was scanned to identify promiscuous MHC binding peptides using virtual matrices designed for 51 HLA-DR alleles [30]. Five peptides (including 3 overlapping sequences) were identified within the N-terminal region of PvMSP9 which were predicted to bind at least 40% HLA-DR alleles tested at a 3% threshold. Synthetic peptides representing such putative T cell epitopes were synthesized at the Laboratory of Biochemical of Proteins and Peptides, Institute Oswaldo Cruz, Fiocruz. The complete sequences of five out of 11 synthetic peptides that induced the highest cellular response were: 1) peptide pE (V147–K159), VVHKLNKKMKSLK; 2) peptide pH (V438–D449), VSLMASIDSMID; 3) peptide pJ (K325–I339), KLKDILLRVLYKTYI; 4) peptide pK (P434–I448), PAEDVSLMASIDSMI and 5) peptide pL, (A443–K456), ASIDSMIDEIDFYEK.
2.6. Antibody Assays
Plasma samples from study participants were screened for the presence of naturally acquired antibodies against the three recombinant proteins PvMSP9-RIRII, PvMSP9-RII, PvMSP9-NT and purified SjGST by ELISA. Briefly, maxisorp 96-well plates (Nunc, Rochester, NY) were coated with 2 μg per ml of each recombinant protein or GST alone as control antigen. After overnight incubation at 4°C, plates were washed with PBS containing 0.05% Tween 20 (PBS-Tween) and blocked with PBS-Tween containing 5% non fat dry milk (PBS-Tween-M) for 2 h at 37°C. Individual plasma samples diluted 1:100 PBS-Tween-M were added in duplicate wells and the plates incubated at room temperature for 1 h. After four washes with PBS-Tween, bound antibodies were detected with peroxidase-conjugated goat anti-human IgG (Sigma St. Louis, MO) followed by o-phenylediamine and hydrogen peroxide. The absorbance was read at 492 nm using an ELISA reader (Spectramax 250, Molecular Devices, Sunnyvale, CA) and specific reactivity was obtained by subtraction of the averaged OD value due to GST alone from the averaged OD value of the same plasma for the recombinant protein. The results for total IgG were expressed as Reactivity Indexes (RI) that were calculated by dividing the mean optical density of tested samples by the mean optical density plus 3 standard deviations of 24 non exposed controls. Subjects were scored positive for serum IgG to a particular antigen if the RI was higher than 1. For determination of IgG subclasses the following peroxidase conjugated monoclonal mouse anti-human antibodies were used: clones HP-6001 for IgG1, HP-6002 for IgG2, HP-6050 for IgG3 and HP-6023 for IgG4 (Sigma) and has been used previously to characterize IgG subclasses in the studied population. Subclass-specific prevalence for each antigen was determined from OD values using 3 SD above the appropriate mean OD of 24 non exposed controls as the cut-off for positivity. To adjust the affinity differences between the IgG subclasses, standard curves were prepared using human IgG kappa myeloma proteins (Sigma) from each of the four subclasses. In addition, standard curves enabled conversion of OD values to concentration (mg/ml) for the comparison of different subclasses. Purified human IgG kappa myeloma proteins from each of the four subclasses (Sigma) were coated overnight at 4°C in PBS at 100 μl per well onto 96-well plates in 1:2 serial dilutions from 24 μg/ml to 2−11 μg/ml. After washing four times, plates were incubated with the appropriate anti-human IgG subclass-specific mAb, washed four times, incubated with peroxidase-labeled goat anti-mouse antibody, washed a final four times, developed with o-phenylediamine and hydrogen peroxide, and measured as described above for subclass-specific ELISA. Subclass-specific OD values were converted to concentration values (μg/ml) using sigmoidal curve-fit equations derived from subclass-specific standard curves. The threshold of positivity was the mean plus 3 SD of concentration values of sera from 24 non exposed controls.
2.7. ELISPOT assays
Cells cultures were carried out in duplicate in nitrocellulose 96 well plates (MAHA S4510 Millipore) coated over night at 4°C with 5 μg/mL capture anti-IFN-γ monoclonal antibodies (MabTech, Stockholm – Clone D1K) or anti-IL-4 (Pharmingen, San Jose, CA -Clone MP4-25D2) in PBS. The plates were blocked with RPMI medium containing 10% fetal calf serum for at least 2 h. 2.5×105 cells were added to the ELISPOT plates in the presence of medium alone, 10 μg/ml of each PvMSP9 peptide or phytohemaglutinin (1mg/mL). Cells were stimulated for 24 h for IFN-γ or 48 h for IL-4 at 37°C, 5% CO2 under sterile conditions. After stimulation, plates were washed four times with PBS containing 0.05% Tween 20 (PBS-T) and incubated with either biotin-anti-human IFN-γ Clone 7-B6-1 (MabTech) diluted in PBS or biotin-anti-human IL-4 Clone 12-1 NON0059 (Biosource International, Camarilla, CA) diluted in PBS-T containing 1% fetal bovine serum (PBS-TF) for 3 h at 37°C. The plates were washed four times with PBS-T and incubated with streptavidin-alkaline phosphatase (MabTech) in PBS-TF for 1 h at 37°C. The plates were washed four times with PBS-T before development with 1-step NBT/BCIP (Pierce, Rockford, IL). Development was stopped by the addition of distilled water. IFN-γ and IL-4 secreting cells appeared as blue spots that were counted with an Immunospot reader (Cellular Technology Ltd, Cleveland, OH) using the Immunospot Software Version 3. ELISPOT responses were expressed as net spot-forming cells (SFC) per 2.5 × 105 PBMCs. Thirty non-exposed individuals were also included in the Elispot test. The assay was subsequently categorized as positive or negative depending on whether the mean number of SFC in the peptide stimulated wells was greater than the mean number plus twice the SD of SFC in the control wells with medium alone from the same donor. When the variability in peptide stimulated wells were very low and control wells were 0 (zero), we imposed the requirement of a peptide stimulated wells average of at least 20 for IFN-γ and 10 for IL-4 more SFCs than control well. This arbitrary value was based in the average numbers of SFCs of 30 non-exposed individuals that ranged from 4.3–9.1 SFCs for IFN-γ (pE= 8.7, pH=4.3, pJ=5.1, pK=7.0, pL=9.1) and 2.8–5.4 for IL-4 SFC (pE=4.3, pH=5.4, pJ=3.4, pK=4.7, pL=2.8).
2.8. Statistical analysis
Analyses were done using Epi Info 2002 (CDC, Atlanta, GA), Prism 4.0 and Instat (GraphPad Software, San Diego, CA) according to the required statistical test. Differences in medians for the study population data were tested by non-parametric Mann-Whitney test when appropriate. Student’s t test was used to compare the means of normally distributed data or normalized transformations were performed on raw data before testing by one-way analysis of variance where appropriate. Differences in proportions were evaluated by chi-square (χ2) test. Relationships between year of residence in the endemic area and number of past malaria episodes or months since last known malaria episode were assessed with Spearman’s rank correlation. Multivariate logistic regression was used to assess the relationship between antigen-specific total IgG responses and the independent variables of gender, age, years of residence in the endemic area, number of past malaria episodes, and months since last known malaria infection.
3. Results
3.1. Epidemiological data
As shown in Table 1, the two community populations are comparable in gender, median age and number of past recalled malaria episodes. They do, though, significantly differ in years of residence in malaria endemic areas and in state of Rondonia, the number of months elapsed since the last malaria episode and the number of malaria infections in the last 6 months. Colina has a significantly higher median number of infections in the last six months and fewer months since the last infection. At the time of the cross-sectional study, P. vivax was the predominant parasite in Ribeirinha, while P. falciparum predominated in Colina. In Ribeirinha we found five individuals with a malaria infection, which were asymptomatic at the time of blood collection (four individuals infected with P. vivax and one with P. falciparum). In Colina all infected individuals found also presented with symptoms of malaria.
3.2. Frequency and reactivity indexes of IgG antibodies to recombinant proteins derived from PvMSP9
The prevalence of anti-PvMSP9 antibodies specific for the three recombinant proteins derived from PvMSP-9 was determined with plasma of 185 individuals from Ribeirinha and 121 from Colina. The results show that a total of 74% of the 306 subjects in both communities presented with IgG antibodies to at least one of the recombinants proteins, which indicated that these proteins are immunogenic during natural human infections. The majority of the individuals in the Ribeirinha community (85%) had IgG antibodies to at least one of the recombinant proteins, while in the Colina community only 58% (χ2=30.247; p<0.0001) were positive to at least one antigen. In Ribeirinha the prevalence of antibodies was significantly higher for all the recombinant proteins tested with the exception of antibodies to PvMSP9-RII, which was similar in both localities (Fig. 2a). The frequency of positive individuals for the recombinant protein consisting of both blocks of repeats (PvMSP9-RIRII) was 73% in Ribeirinha and 50% in Colina (χ2=15.205; p<0.0001). The recombinant protein that included only the second block of repeats (PvMSP9-RII) was positive in 42% of Ribeirinha population and 40% of individuals in Colina (χ2=0.049; p = 0.8252). The frequencies of positive individuals for the N-terminal region (PvMSP9-NT) were 49% in Ribeirinha compared to 16% in Colina (χ2 =33.204; p<0.0001). The magnitude of IgG antibody responses was compared by using the average RI values of positive individuals for each recombinant protein (Fig. 2b). The RI against the recombinant proteins varied from 0.44 to 8.97 and was higher in Ribeirinha than in Colina. The average RI of antibodies against PvMSP9-RIRII in positive individuals from Ribeirinha (2.68 ± 1.95) was significantly higher than the observed in Colina (1.76 ± 0.92; t=4.385 df=194 p<0.0001). The differences between the averages of PvMSP9-RII antibodies was also significant between Ribeirinha (2.95 ± 2.11) and Colina (1.76 ± 0.92; t=2.666 df=123 p=0.0087) and PvMSP9-NT (Ribeirinha = 1.88 ± 0.88 and Colina = 1.22 ± 0.43; t=2.084 df=107 p=0.0212). None of the 24 healthy individuals without previous history of exposure to malaria infections had antibodies against the three recombinant proteins.
Figure 2.

Antibody responses to the recombinant proteins PvMSP9-RIRII, PVMSP9-RII and PvMSP9-Nt in malaria exposed individuals from Colina (CO) and Ribeirinha (RI). (a) Frequency of IgG positive individuals to the recombinant proteins. (*) indicate that the difference was significant (p<0.0001) for a comparison between the frequency of responders from Colina with Ribeirinha by χ2 test; (b) IgG antibodies measured as mean reactivity indexes in responders (mean ± 95% confidence interval). (+) indicate that the difference was significant (p<0.05) for a comparison between the mean RI values responders from Colina with Ribeirinha by Student’s t test; (c) Frequency of IgG subclass distribution in IgG positive individuals. (#) indicates that the difference was significant (p<0.01) for a comparison between a particular IgG subclass over the others IgG subclasses for the same recombinant protein.
3.3. IgG subclasses distribution to PvMSP9 recombinant proteins
The distribution of IgG isotypes was analyzed in the plasma of individuals who were identified being positive for IgG antibodies to each recombinant protein. Antibodies of all IgG subclasses could be detected to the different recombinant proteins even though the predominance of a particular IgG subclass varied for different recombinant proteins and populations (Fig. 2c). Using prevalence as the basis for each recombinant protein in the population as a whole, IgG1 was predominant for PvMSP9-RIRII (63%; p<0.01) and PvMSP9-NT (60%; p<0.0001) while IgG2 was more prevalent for PvMSP9RII (63%; p<0.001). A comparison between IgG subclass distributions in the two communities is presented in Table 2. IgG1 was predominant for PvMSP9-RIRII and PvMSP9-Nt in Ribeirinha whereas in Colina similar frequencies were observed in all IgG subclasses except for IgG4, which was lower in both communities. The subclass distribution for PvMSP9-RII showed a similar pattern in both communities with predominance of IgG2 (Ribeirinha 64% and Colina 63%). Analysis of antibodies levels shows that when concentration values were compared, IgG1 and IgG2 positive responses predominate over other IgGs reactivity with all three recombinant proteins both in Colina and in Ribeirinha. However the mean antibody concentration in Ribeirinha was significantly higher for IgG1 to all recombinant proteins, for IgG2 to PvMSP9-RII and for IgG3 to PvMSP9-RIRII and PvMSP9-RII. No differences were observed in the concentration of IgG4 to all recombinant proteins.
Table 2.
Comparison of IgG subclass-specific concentration against PvMSP9 recombinant proteins in IgG positive individuals in Colina and Ribeirinha communities.
| Recombinant Proteins | Colina (95%-CI) |
Ribeirinha (95%-CI) |
pc | Controlsb (95% CI) |
|---|---|---|---|---|
| PvMSP9-RIRII | n=61 | n=135 | n=51 | |
| IgG1 | 0.306 – 0.485 | 0.557 – 0.687 | 0.025 | 0.120 – 0.131 |
| IgG2 | 0.284 – 0.439 | 0.460 – 0.636 | NSd | 0.095 – 0.109 |
| IgG3 | 0.147 – 0.161 | 0.275 – 0.378 | <0.0001 | 0.100 – 0.117 |
| IgG4 | 0.065 – 0.108 | 0.107 – 0.128 | NS | 0.049 – 0.058 |
|
| ||||
| PvMSP9-RII | n=48 | n=77 | ||
| IgG1 | 0.286 – 0.374 | 0.430 – 0.517 | 0.0133 | 0.129 – 0.139 |
| IgG2 | 0.276 – 0.377 | 0.420 – 0.502 | 0.0025 | 0.095 – 0.107 |
| IgG3 | 0.119 – 0.155 | 0.218 – 0.126 | 0.0091 | 0.098 – 0.111 |
| IgG4 | 0.063 – 0.100 | 0.080 – 0.125 | NS | 0.049 – 0.061 |
|
| ||||
| PvMSP9-NT | n=19 | n=90 | ||
| IgG1 | 0.193 – 0.320 | 0.324 – 0.399 | 0.0356 | 0.121 – 0.139 |
| IgG2 | 0.201 – 0.277 | 0.271 – 0.354 | NS | 0.090 – 0.097 |
| IgG3 | 0.179 – 0.353 | 0.168 – 0.199 | NS | 0.098 – 0.109 |
| IgG4 | 0.130 – 0.141 | 0.121 – 0.150 | NS | 0.055 – 0.060 |
Optical densities (OD) values were converted using standard curves for each IgG subclass (see Material and Methods for details) and 95% confidence intervals of means μg/ml are shown.
All concentration values of non-malaria-exposed donors were significantly lower (p<0.001) when compared with the subclass concentrations of exposed individuals of both communities.
Student’s t test was used to compare differences between Colina and Ribeirinha within each IgG subclass for a single antigen. Statistically significant differences between groups are represented by bold typeface fonts. Log or reciprocal transformations were performed as appropriate to normalize the distribution of data points before parametric statistical analyses.
Not statistically significant (NS).
3.4. Relationship between antibody responses and epidemiological data
To assess if antibody reactivity to the recombinant proteins was associated with epidemiological data related to malaria exposure we calculated Spearman’s correlation coefficients between antibody responses and gender, age, years of residence in malaria endemic area, total number of past malaria episodes, infections in the last 6 months and months since the last infection. In the two localities studied gender did not affect the total IgG antibody response to any of the recombinant proteins. In Ribeirinha, age correlated positively with the IgG reactivity index to PvMSP9-RIRII (r=0.370; p<0.0001) and PvMSP9-RII (r =0.334; p<0.0001) and with IgG1 concentration levels for PvMSP9-Nt (r=0.178; p=0.0028). In our study all volunteers were older than 11 years old and age related changes could not be fully evaluated.
Using years of residence in endemic area and number of past malaria infections reported by donors to define the level of malaria exposure, we observed a direct correlation between time of residence in an endemic area and the reactivity index of IgG antibodies (Fig. 3) for PvMSP9-RIRII and PvMSP9-RII in Colina (r=0.671; p=0.0072 and r=0.718; p=0.0135) and in Ribeirinha (r=0.730, p<0.0001 and r=0.877, p=0.01) whereas a positive correlation between the number of previous episodes of malaria and the reactivity index of IgG antibodies to PvMSP9-RIRII and PvMSP9-RII was observed only in Colina (r=0.278; p<0.0001 and r=0.273; p<0.0001). No association was observed between total IgG positive individuals or RI of antibodies to PvMSP9-Nt in relation to age, years of residence in malaria endemic area, total number of past malaria episodes, infections in the last 6 months, and months since the last infection in both communities. However, years of residence in endemic areas were positively correlated with IgG1 concentration for PvMSP9-Nt in Ribeirinha (r=0.127; p=0.02) and in Colina (r=0.178; p<0.02).
Figure 3.

Spearman’s rank correlation between time of residence in malaria endemic area and the IgG reactivity index against PvMSP9-RIRII and PvMSP9-RII in Colina (a and c) and Ribeirinha (b and d) population.
In order to evaluate if antibody reactivity was associated with protection we used the number of infections in the last 6 months and months since the last infection as indicators of level of protection. In this analysis individuals who either had detectable levels of parasitemia or reported as malaria-naive were excluded. The analysis demonstrated that a positive correlation was observed only between IgG reactivity to PvMSP9-RIRII and months since the last infection in individuals from Ribeirinha (r=0.298; p=0.046).
3.5. IFN-γ and IL-4 responses to PvMSP9 synthetic peptides
Five synthetic peptides, predicted to bind more than 40% of the HLA-DR alleles tested, elicited IFN-γ and IL-4 recall responses. The overall frequencies of IFN-γ and IL-4 responders to at least one of the peptides were 58 % and 41%, respectively. However, as can be seen in Figure 4 the frequencies of positive responses induced by different peptides varied. In Ribeirinha, the frequency of IFN-γ positive individuals was similar for all the peptides while the most frequent response in IL-4 positive individuals was obtained with peptides pE (38%) and pL (37%). In Colina the most frequent IFN-γ responses were obtained with peptides pE (46%) and pL (43%) while IL-4 responses were obtained with peptide pE (14%) and pH (17%). When the pattern of the responses was examined, significant differences were observed between the two communities. In the native population from Ribeirinha, the frequency of positive individuals was similar for IFN-γ and IL-4 responders except for the peptides pE and pL which presented a higher proportion of IL-4 positives individuals when compared to IFN-γ positive individuals (pE: χ2=32.460, p<0.0001 and pL: χ2=6.829, p<0.009). In contrast, the Colina transmigrant population response was biased towards IFN-γ production. The frequency of IFN-γ responses was significantly higher than the frequency of IL-4 to all peptides (pE: χ2=32.365, p<0.0001; pH χ2=6.898, p=0.009; pJ: χ2=12.023, p=0.0001; pK: χ2=20.104, p<0.001; pK: χ2=24.631, p<0.0001). Similarly, the frequency of IL-4 positive individuals in Ribeirinha was significantly higher than in Colina to all peptides (pE: χ2=19.987, p<0.0001; pH: χ2=7.256, p=0.0071; pJ: χ2=17.465, p<0.0001; pK: χ2=9.123, p=0.0025; pL: χ2= 6.968, p=0.0082) while no differences were observed in the frequencies of IFN-γ. In responders, the number of IFN-γ SFC elicited by the peptides in Ribeirinha ranged from 77.1±72.9 to 115.5±146.2 and in Colina 54.3±51.9 to 85±60 SFC. The numbers of IL-4 SFC in responders were lower than the number of IFN-γ SFC and ranged from 23.7±11.5 to 30.1±9.9 in Ribeirinha and 15.5±5.2 to 28.6±5.1 in Colina. Nevertheless individuals from both communities responded equally well to PHA (753±220 IFN-γ and 250±160 IL-4 SFC) and no significant differences were observed between donors.
Figure 4.
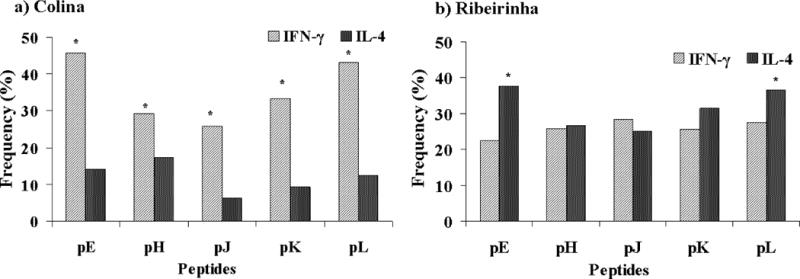
Elispot IFN-γ and IL-4 responses to PvMSP9 synthetic peptides (pE, pH, pJ, pK and pL) in individuals naturally exposed to malaria infection from Colina and Ribeirinha. Frequency of IFN-γ and IL-4 responders for each peptide in individuals from Colina (a) and Ribeirinha (b). (*) Indicates that the difference was significant (p<0,01) for a comparison between the frequency of IFN-γ responders with IL-4 responders for each peptide calculated by χ2 test.
3.6. Relationship between cellular response and humoral immune response
No association was observed between IFN-γ SFC and levels of IgG to all recombinant proteins in both communities. In contrast, in Ribeirinha, the numbers of IL-4 SFC induced by peptides pK and pL correlated positively with the levels of IgG to PVMSP9-RIRII (pK: r=0.707 p=0.0376; pL: r=0.790 p=0.0108) and PvMSP9-RII (pK: r=0.703, p=0.0296; pL: r=0.632 p=0.0159). A positive correlation was also observed in Ribeirinha between the time of residence in Rondonia state and the number of IL-4 responses against pL (r=0.185; p=0.0055).
4. Discussion
Knowledge of the natural immune response to malaria antigens is a prerequisite for the optimal design of a vaccine. Despite being a potential target for vaccine development this is the first report on the naturally acquired humoral and cellular immune responses to PvMSP9 in individuals exposed to malaria infections. In a cross-sectional study carried out in Porto Velho, Rondonia state, Brazil, we assessed naturally acquired humoral and cellular immune responses using cells and sera from individuals living in Ribeirinha, a riverine native community and Colina, a transmigrant community. The two communities were comparable in gender, median age and number of past malaria episodes. However, they significantly differed in years of residence in a malaria endemic area and in Rondonia, months since their last malaria episode, and number of malaria infections in the last 6 months. The presence of asymptomatic infection in Ribeirinha reported here and elsewhere in natives of the riverine community of Rondonia indicates that subjects exposed to both P. vivax and P. falciparum malaria in Brazil also develop acquired resistance to clinical malaria despite different epidemiological and lower transmission levels profile compared to those seen in Africa [31, 32].
Our data shows that the C-terminal and the N-terminal domains of the PvMSP9 are targets of the immune response in individuals naturally exposed to P. vivax malaria transmission. The presence of antibodies in the plasma of individuals that recognize the three recombinant proteins representing PvMSP9, indicate that the expression systems successfully generated epitopes that share similar antigenic determinants as the natural proteins expressed in merozoites. In a previous study we demonstrated that serum from mice immunized with these recombinant proteins recognized the native protein [27]. The overall prevalence of antibodies in individuals that recognize at least one of the recombinant proteins was 74%, with (PvMSP9-RIRII) being the most immunogenic. In Ribeirinha, the frequency of responders and the reactivity indices of antibodies were significantly higher for the majority of the recombinant proteins. The IgG reactivity index to PvMSP9-RIRII and PvMSP9-RII correlated positively with time of residence in endemic region in both localities while correlation with age was observed only in Ribeirinha and the number of previous episodes of malaria only in Colina. The association between antibody responses with age and time of residence in endemic region is a phenomenon which has been frequently reported for various antigens [28, 29, 33, 34], reflecting most likely exposure to malaria parasite and possibly maturation of the immune system over time. However, since in our study mainly adults were studied, age related changes in antibody responses in Ribeirinha could reflect length of exposure as age in this native population represents time of exposure in an endemic region. Curiously, individuals from Ribeirinha had a comparable number of reported past malaria episodes as individuals in Colina despite being natives of the endemic region. However, no significant association was found between number of past malaria episodes and antibody responses. This could be due to imprecise donor recall since individuals from Ribeirinha, may not remember childhood infections or past misdiagnosed malaria episodes or asymptomatic infections. In contrast, individuals from Colina, the migrant population, living for a shorter period of time in the area have likely experienced clinical episodes more recently.
Studies on the humoral immune responses to P. falciparum antigens have consistently shown that immunity to blood-stage antigens is dependent on a specific pattern of immunoglobulin subclass response. Pattern of IgG subclasses consisting of IgG1 and/or IgG3 antibodies to several blood-stage antigens are effective mediators of antibody-dependent cellular inhibition of malaria parasites growth in vitro and have been associated with the acquisition of clinical immunity to malaria [22, 35–37]. However, studies on IgG subclass responses to P. vivax blood-stage antigens in subjects living in endemic areas remain scarce. In Brazil, only a small number of studies have investigated cytophilic antibody responses against P. vivax antigens and the majority is to PvMSP-1 [23, 29, 34, 38–40]. Our results show that using prevalence as a basis, all four subclasses of IgG were detected in both communities but the relative proportion of these subclasses differed for different recombinant proteins. IgG1 was predominant for PvMSP9-RIRII and PvMSP9-NT in Ribeirinha while mixed IgG1, IgG2 and IgG3 responses were observed in Colina for both recombinant proteins. Interestingly, IgG2 polarization was evident for PvMSP-RII in both communities suggesting that the response is epitope driven and seems to be little affected by the time of exposure. However, taking into account the IgG subclasses concentrations as the basis, IgG1 and IgG2 were predominant to all recombinant proteins in both communities although in Ribeirinha the concentrations were significantly higher and IgG1 concentrations to PvMSP9-Nt correlated positively with time of residence and age. Previously reported studies in Brazil showed that IgG1 and IgG3 antibodies to PvRBP1 in the same area were associated with time of residence in the endemic area and the number of months since the last malaria episode suggesting a protective effect of these antibodies [29]. An IgG1 polarized response was also reported to PvAMA1 and this response increased with the number of malaria episodes [41]. In contrast, Morais et al.[42] showed that IgG1 antibodies to PvAMA-1 persisted for seven years among subjects briefly exposed to malaria in an outbreak. From data on subclass antibody responses to the C-terminal of PvMSP-1, Soares et al.[43] showed predominance of IgG1 and IgG3 associated with time of exposure while Morais et al. [42] detected low prevalence of IgG1 and IgG3 among individuals with long term exposure when compared to subjects less or sporadically exposed. Similarly, data with P. falciparum MSP1 antigen reported in malaria endemic areas of Brazil showed that IgG1 to PfMSP-1 was significantly higher among subjects with long term exposure and in asymptomatic infections [44] while others have reported the response was IgG3 biased being little affected by cumulative or current exposure [45].
In our study, in both communities, IgG3 reactivity with the recombinant proteins was not predominant, while high prevalence and levels of IgG1 to PvMSP9-RIRII and PvMSP9-Nt were observed in individuals from Ribeirinha suggesting that cumulative exposure in this native population may account for this profile and may have some protective effect. However, only a weak association was found between IgG1 levels to PvMSP9-RIRII with months since the last malaria episode (r=0.2977; p=0.046). Although this association may suggest a protective effect for this subclass, it was not strong and we are not confident that the majority of individuals recollect precisely the data of their last malaria episode, especially if it occurred more than a year ago. The predominance of IgG2 antibodies reacting with PvMSP9-RII in both communities, although they differ in time of residence in endemic area, seem to be little affected by the time of exposure and could be epitope driven raising the question whether intrinsic features of this block of repeats influences the subclass switching.
Based on in vitro evidence, it has been suggested that IgG2 antibodies compete with IgG1 and IgG3 and interfere with protective immunity [35]. If cytophilic antibodies participate in the protective immunity in P. vivax as has been observed for P. falciparum, it is plausible to argue that the predominance of IgG1 antibodies to the repetitive region (PvMSP9-RIRII) and N-terminal region (PvMSP9-Nt) may participate (at some level) in the elimination of blood stage of P. vivax while the non-cytophilic IgG2 response induced to the second block of repeats (PvMSP9-RII) would interfere with this process. However, Aucan et al. [46] showed that IgG2 specific to P. falciparum RESA and MSP2 are associated with low risk for contracting malaria infection suggesting that IgG2 could be associated with protection. Therefore, the role of antibodies to P. vivax in the development of clinical immunity is still unclear and the effect of each IgG subclass even in P. falciparum is controversial. Studies that evaluate the profile of protective immune responses mediated by cytophilic antibodies are necessary.
Cell-mediated immunity is highly effective in mice with malaria and data from humans show that parasite-specific CD4+ T cells can inhibit parasite growth in vitro [47]. Therefore antigens that are capable of inducing a cellular immune response should be included in malaria vaccine formulation. The existence of functionally distinct CD4 T cells in humans to P. falciparum antigens is indicated by results of in vitro studies with PBMC from donors naturally exposed to P. falciparum [48–50]. Unlike with P. falciparum, very few studies have investigated cellular immune responses to P. vivax antigens and this is the first one with PvMSP9 in humans. We have demonstrated that peptides from the N-terminal region of PvMSP9 induce IFN-γ and IL-4 cytokine recall responses in a significant proportion of individuals indicating the presence of T cell epitopes. The fact that BALB/c mice immunized with the recombinant protein PvMSP9-Nt and individuals naturally exposed to malaria infection exhibited a cellular immune response to PvMSP9-Nt supports the conclusion that this conserved sequence contains helper T-cell epitopes. Indeed, the N-terminus of PvMSP9 is particularly important because it is highly conserved among Plasmodium species [17]. Although we did not determine the HLA haplotype of the individuals, it seems plausible that they represented more than one haplotype since all 5 of the peptides and to some extend the peptides pE, pL, pK and pH appear to have the ability to bind to genetically restricted different HLA haplotypes since they stimulated T-cells to secrete IFN-γ and IL-4 in both natives from the same region in the Amazon and migrant population from different non-malaria regions of Brazil. If that indeed is the case, we would have four promiscuous T-helper cell epitopes confirmed from an asexual blood-stage protein of the parasite.
The response elicited by at least one of the peptides was high for IFN-γ (58%) and for IL-4 (41%) and the numbers of IFN-γ SFC were similar in both communities and were higher than for IL-4. However, the pattern of the response was different between the two communities, where the native population from Ribeirinha, most of the peptides induced both IFN-γ and IL-4 secretion. However, in migrant population from Colina, the response was biased towards IFN-γ production. It is possible that variations in exposure may also explain variations in the type of naturally induced TH1 and TH2 immune responses. Indeed, IL-4 SFC correlated positively with antibody levels to the recombinant proteins and time of residence in the endemic area in Ribeirinha. Our results corroborate with data reported by Troye-Blomberg et al.[51], showing that IL-4 responses to blood-stage antigens are associated with elevated levels of serum antibodies. Farouk et al. [52] measured cellular response to crude P. falciparum antigens by ELISPOT and found a strong association between high IgG and IgE antibodies with increased numbers of IL-4 secreting cells in individuals less susceptible to malaria infection. Similarly, correlations between the production of IL-4 in response to the P. falciparum malaria antigen Pf55RESA and protection against malaria were also reported [53]. Thus, the higher percentage and levels of IL-4 SFC and the correlation between an IL-4 response to pK and pL peptides with IgG levels in Ribeirinha may explain the differences in antibody levels observed between the two communities. Whether naturally induced IL-4 responses associated with elevated antibody levels in Ribeirinha reported in our study is a reflection of immune status is presently unknown.
In conclusion, this study describes for the first time naturally acquired cellular and antibody responses to PvMSP-9 in endemic populations. We have demonstrated that PvMSP9 contains B and T-cell epitopes recognized by antibodies and T cells from individuals in the Brazilian Amazon. The antibody frequency and levels, and malaria-specific IL-4 and IFN-γ responses differed between communities studied and seem to be associated with level of exposure. It still remains to be established whether these responses are indeed protective. The results presented here provide insightful data relevant for understanding potential targets and protective immune responses against P. vivax and further studies towards evaluating PvMSP9 as a vaccine candidate are therefore warranted.
Acknowledgments
This work was supported by Brazilian National Research Council – CNPq, Fiocruz, National Institute of Health, Grant RO1 AI0555994 and the Yerkes Regional Primate Research Center Base Grant. Josue da Costa Lima Junior was the recipient of a CNPq Fellowship.
We are grateful to all individuals that participate in this study for their cooperation and generous donation of blood, which made this study possible. We tank to Eileen Farnon and Jennie Larson for the assistance during the sample collection and Ramira Ribeiro for help with Elisa assays. We thank the Secretary of Health of Rondonia State and the Laboratorio Central – LACEN of Rondonia for providing fieldwork support and the Program for Technological Development in Tools for Health-PDTIS/FIOCRUZ for use of its facilities.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Price RN, Tjitra E, Guerra CA, Yeung S, White NJ, Anstey NM. Vivax malaria: neglected and not benign. Am J Trop Med Hyg. 2007 Dec;77(6 Suppl):79–87. [PMC free article] [PubMed] [Google Scholar]
- 2.Baird JK. Neglect of Plasmodium vivax malaria. Trends Parasitol. 2007 Nov;23(11):533–9. doi: 10.1016/j.pt.2007.08.011. [DOI] [PubMed] [Google Scholar]
- 3.Krotoski WA, Collins WE, Bray RS, Garnham PC, Cogswell FB, Gwadz RW, et al. Demonstration of hypnozoites in sporozoite-transmitted Plasmodium vivax infection. Am J Trop Med Hyg. 1982 Nov;31(6):1291–3. doi: 10.4269/ajtmh.1982.31.1291. [DOI] [PubMed] [Google Scholar]
- 4.Canessa A, Mazzarello G, Cruciani M, Bassetti D. Chloroquine-resistant Plasmodium vivax in Brazil. Trans R Soc Trop Med Hyg. 1992 Sep-Oct;86(5):570–1. doi: 10.1016/0035-9203(92)90120-2. [DOI] [PubMed] [Google Scholar]
- 5.de Santana Filho FS, Arcanjo AR, Chehuan YM, Costa MR, Martinez-Espinosa FE, Vieira JL, et al. Chloroquine-resistant Plasmodium vivax, Brazilian Amazon. Emerg Infect Dis. 2007 Jul;13(7):1125–6. doi: 10.3201/eid1307.061386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Nayar JK, Baker RH, Knight JW, Sullivan JS, Morris CL, Richardson BB, et al. Studies on a primaquine-tolerant strain of Plasmodium vivax from Brazil in Aotus and Saimiri monkeys. J Parasitol. 1997 Aug;83(4):739–45. [PubMed] [Google Scholar]
- 7.Polley SD, McRobert L, Sutherland CJ. Vaccination for vivax malaria: targeting the invaders. Trends Parasitol. 2004 Mar;20(3):99–102. doi: 10.1016/j.pt.2003.12.003. [DOI] [PubMed] [Google Scholar]
- 8.Wipasa J, Elliott S, Xu H, Good MF. Immunity to asexual blood stage malaria and vaccine approaches. Immunol Cell Biol. 2002 Oct;80(5):401–14. doi: 10.1046/j.1440-1711.2002.01107.x. [DOI] [PubMed] [Google Scholar]
- 9.Galinski MR, Medina CC, Ingravallo P, Barnwell JW. A reticulocyte-binding protein complex of Plasmodium vivax merozoites. Cell. 1992 Jun 26;69(7):1213–26. doi: 10.1016/0092-8674(92)90642-p. [DOI] [PubMed] [Google Scholar]
- 10.Chitnis CE, Sharma A. Targeting the Plasmodium vivax Duffy-binding protein. Trends Parasitol. 2008 Jan;24(1):29–34. doi: 10.1016/j.pt.2007.10.004. [DOI] [PubMed] [Google Scholar]
- 11.Tsuboi T, Kappe SH, al-Yaman F, Prickett MD, Alpers M, Adams JH. Natural variation within the principal adhesion domain of the Plasmodium vivax duffy binding protein. Infect Immun. 1994 Dec;62(12):5581–6. doi: 10.1128/iai.62.12.5581-5586.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wertheimer SP, Barnwell JW. Plasmodium vivax interaction with the human Duffy blood group glycoprotein: identification of a parasite receptor-like protein. Exp Parasitol. 1989 Nov;69(4):340–50. doi: 10.1016/0014-4894(89)90083-0. [DOI] [PubMed] [Google Scholar]
- 13.Igonet S, Vulliez-Le Normand B, Faure G, Riottot MM, Kocken CH, Thomas AW, et al. Cross-reactivity studies of an anti-Plasmodium vivax apical membrane antigen 1 monoclonal antibody: binding and structural characterisation. J Mol Biol. 2007 Mar 9;366(5):1523–37. doi: 10.1016/j.jmb.2006.12.028. [DOI] [PubMed] [Google Scholar]
- 14.Hodder AN, Crewther PE, Matthew ML, Reid GE, Moritz RL, Simpson RJ, et al. The disulfide bond structure of Plasmodium apical membrane antigen-1. J Biol Chem. 1996 Nov 15;271(46):29446–52. doi: 10.1074/jbc.271.46.29446. [DOI] [PubMed] [Google Scholar]
- 15.del Portillo HA, Longacre S, Khouri E, David PH. Primary structure of the merozoite surface antigen 1 of Plasmodium vivax reveals sequences conserved between different Plasmodium species. Proc Natl Acad Sci U S A. 1991 May 1;88(9):4030–4. doi: 10.1073/pnas.88.9.4030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Galinski MR, Corredor-Medina C, Povoa M, Crosby J, Ingravallo P, Barnwell JW. Plasmodium vivax merozoite surface protein-3 contains coiled-coil motifs in an alanine-rich central domain. Mol Biochem Parasitol. 1999 Jun 25;(1–2):101. 131–47. doi: 10.1016/s0166-6851(99)00063-8. [DOI] [PubMed] [Google Scholar]
- 17.Vargas-Serrato E, Barnwell JW, Ingravallo P, Perler FB, Galinski MR. Merozoite surface protein-9 of Plasmodium vivax and related simian malaria parasites is orthologous to p101/ABRA of P. falciparum. Mol Biochem Parasitol. 2002 Mar;120(1):41–52. doi: 10.1016/s0166-6851(01)00433-9. [DOI] [PubMed] [Google Scholar]
- 18.Arevalo-Herrera M, Castellanos A, Yazdani SS, Shakri AR, Chitnis CE, Dominik R, et al. Immunogenicity and protective efficacy of recombinant vaccine based on the receptor-binding domain of the Plasmodium vivax Duffy binding protein in Aotus monkeys. Am J Trop Med Hyg. 2005 Nov;73(5 Suppl):25–31. doi: 10.4269/ajtmh.2005.73.5_suppl.0730025. [DOI] [PubMed] [Google Scholar]
- 19.Good MF. Towards a blood-stage vaccine for malaria: are we following all the leads? Nat Rev Immunol. 2001 Nov;1(2):117–25. doi: 10.1038/35100540. [DOI] [PubMed] [Google Scholar]
- 20.Pombo DJ, Lawrence G, Hirunpetcharat C, Rzepczyk C, Bryden M, Cloonan N, et al. Immunity to malaria after administration of ultra-low doses of red cells infected with Plasmodium falciparum. Lancet. 2002 Aug 24;360(9333):610–7. doi: 10.1016/S0140-6736(02)09784-2. [DOI] [PubMed] [Google Scholar]
- 21.Ceravolo IP, Souza-Silva FA, Fontes CJ, Braga EM, Madureira AP, Krettli AU, et al. Inhibitory properties of the antibody response to Plasmodium vivax Duffy binding protein in an area with unstable malaria transmission. Scand J Immunol. 2008 Mar;67(3):270–8. doi: 10.1111/j.1365-3083.2007.02059.x. [DOI] [PubMed] [Google Scholar]
- 22.Metzger WG, Okenu DM, Cavanagh DR, Robinson JV, Bojang KA, Weiss HA, et al. Serum IgG3 to the Plasmodium falciparum merozoite surface protein 2 is strongly associated with a reduced prospective risk of malaria. Parasite Immunol. 2003 Jun;25(6):307–12. doi: 10.1046/j.1365-3024.2003.00636.x. [DOI] [PubMed] [Google Scholar]
- 23.Nogueira PA, Alves FP, Fernandez-Becerra C, Pein O, Santos NR, Pereira da Silva LH, et al. A reduced risk of infection with Plasmodium vivax and clinical protection against malaria are associated with antibodies against the N terminus but not the C terminus of merozoite surface protein 1. Infect Immun. 2006 May;74(5):2726–33. doi: 10.1128/IAI.74.5.2726-2733.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Okech BA, Corran PH, Todd J, Joynson-Hicks A, Uthaipibull C, Egwang TG, et al. Fine specificity of serum antibodies to Plasmodium falciparum merozoite surface protein, PfMSP-1(19), predicts protection from malaria infection and high-density parasitemia. Infect Immun. 2004 Mar;72(3):1557–67. doi: 10.1128/IAI.72.3.1557-1567.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Barnwell JW, Galinski MR, DeSimone SG, Perler F, Ingravallo P. Plasmodium vivax, P. cynomolgi, and P. knowlesi: identification of homologue proteins associated with the surface of merozoites. Exp Parasitol. 1999 Mar;91(3):238–49. doi: 10.1006/expr.1998.4372. [DOI] [PubMed] [Google Scholar]
- 26.Kushwaha A, Rao PP, Duttu VS, Malhotra P, Chauhan VS. Expression and characterisation of Plasmodium falciparum acidic basic repeat antigen expressed in Escherichia coli. Mol Biochem Parasitol. 2000 Mar 5;106(2):213–24. doi: 10.1016/s0166-6851(99)00212-1. [DOI] [PubMed] [Google Scholar]
- 27.Oliveira-Ferreira J, Vargas-Serrato E, Barnwell JW, Moreno A, Galinski MR. Immunogenicity of Plasmodium vivax merozoite surface protein-9 recombinant proteins expressed in E coli. Vaccine. 2004 May 7;22(15–16):2023–30. doi: 10.1016/j.vaccine.2003.07.021. [DOI] [PubMed] [Google Scholar]
- 28.Pratt-Riccio LR, Lima-Junior JC, Carvalho LJ, Theisen M, Espindola-Mendes EC, Santos F, et al. Antibody response profiles induced by Plasmodium falciparum glutamate-rich protein in naturally exposed individuals from a Brazilian area endemic for malaria. Am J Trop Med Hyg. 2005 Dec;73(6):1096–103. [PubMed] [Google Scholar]
- 29.Tran TM, Oliveira-Ferreira J, Moreno A, Santos F, Yazdani SS, Chitnis CE, et al. Comparison of IgG reactivities to Plasmodium vivax merozoite invasion antigens in a Brazilian Amazon population. Am J Trop Med Hyg. 2005 Aug;73(2):244–55. [PubMed] [Google Scholar]
- 30.Singh H, Raghava GP. ProPred: prediction of HLA-DR binding sites. Bioinformatics. 2001 Dec;17(12):1236–7. doi: 10.1093/bioinformatics/17.12.1236. [DOI] [PubMed] [Google Scholar]
- 31.Alves FP, Durlacher RR, Menezes MJ, Krieger H, Silva LH, Camargo EP. High prevalence of asymptomatic Plasmodium vivax and Plasmodium falciparum infections in native Amazonian populations. Am J Trop Med Hyg. 2002 Jun;66(6):641–8. doi: 10.4269/ajtmh.2002.66.641. [DOI] [PubMed] [Google Scholar]
- 32.Coura JR, Suarez-Mutis M, Ladeia-Andrade S. A new challenge for malaria control in Brazil: asymptomatic Plasmodium infection–a review. Mem Inst Oswaldo Cruz. 2006 May;101(3):229–37. doi: 10.1590/s0074-02762006000300001. [DOI] [PubMed] [Google Scholar]
- 33.Baird JK. Age-dependent characteristics of protection v. susceptibility to Plasmodium falciparum. Ann Trop Med Parasitol. 1998 Jun;92(4):367–90. doi: 10.1080/00034989859366. [DOI] [PubMed] [Google Scholar]
- 34.Ceravolo IP, Bruna-Romero O, Braga EM, Fontes CJ, Brito CF, Souza JM, et al. Anti-Plasmodium vivax duffy binding protein antibodies measure exposure to malaria in the Brazilian Amazon. Am J Trop Med Hyg. 2005 Jun;72(6):675–81. [PubMed] [Google Scholar]
- 35.Bouharoun-Tayoun H, Oeuvray C, Lunel F, Druilhe P. Mechanisms underlying the monocyte-mediated antibody-dependent killing of Plasmodium falciparum asexual blood stages. J Exp Med. 1995 Aug 1;182(2):409–18. doi: 10.1084/jem.182.2.409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Cavanagh DR, Dobano C, Elhassan IM, Marsh K, Elhassan A, Hviid L, et al. Differential patterns of human immunoglobulin G subclass responses to distinct regions of a single protein, the merozoite surface protein 1 of Plasmodium falciparum. Infect Immun. 2001 Feb;69(2):1207–11. doi: 10.1128/IAI.69.2.1207-1211.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Topolska AE, Richie TL, Nhan DH, Coppel RL. Associations between responses to the rhoptry-associated membrane antigen of Plasmodium falciparum and immunity to malaria infection. Infect Immun. 2004 Jun;72(6):3325–30. doi: 10.1128/IAI.72.6.3325-3330.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Morais CG, Soares IS, Carvalho LH, Fontes CJ, Krettli AU, Braga EM. IgG isotype to C-terminal 19 kDa of Plasmodium vivax merozoite surface protein 1 among subjects with different levels of exposure to malaria in Brazil. Parasitol Res. 2005 Apr;95(6):420–6. doi: 10.1007/s00436-005-1314-x. [DOI] [PubMed] [Google Scholar]
- 39.Soares IS, Levitus G, Souza JM, Del Portillo HA, Rodrigues MM. Acquired immune responses to the N- and C-terminal regions of Plasmodium vivax merozoite surface protein 1 in individuals exposed to malaria. Infect Immun. 1997 May;65(5):1606–14. doi: 10.1128/iai.65.5.1606-1614.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Soares IS, Rodrigues MM. Immunogenic properties of the Plasmodium vivax vaccine candidate MSP1(19) expressed as a secreted non-glycosylated polypeptide from Pichia pastoris. Parasitology. 2002 Mar;124(Pt 3):237–46. doi: 10.1017/s003118200100110x. [DOI] [PubMed] [Google Scholar]
- 41.Rodrigues MH, Rodrigues KM, Oliveira TR, Comodo AN, Rodrigues MM, Kocken CH, et al. Antibody response of naturally infected individuals to recombinant Plasmodium vivax apical membrane antigen-1. Int J Parasitol. 2005 Feb;35(2):185–92. doi: 10.1016/j.ijpara.2004.11.003. [DOI] [PubMed] [Google Scholar]
- 42.Morais CG, Soares IS, Carvalho LH, Fontes CJ, Krettli AU, Braga EM. Antibodies to Plasmodium vivax apical membrane antigen 1: persistence and correlation with malaria transmission intensity. Am J Trop Med Hyg. 2006 Oct;75(4):582–7. [PubMed] [Google Scholar]
- 43.Soares IS, Oliveira SG, Souza JM, Rodrigues MM. Antibody response to the N and C-terminal regions of the Plasmodium vivax Merozoite Surface Protein 1 in individuals living in an area of exclusive transmission of P. vivax malaria in the north of Brazil. Acta Trop. 1999 Jan 15;72(1):13–24. doi: 10.1016/s0001-706x(98)00078-3. [DOI] [PubMed] [Google Scholar]
- 44.Braga EM, Barros RM, Reis TA, Fontes CJ, Morais CG, Martins MS, et al. Association of the IgG response to Plasmodium falciparum merozoite protein (C-terminal 19 kD) with clinical immunity to malaria in the Brazilian Amazon region. Am J Trop Med Hyg. 2002 May;66(5):461–6. doi: 10.4269/ajtmh.2002.66.461. [DOI] [PubMed] [Google Scholar]
- 45.Scopel KK, Fontes CJ, Ferreira MU, Braga EM. Factors associated with immunoglobulin G subclass polarization in naturally acquired antibodies to Plasmodium falciparum merozoite surface proteins: a cross-sectional survey in Brazilian Amazonia. Clin Vaccine Immunol. 2006 Jul;13(7):810–3. doi: 10.1128/CVI.00095-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Aucan C, Traore Y, Tall F, Nacro B, Traore-Leroux T, Fumoux F, et al. High immunoglobulin G2 (IgG2) and low IgG4 levels are associated with human resistance to Plasmodium falciparum malaria. Infect Immun. 2000 Mar;68(3):1252–8. doi: 10.1128/iai.68.3.1252-1258.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Fell AH, Currier J, Good MF. Inhibition of Plasmodium falciparum growth in vitro by CD4+ and CD8+ T cells from non-exposed donors. Parasite Immunol. 1994 Nov;16(11):579–86. doi: 10.1111/j.1365-3024.1994.tb00313.x. [DOI] [PubMed] [Google Scholar]
- 48.Hviid L, Theander TG, Jakobsen PH, Abu-Zeid YA, Abdulhadi NH, Saeed BO, et al. Cell-mediated immune responses to soluble Plasmodium falciparum antigens in residents from an area of unstable malaria transmission in the Sudan. Apmis. 1990 Jul;98(7):594–604. doi: 10.1111/j.1699-0463.1990.tb04976.x. [DOI] [PubMed] [Google Scholar]
- 49.Ramharter M, Willheim M, Winkler H, Wahl K, Lagler H, Graninger W, et al. Cytokine profile of Plasmodium falciparum-specific T cells in non-immune malaria patients. Parasite Immunol. 2003 Apr;25(4):211–9. doi: 10.1046/j.1365-3024.2003.00628.x. [DOI] [PubMed] [Google Scholar]
- 50.Theander TG, Hviid L, Dodoo D, Afari EA, Jensen JB, Rzepczyk CM. Human T-cell recognition of synthetic peptides representing conserved and variant sequences from the merozoite surface protein 2 of Plasmodium falciparum. Immunol Lett. 1997 Jun;58(1):1–8. doi: 10.1016/s0165-2478(96)02685-5. [DOI] [PubMed] [Google Scholar]
- 51.Troye-Blomberg M, Riley EM, Kabilan L, Holmberg M, Perlmann H, Andersson U, et al. Production by activated human T cells of interleukin 4 but not interferon-gamma is associated with elevated levels of serum antibodies to activating malaria antigens. Proc Natl Acad Sci U S A. 1990 Jul;87(14):5484–8. doi: 10.1073/pnas.87.14.5484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Farouk SE, Dolo A, Bereczky S, Kouriba B, Maiga B, Farnert A, et al. Different antibody- and cytokine-mediated responses to Plasmodium falciparum parasite in two sympatric ethnic tribes living in Mali. Microbes Infect. 2005 Jan;7(1):110–7. doi: 10.1016/j.micinf.2004.09.012. [DOI] [PubMed] [Google Scholar]
- 53.al-Yaman F, Genton B, Taraika J, Anders R, Alpers MP. Association between cellular response (IL-4) to RESA/Pf155 and protection from clinical malaria among Papua New Guinean children living in a malaria endemic area. Parasite Immunol. 1997 Jun;19(6):249–54. doi: 10.1046/j.1365-3024.1997.d01-204.x. [DOI] [PubMed] [Google Scholar]